Professional Documents
Culture Documents
Revista de Biologia
Uploaded by
Ramalde SantanaCopyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Revista de Biologia
Uploaded by
Ramalde SantanaCopyright:
Available Formats
CAPA REVISTA DE EDUCAO BIOLOGIA A SER ELABORADA PELA SECRETARIA DE FORMAO
APRESENTAO
A presente edio da Revista de Educao da APEOESP contem subsdios para os professores da rede pblica estadual, associados do nosso sindicato, que se inscrevero nos prximos concursos pblicos promovidos pela Secretaria de Estado da Educao e que participaro das provas institudas pelo governo. Organizada pela Secretaria de Formao, esta publicao contm as resenhas dos livros que compem a bibliografia dos concursos, realizadas por profissionais altamente qualificados, de forma a contribuir para os professores possam obter o melhor desempenho nas provas. Ao mesmo tempo, no podemos deixar de registrar nossa posio contrria s avaliaes excludentes que vem sendo promovidas pela Secretaria Estadual da Educao que, alm de tudo, desrespeita os professores ao divulgar extensa bibliografia a poucos dias da prova, inclusive contendo vrios ttulos esgotados. Esperamos, no entanto, que todos os professores possam extrair desta edio da Revista de Educao o mximo proveito, obtendo alto rendimento nas provas dos concursos e avaliaes. Nossa luta por mais concursos prossegue, com a periodicidade necessria a uma drstica reduo no nmero de professores temporrios, agregando mais qualidade ao ensino e profissionalizando, cada vez mais, o magistrio estadual. A periodicidade dos concursos a cada quatro anos com ritmo mais acelerado nos prximos dois anos foi uma conquista nossa e vamos exigir que seja efetivada. A diretoria
NDICE DE BIOLOGIA 1. ALBERTS, B.; et al. Fundamentos da biologia celular. 2. ed. Porto Alegre: Artmed, 2006. cap. 1, 4, 6, 7, 8, 10 a 19. 2. BOUER, J. Sexo & Cia: as dvidas mais comuns (e as mais estranhas) que rolam na adolescncia. 2 ed. So Paulo: Publifolha, 2002. 3. CARVALHO F.H; PIMENTEL S. M. R. A clula. Barueri: Manole, 2007. 4. CARVALHO, Isabel C. M. Educao ambiental: a formao do sujeito ecolgico. 3. ed. So Paulo: Cortez, 2008. cap. 1, 3 e 5. 5. DEAN, W. A ferro e fogo: a histria e a devastao da Mata Atlntica brasileira. So Paulo: Companhia das Letras, 1996. 6. GRIFFITHS, A.J. F. et al. Introduo Gentica. 9 ed. Rio de Janeiro: Guanabara Koogan, 2009. cap. 1 a 17, 19. 7. HICKMAN JR., Cleveland P.; ROBERTS, L. S.; LARSON, Allan. Princpios Integrados de Zoologia. 11. ed. Rio de Janeiro: Guanabara Koogan, 2004. 8. KORMONDY, E. J.; BROWN, D. E. Ecologia humana. So Paulo: Atheneu, 2002. 9. KRASILCHIK, M. Prtica de ensino de Biologia. 4. ed. So Paulo: EDUSP, 2008. 10. MARGULIS, L.; SCHWARTZ, K. V. Cinco reinos: um guia ilustrado dos filos da vida na Terra. 3. ed. Rio de Janeiro: Guanabara Koogan, 2001. 11. RAVEN, P. H.; EVERT R. F.; EICHHORN, S. E. Biologia Vegetal. 7. ed. Rio de Janeiro: Guanabara Koogan, 2007. sees 4, 5, 6 e 7. 12. RIDLEY, M. Evoluo. 3. ed. Porto Alegre: Artmed, 2006. 13. SCHMIDT-NIELSEN, K. Fisiologia Animal. Adaptao e meio ambiente. 5. ed. So Paulo: Livraria Santos, 2002. 14. SENE, F. M. Cada caso, um caso... puro acaso Os Processos de evoluo biolgica dos seres vivos. Ribeiro Preto: SBG, 2009. 15. TORTORA, G. J. Corpo humano: fundamentos de anatomia e fisiologia. 6. ed. Porto Alegre: Artmed, 2006.
1. ALBERTS, B.; et al. Fundamentos da biologia celular. 2. ed. Porto Alegre: Artmed, 2006. cap. 1, 4, 6, 7, 8, 10 a 19.
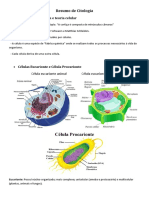
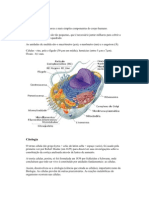
As clulas, unidades bsicas de todos os seres vivos, so envolvidas por membranas e preenchidas por uma soluo aquosa de agentes qumicos (citosol); so dotadas de extraordinria capacidade de criar cpias de si mesmas pelo crescimento e posterior diviso. A possesso de um ncleo por uma clula eucarioteca durante o processo evolutivo significa possuir tambm uma variedade de outras organelas. O ncleo normalmente a organela mais proeminente. Est incluso em duas membranas concntricas que formam o envelope nuclear, e que contm molculas de DNA, polmeros extremamente longos que codificam a especializao gentica do organismo. Essas molculas gigantes tornam-se individualmente visveis no microscpio ptico como cromossomos. O citoplasma de clulas vegetais e animais contm uma variedade de organelas delimitadas por membranas com funes qumicas especializadas. As mitocndrias realizam a oxidao de molculas, liberando energia. Os cloroplastos, em clulas vegetais, realizam a fotossntese. A maioria das organelas est envolvida com a necessidade de importar material bruto e exportar substncias manufaturadas e resduos. Retculo endoplasmtico: montagem, estocagem e exportao de molculas. Aparelho de Golgi: recebe, podendo modificar molculas provenientes do retculo, armazenando-as e exportando-as para a prpria clula ou para o exterior. Lisossomos: digesto intracelular. Peroxissomos: degradao do perxido de hidrognio (gua oxigenada). Filamento protico, o citoesqueleto estende-se por todo o citosol, dando forma e movimento clula. Quase todas as clulas realizam processos bsicos: duplicao do DNA, sntese proteicas, produo de energia etc. Apesar de terem o mesmo material gentico, cada tipo de clula usa, seletivamente, diferentes informaes gnicas de acordo com as funes a que se destina.
Como as clulas obtm energia a partir do alimento
Para liberar a energia de que as clulas necessitam para seu metabolismo, os alimentos energticos passam por trs etapas: 1. Degradao das macromolculas (amido, lipdios e protenas) em subunidades no tubo digestrio; 2. Degradao das subunidades, como a glicose, em piruvato, no citosol da clula; 3. Quebra final da molcula de piruvato nas mitocndrias. A quebra da glicose no citosol (gliclise) libera energia para a produo de 2 ATP e molculas de H, que so captados por um transportador de eltrons, o NAD (Nicotinamida Adenina Dinucleotdeo). O piruvato entra na mitocndria, perde carbonos e convertido na molcula de acetil-CoA. O acetil-CoA passa por uma longa srie de reaes qumicas (ciclo do cido ctrico ou de Krebs) que vo liberando molculas de gs carbnico (descarboxilaes), descartadas, e hidrognios. Esses, pelo NAD formam NADH2, e por outro carregador, o FAD (Flavina Adenina Dinucleotdeo), formam FADH2. Os hidrognios transportados pelos carregadores (NADH2 e FADH2) so levados at as membranas internas das mitocndrias (cristas) onde vo gradualmente passando por molculas aceptoras (cadeia respiratria). Nesse trajeto, ocorrem reaes de oxi-reduo, sendo liberada energia utilizada na converso do ADP + P em ATP. No final desse processo, molculas de 02 que entraram na mitocndria reagem com os hidrognios, formando molculas de gua. Esta a respirao celular ou aerbica: glicose + 02 C02 + H20 + ATP. Na ausncia da glicose, cidos graxos e aminocidos so convertidos em acetil-CoA, e o processo respiratrio prossegue. Sendo essencial vida, os seres vivos armazenam molculas energticas que sero utilizadas em situaes de compensao de ausncia de glicose: gorduras e glicognio nos animais e amido nos vegetais.
A replicao (duplicao) do DNA O potencial do DNA para a replicao e codificao de informaes deve-se ao fato de ser formado por duas longas cadeias ou fitas de nucleotdeos que se ligam atravs de pontes de hidrognio que unem as bases nitrogenadas Adenina com Timina e Citosina com Guanina. Cada fita da molcula de DNA contm uma sequncia de nucleotdeos exatamente complementar sequncia de nucleotdeos da outra fita. Cada lado do DNA atua como um molde para a sntese de uma fita complementar. Para ser duplicada, a dupla hlice, que tem ligao muito estvel, deve ser aberta
(modelo de zper). Protenas iniciadoras ligam-se ao DNA, forando a separao das fitas ao quebrar as pontes de hidrognio. O local onde o DNA aberto chamado de origem de replicao. A enzima DNA-polimerase catalisa a ao de nucleotdeos soltos que vo se encaixando em ambos os lados da molcula de DNA. Uma enzima, a helicase, usando energia do ATP, corre ao longo do DNA abrindo a dupla hlice medida que se move. Cada lado do DNA usado como molde para a formao da fita complementar. Dessa forma, as fitas originais permanecem intactas ao longo de muitas geraes. A replicao semiconservativa, porque cada hlice-fita composta de uma fita conservada e uma recm-sintetizada. Reparo do DNA: erros podem ocorrer durante a replicao, na unio entre as bases A-T/C-G. Quando ocorre um engano (exemplo T-T), a DNA-polimerase retira o nucleotdeo que se encaixou no lugar errado e adiciona o nucleotdeo certo. A estabilidade do gene depende do reparo do DNA. Se a mudana permanece, chamada de mutao, pois modifica a estrutura do gene. Dessa forma, modifica a forma da protena. Modificando a forma, altera a funo da protena codificada por esse gene. Mutaes so raras, porque o mecanismo de replicao e reparo do DNA so to acurados, que mesmo onde nenhuma seleo opera, a mensagem gentica fielmente preservada por milhes de anos.
Do DNA protena A expresso dos genes na clula passa pelos processos de transcrio (DNA RNA) e traduo (RNA protenas). A transcrio a sntese de RNA, catalisada pela enzima RNA-polimerase, a partir de pequenos segmentos de DNA. Antes de sair do ncleo para ir ao citoplasma, o RNA ativado com a retirada de trechos que no so codificados durante a sntese proteicas, chamados de ntrons. O RNA fica menor, mas ativo, porque os trechos ativos, os xons, se unem, tornando o RNA funcional. Esse processo conhecido como splicing de RNA mensageiro. A transcrio e a sntese de protenas ocorrem no citoplasma, no nvel dos ribossomos. O RNAm acopla-se a uma regio do ribossomo expondo trs cdons. Trs bases do RNAm correspondem a um cdon. Cada cdon indica o tipo e a posio dos aminocidos na cadeia de protena. Portanto, o cdigo que determina a relao cdon-aminocido. Com quatro bases nitrogenadas (U-A-C-G) combinadas, so possveis 64 tipos de cdons, e a maioria dos aminocidos reconhecida por mais de um cdon.
RNA transportadores (RNAt) ligam-se a aminocidos especficos. Cada RNAt possui trs bases expostas, o anticdon. Durante a sntese proteicas, cada cdon do RNAm unir-se- ao anticdon do RNAt. A sntese comea sempre com o cdon AUG, que codifica o aminocido metionina. Protenas chamadas de fatores de iniciao comandam esse incio. No ribossomo, dois cdons acoplam-se a dois anticdons do RNAt atravs do pareamento complementar de bases (ex.: cdon AUC, anticdon DAG). Aminocidos trazidos pelo RNAt se ligam por desidratao formando um dipeptdeo. O ribossomo desliga paulatinamente expondo novo cdon, quando o primeiro RNAt se desliga. Novo RNAt se encaixa no novo cdon e ocorre outra ligao entre aminocidos. Assim, a cadeia polipeptdica vai aumentando e termina a sntese quando um ribossomo chega a um cdon de terminao em que no h aminocido correspondente. Nesse local, encaixa-se uma protena chamada fator de liberao, que desliga todos os participantes do processo, soltando a protena.
CROMOSSOMOS E REGULAO CNICA Quase todas as clulas de um organismo multicelular contm o mesmo genoma. As diferenas entre um tipo de clula e outro (ex.: linfcto e neurnio) dependem dos tipos de genes que so expressos em cada uma delas. a expresso gnica. Em cada caso, a clula est usando somente alguns genes de seu repertrio total, que expressam diferentes RNA e protenas. Para que o DNA possa ser facilmente controlado dentro do ncleo, ele compactado e dobrado. A intervalos regulares, a molcula de DNA se enrola sobre grnulos de uma protena, a histona, formando estruturas globulares, os nucleossomos. No perodo da interfase, o cromossomo apresenta-se na forma conhecida como cromonema, na qual um fio de DNA repleto de nucleossomos se enrola helicoidalmente, como um fio de telefone. Em geral, regies do cromossomo que so transcritas em RNA so mais estendidas, enquanto aquelas que so mais compactadas, chamadas de heterocromatina, so inativas. Para um gene comear a se expressar (ou no), depender de uma srie de fatores como: tipo celular, sua circunvizinhana, idade e sinais exteriores. a expresso de um gene dependente de um conjunto de protenas reguladoras de genes que afeta o processo de montagem, facilitando ou diminuindo a velocidade das reaes. Essas protenas reguladoras ligam-se a pequenas sequncias de nucleotdeos (as sequncias regulatrias de DNA), aliviando ou
reprimindo a transcrio do gene. Uma nica protena regulatria, em clulas germinativas, pode disparar a formao de uma nica clula especializada ou mesmo de um rgo inteiro. A maioria das protenas reguladoras dos genes trabalha como parte de um "comit" de protenas regulatrias, todas necessrias para o gene se expressar na forma correta, em resposta a condies corretas, no momento certo e em um nvel requerido. Embora alguns mecanismos de controle se apliquem a clulas bacterianas e eucariticas, estas ltimas, por meio de estrutura cromossmica mais complexa, tm formas de controlar a expresso gnica no disponvel para bactrias. A tecnologia do DNA A tecnologia do DNA recombinante (ou Engenharia Gentica) rene as tcnicas que permitem a manipulao do material gentico, revelando os mecanismos pelos quais a expresso gnica regulada. Para ser estudado, um gene deve ser isolado e purificado. Enzimas bacterianas conhecidas como nucleases de restrio (ou endonucleases) so utilizadas para cortar o DNA em pedaos pequenos. Essas enzimas identificam determinadas sequncias de nucleotdeos de DNA e os cortam. Os fragmentos de DNA obtidos podem ser reunidos por outro tipo de enzima, a ligase. Com ela, pode-se reunir fragmentos de espcies diferentes, formando um DNA recombinante. Clonagem molecular: os segmentos de DNA obtidos pelas endonucleases so separados por eletroforese em gel, quando migram em um campo eltrico. Fragmentos maiores migram mais lentamente que os menores, formando uma escada de bandas discretas, como nos cdigos de barras. Assim eles so separados e isolados. Fragmentos so inseridos dentro de bactrias como a Escherichia coli. Eles se fundem aos plasmdeos (pedaos de DNA soltos) bacterianos que, ao se multiplicarem, duplicam tambm o fragmento de DNA enxertado. A reutilizao desses plasmdeos em outras bactrias produz milhares de cpias do segmento de DNA usado. Genes humanos tambm so clonados por essa tcnica, como o gene para o fator VIII, a protena da coagulao, cujos defeitos causam a hemofilia. Reao em cadeia da polimerase (PCR) - nesse processo, realizado in vitro, usa-se a enzima DNA-polimerase, que permite a fragmentao rpida de vrias cpias de um segmento de DNA extrado de clulas. Transcrio in vitro isolada a enzima transcriptase reversa, que forma uma molcula de DNAmolde a partir de uma fita de RNA (como ocorre em retrovrus, como o HIV), possvel sintetizar em laboratrio um DNA complementar (DNAc). Ele usado como uma sonda para localizar um gene no cromossomo. A Engenharia
Gentica usada para detectar doenas, na cincia forense, estudar protenas raras ou difceis de serem isoladas.
ESTRUTURA DA MEMBRANA
A membrana plasmtica um filme gorduroso to fino e transparente que no visvel no microscpio ptico. Serve de barreira para conter o contedo celular e permite a passagem de substncias entre a clula e o meio circundante. Cresce com a clula, aumentando sua rea sem perda de continuidade. Pode ser furada ou deformar-se sem se romper. As membranas internas que envolvem compartimentos intracelulares so iguais, na composio e propriedades, membrana plasmtica. As membranas celulares consistem numa dupla camada contnua de lipdeos, na qual as protenas esto embebidas. As clulas ajustam a fluidez de suas membranas pelas modificaes da composio lipdica dessas membranas. As molculas de lipdeos da membrana tm tanto regies hidrofbicas ("detestam gua") quanto hidroflicas ("gostam de gua"). Elas se arranjam espontaneamente em bicamadas quando colocadas em gua. Ainda que a bicamada de lipdeos fornea a estrutura bsica da membrana, a maior parte das funes realizada por protenas da membrana, como o transporte de nutrientes, metablitos ou ons. Algumas funcionam como receptores que detectam sinais qumicos no meio ambiente da clula e os transmitem ao interior da clula; outras agem como enzimas especficas. A maioria das membranas reforada e suportada por uma malha de protenas fibrosas chamada de crtex celular. Algumas clulas, como os eritrcitos, necessitam desse crtex para fornecer resistncia mecnica enquanto so bombardeadas atravs dos vasos sanguneos; outras clulas necessitam de seu crtex para capacit-las a mudarem ativamente de forma e a se moverem. Uma estrutura que recobre a membrana o glicoclix, formado por carboidratos que ajudam na proteo e lubrificao da clula, no reconhecimento do vulo por um espermatozoide, no reconhecimento de bactrias pelos neutrfilos (tipo de leuccito) etc.
A clula tem meios de confinar protenas a domnios especficos da membrana e de imobilizar alguns tipos de protenas, ligando-as a macromoleculares intracelulares ou extracelulares.
Transportes de membrana As clulas vivem e crescem pela troca de molculas com o meio, de modo que vrias molculas hidrossolveis devem ser capazes de cruzar a membrana plasmtica. Nutrientes devem que ser importados; produtos residuais devem ser expelidos, e as concentraes de ons como H+, Na+, K* e Ca++ precisam ser ajustadas. Solutos como C02 e 02, molculas lipossolveis e pequenas molculas sem carga podem se difundir simplesmente pela membrana, mas a grande maioria no pode faz-lo. Dependem de protenas especiais. A membrana da clula contm uma variedade dessas protenas transportadoras, cada uma sendo responsvel pela transferncia de um tipo particular de soluto. H duas classes delas: protenas carreadoras e protenascanal. As protenas carreadoras ligam-se a um soluto de um lado da membrana, e o levam para o outro lado. Funcionam como uma catraca, transportando ons e pequenas molculas orgnicas. Podem atuar como bombas ao transportar ons contra um gradiente de concentrao, com gasto de energia. Ex.: a protena gasta energia para transportar ons sdio (Na*) para fora da clula e ons potssio para dentro. Graas a essas protenas, as clulas animais mantm a concentrao de ons sdio cerca de 10 a 15 vezes maior fora da clula do que dentro e quase 10 vezes mais potssio dentro do que fora. o transporte ativo. O transporte por protenas carreadoras pode ser ativo ou passivo. Uma dessas protenas chamada de permease, que transporta molculas de glicose passivamente, sem gasto de energia. As protenas-canal formam minsculos poros aquosos por onde passam solutos; o transporte sempre passivo. Se o canal est aberto, molculas pequenas e com carga apropriada podem passar por ele. Osmose a passagem da gua (solvente) de um meio menos concentrado para um mais concentrado. Se a clula estiver num ambiente mais concentrado (hipertnico) que o seu interior, ela perde gua e murcha. Se o ambiente externo for menos concentrado (hipotnico) que o interior da clula, ela ganha gua, aumenta de volume, podendo arrebentar.
Gerao de energia em mitocndrias e cloroplastos A proviso de uma abundante fonte de energia para a clula foi fundamental para a evoluo da vida no planeta. O mtodo mais eficiente para a sntese de ATP baseia-se no transporte de eltrons pelas membranas, o que ocorre tanto na fotossntese como na respirao aerbica. Fosforilao oxidativa (sntese de ATP) nas mitocndrias: piruvatos (originados da quebra da glicose no citosol) e cidos graxos entram na mitocndria e so quebrados a acetil-CoA. So metabolizados pelo ciclo do cido ctrico (Krebs), produzindo C02, que descartado, e hidrognios, captados por molculas transportadoras, o NAD e o FAD, que os conduziro at a cadeia respiratria. A maior parte da energia da glicose encontra-se nos eltrons dos tomos de hidrognio. Na membrana interna da mitocndria, encontram-se substncias aceptoras de eltrons (cadeia respiratria). Os eltrons dos tomos de hidrognio transportados pelos NADH e FADH2 comeam a percorrer gradativamente os componentes da cadeia, em reaes de oxi-reduo. Nessas passagens, os eltrons vo perdendo energia, que utilizada na sntese de ATP. No final da cadeia respiratria, o oxignio recolhe os eltrons, e combina-se com prtons H+, formando gua. O oxignio o aceptor final do oxignio.
OS CLOROPLASTOS E A FOTOSSNTESE As reaes da fotossntese so agrupadas em duas categorias: reaes de fase clara e reaes de fase escura. Nas reaes de claro, a energia derivada da luz energiza um eltron de clorofila que, excitado, comea a se mover ao longo de uma cadeia transportadora de eltrons da membrana interna do cloroplasto (tilacoide). A clorofila obtm seus eltrons da hidrlise (quebra) da gua, produzindo 0 2 como subproduto. Nessa passagem, h liberao de energia usada na sntese de ATP. No final dessa srie de reaes, os eltrons so carregados, juntamente com H+, pelo NADP, convertendo-o em NADPH2. Nas reaes de escuro, o ATP fornece energia e o NADPH2 participa como redutor, fornecendo os H2 ricos em energia que, numa srie de reaes qumicas, se convertem em glicose.
COMPARTIMENTOS INTRACELULARES E TRANSPORTE As clulas desenvolveram vrias estratgias para segregar e organizar as suas reaes qumicas. Uma estratgia agregar as diferentes enzimas utilizadas para catalisar diversas reaes como sntese de RNA, DNA, protenas. Outra confinar os diferentes processos, bem como as protenas necessrias, em compartimentos delimitados por membranas (lisossomos, aparelho de Golgi, mitocndrias, cloroplastos etc.). As protenas usadas no ncleo so transportadas por meio dos poros nucleares, que funcionam como portes seletivos de molculas. As protenas das organelas so, na sua maioria, produzidas no citosol e transportadas s organelas onde funcionam. Sinais na distribuio da sequncia de seus aminocidos guiam as protenas organela correta. As protenas que operam no citosol no possuem sinais e permanecem onde so sintetizadas. O transporte de protenas entre o local de sntese (ergastoplasma) e o local de armazenagem (Golgi), e desse para outros destinos (lisossomos, gros de zimgeno das clulas do pncreas, etc), realizado por vesculas de transporte, que brotam continuamente de uma membrana e se fusionam outra. Em todas as clulas eucariticas, as vesculas de transporte brotam do aparelho de Golgi, e se fundem membrana plasmtica num processo chamado de exocitose. Essa rota fornece lipdeos e protenas para o crescimento e alargamento da membrana, conduz protenas para serem liberadas do lado externo (secreo) etc. As vesculas endocticas formam-se aps a ingesto de fluidos, molculas e partculas, por invaginao da membrana (fagocitose e pinocitose). Essas vesculas ligam-se aos lisossomos que contm enzimas hidrolticas. Aps a digesto, a maior parte dos componentes dessas vesculas reciclada em vesculas de transporte, de volta membrana plasmtica para reutilizao.
COMUNICAO CELULAR As clulas individuais, assim como organismos multicelulares, precisam sentir e responder aos estmulos do ambiente. Nos multicelulares, as clulas precisam coordenar seu comportamento de vrias maneiras. Para isso, utilizam centenas
de tipos de molculas extracelulares para enviar sinais umas s outras (protenas, peptdeos, aminocidos, nucleotdeos, esteroides e at gases dissolvidos). A converso de informao de uma forma para outra chamada de transduo de sinal. Uma clula sinalizadora produz um tipo especial de molcula (sinal). A clula-alvo recebe o sinal por meio de um receptor (protena especial da membrana). O sinal enviado ao interior da clula e determina o comportamento celular. H quatro tipos de sinalizao entre as clulas: 1. Sinal hormnio: disseminado pelo corpo atravs da corrente sangunea (animal) ou da seiva (planta); 2. Sinalizao parcrna: molculas sinalizadoras atuam a curta distncia, permanecendo na vizinhana da clula que as produziu. So mediadores locais. Ex.: molcula mediadora que coordena a inflamao nos locais de infeco ou na cicatrizao; 3. Sinalizao neuronal: sinais eltricos celulares so convertidos num sinal extracelular qumico (neurormnios), nas sinapses entre neurnios e entre neurnios e msculos ou glndulas; 4. Sinalizao por contato: contato por meio de ligao de uma molcula sinalizadora presente na membrana molcula receptora da membrana plasmtica da clula-alvo. Ex.: ocorre na fase de diferenciao celular da fase embrionria. Uma clula tpica possui uma coleo de diferentes receptores na membrana. As molculas sinalizadoras trabalham em combinao para regular o comportamento da clula. Alguns sinais estimulam o metabolismo celular, outros induzem diviso, outros, diferenciao. Caso privadas de receptores apropriados, a maioria das clulas sofre uma espcie de suicdio celular (morte celular programada ou apoptose).
O CITOESQUELETO uma rede intrincada de filamentos proticos que se estende por todo o citoplasma. Essa rede auxilia na sustentao do grande volume do citoplasma e capacita a clula. Auxilia tambm a se organizar interiormente e adotar uma variedade de formas e movimentos amplos: deslocamento de clulas sobre uma superfcie, contrao de clulas musculares, alteraes na forma celular durante o desenvolvimento embrionrio etc. A presena do citoesqueleto pode ter sido um fator crucial na evoluo das grandes, e estruturalmente complexas, clulas eucariticas. O citoesqueleto constitudo a partir de trs tipos de filamentos proticos: filamentos intermedirios, microtbulos e filamentos de actina. Filamentos
intermedirios - Grande resistncia trao; sua funo principal capacitar as clulas a suportarem tenso mecnica produzida quando estas sofrem estiramento; so os mais resistentes e durveis. Formam uma rede por todo o citoplasma, circundando o ncleo e se estendendo para a periferia da clula. Presentes no axnio dos neurnios, em clulas musculares, so semelhantes a cabos formados por muitos fios longos enrolados juntos. Microtbulos - Tubos longos, ocos e relativamente rijos, formados por uma protena, a tubulina; so capazes de desagregao em um local e reagrupamento em outro. Crescem a partir do centrossomo, estendendo-se para a periferia da clula. Criam um sistema de trilhos, sobre os quais vesculas, organelas e outros componentes celulares podem se locomover. No incio da mitose ocorre a desagregao dos microtbulos e o rearranjo, formando o fuso mittico. Tambm podem formar clios e flagelos. Filamentos de actina - Cada filamento uma cadeia torcida de molculas globulares de actina. Sem esses filamentos, uma clula no poderia rastejar, emitir pseudpodos ou se dividir. Protenas motoras, dependentes da actina, pertencem famlia das miosinas. As miosinas usam a energia do ATP para se deslocarem sobre a actina, promovendo os movimentos musculares de contrao e descontrao.
DIVISO CELULAR CONTROLE DO CICLO CELULAR E MORTE CELULAR O ciclo celular de uma clula eucaritica consiste em vrias fases distintas: perodos G1 S, G2 e M (G = pausa; S = sntese; M = Mitose). A pausa G o estgio que corresponde ao intervalo entre o trmino da diviso (M) e a fase S, em que ocorre a sntese ou duplicao do DNA. Na fase G 1 h intensa atividade metablica na clula. Ocorre sntese de protenas, e a clula cresce em massa. A fase G2 a pausa entre a duplicao dos cromossomos e o incio da diviso. Juntas, as fases G1 e G2 proporcionam um perodo adicional para a clula crescer e duplicar suas organelas citoplasmticas. A fase M comea com a duplicao do centrossomo e a formao do fuso mittico, composto de microtbulos que levam os cromossomos-filhos a plos opostos da clula. Grandes organelas, como o retculo endoplasmtico e o aparelho de Golgi, rompem-se em pequenos fragmentos, assegurando a distribuio idntica entre as clulas-filhas. Mitocndrias duplicam-se. Tm
DNA. Quando o envelope nuclear se rompe, durante a prfase (1a etapa da mitose), os microtbulos do fuso invadem a rea nuclear. Alguns deles capturam os cromossomos duplicados, ligando-os a complexos de protena com o centrmero de cada cromtide-irm. Os microtbulos de plos opostos deslizam, arrastando cada cromtide (agora cromossomo) para os plos da clula. Reorganizadas as cariotecas, a clula, agora com dois ncleos, prepara-se para a diviso do citoplasma (citocinese). Nas clulas animais, forma-se um.anel contrtil de filamentos de actina e miosina (miofibrilas de natureza proteicas caractersticas das fibras musculares) na regio central da clula, que vai se contraindo e estrangulando. Dessa forma, o citoplasma divide-se em duas novas clulas, geneticamente idnticas clula-me.
Controle do ciclo celular e morte celular O controle do ciclo celular consiste num conjunto de protenas especficas que controlam a proliferao, sobrevivncia e morte celular. Esse mecanismo garante que a clula s se divida quando necessrio. Vrias clulas normais morrem durante a vida de um animal, por um processo de morte programada ou apoptose.
TECIDOS A maioria das clulas de um organismo multicelular est organizada em grupos cooperativos, os tecidos, que so compostos no somente de clulas, mas tambm de uma matriz extracelular que elas secretam ao seu redor. essa matriz extracelular que d fora de sustentao ao tecido. A resistncia da matriz se deve a protenas fibrosas chamadas de colgeno. normal a distino de quatro tipos de tecidos em animais: conjuntivo, epitelial, nervoso e muscular. Tecidos conjuntivos (ossos, tendes, cartilagens etc): a principal caracterstica a existncia de diferentes tipos de clulas, separadas por abundante substncia por elas fabricadas. H tambm nmerosas fibras proteicas entremeadas com as clulas, o colgeno e as fibras elsticas. H vrios tipos de tecidos conjuntivos: frouxo, denso, adiposo, sanguneo, sseo e cartilaginoso. A diferena entre eles relacionada ao tipo de componente predominante. As fibras de colgeno assemelham-se a cordas dispostas irregularmente, formando malhas difusas, dando resistncia ao tecido; as fibras
elsticas do elasticidade aos tecidos onde se encontram (pulmo, fgado etc). As clulas do tecido conjuntivo so fibroblastos, mastcitos, macrfagos, clula adiposa, osteoblastos etc. Tecido epitelial: especializado em revestimento e proteo das partes internas e externas do corpo e na formao de glndulas. Suas clulas so firmemente aderidas umas s outras, o que lhe d grande resistncia. No h vasos sanguneos, e a nutrio por difuso. Na maioria dos epitlios aparecem junes ou regies de colagem entre as clulas, as junes aderentes e os desmossomos, formados por filamentos de queratina que estabelecem firme adeso entre as clulas. Esse tecido pode ser simples se tiver uma camada de clulas, e estratificado caso possua mais de uma camada. Os tecidos nervoso e muscular no so tratados aqui. A organizao dos tecidos ativamente mantida por comunicao celular, adeso seletiva clula-clula e clula de memria. Clulas cancerosas surgem do acmulo de mutaes somticas e violam as regras que mantm a organizao dos tecidos.
Os movimentos celulares programados criam o plano de corpo de um animal Embora a estrutura final do corpo de um animal possa ser extremamente complexa, ela gerada por um repertrio limitado de atividades celulares na fase embrionria. A fertilizao induz a clivagem dessa clula em vrias outras menores, as quais realizam uma srie de movimentos. Enquanto algumas clulas movem-se livremente, outras so mantidas juntas. O processo inicial a formao do intestino (gastrulao), o que ocorre em qualquer animal pela invaginao de clulas do exterior do corpo. Aps isso, movimentos celulares formam os rudimentos das principais caractersticas do corpo: cabea, cauda, dorso etc. Em geral, as clulas atuam como fonte de molculas de sinalizao que influem outras clulas nas vizinhanas. Algumas clulas respondem, produzindo novos sinais. Clulas vizinhas, em diferentes distncias da fonte, so expostas a sinais de diferente intensidade, produzindo novos sinais. Clulas de memria preservam traos dos sinais recebidos (ativao ou represso de genes particulares). Esses sinais so protenas sinalizadoras (sonic hedgehog). Clulas do sistema nervoso central respondem a um forte sinal, formando a placa basal, enquanto que as clulas do broto dos membros
respondem formando um dedinho. Graas s clulas de memria, poucos mecanismos bsicos de sinalizao so aparentemente suficientes para dirigir a criao de uma grande variedade de estrutura altamente elaborada.
Sntese elaborada por Maria Aparecida Cassiani
2. BOUER, J. Sexo & Cia: as dvidas mais comuns (e as mais estranhas) que rolam na adolescncia. 2 ed. So Paulo: Publifolha, 2002. Livro de sexo, para qu? Para o autor, falar e escrever sobre o assunto um servio, um espao para tirar dvidas, apesar de ser uma tarefa difcil, por envolver tabus, preconceitos, mitos. Por meio do livro, o autor espera que, de alguma maneira, possa ajudar a resolver as dvidas, informar melhor e quebrar alguns tabus e preconceitos que cercam a sexualidade. Sexo bom, e normal ter dvidas e deve ser cada vez mais natural que as pessoas procurem respostas para aquilo que no sabem. O livro considerado, pelo autor, um verdadeiro manual de sobrevivncia. Nele, voc encontrar atravs de ilustraes, dvidas, depoimentos de adolescentes e tudo sobre: MUDANAS - da cabea aos ps, tudo fica diferente! difcil acreditar que as mudanas que transformam o corpo da gente variam muito de pessoa para pessoa. Na real, quem tem pacincia de esperar?Como controlar a ansiedade de usar um suti ou um barbeador quando todos os amigos da turma j esto nessa? E a sensao de que todo mundo j cresceu e voc continua pequeno? No mole! Mas no h mgica, nem creme, nem reza que faam voc ter seu desenvolvimento antes do planejado. O comeo das mudanas no tm data marcada! Cada um vai ter um calendrio pessoal. preciso ter pacincia para esperar a hora certa. O ritmo de desenvolvimento difere de pessoa para pessoa. - Algumas dvidas e depoimentos para tranqilizar e deixar a natureza seguir seu curso em paz.
Meus testculos so muito pequenos. Meu m dico disse, h uns dois anos, que minhas transformaes estavam comeando (isso depois de ver o meu saco). Agora tenho 16 anos e quero saber quanto tempo mais vou ter de esperar.
Tenho 14 anos e ainda no fiquei menstruada. Todas as minhas amigas j esto nessa fase. Ser que estou muito atrasada.
- ESPINHA - tire a mo delas j! Elas so capazes de provocar estragos no ego de qualquer um. As espinhas so um dos sinais da entrada na puberdade. Elas esto relacionadas com uma ao maior dos hormnios sexuais.
Minhas aulas comeam agora e estou cheio de espinhas. O que fazer para que elas desapaream?
Tenho marcas de espinhas nas costas. So manchas escuras que me deixam com vergonha de usar roupas decotadas. Existe algum creme para clarear essas manchas?
- Importante, para todas as dvidas o melhor consultar um especialista de pele, um Dermatologista.
- PLOS com mais ou com menos, todos normais. Os plos so mais um sinal de que garotos e garotas chegaram puberdade. A maior preocupao com a quantidade. Uns acham que tm muito pouco, outros acham que tm demais. A gentica que conta: o que vai determinar a quantidade de plos no corpo so as informaes genticas que recebemos dos nossos pais. So raros os casos em que a quantidade de plos significa realmente um problema. Normalmente, quando eles acontecem, esto ligados a disfunes dos hormnios. D uma olhada nas dvidas mais comuns:
Tenho 17 anos e passo muita vergonha porque meu corpo tem poucos plos e menos masculino do que os dos meus amigos. Acho que vou tomar vitamina ou algum tipo de hormnio que resolva meu problema.
Estou curiosa para saber se existe algum produto que possa eliminar de vez o buo. Sou mulher e acho esse bigode ridculo.
Eu queria saber se verdade que a plula anticoncepcional retarda o crescimento dos plos. Se for verdade, posso tomar alguma? Qual a mais recomendvel? Tenho 14 anos. Obs: Aconselhvel procurar um endocrinologista para uma avaliao. S esse especialista poder dizer se existe problema hormonal, o que bastante comum. Nunca tome remdios sem prescrio mdica.
- ALTURA - nem sempre possvel ser grande. Tenho 18 anos e 1,70m. Meu irmo tem 16 e j mede 1,75m . Meus amigos so mais altos do que eu. Posso tomar algum remdio ou colocar um aparelho na canela para crescer mais uns 7 cm? Tambm na altura a gentica um fator determinante. E quase nada possvel para mudar essa determinao. No h tratamento milagroso para esticar uma pessoa. Mas o melhor meio de tirar a dvida consultar o pediatra. Outro recurso bastante importante o exame de raios X que indica a idade ssea. E so ultra importantes as boas condies de alimentao e de sade na infncia.
- PESO - pegue leve. O peso tambm motivo de encanao para muita gente. Hoje existe uma espcie de culto ao corpo supermagro. O padro de beleza cruel e muita meninas e at meninos entram em paranoia de que esto gordos, e, na verdade, no esto. Lembrando que na adolescncia, com tantas mudanas corporais, o peso pode demorar um pouco para se normalizar. E um peso saudvel tem a ver com um estilo de vida saudvel. No desenvolvimento, quando se trata de questes que envolvem o peso, preciso muita ateno, pois com a preocupao em aparentar e manter um
corpo perfeito surgem os problemas j conhecidos por todos, que so o uso de frmulas para emagrecer (combinao de substncias) que tiram o apetite, o que gera o risco de se adquirir duas doenas: - a anorexia: cuja caracterstica o fato de que a pessoa deixa de comer e, mesmo estando magra, se acha gorda quando v sua imagem no espelho. Essa distoro da prpria imagem faz com que a pessoa queira emagrecer cada vez mais, e como consequncia, acontecem problemas gravssimos de sade, e at morte. Bulimia: uma ingesto exagerada de alimentos, seguida de uma tentativa desesperada de eliminar esse excesso. A pessoa tem episdios em que come de forma compulsiva tudo o que v pela frente e depois, tomada por culpa e arrependimento, provoca vmitos, faz muitos exerccios, toma diurtico e laxantes. As vttimas potenciais de bulimia so as pessoas que fazem dietas exageradas, sem orientao.. Essas doenas precisam de tratamento especializado, com terapia, orientao nutricional e, muitas vezes, remdios antidepressivos.
- PNIS & CIA rgua para qu? O pnis o campeo de dvidas entre os meninos. pnis pequeno, grande, torto para a direita, para a esquerda, para cima, para baixo... E a pergunta que fica : por que os caras encanam tanto com a forma e o tamanho do pnis? A puberdade traz para a vida mudanas rpidas, com as quais no estamos acostumados. E o pnis muda, cresce, engrossa, comea a liberar esperma etc. o primeiro ensaio para a vida adulta. E os ensaios sempre causam uma certa dose de ansiedade, insegurana e vergonha. Eu tenho 17 anos, e a camisinha fica larga no meu pnis. O que fao? Entre tantas dvidas que surgem, sempre melhor procurar um Urologista, e pode confiar que amigo.
-VAGINA & CIA. Prazer em conhec-la! Est ai uma parte do corpo feminino que as mulheres muitas vezes nem conhecem direito. E os homens, muitos menos. No conhecem, mas deveriam. A vagina o canal que leva ao tero. onde o pnis penetra e por onde saem os bebs. Suas paredes so elsticas; por isso, na hora do parto, d para uma criana de cerca de 3 quilos passar por ela. A vagina um lugar quente e mido e precisa de vigilncia constante para evitar o aparecimento de
Uma das coisas que mais perturbam as mulheres: infeces e corrimentos vaginais. Muitas meninas tm vergonha ou dvidas sobre suas vaginas. Umas as acham grandes, feias e pensam que os namorados no vo aprovar.Outras no sabem direito o que existe dentro dela. A maioria jamais tocou ou olhou sua prpria vagina. Para conhec-la, basta a ajuda de um espelhinho, uma boa conversa com pessoas de sua confiana e a primeira visita a um ginecologista que um especialista, um profissional cuidadoso preparado para responder as dvidas. Ele no morde. -HMEN: voc j sabe o que ? uma pele bem fininha que fica na entrada da vagina e se rompe, em geral, na primeira transa. Segundo a literatura mdica, existem vrios tipos de hmen: -hmen anular: o mais comum; tem um nico orifcio no meio. -hmen complacente: o mais grosso e elstico e pode no se romper na penetrao. -hmen cribiforme: pouco comum. Composto de vrios pequenos orifcios. -hmen imperfurado: raro e a garota vai precisar d uma pequena cirurgia para permitir a passagem do fluxo menstrual.
Existe um limite de tamanho do pnis que uma vagina pode ag uentar? Um pnis muito longo motivo de desconforto ou de prazer para a mulher?
verdade que os absorventes internos no tiram a virgindade? Tenho 14 anos e queria saber se posso usar esse tipo de absorvente sem susto?
- SEIOS - cuidados com os modismos. O tamanho dos seios um assunto que continua a mexer com a cabea de meninas e meninos O aparecimento de brotos mamrios (pequenas salincias que ficam sob as arolas) um dos primeiros sinais de que a garota est entrando na puberdade, e ocorre acontece entre os 10 e12 anos. As mamas comeam a crescer e, em geral, no final da puberdade, adquirem seu tamanho definitivo. Gentica e caractersticas individuais (como a deposio de gorduras no corpo) fazem com que algumas garotas tenham mamas menores ou maiores. Quanto a cirurgia de reduo ou aumento (silicone) no processo de desenvolvimento de uma garota devem ser vistas
com cautela, podendo atrapalhar um processo natural de desenvolvimento. So vrias as dvidas sobre os seios. Como: Tenho 17 anos, sou magra ( 49 quilos), mas tenho seios muitos grandes. Morro de vergonha. Uso creme para diminuir e para firmar os seios. Isso funciona? No, claro. -Importante a conscientizao de que se deve fazer o exame preventivo; muito fcil e indolor. Uma das formas o auto-exame que a garota mesmo pode fazer, apalpando, com cuidado, seus seios durante o banho. Se notar algo, procurar o mdico.
- MENSTRUAO Menstruar normal. Faz parte da vida de qualquer mulher. No doena, mas pode mexer um pouco com sua vida. Algumas garotas sofrem demais com a chegada da menstruao. Ficam tristes, sensveis, irritadas, nervosas, prontas para brigar. o que os especialistas chamam de TPM (tenso pr-menstrual) que uma sndrome, uma srie de sintomas conjuntos, e pode ser provocada por alteraes hormonais, reteno de lquido no organismo e mudana na concentrao de serotonina (transmissor qumico do crebro, portanto no frescura da mulher. Todavia, algumas acabam passando por esses dias sem essas alteraes e sensaes. O autor aborda temas que tambm so bastante complicados na fase da adolescncia, pois envolvem as emoes; so eles: namoro, timidez,vida sexual, pais, ansiedade e depresso. Na adolescncia comum achar que o sexo a parte mais difcil do relacionamento, mas no . Entender as prprias dvidas, incertezas, os questionamentos surgidos nessa fase de transio, sabendo como estabelecer limites, aprendendo a lidar com os outros e sentir-se seguro na hora de tomar decises so tarefas que no se efetivam da noite para o dia. . Tantas coisas acontecendo, muitas pessoas enfrentando problemas srios acabam sofrendo de ansiedade e depresso. . A auto-estima cai, o desempenho na escola piora, as relaes sociais prejudicadas e a vida podem se tornar muito chata. Ficam tristes, desanimadas, infelizes, sem nimo, tomadas de angstia e atitudes estranhas. Alm de lidarem com todas as dvidas comuns da adolescncia, o/a adolescente luta contra uma dificuldade a mais: preconceitos raciais, sexuais, sociais, culturais e religiosos. Mes e pais que no admitem que seus filhos namorem pessoas de outros nveis sociais e raas, colegas que tratam mal pessoas que gostam de gente do mesmo sexo, grupinhos que excluem quem no se encaixa no que a maioria pensa e sente e assim por diante. No entanto,
esse quadro deve ser transformado, pois no podemos admitir que, atualmente, as pessoas sejam discriminadas porque so seguem a norma, o padro. ! E ns precisamos aprender a compreenso e o respeito s diferenas.
Ao falar do tema Sexo, o autor no tem a inteno de fazer um guia completo como o b--b do sexo. A a ideia auxiliar no preparo para a vida sexual de uma forma mais tranqila e responsvel, prazerosa e natural. Faz parte da vida como dormir e comer, entre tantas outras coisas. A escolha pelo melhor momento para se iniciar a vida sexual de cada um, pessoal. Desnecessrio ter pressa. O importante o sentimento de maturidade para se iniciar a vida sexual. As dvidas so muitas: beijo, masturbao, virgindade, a primeira transa, dor e sangramento, ejaculao e ereo, sexo anal, orgasmos, mas at as pessoas experientes tm suas dvidas, problemas etc.
Sexo bom! afirma o autor, eno h dvida. Todavia, ele traz o risco da transmisso de algumas doenas, as sexualmente transmissveis (DST). Desde que o mundo mundo e desde que as pessoas fazem sexo, as DST do sinal de vida. Historicamente, desde a Antiguidade, , h muitas referencias s DST j. O homem convive h sculos com doenas como gonorria, sfilis e outras. Contudo, de duas dcadas para c, estamos convivendo com a doena mais grave de toda a histria: a Aids. Ela j levou milhes de vidas e deixou muitos rfos pelo mundo afora. Em alguns pases, a situao dramtica. No Sul da frica, mais de 30% da populao adulta de algumas naes esta contaminada pelo vrus HIV, causador da Aids. No Brasil, so quase 600 mil pessoas infectadas pelo HIV. Muita gente morreu. E muita gente aprendeu a conviver com ele. Para evitar a Aids, a informao, o combate ao preconceito e o uso regular da camisinha so os maiores aliados. E tambm outras doenas como: Herpes, condiloma (HPV), sfilis, gonorria, chato, candidase, clamdia e tricomonase. Como se pode observar, h vrias doenas sexualmente transmissveis (DST) e, para cada uma delas, existe um tipo de exame
diferente. No h um nico exame que detecte todas ao mesmo tempo. Na dvida, procure um mdico, ele ajudar.
E USE CAMISINHA.
Todos sabem disso, mas na hora H, parecem esquecer e a... Qual outra explicao para que uma em cada cinco adolescentes entre 13 18 anos j tenha enfrentado uma gravidez aqui no Brasil? So diversos os problemas e as implicaes que uma gestao pode trazer para meninas que ainda moram com os pais, mal tm namorado fixo e esto na escola ou comeando uma vida profissional. Ser que a falta de informao faz com que as garotas passem por essa situao? Pouco provvel! Falta, sim, cuidado, preveno, mtodo anticoncepcional e a cabea um pouco mais em cima do ombro quando pinta a paixo ou o teso. E no esquea! a camisinha sua melhor amiga. E fale sempre com o mdico.
E finalizando, DROGAS. A busca de novas formas de prazer e de sensaes diferentes vem de muito tempo atrs em nossa histria. Mas hoje o consumo e a variedade de drogas alcanaram uma proporo nunca vista. A melhor maneira de enfrentar as drogas conhec-las. Saber quais so, o que provocam, que riscos oferecem, por que as pessoas as consomem etc. Alm das drogas ilegais, como a maconha ou a cocana, preciso saber que o fumo e as bebidas alcolicas, que esto por toda parte, tambm so drogas. E, de longe, so as mais consumidas pelo homem. E tambm remdios, prescritos por mdicos para curar problemas, e encontrados em qualquer farmcia, acabam sendo utilizados como drogas. Quem experimenta alguns tipos de droga no se torna um dependente da noite para o dia. No entanto, em geral, de maneira geral, d
para dizer que quem usa droga com frequncia vai enfrentar problemas de sade, dificuldades sociais e perdas importantes. Drogas e sexo andam juntos, e quem consome qualquer tipo de drogas tem sua capacidade de avaliar riscos alterada e pode, entre outrasatitudes perigosas, fazer sexo sem proteo. QUESTES
1) A que tema se refere a frase a seguir: Calma L! No existe hora marcada para as transformaes que acontecem. No corpo. a) Mudanas b) plos c) hormnios d) Drogas e) Nenhuma das anteriores.
2)Alguns sinais indicam que os hormnios comeam a aumentar: espinhas, crescimento das mamas e aparecimento de plos prximos vagina e nas axilas. Para se checar como anda o desenvolvimento do corpo, que profissional consultar? a) Dermatologista b) Endocrinologista c) Ginecologista d) Pediatra e) Todos.
3) Para o autor, o que importante para o (a) adolescente iniciar sua vida sexual? a) Capacidade de assumir responsabilidade b) Maturidade c) Permisso dos pais
d) Confiana no (a) parceiro (a) e) Estar apaixonado(a) 4) Quais as doenas que o autor considera como DSTs ? a) Aids, herpes. b) Condiloma (HPV), Sfilis. c) Gonorria, chato, Candidase. d) Clamdia, Tricomonase e) Todas
5) So consideradas drogas disfaradas: a) Cocana e Crak b) lcool e Cigarro c) Prozac e Maconha d) remdios e)Todas
GABARITO
1. A / 2. C / 3. B / 4. E / 5.B
3. CARVALHO F.H; PIMENTEL S. M. R. A clula. Barueri: Manole, 2007.
Aspectos gerais de estrutura celular
A clula a unidade bsica da vida em que existe uma complementaridade entre estrutura e funo. Essa afirmativa faz parte de uma teoria, denominada teoria celular, estabelecida por Schleiden e Schwann, em 1838-39. A primeira observao de uma clula, no entanto, j havia ocorrido em 1665, ocasio em que, ao examinar um pedao de cortia em um microscpio rudimentar, Hooke dera o nome de clula aos inmeros compartimentos que observara nesse material. Esses compartimentos, na realidade, representavam espaos (celas) ocupados por unidades mortas. Apesar dos fragmentos celulares poderem at desenvolver algumas atividades importantes, somente a clula tem a capacidade de manter vida e de transmiti-la. Pode-se, pois, concluir que os vrus no so unidades de vida, porque no podem se manter, independentemente da clula que infectam. As clulas surgem apenas de outras clulas preexistentes. As formas mais simples de vida so clulas solitrias (organismos unicelulares), enquanto as formas superiores contm associaes de clulas, constituindo colnias de organismos unicelulares mais complexos. Os organismos unicelulares podem ser, estrutural e funcionalmente, mais simples como bactrias, ou mais complexos, como protozorios. Nas associaes de clulas com diferentes especialidades ou diviso de trabalho estar ocorrendo uma contribuio para a sobrevivncia do individuo. O que diferencia colnias de organizaes unicelulares de organizaes multicelulares que, nestas ltimas, as clulas de mesmo tipo podem se apresentar ligadas por uma matriz extracelular, adeses entre membranas ou, ainda, pontes citoplasmticas. As clulas so revestidas por uma membrana plasmtica, tambm denominada plasmalma, de constituio lipoproteica. Clulas mais simples no apresentam ncleo (procariotos), enquanto as mais complexas contm um ou vrios ncleos (eucariotos). Clulas eucariotas so produtos posteriores da evoluo, tendo desenvolvido compartimentalizao do material gentico no ncleo, separado dos constituintes citoplasmticos. As clulas de eucariotos contm maior quantidade de DNA do que as de procariotos. Clulas humanas, por exemplo, contm cerca de 1.000 vezes mais DNA do que clulas bacterianas. Por outro lado, nas clulas eucariotas, dada a sua complexidade, o material gentico requer uma regulao (controle) muito mais complexa do que a das clulas procariotas. Entre o ncleo e o plasmalma existe uma substncia aparentemente amorfa e homognea, se examinada em microscpios mais simples, na qual se
distribuem corpsculos de diversas formas e tamanhos, compartimentalizados por membranas lipoproteicas, as organelas citoplasmticas. Como j mencionado anteriormente, foi apenas com o advento do microscpio eletrnico e de metodologias bioqumicas e fisiolgicas que o conhecimento da subestrutura dessas organelas e de seus atributos funcionais pode ser estabelecido. Ao microscpio de luz, no entanto, podem ser evidenciadas, com metodologia apropriada, regies ocupadas por mitocndrias, lisossomos, peroxissomos, cloroplastos, complexo de Golgi, centrolos vacolos e grnulos de secreo. Os componentes que so encontrados nas clulas podem at ser catalogados como comuns a muitas delas, mas sua estrutura varia conforme cada tipo particular de clula. Embora clulas animais e vegetais tenham muitas caractersticas em comum, uma diferena fundamental a presena de cloroplastos em clulas vegetais, o que lhes permite realizar a fotossntese. Alm disso, clulas vegetais so revestidas por uma rgida parede contendo celulose e outros polmeros.
Formas e tamanhos celulares As clulas podem apresentar estrutura e forma variadas, geralmente associadas a especializaes funcionais. As clulas contm muitas molculas diferentes que interagem em ambiente aquoso e que so compartimentalizadas por membranas lipoproteicas. No estabelecimento de uma forma celular, a organizao de um componente, o citoesqueleto, composto por redes de fibras ou filamentos proteicos, exerce um papel preponderante. De modo geral, as formas celulares dependem da tenso superficial, da viscosidade do protoplasma, da ao mecnica que exercem as clulas contguas, da rigidez da membrana plasmtica e da especializao funcional da clula. A maioria das clulas, especialmente de organismos multicelulares (metazorios), exibe uma forma fixa e tpica. H, no entanto, clulas com forma mutvel, como vrios protozorios e leuccitos. Dentre as clulas de forma fixa, existem aquelas em que a forma regular, seja esfrica [ex., vulo ou linfcito humano], prismtica [ex., clulas vegetais] ou irregular tpica [ex., alguns tipos de clulas vegetais, protozorios, espermatozoides, neurnios, astrcitos, clulas caliciformes e clulas descamadas da mucosa bucal e vaginal.] O tamanho celular oscila entre amplos limites. A maioria das clulas atinge poucos micrmetros de dimetro ou comprimento. H, no entanto, clulas muito maiores, como o vulo humano, com 0,2 mm de dimetro, e vulos de aves, com vrios milmetros de dimetro.
Generalidades sobre o ncleo interfsico
O ncleo, sendo mais facilmente corvel do que os outros componentes celulares e, tambm, graas a seu tamanho, foi descoberto mais cedo, como parte integrante das clulas eucariotas, tendo sido descrito em 1833, por Brown. O estudo do citoplasma foi posterior, pelas dificuldades tcnicas j mencionadas. medida que as tcnicas de estudo foram se aprimorando, foi sendo estabelecida a importncia vital do ncleo para a vida celular, culminando-se com a comprovao de que suas principais funes seriam a de transmisso de caracteres hereditrios e a de superviso da atividade metablica da clula. O ncleo se forma a partir de outro ncleo preexistente, por diviso, que pode ser sincronizada, ou no, com a diviso celular. O ncleo se acha presente em todas as clulas dos eucariotos, exceo daquelas que o perderam em alguma etapa de sua vida (ex.: eritrcitos de mamferos). Nos procariotos, embora no ocorra um ncleo tpico, o DNA se distribui numa regio bem definida, com morfologia caracterstica, denominada nucleoide. Tanto a forma como a posio do ncleo so influenciadas pela prpria forma da clula e pelas condies morfolgicas e funcionais do citoplasma. Nas clulas esfricas e cbicas, o ncleo apresenta forma geralmente esfrica, nas prismticas e fusiformes, elipsoidal ou alongado e, em ambos os casos, est posicionado no centro da clula. Nos leuccitos, pode ter forma bastante irregular. Nos espermatozoides, a forma nuclear pode ser alongada, ou ento, ser irregular, variando conforme o grupo animal. Em lepidpteros e em cigarrinhas-das-pastagens, as clulas glandulares apresentam ncleos estrelados. Em clulas glandulares de outros organismos, geralmente o ncleo se localiza na poro basal celular. Em clulas adiposas de vertebrados, o ncleo alongado e deslocado pelos vacolos de gordura para a periferia celular. A maioria das clulas mononucleada, porm, em hepatcitos, msculo estriado, clulas somticas de muitas espcies de insetos e clulas em cultura, pode ocorrer mais de um ncleo. O tamanho do ncleo tambm pode ser variado, correlacionado ao seu contedo de DNA e ao grau de ploidia da clula, bem como sua atividade funcional, que implica em contedos variveis de RNA e protenas nohistnicas.
MOLCULAS IMPORTANTES PARA A COMPREENSO DA CLULA E DO SEU FUNCIONAMENTO
A anlise e a compreenso da clula e de todo o seu funcionamento vm ganhando importncia nas mais diferentes reas do conhecimento. Todas as formaes da rea biolgica e da sade requerem, como ponto de partida, essa compreenso. Alm destas, o avano da cincia, a crescente importncia da Biologia molecular e a necessidade de um entendimento interativo dos fenmenos da natureza tm agregado a esse grupo profissionais das reas de engenharia, qumica, fsica, jornalismo, direito etc. Essa agregao, cada vez mais nmerosa, faz com que deva existir um conhecimento da clula como uma estrutura dinmica, fisiologicamente ativa, organizada e funcional. Deste modo, inicia-se o estudo da clula a partir de seus constituintes qumicos, principalmente aqueles com organizao macromolecular e que exercem papis fundamentais na sua estrutura e funcionamento.
gua Formada por dois tomos de hidrognio e um de oxignio, trata-se de molcula imprescindvel para a vida em nosso planeta. Em alguns organismos, ela pode representar quase 100% da massa da matria fresca (98% nas guas-vivas) e chega, no ser humano, em torno de 65% da massa corprea do indivduo adulto. A simples presena da gua garante que as demais molculas formem o fluido celular e atinjam seus destinos, seja de forma casual, seja de forma mediada por receptores ou outras maneiras de reconhecimento e de movimentao. A troca gasosa nas clulas animais s se d a partir da dissoluo de oxignio e gs carbnico no sangue e no citosol. Em plantas, a gua responsvel pela presso de turgor que viabiliza o crescimento celular e, por meio de sua maior ou menor disponibilidade no vacolo, pode participar de fenmenos mais complexos, como a abertura e o fechamento das clulasguarda dos estmatos. Normalmente, uma reduo em 20% de seu contedo pode provocar a morte celular. Em contrapartida, em muitas sementes, seus nveis atingem valores inferiores a 10% e isso possibilita que tais estruturas de propagao permaneam em um estado denominado quiescente, com baixssimo metabolismo, e que possam, quando a gua for novamente disponibilizada, germinar e originar um novo indivduo. Em consequncia dessa polaridade, as molculas de gua se atraem umas s outras, criando novas interaes presentes tanto no estado slido quanto no lquido. Essas interaes, denominadas ligaes ou pontes de hidrognio, dificultam a separao entre as molculas e fazem com que haja a necessidade de uma maior quantidade de energia para que ocorra essa
separao. Em outras palavras, isso determina que o seu ponto de fuso e de ebulio seja muito superior ao de outras molculas cuja massa maior ou que apresentam a mesma forma geomtrica. Devido a sua natureza polar, a gua pode atrair regies tambm polares de outras molculas, isto resultando em uma separao dessas molculas, com consequente dissoluo das mesmas. De forma simplificada, imagine i quando uma colher de sal de cozinha (Na+CI-) colocada em um copo com gua: os ons Na+ so atrados pelo oxignio e os ons CI-, pelos hidrognios, havendo assim a dissociao do sal. Raciocnio semelhante pode ser feito para o acar, que apresenta ligaes covalentes, mas tambm atrado pelos plos opostos da molcula de gua e dissolvido na mesma. Desse modo, substncias polares so tambm denominadas hidroflicas. Em muitos casos, a atrao entre a molcula de gua ou uma hidroxila pertence a uma molcula qualquer e a outra molcula polar de tal ordem que a distncia entre elas torna-se pequena o bastante ( entre 0,2 e 0,3 nm) para originar as chamadas ligaes (pontes) de hidrognio e todas as consequncias fsico-qumicas que isso representa. As ligaes de hidrognio tambm ocorrem entre molculas que contenham hidrognio ligado, covalentemente, a tomos fortemente eletronegativos (em geral, biologicamente, oxignio ou nitrognio), com tomos muito eletronegativos da mesma molcula ou outra que esteja prxima. Utilizando a mesma lgica, pode-se concluir que as molculas apolares no so atradas pela gua e, por isso, elas so, em geral, insolveis. Por no se misturarem gua, elas so ditas hidrofbicas. As interaes hidrofbicas so importantes na determinao da estrutura de muitas molculas biologicamente importantes. Finalmente, pode-se classificar um tipo de molculas que apresenta duas regies distintas: uma hidrofbica e outra hidroflica (sabes, por exemplo). Essas molculas so denominadas anfipticas. Sua regio polar pode interagir com a gua e a apolar, apenas com outras molculas apolares.
Molculas e ons inorgnicos A exemplo da gua, outras molculas inorgnicas e ons fazem parte dos organismos vivos, alm do mundo mineral. Esse grupo de substncias, chamado genericamente de sais minerais, apresenta mltiplas funes: quando insolveis, podem ter funo estrutural (p. ex.: clcio e fosfato nos ossos de vertebrados); podem se associar a molculas maiores, como pigmentos (magnsio na clorofila) e protenas (ferro na hemoglobina; ferro e enxofre nos citocromos); podem exercer papel tamponante (bicarbonatos no sangue,
fosfato no citosol), na transferncia de energia qumica (fosfato do ATP), nos impulsos nervosos e equilbrio osmtico das clulas (sdio e potssio), contrao muscular (clcio) e uma infinidade de outras funes.
Molculas orgnicas As molculas orgnicas que fazem parte dos organismos vivos so de natureza bastante variada. Entretanto, em sua maioria, so formadas apenas por seis elementos qumicos: carbono, hidrognio, oxignio, nitrognio, fsforo e enxofre (CHONPS). Esses elementos, associados a outros que aparecem com menor frequncia, formam as substncias necessrias para a vida no planeta, juntamente com a gua e os sais minerais. Dentre os compostos orgnicos, a associao de molculas para a formao de polmeros ou outras molculas maiores comum e as principais classes de substncias dessa natureza so os carboidratos, os lipdios, as protenas e os cidos nucleicos. As demais molculas, a exemplo do que ocorre com os sais minerais, podem ter natureza diversa e exercer mltiplas funes. Mesmo para os seres heterotrficos, grande parte delas pode ser sintetizada pelo prprio organismo ou serem incorporadas na dieta. Como exemplos de importncia, principalmente para o homem, podem ser citados alguns aminocidos noproteicos como a carnitina (que auxilia a entrada de cidos graxos nas mitocndrias) e a ornitina (participa no ciclo da uria); as vitaminas (a maioria delas necessria na dieta) e os hormnios (sintetizados pelo prprio organismo). As vitaminas so necessrias em pequenas quantidades e auxiliam em inmeros processos metablicos. Podem ser subdivididas em hidrossolveis ou solveis em gua (vitaminas do complexo B [B1 ou tiamina, B2 ou riboflavina, niacina, piridoxina, cido pantotnico, biotina, cido flico e B12 ou cobalamina] e a vistamina C) e lipossolveis ou solveis em lipdios (vitamina A, D, E e K). H ainda outras substncias relacionadas s vitaminas, como a colina, o cido p-aminobenzoico, cido lipoico e inositol.
Carboidratos Carboidratos, sacardeos ou acares podem ser definidos quimicamente como poliidroxialdedos ou poliidroxicetonas. Deste modo, os mais simples possuem trs carbonois, com dois grupos hidroxila e um grupo carbonila.
Monossacardeos Biologicamente, encontram-se monossacardeos com 3 7 tomos de carbono que, segundo sua origem aldedica ou cetnica, podem ser classificados em aldoses ou cetoses. Podem tambm receber o prefixo que corresponde quantidade de carbonos da molcula (trioses, tetroses, pentoses, hexoses e heptoses). A seguir so listados alguns monossacardeos bastante abundantes e comuns nos organismos vivos, sua ocorrncia e importncia celular:
a) Pentoses: . ribose e desoxirribose( aldoses) importantes constituintes dos cidos nuclicos (RNA e DNA respectivamente). . xilose e arabionose (aldoses) presentes em glicoprotenas e em paredes celulares de muitas plantas. . ribulose (Cetose) importante na incorporao de CO-2 na fotossntese.
b) Hexoses: . glicose (aldose) tambm conhecida como dextrose, bastante abundante. nica fonte de energia utilizvel pela maior parte dos organismos anaerbios e tambm por alguns rgos e tecidos de animais mesmo em aerobiose (crebro humano por exemplo). produto primrio da fotossntese dos vegetais e est presente em abundncia em muitas frutas onde forma, juntamente com a frutose, o dissacardeo sacarose. . frutose (Cetose) tambm conhecida como levulose, o mais doce dos aucares ( quase duas vezes mais que a glicose) e est fortemente presente em frutas e no mel, muitas vezes ligadas a glicose para formar o dissacardeo sacarose. . galactose (aldose) difere da glicose apenas pela posio da hidroxila do Carbono 4 ( por isso denominada epmero-f da glicose) e seu metabolismo no fgado origina esse acar. Est presente em muitas glicoprotenas, glicolipdios e em muitas paredes celulares vegetais.
Dissacardeos Ligao de dois monossacardeos ocorre entre hidroxilas, com formao de gua. chamada ligao glicosdica e forma os dissacardeos. Os mais comuns so: a) b) c) d) celobiose, maltose lactose sacarose
Oligossacardeos Nome dado molculas que apresentam at cerca de 20 resduos de monossacardeos quando hidrolisadas totalmente. Muitas molculas de glicoprotenas e glicolpidios tm sua poro glicdica composta por oligossacardeos.
Polissacardeos Muitas vezes formados por milhares de unidades monossacardicas, os polissacardeos so importantes macromolculas para os seres vivos de todos os reinos biolgicos. So classificados em homopolissacardeos ou heteropolissacardeos, segundo o produto de sua hidrlise total ser apenas um tipo de monossacardeos ou mais de um. Os polissacardeos mais conhecidos tm papel de reserva energtica e so importantssimos na alimentao dos organismos heterotrficos. Os polissacardeos de reserva nas plantas o amido, enquanto que nos animais, o glicognio, por exemplo, de homopolissacardeos da A- glicose.
Lipdios Contrariamente aos carboidratos, no h nenhum grupamento qumico caracterstico para todos os lipdios. No formam polmeros e a caracterstica que os une a sua pequena solubilidade em gua. Dadas algumas semelhanas estruturais, pode-se classific-los em diferentes grupos, dos quais se destacam:
- Possuidores de cidos graxos:
a) Ceras. b) Gorduras neutras. c) Fosfolipdios. d) Esfingolipdios. - Esteroides derivados do ciclopentanoperidrofenantreno - Terpenoides derivados do isopreno.
cidos graxos so cidos carboxlicos com 4 ou mais tomos de carbono. Em termos biolgicos, so predominantes aqueles com 14 a 22 tomos de carbono em cadeia que pode ser saturada (s simples ligaes) ou instaurada (uma ou mais duplas ligaes) com par de carbonos (sua sntese acontece com a adio de 2 em 2 tomos). Possuem carter anfiptico, ou seja, uma regio polar (do grupo carboxila, que pode se ionizar), e uma regio apolar ou hidrofbica, representada pela poro hidrocarboneto (apenas C e H) da molcula.
a. Ceras So steres de cidos graxos e lcoois graxos. As ceras so bastante apolares e importantes reservas de organismos marinhos, protetoras da pele e anexos de muitos vertebrados (plos e penas de animais aquticos, por exemplo), da superfcie das folhas de muitas espcies, protegendo-as inclusive do ataque de patgenos.
b. Gorduras neutras Correspondem classe mais abundante de lipdios, normalmente presentes como a principal fonte de energia a ser utilizada pelos organismos animais. Um homem adulto, via de regra, tem gorduras neutras suficientes para seu suprido de energia (ATP produzido na respirao aerbica) por vrias semanas enquanto a reserva de acar (glicognio) supre o organismo por cerca de um dia.
c. Fosfolipdios
So molculas resultantes da ligao de um diacilglicerol e um grupo fosfato. Com isso, o carter apolar apresentado pelos triaciglicerois perdido e volta-se ao carter anfiptico: uma regio hidrofbica (apolar) representada pelos dois cidos graxos esterificados no glicerol e uma regio hidroflica (polar) do grupo fosfato.
d. Esfingolipdios So formados pela ligao de um cido graxo com o grupo amina do amino-lcool esfingosina (que tem uma cadeia com 18 tomos de carbono e uma dupla ligao), formando uma amida.
Esteroides Derivados do ciclopentanoperidrofenantreno, que tem uma estrutura cclica composta por 4 anis (um com cinco tomos de carbono e os demais com seis). Comumente apresentam pelo menos uma hidroxila (R-OH) e, muitas vezes, grupo(s) carbonila (R-C=0).
Terpenoides Grupo de lipdios derivados do isopreno: Os carotenoides so os representantes mais comuns desse grupo. Dentre eles, a vitamina A, presente em pequenas quantidades no organismo humano e o B-caroteno , presente dos cloroplastos das clulas vegetais ou acumulado como reserva em algumas plantas (cenoura por exemplo) so os mais abundantes e representativos.
Protenas So polmeros de aminocidos que tm funo biolgica.
Aminocidos H vinte aminocidos que podem participar da formao das protenas e serem incorporados durante a sua sntese nos ribossomos. Todos eles so aaminocidos e se apresentam na forma L (o carbono a assimtrico).
Em pH [acido (menor que 7), o grupo carboxila se ioniza, ou seja, perde seu prton (em pH2 a 2,5, 50% deles j se encontram ionizados), enquanto o grupo amina est protonado (tem fraco carter cidom e a maioria fica ionizada apenas em pH acima de 9).
Peptdeos A ligao entre o grupo carboxila de um aminocido (aa) e o grupo amina de outro denominada ligao peptdica e forma dipeptdeos (2 aa), tripeptdeos (3 aa), oligopeptdeos (vrios AA ) e polipeptdeos (dezenas a centenas de aminocidos).
Protenas So as mais abundantes molculas presentes nos organismos vivos exceo da gua, podendo perfazer 50% da matria seca deles. No ser humano, por exemplo, cerca de 15% da massa corprea vem das protenas. Grande parte das protenas formada apenas por aminocidos e, por isso, elas recebem o nome de protenas simples. Entretanto, h protenas denominadas protenas conjugadas, pois possuem outras molculas ou tomos (grupo prosttico), alm dos seus aminocidos formadores. As propriedades fsico-qumicas e as funes das protenas tm relao direta com a sua composio em aminocidos. Por esse motivo, de extrema importncia o conhecimento da sequncia com que eles so incorporados molcula durante a sntese proteica para formar a estrutura tridimensional do polipeptdeo. Didaticamente, a sequncia de aminocidos de uma protena denominada estrutura primria da protena. Em decorrncia, medida que novos aminocidos so incorporados molcula de protena que est sendo sintetizada, comear a haver a atrao entre o oxignio e o H de ligao peptdicas distintas. Essas interaes determinam uma estrutura especial caracterstica. Que denominada estrutura secundria da protena. Assim, estrutura secundria de uma protena a forma que ela toma em decorrncia das pontes de hidrognio entre oxignio e hidrognio ligado ao nitrognio de ligaes peptdicas distintas. A estrutura secundria, resultante das interaes por ligaes de hidrognio especficas dos tomos das ligaes peptdicas, posiciona os grupos R dos diferentes aminocidos, de tal modo que pode provocar novos tipos de interao. Assim, a estrutura tridimensional total de molcula, agora
com o somatrio dessas interaes, denomina-se estrutura terciria da protena. As interaes mais comuns entre os diferentes grupos so: a) ligao ou interao eletrosttica: um grupo R carregado com a carga contrria a outro grupo R o atrai; b) ligao ou interao covalente: especfica da cistena, cujo grupamento sulfidrila (-S-H) facilmente oxidado com outro grupamento S-H de outra cistena formando pontes S-S- (pontes dissulfeto) e dando origem ao conjunto chamado cistina (duas cistenas ligadas por uma ponte dissulfeto); c) ligaes de hidrognio: ocorrem entre um tomo muito eletronegativo de um grupo R e outro tomo eletronegativo ligado a hidrognio de um outro grupo R; d) ligao ou interao hidrofbica: realizada entre grupos R apolares. Geralmente, os aminocidos portadores de grupos R apolares ocupam a regio mais interna da protena e interagem seus grupos R. Os aminocidos com grupo R polares ocupam a poro mais exterior da molcula, na qual podem interagir com a gua. Algumas protenas, para adquirirem seu estado funcional, necessitam mais de uma cadeia polipeptdica (com sua estrutura tridimensional definida). Ao estado funcional, que conta com as estruturas tercirias de dois ou mais polipeptdeos unidos, geralmente, a um grupo prosttico, d-se o nome de estrutura quaternria da protena. O exemplo mais conhecido de protena que tem estrutura quaternria a hemoglobina, formada por 4 cadeias (duas denominadas a e duas b), cada uma delas envolvendo um grupo heme contendo ferro.
cidos nucleicos So polmeros de nucleotdeos que, por sua vez, so molculas formadas por uma base nitrogenada heterocclica, uma pentose (acar) e um fosfato. juno da base com o acar (sem o fosfato) d-se o nome de nucleosdeo. Segundo o tipo de acar formador, tem-se o cido ribonucleico (RNA ou ARN) ou cido desoxirribonuclico (DNA ou ADN).
Nucleosdeos OS nucleosdeos podem ser hidrolisados em uma pentose e uma base heterocclica.
A pentose a ribose para os nucleosdeos de RNA ou a desoxirribose para os chamados desoxiribonucleosdeos, e tem como diferena o carbono 2 do acar no hidroxilado: As bases heterocclicas podem ser derivadas de dois compostos: purina e pirimidina. Segundo essa origem, tem-se as bases pricas (adenina e guanina) e as bases pirimdicas (citosina, uracila e timina).
Nucleotdeos ou nucleosdeos-fosfato A adio de fosfato ao nucleosdeo origina um nucleotdeo ou um nucleosdeofosfato. Adicionar um (nucleosdeo monofosfato), dois (nucleosdeo difosfato) ou trs fosfatos (nucleosdeo trisfosfato). O fosfato adicionado ao carbono 5 do acar, com liberao de gua.
cido desoxirribonucleico DNA ou ADN Polmero formado por monmeros dos nucleotdeos (originrios de bases pricas), de bases pirimdicas. o responsvel pela informao gentica contida nos organismos vivos e pela sua transmisso s clulas filhas. Em envoltrio nuclear, alm de existir em pequena quantidade nas mitocndrias e cloroplastos. Nos procariotos, ocupa preferencialmente uma regio denominada nucleoide, apesar de no estar fisicamente delimitado por membrana. No DNA, h o pareamento de uma fita com outra, formando uma dupla fita ou dupla hlice. Essa interao, estvel apenas quando no sentido antiparalelo ( uma fita 5 --->3 e a outra no sentido 3> 5), feita por meio de ligaes de hidrognio entre uma base prica com uma pirimdica. O pareamento se d entre uma Adenina e uma timina ( duas pontes de hidrognio) ou entre uma guanina e uma Citosina (trs pontes de hidrognio). A reao de polimerizao do DNA complexa e envolve muitas enzimas, com destaque para as DNA polimerases. Se d sempre no sentido 53 da molcula me e semi conservativa, ou seja: cada fita da molcula original serve de molde para as novas molculas formadas. O processo de sntese de DNA a partir do DNA j existente na clula denominado replicao e ocorre em quase sua totalidade em uma etapa da interfase do ciclo celular denominada fase S .
cido ribonucleico RNA ou ARN
Polmero formado por monmeros dos nucleotdeos. Sua sntese ocorre a partir da molcula de DNA que lhe serve de molde e se d no sentido 5 3 dessa molcula. Assim dos eucariotos, onde ocorre a sua transcrio a partir do DNA. O RNA transportador (RNAt) responsvel pelo reconhecimento, ligao e transporte dos aminocidos presentes no citoplasma para a sntese proteicas (traduo). A outra famlia de RNA, transcrito pelo DNA, refere-se aos inmeros RNA mensageiros (RNAm). Normalmente fita simples, esses RNA apresentam tamanho variado conforme a protena que codificam para a sntese. Em procariotos, tm uma srie de bases iniciais que os auxilia na ligao com o ribossomo e no correto posicionamento para o incio da sntese proteica. Em eucariotos, a sntese mais complexa e sua extremidade 5 protegida por uma srie de protenas. Finalmente, o DNA transcreve RNA ribossomal (RNAr) que, juntamente com dezenas de protenas, forma o ribossomo de procariotos ou de eucariotos. O RNA ribossomal sintetizado em regio repetitiva do DNA (que tem bases suficientes para transcrever simultaneamente dezenas de molculas desse RNA), denominada regio organizadora do nuclolo.
MICROSCPIAS As clulas so pequenas e complexas, sendo difcil a observao de sua estrutura e composio macromolecular. Ainda mais difcil elucidar como elucidar como funcionam seus diversos sub-compartimentos. H uma grande variedade de procedimento experimentais para esse estudo, sendo que as possibilidades e limitaes dessas tcnicas tm determinado, em parte, nossa concepo atual da clula e dos tecidos. Portanto, para compreender melhor a clula,, necessria a compreenso geral dos mtodos que foram desenvolvidos para o seu estudo.
Microscopia de luz Os efeitos da interao da luz com o meio por ela percorrido decorrem de sua natureza corpuscular ou fotnica e ondulatria (radiao eletromagntica). Dois efeitos, absoro e refrao, ocorrem como consequncia da interao da frente de onda, fotnica ou eletromagntica, com os componentes do material a ser analisado. Pode-se dizer que absoro e refrao so duas faces do
mesmo fenmeno e so importantes fatores a serem considerados na formao das imagens aos microscpios. Os microscpios so equipamentos que tm por objetivo produzir imagens aumentadas de objetos to pequenos que so importantes vista desarmada ou que, vistos a olho nu, no revelariam aspectos texturais mais detalhados. A formao de imagens pelos microscpios fundamenta-se em um sistema de lentes combinadas, que so colocadas de forma a ampliar a imagem do objeto. importante lembrar mais uma vez que, para que os objetos sejam vistos microscopia, dois requisitos fundamentais tm de ser cumpridos: a interao da luz com o espcime tem de gerar absoro ou refrao dos raios de luz, criando contrastes entre o objeto e o meio que envolve. A teoria da formao da imagem nos microscpios regida pelas Leis da Fsica. Entretanto, como as lentes dos microscpios comportam-se como sistemas biconvexos, ou seja, lentes convergentes, a formao da imagem depende da posio do objeto em relao ao plano focal (f) ou ao centro focal (c) da lente. Assim, teremos imagens reais invertidas ou virtuais direitas.
Microscopia de contraste de fase Esse tipo de microscopia baseia-se nos princpios da difrao da luz, isto , o caminho do feixe luminoso, na formao da imagem por esse tipo de microscpio, sofre um retardo ptico, permitindo assim que seja possvel observar materiais biolgicos sem a colorao. Esse tipo de microscopia foi desenvolvido pelo holands Zernike, na dcada de 1950. Isso permitiu grandes avanos nos estudos de clulas vivas pois, a partir desse tipo de microscpio, pode-se observar preparados no corados que, microscopia de luz convencional, apresentam-se transparentes ou com pouco contraste.
Microscopia de contraste interferencial Esse tipo de microscopia funciona base de prismas pticos posicionados no caminho da luz. Esses prismas modificam a fase da onda luminosa, que aparecer contrastando-se com o meio em que se encontra o material a ser analisado. O microscpio de interferncia requer uma construo especfica, diferente do microscpio de contraste de fase, que apenas apresentava anis do caminho da luz. Esse microscpio no requer objetivas especiais, mas o revlver deve conter ranhuras para alojar os prismas de interferncia, de modo a gerar as cores de interferncia, que promovem a visualizao do material. A
microscopia interferencial mais difundida na rea biolgica a chamada microscopia de Normarski. Esse tipo de contraste interferencial trabalha com a defasagem dos comprimentos de ondas.
Microscopia de polarizao O microscpio de polarizao apresenta dois prismas, ou filtros, chamados polarizador e analisador. Esses filtros esto posicionados estrategicamente entre a fonte de luz e o condensador (filtro polarizador) e entre a objetiva e a ocular (filtro analisador). Na microscopia de luz comum, os feixes de ondas luminosas apresentam direo de vibrao em todos os planos. Os filtros polarizadores promovem a seleo de apenas um plano de direo de vibrao das ondas luminosas, o que conhecido por plano da luz polarizada (PPL). As anisotropias pticas so fenmenos de ordem espectral conhecidos por dicrosmo e birrefringncia. O dicrosmo ocorre quando apenas um filtro polarizador colocado no sistema. Ele expresso pela diferena de absoro do objeto em duas direes de deslocamento do feixe de luz no objeto (um perpendicular ao outro). A birrefrindncia ocorre quando cruzamos perpendicularmente os dois filtros, o polarizador e o analisador, dependendo da diferena entre os ndices de refrao do objeto.
Microscopia de campo escuro Os microscpios de campo escuro apresentam um sistema especial de condensador. Esse condensador permite luz ficar, de tal modo inclinada, que no atravessa o material. A luz atinge o espcime a ser analisado, e somente os feixes desviados pelo objeto percorrem o resto do sistema, isto , as objetivas e as oculares, formando a imagem. Esse tipo de microscopia utilizado somente para pequenos materiais, como plncton, bactrias, pequenos cristais, gros de plen e outros objetivos transparentes microscopia de campo claro convencional.
Microscopia de fluorescncia A microscopia de fluorescncia est baseada na propriedade fsica de algumas substncias absorverem a luz, em um determinado comprimento de onda, e emitirem luz, com comprimentos de onda maiores e nveis energticos mais
baixos. Existem componentes celulares ou moleculares naturalmente fluorescentes, e outros que podem se ligar a substncias fluorescentes ( fluorocromos). Uma estrutura fluorescente dever, em ltima anlise, emitir brilho contra um fundo escuro. O microscpio de fluorescncia diferente dos demais aparelhos j citados , por precisar de um sistema ptico que interaja pouco com a luz. A luz que alimenta seu sistema ptico uma luz de mercrio de alta presso, cujos picos mais caractersticos variam entre 312 e 579 nm. Outra peculiaridade do microscpio de fluorescncia so os sistemas de filtros requeridos para detectar o brilho do material contra o fundo negro. So os chamados filtros de excitao e filtros de barragem. Os filtros de excitao localizam-se logo aps a sada da fonte de luz e antes do condensador, tendo como finalidade selecionar o comprimento de onda desejado. Os filtros de barragem localizam-se entre a objetiva e a ocular, isto , aps o objeto, tendo como funo primordial deixar passar somente a luz fluorescente emitida pelo espcime analisado, barrando assim a luz de excitao. Assim, o material fluoresce contra um fundo escuro.
Microscopia confocal a laser Esse microscpio, desenvolvido recentemente e comercialmente disponvel a partir de 1987, tem suas peculiaridades, permitindo, por exemplo, a observao de materiais espessos, sem colorao prvia, vivos ou pr-fixados. Esse aparelho pode obter imagens de planos focais especficos ou cortes pticos. Esses cortes pticos so, ento, estocados em um computador e podem ser utilizados na reconstruo tridimensional e visualizao da estrutura como um todo. Desta forma, os microscpios tradicionais trabalham com imagens analgicas, enquanto o confocal a laser, com imagem digital. Esse aparelho trabalha com a ptica de um microscpio de fluorescncia, mas utiliza laser como fonte de luz alimentadora do sistema. O microscpio confocal permite o detalhamento de estruturas subcelulares que no apresentam limite de resoluo compatvel ao da microscopia de luz fluorescente convencional, como microtbulos e outros elementos fibrilares do citoesqueleto e elementos finos da matriz extracelular.
Aplicao da microscopia de luz nas anlises quantitativas e padres texturais Para a quantificao de elementos ou macromolculas na clula, existem vrios sistemas. O principal deles o conhecido citofotmetro ou microespectrofotmetro. Esse aparelho comporta-se como um microscpio
comum, em seu sistema ptico, entretanto ele apresenta uma fotoclula que capta os sinais luminosos e os transfere para um terminal fotomtrico ( semelhana de um espectrofotmetro utilizado nas dosagens bioqumicas), que transformar os valores absorciomtricos em valores quantitativos. A citofotometria tem tido ampla aplicao na rea de Biologia celular, principalmente para a quantificao de DNA e de protenas, dente outras macromolculas. Aos microscpios de luz, podem ainda ser acoplados sistemas analisadores de imagens, consistindo basicamente de microcmeras de vdeo que captam a imagem e a transferem para um terminal de computadores tipo PC. Softwares especficos permitem a anlise de padres texturais dos componentes morfolgicos nos tecidos, como a distribuio das massas cromatnicas em ncleos ou, at mesmo, padres de arranjo das fibras da matriz extracelular.
Microscopia eletrnica Os princpios que regem a ptica da microscopia eletrnica so os mesmos descritos para a microscopia de luz, sendo que o primeiro apresentase de maneira invertida, isto , a fonte geradora dos feixes de eltrons est na poro superior do aparelho. A microscopia eletrnica, como a microscopia de luz, tambm apresenta vrios tipos de aparelhos com especificidades quanto ao funcionamento e utilizao. Basicamente, pode-se dizer que existem duas formas de microscopia eletrnica: a microscopia eletrnica de transmisso e a microscopia eletrnica de varredura.
Microscopia eletrnica de transmisso O funcionamento do microscpio eletrnico de transmisso est relacionado, principalmente, natureza dos feixes de eltrons, utilizados na formao da imagem. Nesse tipo de microscopia, os eltrons tm de interagir com o objeto para fornecerem a imagem. O objeto deve ser exatamente fino para permitir a passagem dos eltrons.
Microscopia eletrnica de alta voltagem A utilizao do microscpio de alta voltagem veio, de certa maneira, revolucionar a Biologia estrutural. Pois graa a esse aparelho, muitas estruturas subcelulares puderam ser descritas, como, por exemplo, a
organizao tridimensional dos componentes do citoesqueleto, graas a possibilidades de se utilizarem espcimes com espessura, na casa dos micrmetros, e seria possvel na microscopia eletrnica de transmisso convencional.
Microscopia eletrnica de varredura A microscopia eletrnica de transmisso fornece informaes a partir de cortes ultra-finos de clula ou tecidos, pois a obteno da imagem depende da interao dos eltrons com o material, ao ser atravessado por eles ou no. O microscpio eletrnico de varredura pode velar feies topogrficas de uma superfcie em grande nitidez de detalhes. Esse aparelho fornece imagens tridimensionais, tanto de objetos relativamente grandes, como vermes e insetos, quanto de clulas livres, como tecidos animais e vegetais ou, at mesmo, embries e fragmentos geolgicos em anlises de granulometria e textura de solos.
Microscopia de tunelamento quntico e de fora atmica O microscpio de fora atmica assemelha-se ao de tunelamento quntico, com a diferena que este ltimo apresenta um microespelho e um feixe de laser sobre a agulha. Esse fato permite uma menor agressividade amostra e a deteco de detalhes de superfcies, sem maiores interferncias com a amostra. Esse tipo de instrumento permite tambm a obteno de imagens em soluo ou de sequncias que representam reaes qumicas ou modificaes estruturais ao longo do tempo. Nesse tipo de aparelho tambm pode ser avaliada a estrutura atmica de biomolculas.
MTODOS DE ESTUDO DA CLULA
Para a obteno de um bom preparado citolgico, ou histolgico que seja passvel de anlise ao microscpio, imprescindvel que se tomem alguns cuidados durante todas as etapas do processamento. Esses s cuidados vo desde a escolha e coleta do material, at a montagem do preparado permanentemente em lmina histolgica.
Coleta do material biolgico
Essa etapa de importncia fundamental para o estudo dos componentes celulares e subcelulares, pois nesta fase que so definidas as formas de anlise dos preparados citolgicos.
Montagem total Consiste em coletar o material, que dever ser fino ou transparente o suficiente para que o mesmo possa ser colocado diretamente sobre uma lmina, e, assim, promover as tcnicas subsequentes de fixao e colorao.
Esfregao Clulas livres presentes nos fluidos corpreos, como sangue, linfa, smen, lquor, hemolinfa, podem ser dispostas sobre lmina em fina camada de maneira que possam ser observadas em microscpio de luz. O mtodo de esfregao consiste em colocar uma gota do material a ser analisado sobre uma lmina, e, com o auxlio de uma outra lmina, promover o deslizamento do lquido sobre a primeira lmina, de forma que se faa uma camada do lquido com clulas sobre o vidro.
Espalhamento O espalhamento, erroneamente chamado de esfregao, consiste em promover uma raspagem das camadas superficiais de membranas mucosas com uma pequena esptula ou palitos e, posteriormente, deslizar esse material raspado sobre a superfcie limpa de uma lmina. Esse tipo de prtica muito utilizada para a avaliao de mucosas vaginais, no conhecido exame preventivo de cncer denominado Papanicolau.
Esmagamento Esse mtodo consiste em esmagar, entre lmina e lamnula, o material a ser analisado. Pode-se promover a colorao concomitantemente ao esmagamento, ou promover a retirada da lamnula com auxlio de nitrognio lquido, e fazer a colorao posteriormente. O nitrognio lquido congela rapidamente o material e a lamnula pode ser retirada sem que o material esmagado seja removido da lmina. Esse mtodo de coleta muito eficiente para estudos de divises celulares em tecido com alta taxa de diviso (raiz de cebola, testculos e glndulas salivares de insetos). Tambm, por esta maneira de coleta, pode-se estudar ncleos interfsicos inteiros com a vantagem de
obteno de preparados na ntegra, podendo assim fazer quantificaes de DNA e protenas nucleares.
Decalque O objetivo dessa tcnica de coleta de colocar sobre uma lmina, devidamente limpa, ncleos inteiros de rgos com relativa consistncia mole, como, por exemplo, o fgado, o bao, os rins, o timo. A tcnica consiste em retirar do animal um fragmento do rgo a ser estudado, e a face que foi cortada dever ser levada em soluo salina, para retirada do sangue, e secada em papel de filtro posteriormente. Ao promover esses movimentos, os ncleos ficaro impressos na lmina, podendo ser feitos os passos seguintes de fixao e colorao. Essa tcnica relativamente simples muito til para o estudo de quantidade de DNA, interaes moleculares entre complexos DNA/ protena, textura cromatnica e anlise de imagens dos fentipos nucleares.
Corte histolgico Existem muitos estudos em que a inter-relao entre as clulas e a topografia dos tipos celulares deve ser respeitada. Assim, necessrio promover cortes extremamente finos dos rgos a serem analisados. Para isto, utiliza-se o corte histolgico. A tcnica histolgica consiste na obteno de cortes extremamente finos (na casa dos micrmetros de espessura) e na sua colocao sobre a lmina. Entretanto, para a obteno desses cortes, preciso que o material a ser analisado passe por um tratamento de incluso em parafina, resina ou gelatina, ou simplesmente seja congelado. Com qualquer um desses tratamentos, o material ficar uniformemente duro, podendo ser facilmente cortado pelo micrtomo (equipamento utilizado na obteno dos cortes; para cortes de parafina e historresinas utilizado o micrtomo rotativo manual e para cortes ultrafinos para microscopia eletrnica utilizado o ultramicrtomos).
Fixao biolgica e agente fixadores A fixao constitui uma das etapas mais importantes dos processamentos citolgicos e histolgicos, pois depende de processos fsico-qumicos nos quais os componentes macromoleculares dos tecidos e das clulas passam por um processo de insolubilizao, que inativa os constituintes moleculares dos compartimentos teciduais ou celulares de origem. Em ltima anlise, o processo de fixao biolgica promove uma preservao das caractersticas morfolgicas e macromoleculares dos tecidos ou clulas. A fixao tambm
tem funo impedir a autlise ou degradao bacteriana do material biolgico a ser analisado ao microscpio. A fixao um passo importante de tcnica citolgica e histolgica, pois a anlise satisfatria de um determinado preparado depende da preservao adequada do que se quer analisar. Os fixadores so agentes qumicos das mais diversas funes orgnicas, que reagem quimicamente com os componentes celulares, promovendo a sua estabilizao molecular. Os principais componentes celulares que podem ser preservados so as macromolculas (protenas, cidos nucleicos, polissacardeos e lipdios). Na maioria das vezes, os fixadores agem sobre essas molculas coagulando as ou tornando as insolveis e, consequentemente, precipitando as nos tecidos de origem.
Preparao do material para microscopia eletrnica de transmisso Existem diferenas fundamentais entre o processamento de fragmentos de rgos para a microscopia de luz e para a eletrnica. Sero abordadas, neste tpico, as principais diferenas entre as etapas para a tcnica rotineira de obteno de cortes ultrafinos. As diferenas no processamento residem no fato de serem necessrios maiores cuidados com a preservao do material, visto que o poder da resoluo da microscopia de luz. Estas diferenas referem se ao processo de fixao, incluso, corte e colorao. As clulas e os tecidos biolgicos, assim como toda a matria viva, so constitudos de elementos qumicos de baixssimo peso molecular, como, por exemplo, o carbono, o oxignio, o nitrognio, o hidrognio, dentre outros. Dessa forma, a visualizao da matria orgnica ao microscpio fica comprometida devido a seu baixo contraste. Alm disso, o material a ser examinado ao microscpio de luz deve, em princpio, ser bem fino e transparente, para que a luz possa interagir com este para a formao da imagem. Partindo dessas colocaes iniciais, se faz necessrio preparar o material biolgico por coloraes ou reaes que resultem numa resposta colorida dos elementos a serem analisados para que possamos visualizar o material com maior clareza de detalhes ao microscpio de luz. A citoqumica a rea da Biologia celular e estrutural dedicada aos estudos dos mtodos de colorao dos tecidos e constituintes celulares ou subcelulares, preocupando se no somente com os princpios qumicos das reaes de colorao, mas tambm com os procedimentos protocolares para obteno de preparados a serem avaliados aos microscpios.
Reaes citoqumicas mediadas por ligaes eletrostticas
Nestas reaes, dizemos que, por afinidade eletrosttica, um corante ionizado em uma soluo reage com um substrato (nome dado ao componente a ser avaliado pela reao) de carga inica aposta. Assim, podemos enumerar dois fenmenos citoqumicos: acidofilia e basofilia. Entendese por acidofilia o fenmeno citoqumico no qual um substrato carregado positivamente, chamado de substrato catinico, reage eletrostaticamente com um corante carregado negativamente, dito aninico. Assim, por ligao inica esses dois elementos reagem e formam um composto colorido, que ser evidenciado ao microscpio de luz. Entende-sepor basofilia como o fenmeno citoqumico no qual substrato carregado negativamente, chamado de substrato aninico, reage eletrostaticamente com um corante carregado positivamente, dito catinico.
Reaes citoqumicas mediadas por ligaes covalentes Muitas so a reaes citoqumicas mediadas por ligaes covalentes, principalmente aquelas que necessitam ser facilitadas por uma molcula de componente metlico, que chamado mordente. Os tricrmicos so conhecidos como tcnicas citoqumicas para demonstrao diferencial dos tecidos conjuntivos. Os exemplos mais clssicos de reaes citoqumicas mediadas por ligaes covalentes so, entretanto, a reao de feulgen, para DNA e o teste do cido peridico Schiff (PAS), para polissacardeos neutros. Ambas as reaes so obtidas a partir de um mesmo reagente chamado de Reativo de Schiff, que consiste num leucoderivado do corante Fucsina Bsica. Esse reagente, quando em presena de aldedos livres ou combinados nos tecidos, promove a colorao magenta caracterstica. A reao de feulgen tem, como pr-tratamento do material a ser submetido ao reativo de Schiff, uma hidrlise cida pelo HCI, na qual a molaridade do cido, o tempo e a temperatura podem variar na dependncia do material a ser estudado.* importante saber que essa hidrlise remove da molcula de DNA as bases pricas promovendo, assim, a depurinao do DNA sem as purinas, apresenta grupamentos aldedicos, gerados a partir da instabilidade das desoxirriboses, sendo estes reativos ao reativo de Schiff. O teste do ps muito utilizado para avaliaes citoqumicas de polissacardeos neutros, como o glicognio, o amido e a celulose, alm de glicoprotenas. Baseia-se na capacidade do cido perodo (HIO 4) oxidar as ligaes carbono carbono das sequncias 1 2 glicol dos carboidratos produzindo aldedos. Aps esse tratamento prvio com o cido peridico, o
material ser submetido ao reativo de Schiff e, assim como na reao de Feulgen, os aldedos sero evidenciados em cor magenta.
Reaes citoqumicas mediadas por interaes hidrofbicas Essas reaes so especficas para lipdios no polares como, por exemplo, os triglicerdeos e derivados do colesterol, sendo importante para o estudo de clulas adiposas e de elementos do tecido adiposo ou at mesmo para estudo dos lipdios da bainha de mielina nos nervos e em clulas hepticas. Essas coloraes partem do princpio que os lipdios no polares so altamente hidrofbicos, portanto, as reaes devem ser livres de gua, consequentemente, os corantes a serem utilizados devem ser diludos em solues a base de lcool ou acetona. Assim esses corantes interagem com os lipdios hidrofobicamente. Dentre os corantes especficos para lipdios, podem ser citados o sudan Black, o sudan III e o azul de Nilo.
Citoqumica enzimtica Os estudos citoqumicos de enzimas baseiam-se, principalmente, na possibilidade de averiguar in situ a atividade delas. Para isto, so necessrias alguns pr-requisitos como a preservao da integridade molecular da enzima, com fixaes brandas e/ou cortes por congelao, ou ainda tratamento em bloco. A imunocitoqumica utiliza a especificidade dos anticorpos na localizao de molculas ou de regies de molculas nas clulas ou tecidos. A visualizao dos anticorpos depende do acoplamento deles com marcadores detectveis que podem ser de diferentes tipos, dependendo dos interesses especficos e dos microscpios disponveis para as anlises. A imunocitoqumica consiste em uma excelente ferramenta para estudos sobre a clula, garantir grande especificidade, associada identificao das relaes estruturais entre a molcula de interesse e outros componentes e/ou compartimentos celulares. Normalmente, se marca o ncleo das clulas com um corante ou fluorocromo numa etapa final de contracolorao, para melhor localizar o produto da reao imunocitoqumica.
Anticorpos policlonais x anticorpos monoclonais Quando uma macromolcula isolada, ou um conjunto de molculas, injetada em um animal, ela elcita a produo de anticorpos pelo organismo. Os anticorpos produzidos pelo organismo so especficos contra as diferentes
molculas da mistura ou a diferentes regies de uma mesma macromolcula e esto todos presentes no soro. A especificidade na utilizao desses anticorpos s garantida quando antgeno injetado altamente purificado, sendo os anticorpos denominados monoespecficos. Quando se utiliza soro do animal hospedeiro, que pode ser coelho, rato, camundongo, cabra, burro, cavalo, porco, macaco e, at mesmo, galinha, tem se uma preparao chamada de policlonal, j que os anticorpos so produzidos por diferentes clones de linfcitos. Em contraste, os lintcitos B dos animais utilizados podem ser isolados e fundidos a clulas de um tipo de plasmocitoma (linfcitos B tumorais), adquirindo, destas ltimas, grande capacidade proliferativa. clula hbrida formada denomina-se hibridoma. Aps as etapas de clonagem (isolamento de uma nica clula) e de propagao dos clones que produzem o anticorpo de interesse, obtm-se uma linhagem celular que produz um nico tipo de anticorpo, dirigindo a um nico antgeno que corresponde a uma molcula da mistura inicial ou a um segmento da macromolculas que serviu para imunizar o animal. Embora monoclonais possam ser obtidos a partir de diferentes espcies, os camundongos so os mais comumente utilizados.
Imunocitoqumica direta x imunocitoqumica indireta Para serem localizados, os anticorpos precisam estar acoplados a marcadores que permitam a sua localizao, uma vez que tenham se ligado ao antgeno especfico. Quando o anticorpo marcado por um marcador qualquer e pode ser observado imediatamente, o processo recebe o nome de imunocitoqumica direta. Dessa forma, a estratgia comumente utilizada para evitar essa perda de anticorpos, que so destinados localizao dos antgenos, utilizar um segundo anticorpo (anticorpo secundrio), este sim, acoplado a um marcador, na localizao do anticorpo primrio. Nesse caso, tem-se a imunocitoqumica indireta possibilita tambm uma ampliao do sinal obtido, pois, a cada anticorpo primrio, podem se ligar vrios anticorpos secundrios.
Imunoflurescncia x imunoperoxidase Quando o composto utilizado na marcao do anticorpo um fluorocromo, a imonocitoqumica recebe, s vezes a designao imunoflurescncia. Embora os fluorocromos fluorescena (que emite fluorescncia verde) e rodamina (que emite fluorescncia vermelha) sejam os mais conhecidos dos mercadores utilizados em imunocitoqumica, vrios substitutos com melhores propriedades espectrais ou de estabilidade tm sido disponibilizados comercialmente. No caso desses dois, o reagente utilizado na marcao do
anticorpo o insotiocinato de fluorescena ( FITC) e o isotiocinato de tetrametilrodamina ( TRITC). Alm da necessidade da utilizao de um microscpio de fluorescncia para observao das reaes obtidas com uso de sondas fluorescentes, a preparao obtida tem vida limitada, principalmente devido destruio dos fluorocromos pela exposio luz de excitao.
Fracionamento celular Em eucariotos, grande parte das vias metablicas est compartimentalizada em organelas. Desse modo, se quisermos isolar uma enzima que catalisa uma reao especfica, ou mesmo purificar uma determinada protena que reside em determinada organela, necessrio isolar est ltima das outras organelas celulares. O conjunto de procedimentos que levam a separao das organelas celulares chama-se fracionamento celular. Para se estudar os componentes presentes em uma determinada organela, necessrio que se rompa a sua membrana por choque osmtico, ultrasom ou passando a suspenso de organelas atravs de pequenos orifcios. No caso do ncleo, pode ser usado um detergente, como tween ou Triton, que, ao solubilizar os lipdios da membrana permite a sada do contedo no clear, constitudo de nuclolo, acido nuclico e protenas. O tratamento com ultrassom alm de ser utilizado para romper o envelope nu clear, empregado para romper mitocndrias.
Cromatografia liquida e eletroforese O grande nmero de macromolculas, a variedade de suas atividades biolgicas e as diferenas qumicas, entre elas organismos diversos, tornaram as tcnicas bioqumicas de extrao, purificao e caracterizao, prticas comuns nas pesquisas em Biologia celular. Entre os processos de purificao, a cromatografia lquida um mtodo de separao muito utilizado. Diferentes mtodos cromatogrficos podem ser empregados, levando em considerao as caractersticas da molcula de interesse. O resultado final pode ser a obteno de um componente com um alto grau de pureza.
Cromatografia em gel filtrao. Nesta cromatografia, as molculas maiores no conseguem entrar nas esferas do polmero, e por isso so as primeiras a serem eludas da coluna. J as de tamanho menor penetram nas esferas e, em consequncia, sua migrao
mais lenta. As molculas pequenas atravessam vrias esferas, retardando ainda mais sua migrao. Dependendo do tamanho das diferentes molculas, elas podem ser eluidas da coluna em momentos diferentes, possibilitando que sejam separadas e at purificadas.
Cromatografia de troca inica A separao nessa cromatografia depende, principalmente, da carga eltrica das molculas. As matrizes utilizadas nessa cromatografia so variadas (celulose, cepeharose etc) com grupos carregados positivamente, como o dietilaminoetil, ou negativamente como carboximetil. As matrizes modificadas com grupos DEAE so trocadoras de nions, enquanto aquelas modificadas com CM so trocadoras de ctions. Nos dois casos, as molculas sero eludas com um gradiente crescente de cloreto de sdio.
Cromatografia por afinidade Nessa cromatografia, a separao das molculas (geralmente protenas) baseada em sua afinidade por um ligante.
Eletroforese Uma forma de se analisar as protenas obtidas e um processo de extrao ou presentes em fraes de uma cromatografia pela eletroforese. Essa tcnica consiste em separar molculas colocadas em gel submetido a uma diferena de potencial eltrico.
Tcnicas de DNA Recombinante Esto cada vez mais disponveis mtodos que permitem avaliar com preciso a estrutura e expresso gnicas. A versatilidade e, em alguns casos, a simplicidade dessas metodologias, vem permitindo uma importante ampliao do conhecimento bsico da estrutura e funo gnicas, alm de possibilitar o estudo de anormalidades genticas, podendo envolver alteraes em oncogenes e genes supressores de tumor, diagnstico de agentes infecciosos, identificao de indivduos, melhoramento animal e vegetal em Biologia do desenvolvimento, evoluo de organismos e produo de protenas
recombinantes. Finalmente, o desenvolvimento da tecnologia para gerao de organismos transgnicos uma aplicao direta da tecnologia do DNA recombinante. O DNA consiste de um polmero dupla fita formado por resduos de nucleotdeos trifosfatados unidos por ligaes covalentes do tipo fosfodister. A adio de um ou mais grupos fosfatos na posio 5 ou 3 da desoxirribose origina um nucleotdeo mono, de ou trifosfatado, dependendo do nmero de fosfatos. A determinao da quantidade de DNA em diferentes organismos mostra, em geral, uma proporcionalidade entre esta, o tamanho do genoma e o estgio evolutivo. O ganho de DNA entre bactrias e mamferos, de trs ordens de magnitude, muito alm do aumento do espectro de protenas presentes, Isso sugere o acmulo de DNA com funes distintas daquela de codificar protenas (genes estruturais). A expresso dos genes se d pela sntese de um RNA, cido ribonucleico , que difere do DNA apenas por conter uracila, em vez de timina, e ribose, em vez de desoxirribose. Entre os trs tipos de RNA, sintetizados a partir do DNA, o mensageiro o que codifica protena. A dupla fita de DNA mantida por pontes de hidrognio entre as bases nitrogenadas adenina/timina e citosina/guanina denominadas interaes Watson Crick. Essas mesmas interaes permitem que a sntese de uma molcula de RNA, a partir da molcula de DNA, seja uma cpia fiel da mesma, pois para a sntese de RNA, o DNA aberto por uma complexo enzimtico especfico e uma das fitas serve como molde para a maquinaria que sintetiza o RNA.
Hibridao in situ (HIS) Como o nome indica, esta metodologia permite a identificao de genes ou de seus transcritos simultaneamente localizao celular, sendo tambm efetuada em preparaes cromossmicas. Uma vez que pode ser realizada em tecidos fixados, embebidos em parafina, a hibridao in situ principalmente empregada em estudos retrospectivos. Comparado s hibridaes em filtro ou PCR.
Reao em cadeia da polimerase (PCR) A reao de PCR utiliza certas caractersticas do processo de replicao do DNA. A DNA polimerase, enzima responsvel pela replicao, utiliza uma molcula de DNA fita simples como molde para sintetizar uma nova fita complementar. Essa enzima tambm tem requerimento de um pequeno
segmento de DNA dupla fita para que, a partir deste ponto, ela possa iniciar a sntese. Durante o processo de sntese, faz-se necessria a presena de desoxirribonucleotdeos trifosfatados (dNTP), como precursores, e sal de magnsio em concentrao especfica com o co-fator para a DNA polimerase. A primeira etapa da replicao do DNA in vitro a desnaturao do molde de DNA, ou seja, a separao da dupla fita, que pode ser obtida por aquecimento. Em seguida, preciso que ocorra o anelamento dos iniciadores ( primers) em pontos especficos da cadeia de fita simples do DNA molde, formando um pequeno segmento dupla fita que fornece para a DNA polimerase uma extremidade 3OH livre da ribose para a adio de um novo nucleotdeo. Desse modo, em uma reao de PCR, pode-se direcionar a DNA polimerase para que ela sintetize uma regio especfica do DNA molde de interesse. Durante esse processo, as duas fitas do DNA servem como molde para a sntese, catalisadas pela DNA polimerase. Para que isso ocorra, faz-se necessria a presena de oligonucleotdeos iniciadores para cada fita do DNA molde. Os iniciadores so escolhidos para flanquearem na regio do DNA que vai ser amplificado, de maneira que as novas fitas de DNA sintetizadas, iniciando-se sempre a partir de um iniciador, sejam estendidas at a posio do outro iniciador na fita oposta. Desse modo, novos stios para a ligao dos iniciadores so gerados em cada nova molcula de DNA sintetizada. Se essa mistura de reao for novamente aquecida para separar as fitas moldes, tanto as fitas originais quanto as recm-sntetizadas, esto agora disponveis para o anelamento com os iniciadores e a sntese de novas fitas complementares. Esse comportamento cclico pode ser repetido n vezes, sempre seguindo esse padro de desnaturao, pareamento e extenso.
PCR in situ A tcnica de PCR in situ tem o mesmo objetivo de uma hibridao in situ, porm utilizada quando o DNA alvo muito raro, ou seja, poucas cpias esto presentes no tecido ou na clula, tornando sua deteco por hibridao in situ muito difcil. Nesse caso, aplica-se uma combinao do mtodo de PCR e de hibridao in situ.
Sequenciamento de DNA O mtodo de Maxam e Gilbert inicia-se com a marcao do DNA a ser sequenciado no final de uma das fitas com fsforo (P) . A enzima polinucleotdeo cinase usualmente utilizada para adicionar P na extremidade hidroxil 5 da fita do DNA. O DNA marcado ento quebrado, preferencialmente em um dos quatro nucleotdeos, pelo emprego de reagentes qumicos que especificamente modificam e removem as bases a partir de seus
acares. As condies de quebra so controladas de tal maneira que, em mdia, se obtenha uma quebra por cadeia. Os fragmentos de cada mistura so separadas por eletroforese em gel de poliacrilamida desnaturante; o gel seco , ento, exposto a filme de raio X e o auto-radiograma, analisado. Essa anlise baseada no fato de que os fragmentos de menor tamanho correspondem ao nucleotdeo mais prximo da marcao radioativa e, consequentemente, seria a extremidade 5 da fita de DNA que est sendo sequenciada.
BIOMEMBRANAS
As biomembranas so estruturas laminares, compostas principalmente de lipdios e protenas, que definem os limites entre as clulas e o ambiente extracelular. Elas formam barreiras de permeabilidade seletiva, que regulam a composio molecular e inica do meio intracelular. Tambm tornam possvel a compartimentalizao das atividades metablicas, por meio da segregao de enzimas e outros componentes nas diferentes organelas membranosas presentes na clula. As biomembranas possuem muitas enzimas e sistemas de transportes importantes. Alguns tipos especializados de membranas geram gradientes inicos que podem ser utilizados para sintetizar ATP ou para produzir e transmitir sinais eltricos. Alm disso, na superfcie externa das clulas, esto localizados muitos stios receptores ou de reconhecimento, que podem interagir com outras molculas ou mesmo com outras clulas. Apesar dessas diferentes funes, as membranas tm em comum a sua estrutura geral: uma bicamada composta de lipdios e protenas, mantidos juntos principalmente por interaes hidrofbicas.
Estrutura das membranas biolgicas No sculo passado, Klliker observou que clulas animais e vegetais, quando colocadas em solues inicas concentradas, permitiam a passagem de gua, mas no de molculas solveis. Isso sugeria a existncia de uma barreira semipermevel envolvendo as clulas. Em 1925, Gorter e Grendel extraram fosfolipdios de eritrcitos humanos e os colocaram em um recipientes com gua, Foi observado que esses lipdios formavam uma camada na interface entre a gua e o ar.
Em 1935, com a descoberta de que as membranas tambm eram compostas por protenas, foi proposto, por Danielli e Davson, um modelo no qual protenas globulares encontravam-se na periferia das membranas, interagindo com as cabeas polares dos fosfolipdios. O modelo de Danielli-Davson-Robertson, apesar de apresentar os seus mritos, era sujeito a crticas, como a limitao da permeabilidade que as camadas compactas de protenas representavam. Por volta da dcada de 1970, observou-se que muitas protenas de membrana apresentavam domnios estruturais compostos por a-hlices hidrofbicas. Isso levou sugesto de que algumas protenas de membrana apresentavam-se inseridas na bicamada lipdica. Alm disso, foi mostrado que antgenos de superfcie eram capazes de se deslocar na superfcie celular. Com esses conhecimentos em mos, em 1972, Singer e Nicolsom propuseram o modelo do mosaico fluido para as membranas biolgicas. Esse modelo pressupunha que as protenas estariam embebidas na bicamada lipdica, de forma que as pores hidrofbicas das protenas encontrar-se-iam no interior tambm hidrofbico da bicamada, ambos protegidos do contato com a gua. Isso indicava que essas protenas apresentariam, portanto, trs domnios distintos: 2 hidroflicos, nas faces externas da membrana, e um hidrofbico, no interior dela. O modelo propunha tambm que as protenas, dada a fluidez dos lipdios, estivessem em movimento constante pela bicamada lipdica.
Composio qumica As biomembranas so compostas basicamente por lipdios e protenas, sendo que alguns desses componentes esto covalentemente ligados a carboidratos.
Lipdios So substncias orgnicas insolveis em gua e solveis em solventes orgnicos. Os lipdios so molculas pequenas, sendo que h cerca de 50 molculas de lipdios para cada molcula de protena nas biomembranas. Dentre os vrios tipos existentes, os mais abundantes nas membranas biolgicas so os fosfolipdios, que pertencem classe dos fosfoglicerideos. Em menor quantidade, so encontrados os esfingolipdios e o colesterol. Os fosfolipdios apresentam uma cabea polar ou hidroflica, e duas caudas apolares, ou hidrofbicas. Dessa forma, pelo fato de apresentarem regies hidroflicas e hidrofbicas, os lipdios de membrana so molculas anfipticas. A cabea polar composta pelo glicerol (um trilcool), um fosfato e um lcool, como a colina, a etanolamina, o inocitol ou a serina. As caudas apolares so compostas pelas cadeias carbnicas dos cidos graxos.
Protenas Embora a estrutura bsica de uma biomembrana seja dada pela bicamada lipdica, a maioria dessas funes especficas realizada por protenas. Entre essas funes, pode-se citar o transporte de ons e molculas polares, interao com hormnios, traduo de sinais por meio de membranas e at estabilizao estrutural. A razo entre protenas e lipdios nas membranas varia de acordo com atividade funcional da mesma. As protenas de membrana podem se associar a lipdios de diversas formas. Algumas protenas interagem muito fortemente com as pores hidrofbicas dos lipdios de membrana. Essas protenas s podem ser extradas com o uso de agentes que quebrem essas interaes, solubilizando as membranas, como os detergentes. Essas protenas so conhecidas como protenas intrnsecas. As protenas intrnsecas esto inseridas na bicamada lipdica e apresentam domnios citoplasmtico, transmembrana e nocitroplasmatico. Os domnios que passam pelo interior das membranas e que fazem parte de um ambiente hidrofbico possuem, em sua maioria, resduos de aminocidos hidrofbicos.
Os carboidratos Os carboidratos so de grande importncia para a fisiologia das biomembranas. Eles ocupam espao relevante da superfcie das membranas. No caso da membrana plasmtica, os carboidratos presentes na superfcie celular compem um tipo especial de camada, com cerca de 10 a 20 nm, conhecida como cobertura celular ou glicoclix. Uma vez que os carboidratos carregados negativamente, em especial o cido silico, apresentam-se em quantidades significativas, o glicoclix , em grande parte, responsvel pela carga eltrica negativa encontrada na superfcie da clula. A funo mais importante dos carboidratos nas membranas o reconhecimento molecular, o que permite tambm a comunicao intercelular.
Aspectos funcionais Fluidez das membranas Dada a ausncia de ligaes qumicas entre os diferentes componentes das biomembranas, eles possuem relativa liberdade com respeito aos adjacentes e so livres para se mover no plano da bicamada. A fluidez a capacidade de movimentao dos diferentes componentes na bicamada lipdica.
A movimentao dos lipdios de extrema importncia. Alm de diminuir a rigidez das membranas biolgicas, ela permite a difuso dos seus diferentes constituintes. Na verdade, a maior parte dos fenmenos associados fisiologia das membranas, entre eles a atividade enzimtica e o transporte de solutos, profundamente afetada pela fluidez. A fluidez uma caracterstica dada pela composio lipdica. Dentre os fatores que podem interferir na fluidez esto (1) a presena ou no de insaturaes nas cadeias dos cidos graxos, (2) a temperatura ambiental, (3) a presena de molculas interpostas na bicamada lipdica, e (4) a prpria dieta alimentar. . Presena de insaturaes: as cadeias carbnicas dos cidos graxos podem ser saturadas ou insaturadas. As insaturaes fazem com que os cidos graxos ocupem um maior espao no plano da membrana, possibilitando assim um maior espao no plano da membrana. Possibilitando assim uma maior movimentao dos lipdios e consequentemente das protenas. . Temperatura: a temperatura interfere na fluidez das membranas porque os cidos graxos que os compem apresentam um determinado ponto de fuso e, consequentemente, uma transio de fase. O ponto de fuso dos cidos graxos a temperatura em que a molcula passa do estado gel para um estado lquido-cristalino. Em uma biomembrana no estado gel, os fosfolipdios esto completamente estendidos e alinhados, agrupando-se de forma bastante fechada e perpendicular ao plano da bicamada. A movimentao dos lipdios, nesse caso, fica bastante restrita, o que torna a membrana bastante rgida, compacta e impermevel. Por outro lado, o estado lquido cristalino caracterizado por uma intensa movimentao dos cidos graxos, o que representa uma maior fluidez e maior permeabilidade. . Presena de molculas interpostas: a presena de molculas entre os fosfolipdios, como o colesterol, capaz de interferir na fluidez, pois altera o grau de compactao normal dos cidos graxos e dificulta a movimentao destes no plano da bicamada. O colesterol quebra a estrutura altamente ordenada das cadeias dos cidos graxos. Assim, em uma dada temperatura, por impedir a aproximao e a associao lateral, o colesterol mantm as cadeias de hidrocarbonetos dos fosfolipdios em um estado fluido intermedirio entre o gel e o lquido-cristalino. . Dieta alimentar: outro fator que pode interferir na composio lipdica das membranas celulares e, como consequncia, na sua fluidez, a dieta. Os lipdios obtidos via cadeia alimentar, entre eles cidos graxos saturados, insaturados, poliinsaturados e o prprio colesterol, so incorporados s membranas. Desta forma, a dieta observada em uma dada populao, seja animal ou humana, capaz de interferir de forma bastante acentuada na fisiologia das membranas biolgicas.
Permeabilidade Se analisarmos a permeabilidade de membranas lipdicas sintticas, veremos que elas bloqueiam a passagem da maioria das molculas polares, de molculas apolares grandes (de alta massa molecular) ou molculas carregadas eletricamente. Essa barreira de crucial importncia, pois permite clula manter diferentes concentraes de solutos no citoplasma em relao ao fluido extracelular. O transporte mediado por protenas nas membranas pode ser feito de trs formas diferentes: . Uniporte: quando uma nica unidirecionalmente atravs da membrana; molcula transportada
. simporte: quando duas molculas so transportadas simultaneamente em uma mesma direo; . Antiporte: quando duas molculas so transportadas, simultaneamente, em direes opostas. O transporte atravs das biomembranas ocorre por dois mecanismos bsicos. Por difuso (transporte passivo) ou por transporte ativo. A difuso caracterizada por no existir gasto energtico durante o transporte. Ela pode se dar por difuso simples ou por difuso facilitada. O transporte ativo caracterizado basicamente envolver gasto energtico. Vejamos agora as principais formas de transporte atravs das biomembranas. Difuso simples: ocorre a favor de um gradiente de concentrao de soluto, at que seja atingido equilbrio entre os dois comportamentos. A velocidade da difuso depende da solubilidade dos solutos em relao aos lipdios e do tamanho das molculas. Quanto a maior a solubilidade das molculas nas membranas, maior ser a permeabilidade e mais rpido o transporte dessas molculas. Por exemplo, as solubilidades do oxignio e do nitrognio so extremamente altas. Desse modo, esses compostos se deslocam pelas membranas como se elas no representassem qualquer barreira. Molculas com a mesma solubilidade tero como fator limitante ao transporte o seu tamanho. As que forem menores permeiam mais facilmente pelas membranas. . Difuso por canais proteicos: os canais proteicos constituem vias aquosas para a passagem de solutos. Esses canais so altamente seletivos passagem de solutos de ons ou molculas. Essa seletividade consequncia das prprias caractersticas dos canais, como o seu dimetro e a disposio de cargas eltricas ao longo deste. Os canais de slido, por exemplo, apresentam dimetro entre 0,3 e 0,5 nm e so riscos em cargas negativas. Essas cargas atraem os ons Na+ em maior quantidade que outros compostos, pois a relao
entre a fora de atrao e eo dimetro inico maior para o sdio que para outros ons. J os canais de potssio no apresentam cargas eltricas, de modo que no exercem atrao molecular. Uma vez que o on Na+ menor que o K+, sua densidade de cargas e seu campo eltrico so mais fortes. Assim, a forma hidratada do Na+ maior em dimetro que o K+. Como consequncia, os ons potssio hidratados so menores e arrastam uma menor quantidade de gua. Desta forma, eles passam por seus canais, enquanto os ons sdio hidratados so retidos em decorrncia de suas dimenses. Os canais proteicos nem sempre esto abertos. Existe um sistema de fechamento, semelhante a comportas, que os obstruem. Esse sistema de fechamento representa um mecanismo fisiolgico que permite o controle da permeabilidade das biomembranas. Esses canais podem ser abertos por (1) potenciais eltricos gerados nas membranas ou (2) por interaes com outros tipos moleculares, como hormnios ou neurotransmissores. . Difuso facilitada: um grande nmero de componentes, como acares e aminocidos, atravessam as biomembranas com uma taxa maior do que a esperada em relao ao seu tamanho, carga eltrica ou concentrao. Esses compostos so translocados para o interior da clula por meio da difuso facilitada. Esta ltima uma forma passiva de transporte mediada por carreadores que apresentam especificidade para os solutos presentes no meio extracelular; entretanto, por se tratar de um transporte mediado por protenas de membrana, ela apresenta um limite de saturao. Acima desse ponto, o aumento na taxa de transporte no mais observado, embora a concentrao do soluto possa aumentar. Apesar da difuso facilitada proporcional, um deslocamento mais rpido que as outras formas passivas de transporte, ela incapaz de carrear molculas pelas membranas contra um gradiente de concentrao. A distribuio de Na+ e K+, por exemplo, depende de transporte ativo, que consome de 25% da energia da maioria das clulas e at 60% da energia de clulas excitveis, como neurnios. . transporte ativo: esse transporte feito s custas de gasto energtico. Ele mediado por protenas carregadoras, da mesma forma que na difuso facilitada. No entanto, no transportado ativo o carreador consome energia qumica (ATP) para promover o transporte de molculas contra gradientes de concentrao ou eletroqumicos. O exemplo mais comum de transporte ativo a bomba de sdio e potssio, mediada pelo carregador Na+/K+- ATPase. Esse macanismo transporta ons Na+ do interior da clula para fora e ons K+ do ambiente extracelular para o citoplasma. Embora o Na+/K+- ATPase seja o exemplo mais conhecido de transporte ativo, ele no o nico que ocorre na clula. A bomba de Ca um carreador encontrado na membrana plasmtica e em endomembranas, como as mitocndrias e do reticulo sarcoplasmtico (o retculo endoplasmtico das clulas musculares). Esse carreador, como o prprio nome sugere, promove o transporte de Ca+. Outro carregador que
trabalha mediante gasto energtico a bomba de H+, presente nas membranas lisossomais. Assim como no caso da difuso facilitada, o transporte ativo tambm apresenta um ponto de saturao, que atingido quando todos os carregadores de membrana esto trabalhando em atividade mxima. O transporte ativo de extrema importncia para a fisiologia da clula, embora seja bastante dispendioso em termos energticos. Uma boa parte da energia celular gasta em processos dessa natureza.
Especializaes de membrana
Microvilosidades As microvilosidades so especializaes digitiformes da membrana plasmtica responsveis pela ampliao da superfcie celulares visados trocas entre a clula e o meio. Essas estruturas esto especialmente presentes nas clulas absortivas, como as clulas epiteliais ou dos tbulos proximais dos rins. Estruturalmente, as microvilosidades assemelham-sese a um cilindro recoberto pela membrana plasmtica. Um feixe de filamentos de actina constitui o cerne da microvilisidade, dando-lhe o formato caracterstico. Os filamentos de actina estendem-se em direo ao citoplasma e conectam se a uma rede de filamentos de actina que constituem a trama terminal. Recobrindo externamente a vilosidade, existe um glicoclix desenvolvido, principalmente no intestino. O tamanho e a densidade das microvilosidades variam conforme o tipo celular.
Estereoclios semelhana das microvilosidades, os estereoclios tambm tem importncia no aumento da superfcie de trocas entre a clula e o meio que a circunda. Os estereoclios so mais longos e mais finos que as microvilosidades e no apresentam o feixe de actina caracterstico destas ltimas. No epiddimo, os estereoclios (pontas de seta) esto envolvidos com a modificao da composio do lquido em que ficam imersos os espermatozoides (seta) durante a sua capacitao.
Bainha de mielina
A bainha de mielina corresponde a expanses da membrana plasmtica das clulas de Schwann, que se enrolam ao redor de alguns tipos de axnios (Ax). A funo principal da bainha de mielina garantir um isolamento eltrico do axnio, que contribui para aumentar a velocidade de propagao do impulso nervoso. Entre uma clula de Schwann e outra h uma descontinuidade da bainha de mielina (conhecida como ndulo de Ranvier). O empacotamento das membranas bem estreito e a extenso bastante longa, de forma que a passagem de nutrientes ocorre atravs de junes comunicantes que apaream nos nveis das incisuras de Schmidt Lantermann.
JUNES CELULARES
As clulas dentro de um tecido, ou rgo, esto interconectadas com outras clulas e com a matriz extracelular por meio de estrutura especializadas da membrana plasmtica, denominadas junes intercelulares e junes clula matriz respectivamente. Estruturalmente, as junes intercelulares so classificadas em quatro diferentes componentes: juno de ocluso ( zonula occludens ou tght junction), juno aderente (ou zonula adherens), desmossomo (ou maculae adherens) e juno comunicante (nexus ou gap juction). Quanto s junes clula matriz, estas podem ser divididas em dois tipos: a juno de adeso focal e o hemidesmossomo. Todas essas junes celulares no so necessariamente encontradas em todos os tipos celulares do organismo; as clulas do sangue, por exemplo, so desprovidas dessas estruturas. Em outros casos, como no dos epitlios de revestimento, as junes aparecem bem desenvolvidas. Tem sido demonstrado que essas especializaes da membrana se constituem em elementos altamente dinmicos e regulveis da membrana celular, sendo modulados por substncias e tratamentos diversos. Vrias protenas integrantes das junes j foram identificadas, isoladas e bioquimicamente caracterizadas, tendo sido evidenciado o envolvimento dessas estruturas em vrios processos fisiopatolgicos. A disfuno de algumas junes pode contribuir na etiologia do processo de infeco bacteriana intestinal, cncer e metstase, doenas dermatolgicas de origem auto-imune, neuropatias etc.
Junes intercelulares
Junes intercelulares so definidas como especializaes da membrana plasmtica que interconectam clulas vizinhas dentro de um tecido. As junes foram originalmente denominadas barra terminal, pois apareciam como uma srie de pontos mais corados das membranas laterais de clulas adjacentes, imediatamente abaixo da superfcie apical do epitlio intestinal, observado por microscopia de luz. Acreditava-se que, neste local ocorria depsito de um cimento intercelular que servi para selar o espao intercelular. Com o advento da microscopia eletrnica, entretanto, a barra terminal (termo, agora, em desuso) foi identificada como um complexo de estruturas morfologicamente distintas, localizadas na membrana lateral da clula. No caso tecido epitelial, a juno de ocluso (localizada na poro mais especial), pela juno aderante e pelo desmossomo, foram uma trade na membrana lateral das clulas, conhecida como complexo Unitivo. Funes especficas foram atribudas a cada um dos componentes das junes intercelulares, baseando-se na sua ultra-estrutura visualizada por microscopia eletrnica. A juno de ocluso, a juno ardente e os desmossomos representam dispositivos de adeso intercelular. No epitlio de revestimento, a juno de ocluso age como uma barreira de difuso, limitando a permeao de H2O, ons e pequenas molculas entre as clulas (ou via paracelular). A juno comunicante desempenha uma funo importante no processo de comunicao intercelular. A estrutura bioqumica do complexo Unitivo tem sido intensamente estudada, nos ltimos anos, tendo sido identificadas vrias protenas integrantes a essas estruturas.
ENVOLTRIO NUCLEAR
A existncia de um sistema que delimita o ncleo celular foi sugerida j com as primeiras observaes do prprio ncleo celular. Posteriormente, a presena de um sistema membranoso foi reforada por experimentos de permeabilidade, de micromanipulao e com o emprego da microscopia de polarizao. Em ingls, o termo nuclear envelope tem sido empregado para designar esta complexa estrutura que delimita o ncleo celular. Em nosso idioma, parece que envoltrio nuclear termo mais adequado que envelope nuclear, carioteca, invlucro nuclear ou membrana nuclear, por melhor expressar a complexidade apresentada por esta organela e sua participao na estrutura e fisiologia nucleares. A compartimentalizao do material gentico uma caracterstica fundamental das clulas eucariticas e esta parece ser a funo primordial do
envoltrio nuclear. Porm, outras funes tm sido descobertas. Assim, podese atribuir ao envoltrio nuclear papeis (1) na separao dos processos de transcrio (a sntese de RNA a partir de um molde de DNA), que acontece no ncleo, e de traduo (a sntese de protenas com base de um molde de RNA mensageiro, com a participao dos ribossomos e outros componentes), que acontece no citoplasma; (2) na organizao espacial do material gentico no interior do ncleo, sendo que cromossomos e genes ocupam posies definidas no ncleo interfsico e (3) na determinao da forma e proteo mecnica do contedo nuclear, deixando-o menos sujeito aos movimentos celulares e/ou do citoplasma, ocasionados pelo citoesqueleto. O envoltrio nuclear tambm representa uma barreira seletiva, criando diferenas na distribuio de protenas e ons entre o ncleo e o citoplasma. Esta funo depende de trocas ncleo-citoplasmticas obrigatrias vida celular, existindo mecanismos de importao-exportao de componentes produzidos em um dos compartimentos e destinados ao outro. Outra caracterstica do envoltrio nuclear da maioria das clulas eucariticas a sua desintegrao e reestruturao durante o ciclo celular, dois eventos finamente regulados pelos mecanismos de controle da diviso celular. A complexidade do envoltrio nuclear j era notada pela identificao ultraestrutural do conjunto das duas membranas, do prprio espao delimitado por elas (denominado espao perinuclear), dos pontos de interrupo e fuso das duas membranas em estruturas denominadas complexo de poro, e da presena de uma lmina densa nuclear interna de espessura varivel, a lmina nuclear.
O transporte atravs dos complexos de poro Aps a identificao dos complexos de poro, com o auxlio da microscopia eletrnica, passou-se a acreditar que partculas de pequeno tamanho passam livremente atravs do envoltrio nuclear. O limite desse tamanho seria ditado no pelo dimetro do poro propriamente dito, mas pelos componentes aparentemente fibrilares que se estendem em direo ao seu centro. Alguns experimentos, utilizando ons ou solutos de baixa massa molecular, demonstraram que partculas com at 4,2 kDa difundem-se, passivamente, atravs do envoltrio nuclear, o que seria compatvel com a existncia de canais aquosos com cerca de 9 a 10 nm de dimetro.Componentes de 12 a 50 kDa tambm se difundem atravs do envoltrio nuclear, porm com velocidades inversamente proporcionais aos seus tamanhos.
Entretanto, j na dcada de 1960, experimentos com microeletrodos foram capazes de determinar a existncia de uma diferena de potencial estabelecida pelo envoltrio nuclear entre o citoplasma e o ncleo. Isso indicava que o envoltrio nuclear no s constitua uma barreira passagem de ons, mas tambm mantinha uma diferena em suas concentraes, apesar da existncia dos complexos de poro confirmada pela microscopia eletrnica. Alm disso, mais recentemente, grande ateno foi dispensada identificao dos mecanismos de controle da concentrao nuclear de clcio, face importncia desse on na regulao de diversos processos nucleares, destacando a ativao de endonucleases, que tem papel importante em processos como a morte celular programada (apoptose), por exemplo. Em vrias situaes, foram demonstradas variaes na concentrao nuclear de clcio durante alteraes fisiolgicas da clula. Por outro lado, em outros tipos celulares, como em alguns ocitos, parece no existir qualquer diferena entre as concentraes nucleares e citoplasmticas desses ons. Ento, pode-se concluir que essas variaes resultem de variaes funcionais especficas do tipo celular e do momento fisiolgico, alm de reforar a ideia da existncia de mecanismos de controle das concentraes inicas associados ao envoltrio nuclear, no necessariamente exclusivos dos complexos de poro. Acredita-se, por exemplo, que o clcio acumulado no espao perinuclear tenha importncia na regulao da concentrao intranuclear de clcio, sendo necessrio, para isto, que haja transportadores de clcio em nvel da membrana nuclear interna. Embora j tenha sido observado o brotamento de vesculas das membranas nucleares, o transporte de macromolculas basicamente depende das trocas que ocorrem no complexo do poro. Protenas e RNA so transportados custa de gasto de energia atravs dessas estruturas. A translocao de componentes na direo nuclear ocorre em um processo de duas etapas. A primeira corresponde ligao com o complexo de poro e a segunda, translocao propriamente dita atravs do canal. Como mencionado, macromolculas, maiores que o dimetro do poro, so tambm translocadas, sofrendo modificaes conformacionais necessrias que ocorrem com gasto de energia. A existncia desse sistema de duas etapas confirmada pela demonstrao de que a depleo de energia impede a translocao do material com destino nuclear, mas no a sua ligao com envoltrio nuclear. O transporte de protenas citoplasmticas para o ncleo depende da existncia de uma sequncia de localizao nuclear (NLS, nuclear localization sequence), que corresponde a um conjunto de aminocidos, geralmente de carter bsico, que so reconhecidos por receptores citoplasmticos, conjuntamente denominados importinas, e ento direcionados ao complexo de poro, o que garante a sua internalizao. Alguns dos mecanismos reguladores
da translocao de protenas residem na exposio ou bloqueio dessas sequncias, o que pode ocorrer por alteraes na confirmao da protena, promovidas por modificaes ps-traducionais ou pela associao com fatores especficos (que pode ser um hormnio esteroidal, por exemplo). Alm disso, tem sifo demonstrado que modificaes covalentes por fosforilao tambm so importantes na regulao do transporte. Se a fosforilao ocorre em um dos aminocidos da sequncia de localizao nuclear, o transporte reduzido e a protena acumula-se no citoplasma. Se a fosforilao ocorre em stios adjacentes, mas a uma certa distncia da NLS, o transporte acelerado. Em outras situaes, a sequncia pode ser apenas estrutural e no residir necessariamente na sequncia primria da protena. Isso significa que a sequncia de localizao nuclear somente formada devido a um arranjo especfico encontrado em certa conformao da protena. Alm disso, algumas protenas podem tambm ser transportadas para o interior do ncleo sem possuir a NLS, atingindo o ncleo por interaes com outras protenas competentes para a translocao. O complexo protena e importina, aps ser translocado para o ncleo, desfeito pela associao com uma protena denominada RN-GTP. Essa molcula associa-se importina, desfazendo o complexo. No citoplasma, o GTP hidrolisado a GDP e o complexo se desfaz, disponibilizando a importina para um novo ciclo de transporte para o interior do ncleo.
As vias de transporte de molculas entre o ncleo e o citoplasma Diferentes molculas atravessam os complexos de poro. A existncia de molculas que se concentram no citoplasma ou no ncleo demonstra a existncia de mecanismos de transporte na direo do compartimento alvo e/ou mecanismos de reteno, que impedem o transporte enquanto a molcula no est madura ou mesmo o seu retorno ao compartimento de origem. Pelo processo de difuso, molculas que possuem massa molecular at 50 kDa passam livremente pelo complexo de poro, estabelecendo um equilbrio entre os dois compartimentos, a no ser que os mecanismos de reteno, mencionados acima, concentrem-nas num dos lados do envoltrio nuclear. Quando as molculas apresentam massa molecular acima de 50 kDa, a sua passagem pelo complexo de poro depende de outros fatores alm de sua maior concentrao em um dos compartimentos. No processo de difuso facilitada, as molculas a serem transportadas interagem com componentes do complexo de poro, que auxiliam a sua passagem para o outro compartimento. Aparentemente, no h restrio de tamanho para a passagem dessas molculas, cujo transporte depende da ao do complexo de poro, mas a passagem se d no sentido de equilibrar as suas concentraes nos dois
compartimentos. Por outro lado, para aqueles componentes que so concentrados no ncleo, h o processo de importao mediada por sinal. Esses componentes no passam por difuso simples atravs do complexo do poro e dependem de sinais especficos (sinais de localizao nuclear NLS), reconhecidos por receptores citoplasmticos, que medeiam a sua interao com o complexo de poro e facilitam a sua translocao para o ncleo. Da mesma forma, para aqueles componentes que se originam no ncleo e precisam ser transportados para o citoplasma, existe o mecanismo de exportao mediada por sinal (dependentes de sinal de exportao nuclear NES). Neste caso, so transportados, principalmente, os complexos RNAprotenas que se destinam ao citoplasma. Esses componentes s entram no sistema de exportao nuclear, quando os mecanismos de reteno so removidos, aps o apropriado processamento do RNA. importante ressaltar que os receptores (e carreadores associados a eles) so transportados de volta para o compartimento de origem, sendo reutilizados na mesma via de transporte.
Mutaes nos genes que codificam protenas da lmina nuclear so causas de distrofias So vrias as protenas que fazem parte da lmina nuclear. As laminas A/C e outra protena da lmina nuclear, a emerina, tm sido implicadas em um tipo de distrofia muscular, a Distrofia Muscular de Emery-Dreyfuss. No caso das mutaes dos genes das laminas, a distrofia tem carter autossmico, enquanto a associada emerina tem carter ligado ao cromossomo X. Esse tipo de distrofia muscular atinge adolescentes e se caracteriza por encurtamento dos msculos da perna e do antebrao. Na maioria dos casos, os problemas cardacos manifestam-se entre os 20 e 40 anos. Nos casos mais graves h severa disritmia ventricular (levando morte sbita). O aumento da creatinina plamtica revela, na maioria dos casos, leses nas clulas musculares. Essas clulas apresentam tamanho varivel e as do tipo I so atrficas. A distrofia muscular decorrente de mutaes nos genes de protenas associadas lmina nuclear no bem entendida. Acredita-se que as protenas da lmina nuclear (1) interajam com fatores de transcrio especficos que so necessrios para a manuteno da integridade da clula muscular, (2) faam parte das estruturas de reforo do ncleo que protegem a clula muscular do estresse mecnico e (3) sejam responsveis pela ancoragem de heterocromatina superfcie interna do ncleo, afetando a expresso gnica. Embora todas essas possibilidades ainda no estejam comprovadas, a falha de qualquer uma delas pode ser a responsvel pelos danos das clulas musculares. Entretanto, uma caracterstica comum das clulas afetadas a desestruturao do envoltrio nuclear e o extravasamento do contedo nuclear para o citoplasma. Uma questo relevante a razo pela
qual as clulas musculares so alvo das modificaes, uma vez que todas as clulas carregam a mesma alterao gentica. Uma provvel explicao est no fato de que as clulas musculares tm pouqussimas laminas do tipo B1, que so bastante abundantes nos outros tipos celulares e, aparentemente, contribuem para diminuir o efeito de mutaes deletrias nos genes das laminas A/C. Esse tipo de distrofia pode ser reproduzido experimentalmente por meio da produo de camundongos knock out para o gene das lminas A/C. Por outro lado, alguns tipos de mutaes nos genes das lminas A/C esto associados lipodistrofia familiar do tipo Dunnigan. No surpreendentemente, os knock out para as laminas A/C tambm se caracterizam por uma perda progressiva dos adipcitos. Outra manifestao da ausncia de laminas A/C funcionais na neurodegenerao perifrica que ocorre em pacientes com a sndrome de Charcot-Marie-Tooth . Mutaes nos genes das laminas B tambm levam a alteraes nucleares. A anomalia de Pelger-Huet foi recentemente relacionadaa mutaes nestes genes. Sua principal caracterstica a presena de granulcitos com ncleos arredondados, ao invs de apresentarem a tpica lobulao observada nestas clulas. Os pacientes podem apresentar tambm encurtamento de alguns dos ossos longos.
Especializaes do envoltrio nuclear
Formas irregulares, sulcos da superfcie e retculo nuclear Algumas clulas, como aquelas das glndulas salivares de alguns insetos, apresentam ncleo com formato bastante irregular. Esse formato adotado pela ao do citoesqueleto de actina e sua forma aparentemente mantida pela lmina nuclear. Em clulas de mamferos, tambm so encontradas algumas variaes, como sulcos na superfcie ou canais que se estendem para dentro do ncleo, mantendo-se revestidos pelas membranas do envoltrio e, s vezes, contendo complexos de poro. Em todos esses casos, acredita-se que se tratem de especificaes que favorecem as trocas ncleo-citoplasmticas, promovendo uma ampliao da superfcie nuclear. Em algumas situaes, fica clara a associao desses canais com domnios nucleares. Anlises fisiolgicas mais detalhadas demonstraram a continuidade desses canais com o envoltrio nuclear e com o RE. Alm disto, existe um mecanismo de liberao de clcio para o interior do ncleo, regulado por molculas sinalizadoras e que permitem a disponibilizao desse on, que controla a funo de diversas
molculas para domnios nucleares especficos, e no para o ncleo como um todo. Este sistema recebeu o nome de reticulo nuclear.
A funo de nucleao de microtbulos em plantas A diviso celular em vegetais denominada acntrica, dada a ausncia de centrossomos. J h algum tempo, entretanto, foi demonstrado que as molculas que desencadeiam a polimerizao dos microtbulos esto associadas superfcie nuclear. Desta forma, ncleos de clulas vegetais isoladas so capazes de nuclear a polimerizao de microtbulos. Foi tambm demonstrado que, pelo menos algumas das molculas, existentes na superfcie nuclear das clulas vegetais, apresentam reao cruzada com anticorpos produzidos contra molculas associadas ao centrossomo das clulas animais. Alm disto, foi tambm demonstrada a participao de tubulina e, como elemento auxiliar, associada superfcie do ncleo.
A desestruturao do envoltrio nuclear no inicio da diviso celular At pouco tempo, acreditava-se que o envoltrio nuclear era desestruturado de maneira uniforme, dada a desestruturao da lmina nuclear. Alguns estudos demonstraram, entretanto, que o envoltrio nuclear rompe-se inicialmente num ponto localizado da superfcie do ncleo. Foi tambm demonstrada uma ntima associao do centrossomo com depresses da superfcie nuclear. Os estudos demonstraram ento que a regio de rompimento do envoltrio nuclear diametralmente oposta regio de associao do centrossomo com o ncleo e que, aparentemente, nesta associao, ocorre uma distenso do envoltrio nuclear na regio oposta, o que, em associao com a desestruturao da lmina nuclear, levaria ao rompimento do envoltrio. Sabe-se tambm que h uma participao bastante ativa dos microtbulos do fuso em formao, assim como de molculas de dinena que se ancoram na superfcie nuclear e movimentam-se sobre os microtbulos, auxiliando na trao das membranas nucleares na direo do sulco em que se encontra o centrossomo.
Cromatina e cromossomos
Cromatina o complexo de DNA, protenas histnicas e no-histnicas, presente no ncleo de clulas em interfase. Molculas de RNA podem fazer parte temporariamente desse complexo. Durante a fase de diviso celular, a
cromatina sofre alteraes em sua morfologia, composio e funo, apresentando se sob a forma de unidades individualizadas conhecidas por cromossomos. Nos ncleos interfsicos, a cromatina pode se apresentar diferentemente compactada, granulosa ou filamentosa e com distribuio textural variada, quando se consideram clulas de mesmo tecido ou, at mesmo, tipos celulares em diferentes momentos fisiolgicos, sendo assim identificados diferentes fentipos nucleares. Com frequncia, a cromatina se acha ligada matriz nuclear, uma estrutura filamentosa proteica com diversos papis, entre os quais os de compactao e organizao de cromatina, o de regulao da expresso gnica e o da replicao de DNA. Em nvel ultra-estrutural, a cromatina mostra se constituda por uma estrutura filamentosa com cerca de 10 a 30 nm de espessura, que sofre nveis adicionais de empacotamento. Como esse filamento se encontra organizado no interior dos ncleos ou mesmo constituindo os cromossomos, somente pode ser compreendido com a associao de tcnicas bioqumicas, de Biologia molecular e de microscopia eletrnica mais moderna, tendo sido muito importante, em todos os casos, o emprego de enzimas especiais, como as de endonucleases.
Composio qumica DNA Dentre os componentes da cromatina, o DNA o branco de informaes genticas da clula e seu vetor pelas vrias geraes celulares. O DNA encontra-se na cromatina na forma de macromolculas. Constitui-se de duas cadeias helicoidais de polinucleotdeos complementares (purina pirimidina), que associam por ligaes de hidrognio, girando para a direita ao redor de um eixo central imaginrio, lembrando uma escada helicoidal. Enquanto a disposio das bases nitrogenadas pareadas corresponderia aos degraus dessa escada, perpendiculares ao eixo central, as cadeias de acar-fosfato corresponderiam ao seu corrimo. Forma-se, assim, um duplex de 20A de dimetro, conforme estabelecido no modelo clssico de Watson e Crick. O modelo mencionado refere-se conformao B do DNA. Outras conformaes so conhecidas, como a A e a C, variando-se nestas os graus de hidratao do meio, o passo da hlice da macromolcula, o nmero de pares de bases nitrogenadas por volta e o ngulo que as bases nitrogenadas fazem com o eixo imaginrio, ao redor do qual a macromolcula se estende. Existe uma conformao chamada Z, facilitada por uma sequencia especial de
bases (-CGCGCG- ou ACACA-CAC-) e com sentido da hlice voltado para a esquerda, fazendo com que a disposio do esqueleto acar-fosfato do DNA adquira uma geometria em zig-zag, da sua denominao. O DNA pode ainda se apresentar in vitro com outros tipos de estrutura, como o caso do DNA H (hlice tripla). importante mencionar que, embora vrias configuraes j tenham sido descritas para o DNA, a configurao B que se admite quando so apresentados modelos de estrutura cromatnica. O contedo de DNA, no interior de um ncleo diploide, razoavelmente constante para as diferentes clulas de uma mesma espcie. Nas clulas germinativas (vulo e espermatozoide), o contedo de DNA costuma ser haploide, ou seja, a metade do contedo encontrado nas clulas diploides correspondentes. Uma clula pode existir com menos de 500 genes (em torno de 580.070 pb de DNA), como na pequena bactria Mycoplasma genitalium, que possui o menor genoma conhecido. Embora clulas de organismos superiores tenham contedos de DNA vrias ordens de grandeza maiores do que os de organismos unicelulares, no h uma razo direta entre espcies filogeneticamente superiores e contedo de DNA. Assim, insetos de mesma ordem podem diferir em at 10 vezes em valores absolutos de contedo de DNA por clula diploide, anfbios podem apresentar muito mais DNA do que o gato domstico.
Histonas As histonas so protenas bsicas nucleares de alto ponto isoeltrico, encontradas nos eucariotos. So importantes componentes da estrutura da cromatina , participando no somente como repressoras, mas tambm como ativadoras da transcrio do DNA. Como o DNA, as histonas so sintetizadas na fase S do ciclo celular. A rezo de massa DNA/histona igual a 1,0. So cinco as classes principais de histonas (H1, H2A, H2B, H3 e H4) que so classificadas com base em seu teor em lisina e arginina. A histona H1 muito rica em lisina, enquanto as histonas H2A e H2B so moderadamente ricas em lisina, e as histonas H3 e H4 so ricas em arginina. Em termos de evoluo, as histonas so muito conservadas, o que significa que variam pouco em sequncia de aminocidos nas diferentes espcies consideradas. No entanto, esto entre as protenas mais modificadas, o que acontece por fosforilaes, acetilaes, metilaes e ubiquitinizao. As modificaes diminuem as cargas positivas das histonas, alterando as interaes DNA-histonas. Muitas das modificaes so reversveis.
Dentre as histonas, a mais varivel em estrutura primria e tamanho H1, sendo, pois, a menos conservada em termos evolutivos. Em contrapartida, as histonas H3 e H4 so as mais conservadas evolutivamente. As histonas H4 de bovinos e ervilhas, espcies que divergiram 1,2 bilhes de anos atrs, diferem pr apenas dois resduos. A molcula da histona H1 apresenta trs regies bem definidas: a amino-terminal, com 40 resduos de aminocidos; a central, globular e hidrofbica, com cerca de 80 resduos; e a carboxiterminal, com 108 resduos e a mais rica em aminocidos bsicos, especialmente lisina. Os outros tipos de histona apresentam maior quantidade de aminocidos hidrofbicos nas regiescarboxiterminal e globular. Em eritrcitos nucleares de peixes, aves, rpteis e anfbios, para da histona H1 aparece substituda por outra protena da mesma famlia, H5, que exerce papel especial na estabilidade fsico-qumica e condensao cromatnica dessas clulas, restringindo sua atividade gnica. Nos espermatozoides, as histonas somticas so substitudas por (ou acrescentadas de) outros tipos de protenas nucleares bsicas. Essas podem ser do tipo caracterizado por uma riqueza especial em arginina, a ponto de o polipeptdeo poder ser considerado quase uma poliarginina, como acontece no salmo. Nesse caso, a protena conhecida como protamina ou clupena, tem uma massa molecular em torno de 4.000 dltons e uma afinidade muito alta por DNA em dupla fita, tornando o complexo DNA-protena muito estvel e de difcil dissociao. Em outros tipos de espermatozoides, como nos mamferos, pode ocorrer uma protena rica em arginina, classificada como semelhante protamina ou queratinosa, por conter tambm o aminocido cistena. H ainda espermatozoides em que a protena nuclear bsica uma variante de H1, muito rica em lisina e com variada presena de regies de a-hlice. Esse tipo ocorre em ourios-do-mar, em abelhas e em alguns anfbios. Embora se tenha tentado estabelecer uma correlao entre tipos de protena bsica nuclear presentes em espermatozoides e filognese, at o momento so se logrou xito.
Protenas no- histnicas Participam ainda da composio da cromatina as protenas no-histnicas (NHP). Admite-se que essas protenas desempenhem variados papis na cromatina, desde o estrutural at o enzimtico, participando da regulao da atividade gnica. Fazem parte desse grupo as enzimas que atuam nos
processos de transcrio, replicao e reparo do DNA e nos processos de condensao e descondensao cromatnica. A massa molecular das protenas nucleares NHP varia de 10 a centenas de kDa, a razo de resduos cidos/bsicos delas varia 1,2 a 1,6 e seu ponto isoeltrico, de 3,7 a 9,0. O nmero de protenas nucleares no-histnicas, analisadas por eletroforese em gel de poliacrilamida com SDS, varia conforme o tecido num mesmo indivduo, tendo j sido encontrados mais de 1.000 tipos dessas fraes proteicas. Pouco ainda conhecido com respeito distribuio das protenas NHP na cromatina. De certo, sabe-se que, aps a extrao de DNA. Histonas e RNA e mesmo de muitas protenas NHP, permanece um esqueleto ou arcabouo, especialmente visto em cromossomos, cuja arquitetura lembra a das prprias unidades cromossmicas. O DNA de regies precisas da cromatina se ligaria a essas protenas.
RNA O RNA integrado de modo transitrio estrutura da cromatina, constituindo em torno de 3% da sua composio e correspondendo s cadeias recmtranscritas em vrias fases do seu processo de elongao.
Estrutura da Cromatina Embora desde 1956, houvessem sido apresentadas propostas de como ocorreria a associao do DNA com protenas, organizando as fibras de cromatina, somente a partir de 1974, com os trabalhos de Olins e Olins e de Kornberg et al., pode ser estabelecido um modelo de estrutura cromatnica mais aceito. Olins e Olins, nos EUA, submeteram ncleos de diferentes tecidos a um choque osmtico com KCI 0,1 M. Esse tratamento rompeu os ncleos, liberando fibras de cromatina que, ao microscpio eletrnico, exibiam uma distribuio linear de pequenas unidades aproximadamente esfricas (nubodies), com um dimetro de 7 nm, unidas entre si por um filamento com 1,5 nm de espessura. Na poca, essa estrutura foi chamada de colar de contas. Ao mesmo tempo, na Inglaterra, um grupo de bioqumicos, liderados por Kornberg, comprovou que a fibra cromatnica era constituda por unidades repetitivas compostas por duas molculas de cada uma das histonas H2A, H2B, H3 e H4 e de cerca de 200 pares de bases (PB) de DNA.
Em 1975, Oudet et al, denominaram a unidade estrutural repetitiva da cromatina de nucleossomo. Este mostrou ser constitudo por um nucleoide (core nucleossmico) ligado unidade seguinte por um filamento espaador (espaador internucleossmico). O nucleoide estaria composto por 146 pb de DNA, que descreveria 1 e de volta de uma hlice ao redor de um octmero de histonas (2 molculas de H2A, 2 de H2B, 2 de H3 e 2 de H4). O filamento espaador composto por uma sequencia varivel em nmero de PB de DNA. Chegou-se a concluso de que o nucleoide teria a forma aproximada de um cilindro achatado, com 11 nm de largura e 5,5 nm de altura, e que podia ser clivado por uma digesto suave com nuclease microccica. Graas ao achado de que o DNA do nucleoide de fcil acesso digesto enzimtica por endonucleases, como o DNase I, de origem pancretica, demonstrou-se que o DNA ocuparia uma posio perifrica, em relao ao octmero de histonas. Esta ideia foi confirmada por difrao de nutrons. Segundo a idealizao de Oudet, faria parte do nucleossomo a histona H1. No entanto, na literatura que se seguiu ao trabalho de Oudet, encontra-se muitas vezes sob a definio de nucleossomo a unidade cromatnica, sem se incluir a molcula de H1. Assim, para se evitar dvidas na conceitualizao, prefervel que se faa referncia unidade repetitiva da cromatina, ao invs de nucleossomo, e se defina, em termos bioqumicos, o que se est considerando. tambm frequente na literatura moderna, o uso do termo cromatossomo para indicar a unidade cromatnica, na qual se inclui uma cpia de H1. H locais bem definidos de interao do DNA com as histonas do core, o que se demonstra pelo acesso do DNA ao enzimtica por DNase I (do pncreas) e DNase II (do bao). Quando o DNA no complexado a histonas clivado por nucleases, so produzidos fragmentos com tamanhos variados, que se manifestam em gel de agarose ou poliacrilamida com um padro caracterstico de bandas. No entanto, quando a cromatina submetida ao mesmo tratamento, os fragmentos de DNA so clivados segundo um padro regular em mltiplos de 10,4 nucleotdeos, isto porque o DNA est ligado s histonas em stios especficos, com repetitividade. O DNA espaador, ou seja, aquela poro da unidade repetitiva da cromatina no contida no nucleoide, interage com as histonas H2A e H2B na entrada e na sada do DNA do nucleoide. Assim, as histonas H2A e H2B estariam, de alguma forma, estabilizando a ligao DNA-H3H4, mas tambm se ligando ao DNA espaador. Alm disso, h evidncias de que as histonas H2A e H2B possam se ligar tambm histona H1. As histonas H2A-H2B apareceriam formando heterodmeros, enquanto as histonas H3-H4 constituiriam um tetrmero. Esse tetrmero um complexo estvel de dois heterodmeros de H3-H4.
A posio exata da histona H1 na unidade repetitiva da cromatina tem sido muito discutida. Embora tenha sido admitido por muito tempo que H1 se posicionaria lateralmente ao nucleoide, ligando-se ao DNA espaador e participando na compactao do filamento cromatnico, modernamente tambm se admite que a histona H1 possa se ligar ao DNA e as histonas do core, no por fora, mas por dentro da estrutura nucleossmica. Apesar do DNA ser menos compctado em cromatina deficiente em histona H1, o papel dessa histona parece estar mais relacionado com a reunio de complexos nucleoproteicos reguladores especficos, que participam tanto da represso quanto da ativao da transcrio.
Nveis hierrquicos de organizao cromatnica Vrios nveis de compactao do filamento cromatnico ocorrem no ncleo interfsico. O nucleofilamento a fibra cromatnica de 10 nm de espessura, com a sequncia linear das unidades repetitivas da cromatina. O nucleofilamento sofre uma organizao helicoidal com 5 a 6 unidades repetitivas da cromatina por volta de hlice, constituindo uma fibra de 20-30 nm denominada solenoide. Admite-se que o solenoide seja estabilizado no apenas graas interao entre molculas de H1, mas tambm por interao entre as faces superior e inferior das unidades repetitivas, especialmente pelas caudas das histonas dos nucleoides. Tem sido tambm relatado que, no arranjo das unidades cromatnicas em sequencia, devam fazer parte certas regies curtas descontnuas, geralmente com 100 a 400 pb de DNA de comprimento, mais acessveis clivagem por DNase I e reagentes. Essas regies corresponderiam a stios gnicos regularidades, acessveis a fatores de transcrio, podendo estar associadas matriz nuclear. Se tais regies de DNA se associarem matriz nuclear na cromatina de ncleos interfsicos, sero chamadas de MAR ( matrix attchment regions); se a associao ocorrer em cromossomos de clulas em diviso, as regies sero denominadas de SAR (scaffold attchment regions). Segundo Worcel, as molculas de H1 estariam com a sua polaridade alternada ao longo do nucleofilamento, ou seja, a regio N-terminal de uma molcula de H1 estaria prxima regio C-terminal de outra, situada na volta adjacente da super-hlice do nucleofilamento. As interaes H1 H1 poderiam, assim, estabilizar a fibra de 20 30 nm (solenoide). Admita-se que, a cada dez unidades repetitivas da cromatina, a molcula de H1 aparea substituda pela protena UH2A (ubiquitina ligada H2A), o que conferiria maior flexibilidade fibra solenoidal. O papel de outras
protenas no histnicas na estrutura da cromatina ainda pouco compreendido. O conhecimento sobre os nveis hierrquicos superiores da estrutura cromatnica ainda limitado. Existem evidncias de que a cromatina esteja dividida em domnios funcionais, cujos limites estariam em associao com a matriz nuclear. Foram alcanados avanos pelo uso da microscopia fluorescncia tridimensional, associada a sondas para regies cromossmicas ou cromossomos especficos, possibilitando que fosse admitida a organizao de territrios cromossmicos e de movimentos e mesmo rotao de unidades cromossmicas e regies cromossmicas no ncleo interfsico. Certas situaes patolgicas parecerem mesmo estar associadas com mudanas de composio dos cromossomos no ncleo, como o caso da posio do cromossomo X em focos epilpticos. Nas clulas em diviso, o empacotamento dos complexos DNA protena maior, a ponto do ndice DNA/cromossomos ser ao redor de 5.000/1, enquanto, no solenoide, da ordem de 50:1 e, na unidade repetitiva da cromatina, 7:1. Durante a fase S do ciclo celular, ocorre a replicao do DNA, segundo o modelo semiconservativo. Tambm nessa fase ocorrer agregao de novas histonas recm sintetizadas. A natureza dessa agregao tem sido muito discutida. Uma das hipteses mais aceitas a de que as molculas de histonas novas e paternas se distribuam ao acaso nos filamentos que se replicam. Quanto estrutura cromatnica durante a transcrio do DNA, h indicaes de que as unidades repetitivas da cromatina sofram algum tipo de alterao estrutural, com reposicionamento do octmero de histonas em relao ao DNA ou mesmo completa liberao do DNA a ser lido pela RNA polimerase.
Os cromossomos metafsicos Os cromossomos autossmicos geralmente ocorrem aos pares (2 lotes cromossmicos, 2n) nas clulas somticas, tanto de animais como de vegetais. O nmero de cromossomos de uma espcie constante e se mantm como tal durante os ciclos repetidos de diviso celular. Durante a meiose, os cromossomos sofrem reduo em nmero. No final do processo, sero obtidas clulas com um lote cromossmico (clulas haploides ou n). Dado que ocorre o fenmeno de recombinao dos genes, as
unidades cromossmicas, geradas no final do processo, no sero idnticas s de origem materna ou paterna. A determinao do nmero de cromossomos de uma espcie geralmente efetuada na metfase, perodo no qual ocorre a condenao mxima das unidades cromossmicas, facilitando a contagem. Nas metfases meiticas (I ou II), a condensao dos cromossomos ainda maior do que na mitose. Os cromossomos apresentam tamanho relativamente constante nas diferentes clulas de uma mesma espcie, em mesma fase do processo de diviso. No mesmo lote cromossmico, no entanto, os cromossomos podem ser apresentar com extrema variabilidade em tamanho. O comprimento dos cromossomos pode variar de 0,2 a 50 um e o seu dimetro de 0,2 a 2 um. Na mesma espcie humana, os cromossomos atingem de 4 a 6 um de comprimento. O conjunto das caractersticas morfolgicas que permite a caracterizao dos lotes cromossmicos de um indivduo denominado de caritipo. E acostuma se ordenar os cromossomos, fotografados ou esquematizados, aos pares de homlogos, numa srie decrescente de tamanho. Esse arranjo do caritipo, que revela o nmero, a forma e os tipos de cromossomo, denominado cariograma. Pode-se tambm representar o caritipo esquematicamente, construindo um idiograma. Essa forma de representao dos cromossomos utiliza os valores mdios do tamanho relativo e da posio de centrmero obtidos por medidas tomadas para vrios caritipos de diferentes indivduos de uma espcie. A cada das metades cromossmicas observadas durante a diviso celular e que iro constituir um novo cromossomo, se d a denominao de cromtide. As cromtides sero irms se forem de um mesmo cromossomo, ou homlogas, se situadas em diferentes cromossomos do mesmo par (paterno e materno). Denomina-se cromonema a unidade cromossmica filamentosa, ou seja, o prprio cromossomo. Centrmeros, constries secundrias e telmeros so as principais diferenciaes morfolgicas naturais dos cromossomos. O centrmero ou constrio primria do cromossomo a regio em que situa o cinetcoro, estrutura organizadora da polimerizao das fibras cromossmicas do fuso mittico. A posio do centrmero em um determinado cromossomo constante, permitindo que este possa ser classificado como metacntrico, se localizado na poro mediana do cromossomo, submetacntrico, se deslocado para um dos braos cromossmicos, e acrocntrico ou telocntrico, se
posicionado em uma das extremidades do cromossomo. H espcies, como o roedor Calomys SP, em que a maioria dos cromossomos acrocntrica. Graas obteno de sondas especficas e ao mtodo de FISH (florescence in situ hybridization), possvel identificar a regio centromrica de um cromossomo em especial, inclusive em ncleos interfsicos. Outras constries presentes nos cromossomos so chamadas de secundrias e podem a conter a regio organizadora do nuclolo, geralmente se associando a ele. As extremidades cromossmicas so denominadas telmeros. Se acidentalmente ocorrer perda de telmeros em diferentes cromossomos, estes podero se fundir. Os telmeros consistem de sequncias de DNA ricas em G, repetidas centenas de vezes e altamente conservadas em termos evolutivos. As extremidades dos cromossomos no se replicam por ao normal da DNA polimerase, mas sim por mecanismo especial que envolve atividade de uma transcriptase reversa (telomerase), que carrega consigo seu prprio molde de RNA, complementar s sequncias repetitivas de DNA do telmero. O uso deste RNA permite telomerase entender a extremidade 3 do DNA cromossmico de uma unidade de repetio, para alm de seu comrpimento original. A falha na manuteno do nmero das repeties telomricas, por deficincia na ao das telomarases, parece estar relacionada ao envelhecimento celular. Nas clulas tumorais, a atividade telomersica parece se manter elevada e o tamanho do telmero, por conseguinte, no se altera. H mtodos especiais de tratamento e de colorao que permitem identificar nos cromossomos segmentos ou bandados importantes na sua identificao e caracterizao. As bandas aparecem por diferenas na distribuio de componentes cromatnicos ou mesmo por diferena na composio qumica da cromatina ao longo do cromossomo. As bandas Q aparecem fluorescentes com quinacrina. As bandas G aparecem como segmentos mais corados com Giemsa, aps tratamento com tripsina ou tampo fosfato. As bandas C aparecem aps tratamento com solues cidas ou alcalinas, seguindo de tratamento com soluo salina 2SSC e colorao com Giemsa. De fato, a denominao de banda C se baseia nas primeiras observaes de que o mtodo evidenciava regies centromricas. As alteraes nos padres da distribuio de bandas geralmente esto associadas a anomalias de cartergentico. Os padres de banda tambm permitem estudos de filognese animal e vegetal. Com o projeto Genoma Humano,dados de mapeamento do DNA do genoma humano puderam ser cruzados aos de mapas de bandado cromossmico obtido ao nvel citogentico. Essa informao se encontra disponibilizada em site do national Center for Biotechnology Information (USA) (http://www.ncbi.nih.gov). Como os genes por cromossomos so muito
nmerosos, para serem representados em sua totalidade, as imagens fornecidas buscam informar algum aspecto particular no qual o pesquisador tenha interesse. Por exemplo, cromossomo humano 17 foram localizados 1.679 genes distribudos segundo as suas diferentes regies; destes genes, apenas uma parte tem funo conhecida. Os detalhes de localizao dos genes no respectivo mapa cromossmico mostram que, dentre muitos que poderiam ser citados, o gene que codifica para a queratina 15 se situa na regio 17q21.2, o que codifica o antgeno 5 associado ao espermatozoide, na regio 17q11.1 e o que codifica a serina/ treonina cinase 12, na regio 17p13.1.
Os cromossomos gigantes: plumosos e politnicos Tanto os cromossomos plumosos quanto os politnicos apresentam-se com tamanhos muito acima dos comumente observados. Os plumosos chegam a atingir 800um de comprimento. Os politnicos atingem 150 a 250um de comprimento, porm, se as clulas em que ocorrem estiverem infectadas por vrus ou microspordeos, chegam a atingir 1.500um. Os cromossomos plumosos foram descobertos 1892 por Rckert, em ocitos de tubaro. Podem tambm ocorrer em ocitos de outros peixes, bem como de anfbios e rpteis, em algumas clulas vegetais e no espermatcito de Drosophila (cromossomo Y). Tambm, em espermatcito de gafanhoto, a estrutura cromossmica, quando vista ao microscpio de luz, lembra o aspecto plumoso. Os cromossomos politnicos ocorrem em clulas somticas de vrios tecidos de dpteros (Drosophila, Rhynchosciara, Sciara, Chironomus e outros), em insetos colemboldeos, em protozorios ciliados e no ligamento suspensor do feijo. Os cromossomos plumosos ocorrem na prfase meitica ((diplteno), quando so observados dois homlogos mantidos unidos por quiasmata. Distribudas ao longo dos cromossomos, aparecem centenas de regies granulosas, com maior espiralizao de nucleofilamentos, os chamados crommeros. Quando os crommeros e desespiralizam, projetam-se alas finas, distribudas aos pares, simtricas e com caractersticas morfolgicas prprias e constantes. Alas se desespiralizam de ambas cromtides de cada homlogo. Nas alas dos cromossomos plumosos, h uma intensa sntese de RNAm, que ser liberado do cromossomo no interior de micronuclolos e ir comandar a sntese de protenas. Nas alas, portanto, so incorporados precursores de RNA. A atividade de cada cromossomo independente e distinguvel da dos demais, por representar um lcus gnico.
Os cromossomos plumosos esto to espessamente recobertos por RNA e protena que o seu DNA chega a representar apenas 1% de sua massa seca total. O padro de transcrio simultnea de muitas molculas de RNA, nesses cromossomos, lembra as imagens de rvore de Natal vistas no nucleolo, durante a transcrio do precursor maior dos RNA pesados. Alm disso, possvel que uma mesma ala possua diversas unidades de transcrio. s vezes, vista uma ala desespiralizada apenas em um dos lados da estrutura plumosa, indicando diferentes manifestaes de dois genes alelos em um indivduo heterozigoto (herana mendeliana simples). Muito desenvolvidas nos ocitos jovens, as alas desaparecem com o decorrer da ovognese, quando a liberao das fibrilas de RNP de suas matrizes no nucleoplasma e consequentemente retrao do filamento de DNA protena. Nas alas dos cromossomos plumosos cujo DNA transcreve para RNAm de histonas,foi observado que este RNA ocupa apenas um curto segmento do transcrito. No se conhece o significado do restante da cadeia de RNA transcrita em cada fibrila. O cromossomo plumosos Y, da prfase meitica de Drosophila, apresenta ao menos 5 pares de alas essenciais para a diferenciao do espermatozoide vivel. Comprovou se que a ausncia de uma dessas alas induz o aparecimento de espermatozoide com encurtamento de cauda e com imperfeies no axonema e nos derivados mitocondriais. J em relao aos cromossomos politnicos, estes se originam por pareamento ponto por ponto dos cromossomos homlogos de modo que, embora em uma espcie o nmero 2n de cromossomos possa ser igual a 8, ocorram 4 unidades morfolgicas individualizadas. Essas unidades iniciam, ento, uma srie de ciclos de replicao, sem que as cromtides se separem (pareamento secundrio). O nmero de ciclos de replicao varia conforme o tecido e a espcie considerados. Em glndulas salivares de Drosphila virilis e de Rhynchosciara americana ocorrem 10 ciclos, organizado mais de 1.000 cromtides por cromossomo politnico (2 = 1.,024); em Sciara coprophila ocorrem 12 ciclos e em Chironomus. Geralmente, ocorre um grau de politenizao mais elevado nas glndulas salivares. O volume dos nuclolos com cromossomos politnicos aumenta na medida em que aumentam os ciclos de politenizao. Da mesma forma, aumenta o tamanho das clulas e dos rgos em que se encontram. Foram j registrados casos em que a politenizao no permanente, ou seja, pode ser seguida por uma fase em que as cromtides se separam e se dispersam, dando ao ncleo um aspecto interfsico mais usual.
Nos insetos colemboldeos, os homlogos no se pareiam; politenizam-se em separado. Ao longo do comprimento dos cromossomos politnicos, ocorrem em regies com maior espiralizao cromatnica (crommeros) alternadas com regies de menor espiralizao. Durante a politenizao, os crommeros de cromtides dispostas paralelamente ficam justapostos, gerando estrias transversais em registro, as bandas. As bandas se separam entre si pelas interbandas. As bandas ocorrem em nmero, posio e distribuio constantes para os mesmos cromossomos de diferentes tecidos, o que favorece a elaborao de mapas cromossmicos. De pose desses mapas possvel constar- se alteraes estruturais e fisiolgicas ao longo dos cromossomos. Experimentos com radioautografia mostraram incorporao de timidina tritiada ao longo do filamento cromossmico dos cromossomos politnicos nos vrios ciclos de politenizao, demonstrando que o DNA contnuo de uma extremidade a outra do cromossomo politnico. Admite-se que uma banda no represente propriamente um gene. Um gene pode estender-se de banda a interbanda ou at ser compreendido numa interbanda. Algumas bandas contm diversos losi gnicos e uma s banda pode se apresentar como duas, aps um esmagamento cromossmico mais forte. Algumas regies dos cromossomos politnicos sofrem intumescimentos com desepiralizao dos filamentos cromatnicos, seguindo-se regresso do fenmeno durante o desenvolvimento do organismo. Esses intumescimentos so denominados pufes, anis de Balbiani e bulbos, e parecem no ser diagnosticveis nos protozorios e no feijo. Nos anis de Balbiane, estruturas que aparecem nos cromossomos politnicos do gnero Chironomus, no se verificam regresso do intumescimento. Quanto aos bulbos, tratam-se de pequenos pufes de RNA que ocorrem predominantemente em sciardeos. Embora possa ocorrer transcrio ao longo de todo cromossomo politnico, nos pufes que se d a sntese mais intensa de RNA. O padro de pufao vria conforme o tecido e o rgo considerado. As modificaes morfolgicas e fisiolgicas nas regies de pufe, associadas a modificaes de carter celular funcional, permitem a obteno de alguns dados sobre a fisiologia cromossmica e sua regulao. Em poucos casos pde ser estabelecida uma relao direta entre um pufe e uma funo especfica. Um exemplo clssico foi a observao de Beermann, em 1961 de que quatro clulas especficas das glndulas salivares de Chironomus pallidivittatus apresentavam o cromossomo IV com o anel de Balbiani BR4 bem desenvolvidos, ao mesmo tempo em que essas clulas secretavam grnulos de protena. Em Chironomus tentans, no foi constatado anel de Balbiani nem secreo de grnulos proteicos nas clulas
correspondentes s de C.pallidivittatus. hibrido dessas espcies recebia anel BR4 apenas num dos lados do cromossomo politnico IV, bem como metade da secreo proteica verificada em C. pallidivittatus. Assim, a herana desses grnulos proticos foi demonstrada como sendo devida a um fator mendeliano simples. A utilizao de eletroforese, associada a tcnicas genticas convencionais, possibilitou que se correlacionassem os pufes 3C e 68C de cromossomos pilinicos de glndulas salivares de Drosophila melanogaster com a sntese de duas fraes proteicass especficas. tambm notrio que alguns pufes, alm de descreverem RNA, contam com uma replicao local, por amplificao gnica, de um DNA adicional. Esses pufes so denominados pufes de DNA e foram descobertos por BREUER e PAVAN, 1955, em cromossomos de Rhynchosciara americana e, mais tarde, comprovados em outros dpteros i ciardeos dpteros sciardeos. Quando da regresso do pufe de DNA, o DNA adicional no eliminado e, portanto, essa regio do cromossomo se torna aumentada em espessura, o que conhecido por cicatriz. Foi demonstrado, pelo grupo de Lara, que o DNA amplificado nos pufes de DNA de Rhynchosciara americana estaria codificando a transcrio de um RNAm responsvel pela sntese de protenas de secreo salivar do inseto, envolvidas na fabricao do seu caso comunal. Admite-sese que a pufasso pufao seja induzida por ao hormonal, especialmente de ecdisona, hormnio de muda dos insetos. A resposta ao hormonal variaria conforme a poca do desenvolvimento do inseto e do rgo especfico considerado, bem como da concentrao e tempo de ao do hormnio. Foi tambm demonstrado que outras substncias podem induzir alguns pufes especficos; entre elas estariam ons metlicos, antibiticos, vitaminas e galactose. Choques de temperaturas so tambm muito conhecidos como indutores de pufao. Larvas de Drossophilas crescidas a 25C, quando transferidas para meio a 37C, chegam a exibir 9 a 10 em 30 a 40 min. A formao de um pufe envolve o acmulo de protenas no-histnicas na interbanda adjacente ou na prpria banda que origina o pufe. O contedo das protenas no-histnicas acompanha o crescimento dos pufes, bem como sua regresso. H hipteses que buscam interpretar o papel dessas protenas nos pufes, entre as quais semencionam: empacotamento e transporte de RNA recm transcritos, ativao de genes presentes no prprio pufe ou em regies a ele adjacentes, ou ainda transcrio de RNA. Quanto as protenas histnicas em pufes de DNA, so elas adicionadas regio de pufe quando o DNA adicional amplificado.
Heterocromatina e eucromatina O termo heterocromatina, cunhado por Heitz, foi proposto para definir, em termos morfolgicos, a cromatina de regies especficas de certos cromossomos, que permanecia condensada (heteropicntica) durante toda a interfase, com uma afinidade to intensa por corantes tanto quanto aquela de cromossomos metfasicos. Esse comportamento se diferenciaria daquele da maioria da cromatina que, aps a diviso celular, torna-se mais descompactadas e difusa (eucromatina). Alm das caractersticas estruturais, caractersticas fisiolgicas, qumicas e genticas passaram a se somar para definir heterocromatina seja a cromatina condensada, com replicao de DNA mais lenta e certa inatividade gentica. A heterocromatina pode ser classificada em constitutiva e facultativa. A heterocromatina constitutiva aquela de regies centromricas, telomricas e, algumas vezes, intercalares de regies homlogos, geralmente ricas em DNA repetitivo ou satlite e predominantemente no codificadora. Um conjunto de pores heterocromticas constutivas pode associar se entre si, constituindo corpos maiores, mais facilmente evidenciveis, denominados cromocentros. Estes so muito conspcuos em clulas de insetos, em hepatcitos de camundongo e em clulas portadoras de cromossomos politnicos. A caracterstica de no codificao por parte do DNA da heterocromatina constitutiva parece no ser to rgida assim, uma vez que algumas de suas reas tm sido vistas em associao ao ncleo e, sob condies de estresse, como choques de temperatura ou infeces, podem vir apresentar descondensao, como acontecem cromossomos politnicos e em clulas de triatomneos. As sequencias ativas, em termos de transcrio, geralmente se encontram na eucromatina. possvel que a condensao cromatnica dificulte o acesso da RNA polimerase ao DNA, porm outros fatores podem estar envolvidos. A simples observao de condensao cromatnica ao microscpio de luz (nvel de supra organizao cromatnica em nvel nucleossomal. A heterocromatina facultativa no apresentaria diferenas na composio de DNA em comparao com a eucromatina; seria mais um estado fisiolgico reprimido do DNA numa regio condensada da cromatina, ocorrendo em apenas um dos cromossomos homlogos, e podendo reverter ao estado no condensado. Exemplos: a heterocromatina de um dos cromossomos X de mamferos placentrios XX e todos os cromossomos paternos de machos de planococcus sp (pulges de farinha). No caso heterocromatina facultativa de um dos cromossomos X, a heterocromatizao instala se em um dos
cromossomos do par, ao caso, aps alguns ciclos de diviso celular do embrio, e reverte ao estado eucromtico nas clulas germinativas (ocitos). Uma vez que os conceitos clssicos de heterocromatina e eucromatina no so exatamente precisos para distinguir muitas regies cromatnicas com funo diversa, talvez alguma reviso desses conceitos se faa necessria. O termo heterocromatina parece englobar diferentes tipos de estrutura cromatnica, cujo denominador comum um alto grau de organizao, e possivelmente distinguidos por ocorrncia de diferentes tipos de modificaes de histonas e de agregao de protenas no histnicas. Atualmente, vem-se propondo que a heterocromatina possa fornecer um mecanismo de defesa contra elementos mveis de DNA, empacotando tais estruturas e impedindo que as mesmas sofram uma proliferao subsequente.
Fentipos nucleares A observao de ncleos interfsicos ao microscpio de luz permite que se descrevam suas caractersticas geomtricas aliadas distribuio e aos estados de empacotamento (supra-organizao) da cromatina. Tais caractersticas, que definem os fentipos nucleares, so realadas pelo emprego de testes citoqumicos apropriados, de modo que as imagens produzidas ganham melhor poder discriminativo. A moderna anlise dos fentipos nucleares incorporou procedimentos obtidos com o uso de analisadores de imagem, permitindo a incorporao de inmeros parmetros matemticos anlise qualitativa ou semi-quantitativa mais simples, e e em ganhando notoriedade no campo da Biologia celular. Identificando diferentes funcionalidades nucleares e celulares e, no campo da citopatologia, como ferramenta de reconhecimento de leses celulares, auxiliando em muitos casos o citodiagnstico. Clulas epiteliais mamrias humanas em cultura, quando transformadas e tornadas tumorignicas pela ao de carcingenos, como o benzo[a]pireno, um dos elementos presentes no fumo, sofrem alteraes nos seus fentipos nucleares. Salienta o aumento em condensao cromatnica nos ncleos das clulas tumorignicas em relao aos das clulas no-transformadas (controle).
NUCLEOOLO
O nucleolo a estrutura celular mais facilmente visvel, mesmo sem colorao e in vivo, em microscopia de luz comum, o que possvel graas ao seu ndice de refrao mais elevado do que o dos outros elementos do ncleo e do citoplasma. Embora j tivesse sido descrito por Fontana, em 1781, sua denominao, como a conhecemos hoje, foi dada por Valentin, somente em 1839. O nucleolo a organela celular cuja funo produzir ribossomos. Seu tamanho e forma dependem do estado funcional celular, variando conforme a espcie e, dentro de uma espcie, e dentro de uma espcie, de tecido para tecido e mesmo de clula para clula. Muitas vezes o nuclolo visto prximo periferia nuclear, porm essa no uma regra fixa. Quanto mais forte a sobrecarga funcional celular, maior ser o nuclolo. o que ocorre em clulas em processo de secreo (clulas glandulares e neurnios) e em muitas clulas tumorais. Por outro lado, como exemplo de clulas com nucleolos pequenos, temos as clulas endoteliais e as da glia. Podem ser observados um ou mais nucleolos por ncleo, porm a maioria das clulas possui apenas um nuclolo. Hepatcitos, clulas vegetais e clulas animais em cultura so alguns exemplos de clulas em que ocorre mais de um nuclolo. No caso extremo de ocitos de anfbios, podem ser encontrados, em algumas circunstncias, at 3.000 nuclolos por ncleo. Ncleos poliploides, ou seja, com vrios lotes do genoma, geralmente contm mais nuclolos do que ncleos diploides. A falta de uma membrana ao redor do ncleo pode significar que no exista barreira para difuso entre nucleolos e nucleoplasma. O nucleolo se associa a stios cromossmicos especficos (zonas organizadoras do nuclolos, NOR) que carregam os genes codificadores do RNAr mais pesados. Pode ocorrer uma nica NOR por lote cromossmico haploide. No entanto, dois nucleolos podem se fundir ou um zona organizadora do nucleolo pode se encontrar distribuda em mais de um cromossomo do lote haploide. Nos seres humanos, por exemplo, os genes para RNAr se situam nas extremidades de cinco diferentes pares cromossmicos. comum tambm se observar uma regio de heterocromatina em ntima associao com a NOR, Em hepatcitos de roedores, a heterocromatina se distribui ao redor do ncleo, enquanto o inverso ocorre em hempteros sugadores de sangue. Durante o ciclo celular, podem ocorrer alteraes na forma e tamanho dos nucleolos. Costuma-se afirmar que, durante a diviso celular, os nucleolos desaparecem a partir do fim da prfase, reaparecendo no final da telfase. H, no entanto, excees regra.
Ultraestrutura e classificao dos nuclolos Ao microscpio eletrnico, so detectveis nos nucleolos reas ricas em elementos granulares (grnulos com dimetro ao redor de 15 a 20 nm) e reas predominantemente fibrilares (3 a 4nm de espessura), que variam em disposio tridimensional, conforme o tipo celular analisado. Com base nessas observaes, foram propostas vrias classificaes dos nucleolos, sendo a mais simples e objetiva aquela que contempla a grande maioria dos casos estudados e que pressupe trs categorias: 1. nucleolos reticulados, com nucleolonema, estrutura filamentosa trabeculada com aproximadamente 1.000 nm de espessura e contendo em seu corpo, predominantemente , elementos granulares de ribonucleoprotena (RNP), mas tambm elementos fibrilares; 2. nucleolos compactos, no qual os elementos se superpem e se anastomosam numa massa nica compacta. So observados, principalmente, em clulas em proliferao, podendo se tratar de uma primeira etapa de desenvolvimento de um nucleolo que ser do tipo 1, ou mesmo surgir em condies que resultem em inibio ou bloqueio de sntese de RNA nucleolar; 3. nucleolos com camadas concntricas, em que a poro central representada pelo elemento fibroso e a camada cortical, perifrica, contm os elementos granulares. Esse tipo de nucleolo muito comum em ocitos de anfbios. Nessas clulas, as duas camadas, central e cortical, podem ser muito bem discriminadas at com um microscpio de luz (microscopia de fase) e separadas por choque osmtico com solues salinas de NaCI ou KCL de baixa molaridade, ou gua deionizada, a partir de nuclolos previamente isolados com pinas, sob lupa.
Organizao molecular e papel fisiolgico na biognese de ribossomos O nucleolo encontra-se envolvido na biognese de ribossomos, no unicamente com respeito sntese dos RNAr mais pesados, mas tambm quanto ao seu processamento ps-transcricional e sua reunio, seja com RNAr mais leves, seja com protenas. Ocorrem muitas modificaes qumicas no precursor de RNAr antes que esse RNA venha a se tornar parte integrante do ribossomo. Essas modificaes provavelmente auxiliem na dobradura e na agregao dos RNAr finais e alterem a funo dos ribossomos.
O nuclolo durante a mitose
Segundo descries clssicas, o nucleolo desaparece no fim da prfase e reaparece na telfase, junto aos stios NOR. A reformulao ps-mittica dos nuclolos dependeria da interao de duas entidades separadas, as regies NOR e os corpos pr-nucleolares, que aparecem na telfase. Mais recentemente, constatou-se que os componentes nucleolares, RNAe e protenas no se dissolvem no citoplasma, durante a diviso celular. O RNA permanece como um envoltrio ao redor dos cromossomos na placa metafsica, porm, quando da migrao destes para os plos, desloca-se para a regio junto aos microtbulos do fuso entre os dois blocos cromossmicos em movimento, assim permanecendo at a telfase. Parte das protenas nucleolares, na metfase, tambm se posiciona em volta dos cromossomos, porm permanece acompanhando os respectivos blocos cromossmicos, em sua migrao para os plos. Supe-se que esses componentes, ou parte deles, participem da reorganizao dos nuclolos na telfase. Admite-se tambm, ao menos com relao aos componentes que permanecem em ntima associao aos cromossomos, que formem uma estrutura organizada tal que desempenhe um papel protetor do material gentico, numa situao em que a lmina nuclear se encontra dissociada e espalhada pelo citoplasma, alm de se constituir em stio de armazenamento de fatores de maturao de RNAr.
MATRIZ NUCLEAR, CROMOSSMICOS
DOMNIOS
NUCLEARES
TERRITRIOS
O ncleo interfsico foi a primeira estrutura intracelular a ser observada ao microscpio, mas vrios aspectos de sua organizao funcional ainda so muito pouco conhecidos. Recentemente, tem sido demonstrado que o ncleo em intrfase altamente compartimentalizado e extremamente dinmico. Muitos fatores nucleares distintos so localizados em compartimentos especficos no ncleo que, por tcnicas especficas, podem ser estudados ao microscpio de luz e eletrnico. Enquanto um significativo progresso no entendimento das propriedades especficas dos genes e de sua regulao ocorreu nas ltimas dcadas. Como j visto em captulos anteriores, a arquitetura do ncleo interfsico est sustentada basicamente pelos componentes, morfologicamente distintos: envoltrio nuclear, lmina nuclear, cromatina, nuclolo e tambm por uma atriz proteicas conhecida por matriz nuclear.
Atualmente sabe-se que o ncleo interfsico no uma estrutura homognea, e os cromossomos, que so individualizados na diviso celular, no ncleo interfsico, apresentam domnios prprios e microambientes regulados por protenas desta matriz nuclear, constituindo, assim, territrios cromossmicos especficos. A manuteno desse microambiente de fundamentarl importncia para ao e expresso gnicas, pois os genes ocupam posies especficas no ncleo interfsico, ao contrrio do que se pensava h poucas dcadas, e este posicionamento especfico dos genes no ncleo interfsico chamado de domnio nuclear.
Definio Beresney e Coffey definiram a matriz nuclear como sendo uma estrutura altamente residual, obtida dos ncleos por extraes salinas sequenciais, tratamento com detergentes no-inicos ou nucleases. Essa definio reconhecida como a primeira definio original do termo matriz nuclear. A matriz nuclear consiste de uma poro morfolgica e bioquimicamente distinta, por se apresentar como uma estrutura proteica fibro-granular, que alicera o ncleo, distinguindo-se dos outros componentes da cromatina. Ela associa-se ao DNA quando este contm sequencias ricas em A-T, sendo estas conhecidas como Regies de Associao de Matriz (MAR) ou, do ingls, Scaffold associated region (SAR). Esse arcabouo, proteico prende o DNA durante os processos de duplicao e regula a transcrio nos eucariotos, juntamente com as histonas.
Composio qumica A matriz nuclear pode associar-se a at 80% do DNA genmico, estando ele na forma de filamentos cromatnicos ou cromossmicos. Estudos iniciais revelaram que, dos componentes da matriz nuclear em fgado de rato, o material mais abundante de origem proteica, seguido de RNA, fosfolipdios e DNA. Os polipeptdeos da matriz nuclear variam de peso molecular entre 30 e 190 kDa. Dentre as protenas da matriz nuclear, pode-se citar genericamente as matrinas e as metaloprotenas. As matrinas, em linhas gerais, so as principais e maiores protenas da matriz nuclear, distinguindo-se bioquimicamente das laminas A, B e C da lmina densa e das protenas nucleolares, como a B-23, e tambm das hnRNP (ribonucleoprotenas heterogneas), j identificadas no ncleo interfsico.
As matrinas so protenas de peso molecular ponto isoeltrico variveis e foram descritas a partir de estudos com eletroforese bidimensional. A nomenclatura foi adotada de acordo com o peso molecular, ou seja, a matrina 1 a mais pesada (190 kDa), e as numeraes subsequentes so de acordo com o peso molecular decrescente. Muitos subtipos dessas protenas tm sido descritos ano a ano por pesquisadores da rea, que atribuem funes especficas a essas matrinas na fisiologia nuclear. As metaloprotenas so protenas que garantem a integridade estrutural da matriz nuclear sem, entretanto, impedir as relaes entre os componentes da matriz e os da cromatina. A descoberta das metaloprotenas na matriz nuclear permitiu o entendimento de que os ons cpricos associados a essa protenas que promovem a maior estabillizao de algumas interaes molecular durante a ao de genes, seja na ativao ou no silenciamento gnico. Outra classe de protenas que tem tomado grande importncia na matriz nuclear so as glicoprotenas, podendo assumir um papel funcional no transporte e reconhecimento de sinais na matriz nuclear. No se pode deixar de mencionar as enzimas do metabolismo cromatnico, que podem fazer parte da trama arquitetural da matriz nuclear. Como exemplo, citam-se as topoisomerases, que atuam efetivamente no processo de duplicao do DNA e j foram isoladas como sendo constituintes da matriz nuclear. O RNA o segundo componente em abundncia na matriz nuclear. Essas molculas encontram-se sob a forma de hnRNP ou tambm em pequenas ribonucleoprotenas nucleares (snRNP). Esses elementos so oriundos do processamento e clivagens de RNA cromatnicos e nucleolares. Recentemente, essas estruturas tm sido denominadas de nuclear speckles, e tem-se assumido que esses compartimentos subnucleares sejam a expresso morfolgica do dinamismo cromatnico/cromossmico das clulas eucariotas. Nas clulas eucariotas, foram mapeados entre 10 e 30 stios de nuclear speckles difusamente distribudos no nucleoplasma e matriz nuclear. Recente reviso realizada por Angus Lamond e David Spector prope uma redefinio dos compartimentos nucleares, baseada no modelo destes speckles e propem um modelo de organelas nucleares. Isto logicamente baseado na distribuio e quantidade dessas estruturas por ncleo, avaliandose por meio de microscopia de fluorescncia e confocal.
Morfologicamente, esses speckles se organizam em grnulos intercromatnicos, por vezes, estruturados em cachos ou filamentos pericromatnicos. Outros componentes, como DNA e fosfolipdios, por apresentarem uma baixa expresso volumtrica nas extraes bioqumicas, so considerados por muitos autores como contaminantes.
RIBOSSOMOS E SNTESE PROTEICAS
Nas clulas, a informao passa do DNA para o RNA e deste para as protenas. Cada gene consiste de uma sequencia linear de nucleotdeos que determina a sequencia de aminocidos em um polipeptdeo. Dois processos so fundamentais para que isso ocorra: a transcrio e a traduo. Durante a transcrio, a informao contida na sequncia de nucleotdeos do gene codificada em molculas de RNAm. Durante a traduo, a sequencia de cdons (cada trs nucleotdeos consecutivos) do RNAm utilizada para adicionar aminocidos especficos, um a um, para a formao de uma cadeia polipeptdica. A traduo do RNAm ocorre nos ribossomos, uma organela constituda da RNAr e protenas. A traduo requer tambm molculas de RNAt, que se ligam ao aminocido especfico de acordo com o anticdon ( sequencia de trs nucleotdeos) e este se associa ao cdon do RNAm no ribossomo, trazendo assim os amonocidos para serem incorporados cadeia polipeptdica na ordem precisa determinada pelo DNA.
O DNA de cada clula de todos os organismos contm, pelo menos, uma cpia (ou raramente, vrias) dos genes que carregam a informao para produzir cada protena que o organismo necessita. O primeiro passo para a expresso dessa informao gentica a transcrio dos genes em molculas complementares de RNA. Para esse processo, so necessrios diferentes ribonucleotdeos: ATP, GTP, CTP e UTP e as enzimas polimerases, que catalisam a adio de cada um desses nucleotdeos de acordo com a sequncia de bases do DNA, ou seja, onde houver timina (T) no DNA, ser adicionada adenina (A) no RNA, C-G, G-C e A-U. Pelo processo de transcrio, todos os RNA celulares so produzidos a partir do DNA. Os trs principais tipos de RNA transcritos que participam conjuntamente na sntese proteicas so: RNA mensageiro (RNAm), RNA transportador (RNAt) e RNA ribossmico (RNAr).
A sntese de protenas no feita diretamente do DNA. Para que a informao, contida na sequncia de bases do DNA, seja traduzida em uma sequncia de aminocidos da protena (estrutura primria), um tipo especial de RNA utilizado como intermedirio, o RNAm. A sequncia de nucleotdeos dos diferentes RNAm lida pela maquinaria celular da sntese proteicas para produzir milhares de protenas. Esse processo ocorre em organelas celulares denominadas ribossomos e conhecido como traduo.
O ribossomo Esta estrutura celular constituda de molculas de RNAr e protenas. Logo que o RNAr transcrito, associa-se a protenas e, nos eucariotos, fica retido temporariamente em torno da regio cromatnica no qual esto localizados os genes ribossmicos, formando o nucleolo. No nucleolo, os RNAr precursores transcritos sero processados e originaro as duas subunidades ribossomais. O RNA 5S, que faz parte da subunidade maior, transcrito em outra regio cromossmica, migrando para os nucleolos. Assim, as subunidades prontas atravessam o complexo de poro do envoltrio nuclear e se associaro no citoplasma apenas no momento da sntese proteicas, originando o ribossomo funcional. Mitocndria e decloroplasto tambm contm ribossomos que, em alguns organismos, so menores que os de procariotos. Mitocndrias de organismos eucariotos unicelulares (leveduras, Neurospora, Aspergillus nidulans, Euglena gracilis, entre outros) contm ribossomos com coeficiente de sedimentao semelhante aos de Escherichia coli, isto , 70S A 74S (subunidades 50S e 40 ou 30S; RNA 21 24S e 14 15S), enquanto em animais (por exemplo, no homem, no rato, no anfbio Xenopus laevis, no gafanhoto lucusta migratria e no inseto Drosophila melanogaster), os ribossomos so de 55 a 60S (subunidades 40S e 30S; RNAr 16 a 17S e 12 a 13S). Nas mitocndrias das clulas humanas, o ribossomo constitudo das molculas de RNAr 16S e 12S, porm, nas plantas, contm, alm desses dois, tambm um RNAr 5S. Por outro lado, os ribossomos 70S dos cloroplastos apresentam 4 tipos de RNAr, 23S, 5S e 4,5S na subunidade maior 50S e o 16S na subunidade menor 30S. Nos eucariotos, os ribossomos podem ser encontrados livres no citoplasma ou associados membrana do retculo endoplasmtico e membrana externa do envoltrio nuclear. A nica diferena entre esses ribossomos est relacionada ao destino das protenas que esto sitentizado. Os ribossomos apresentam de 20 a 30 nm de dimetro e podem ser observados ao microscpio eletrnico de transmisso como centenas, s vezes milhares, de pequenos pontos. Escherichia coli, por exemplo, contm cerca de 20.000 ribossomos, o que representa 25% do peso seco da clula bacteriana, e
clulas de mamferos em proliferao contm cerca de 10 milhes de ribossomos. A forma dos ribossomos foi deduzida a partir de estudos por mtodos fsicos, como o espelhamento de nutrons, a difrao de raio X e a observao ao microscpio eletrnico com reconstruo tridimensional, entre outros. A localizao das protenas no ribossomo tem sido possvel por imuno eletroscopia e anlise bioqumica. A anlise por criomicroscopia eletrnica mostrou que os ribossomos so constitudos de densos filamentos com aspecto contorcido, de forma que o ribossomo como todo exibe um aspecto rugoso. Nessa imagem do ribossomo no possvel distinguir as molculas de RNA e de protenas, mas possvel visualizar a posio do aminoacil RNAt entre as subunidades maior e menor.
A sntese proteicas A sntese de protenas pode ocorrer em polissomos (vrios ribossomos + um RNAm) livres ou associados ao retculo endoplasmtico. O mecanismo bsico da sntese o mesmo, independente de onde ela ocorra. Vrios componentes so importantes durante a sntese, mas alguns so primordiais, como as molculas de RNA. Alm do RNAr presente nos ribossomos, atuam na sntese proteicas RNA mensageiro (RNAm) e o RNA transportador (RNAt). Como visto anteriormente, o RNAm contm a sequncia de bases que vai determinar a ordem em que os aminocidos sero adicionados durante a sntese de protenas. A leitura do RNAm vai ocorrer na direo 5 3, como ser visto mais adiante. Um outro cido nucleico de fundamental importncia durante a sntese proteica o RNAt, que vau se ligar a um determinado aminocido e tambm vai ser reconhecido por um grupo de 3 nucleotdeos na molcula de RNAm. Enquanto a molcula de RNAm alongada em toda a sua extenso, a molcula de RNAt apresenta em sua estrutura tridimensional, vrias dobras, que resultam dos emparelhamentos entre as bases nitrogenadas(A-U, C-G) que esto sobre o mesmo filamento de polinucleotdeo. Tambm existem domnios, como se fossem duplas hlices, levando molcula a assumir as conformaes mostradas na figura 11.4. Duas regies da molcula de RNAt tm importncia especial no processo de sntese proteica. Uma dessas regies contm uma sequncia de trs nucleotdeos, CCA, em sua extremidade 3 que vai se ligar covalentemente a uma molcula de aminocido, e a outra regio contm uma sequncia de 3 bases (denominada anticdon), que ir se emparelhar com o cdon presente na molcula de RNAm. No entanto, se cada cdon tivesse um anticdon correspondente, haveria 61 RNAt diferentes. Porm, foram encontrados pouco mais de 30 RNAt. Isso acontece porque o
pareamento da primeira base do anticdon do RNAt(dita oscilante), na posio 5, com a terceira base do cdon (na posio 3) do RNAm no segue o padro. Essa primeira base do anticdon pode ser uma guanina (G) modificada, denominada insnia (I), e pode emparelhar se com C, A e U. E ainda, se a primeira base do anticdon for U, pode reconhecer A e G na terceira posio de cdon, e se for G, reconhece C e U. Por exemplo, os cdons 5 - UUU-3 ou UUC no RNAm especificam fenilalanina-RNAt ( um RNAt com um aminocido fenilalanina ligado) e podem se complementar com o anticdon 3 AAI-5. As duas primeiras bases SAP as mesmas em ambos os cdons e determinam o aminocido; a terceira pode se emparelhar com qualquer anticdon do RNAt. A estrutura tridimensional dos diferentes RNAt faz com que essas molculas sejam reconhecidas por enzimas especficas, que vo catalisar a ligao de cada RNAt a um determinado aminocido. Uma caractersticas do RNAt a expressiva quantidade (cerca de 10%) de nucleotdeos covalentemente modificados. Essas modificaes afetam a conformao da molcula de RNAt, implicando no reconhecimento pela enzima que catalisa a ligao do RNAt com o aminocido e tambm no emparelhamento correto com o RNAm. Antes de ser iniciada a sntese, o aminocido passa por um processo de ativao, no qual este se liga a uma molcula de RNAt em uma ligao catalisada pela enzima aminoacil RNAt sintetase . A ligao se d entre o grupo hidroxila presente na extremidade 3 da molcula, no adenilato da sequencia CCA, e o grupo carboxila do aminocido. A ativao do aminocido catalisada pela aminoacil RNAt sintetase, na verdade, se d em duas etapas. Primeiramente, o aminocidose liga ao AMP, formando um aminoacil adenlico: Ainda no stio ativo da enzima, o aminoacil transferido para o RNAt: Aminoacil-AMP + RNAt Aminoacil RNAt + AMP O aminocido ligado ao seu RNAt est em condies de ser adicionado cadeia de polipeptdeos que est sendo sintetizada no polissomo. No caso de a sntese estar sendo iniciada, alm da ativao do aminocido, outros eventos ocorrem como a associao do RNAm subunidade menor do ribossomo e tambm associao das duas subunidades dos ribossomos. Esta etapa de iniciao, que bastante complexa envolve a participao de protenas especificas chamadas de fatores de Iniciao (IF). Como j foi visto, as subunidades ribossomais somente se associam durante o processo da sntese proteicas. No caso de procariotos, a subunidade maior s vai se associar subunidade menor depois que esta ltima se ligar ao RNAt iniciador carregando o aminocido metionina ligado a um radical formil (f-met). O RNAt iniciador sempre carrega f-met, no caso de procariotos, e met, no caso de eucariotos. Em procariotos, a ligao do RNAt iniciador ocorre no stio P da
subunidade menor e s depois desse complexo formado, que o RNAm se associa. Em procariotos, o incio da sntese se d com o N- formil- metioninaRNAt, que vai se instalar no stio P do ribossomo. O cdon de iniciao quase sempre AUG (em algumas bactrias pode ser GUG). Um questionamento que surgiu, quando se iniciarem os estudos sobre sntese proteicas, foi de como o ribossomo iria reconhecer o stio do RNAm para iniciar a sntese, uma vez, que existem vrias sequncias de AUG ao longo da molcula de RNAm. No demorou muito para que fosse identificada uma sequncia no RNA 16S presente na subunidade menor do ribossomo, a qual se pareava com uma sequncia (sequncia Shine Dalgarno) no RNAm, que precedia a sequncia AUG que deveria, ento, servir de ponto inicial para a sntese de protenas. Em eucariotos, primeiramente o RNAt transportando a metionina se associa subunidade menor do ribossomo, com a ajuda de protenas chamadas fatores de inciao, conhecidas pela sigla eIF (do ingls eucaryotic Initiation Factors). Em seguida, esta subunidade se liga extremidade 5 do RNAm, aps reconhecer uma base modificada a 7- mentil guanosina, e dois fatores de iniciao, eIF-4g e eIF-4E, que j estavam associadas extremidade 5(cap) do RNAm. A subunidade menor ento se move ao longo do RNAm com a ajuda de outros fatores de iniciao, at encontrar uma sequncia AUG. Neste ponto, os fatores de iniciao so desligados e a subunidade maior se associa ao complexo Metional-RNAt-subunidade 40SRNAm, formando o de ribossomo completo funcional. Pelo menos 10 fatores de iniciao participam nessa fase inicial da sntese proteica em eucariotos. J em procariotos, atuam apenas trs fatores de iniciao, IF-1,IF-2,IF-3 para uma visualizao da iniciao da sntese em procariotos. Tanto em procariotos como em eucariotos, a incorporao da subunidade maior do ribossomo, formando o ribossomo funcional, ocorre aps a ligao do aminocil-RNAt com a subunidade menor do ribossomo e com o RNAm. O conjunto formado pelo ribossomo completo, f-met-RNAt ou Met-RNAte RNAm, conhecido como complexo de iniciao 70S e 80S e, a partir do momento em que esse complexo formado, a sntese tem prosseguimento, com a adio de novos aminocidos. Segue se a formao da ligao peptdica e a translocao do ribossomo sobre a fita de RNAm, caracterizando a etapa de enlogao. No ribossomo, so reconhecidos trs stios: o stio A, que recebe os complexos de aminoacil RNAt, o stio P. como pde ser visto na etapa da iniciao, o primeiro aminocido ocupa o stio P, deixando o stio A livre para receber o prximo aminoacil RNAt. Este escoltado at o ribossomo por uma protena chamada fator de elongao (EF-Tu em procariotos e EF-1 em eucariotos), que se complexa com o GTP. Aps o aminocil RNAt estar corretamente emparelhado com o cdon do RNAm no
stio A, oGTP sofre hidrlise, o EF-Tu GDP liberado e o primeiro aminocido metionina se liga, por seu grupo carboxil, ao grupo amino do aminoacil RNAt que ocupa o stio A. O EF-Tu GDP agora inativo, pode ser novamente ativo, complexando-se com um fator EFTs, que permite a troca de GDP por GTP, regenerando o EF-Tu-GTP e a sua capacidade de novamente interagir com um outro aminoacil-RNAt at alcanar o ribossomo. A formao da ligao peptdica, seja no caso da f-met-RNAt e no de Leu-RNAt,ou entre qualquer peptidil RNAt e o prximo aminoacil RNAt que estiver ocupando o stio A, catalisada por um complexo enzimtico, peptidil transferase, presente na subunidade maior do ribossomo. Vrios estudos tm mostrado que essa reao mediada no por uma protena, mas pelo maior RNAr presente na subunidade maior do ribossomo. O passo seguinte, ainda dentro do processo de elongao, consiste na migrao ou translocao do ribossomo ao longo do RNAm na direo 5 3, deixando, dessa forma, o stio A livre para a chegada de um novo aminoacilRNAt. A translocao depende da presena de outro fator, EF-G, e consequente hidrlise de GTP, resultando em mudana conformacional do ribossomo e na sua translocao ao longo do RNAm. A etapa de elongao muito semelhante entre procariotos e eucariotos, deferindo apenas em relao aos seus fatores de elongao. O crescimento da cadeia polipeptdica, pela adio de aminoacil-RNAt, ocorre at que o stio A do ribossomo encontre um dos cdons UAA, UAG e UGA, conhecidos como stop codon, para os quais no existe RNAt com anticdon capaz de formar emparelhamento. No entanto, aqueles cdons so reconhecidos por protenas chamadas de fatores de liberao. Esses fatores vo encerrar a sntese proteica, pois ocupam o stio A e alteram a atividade da enzima peptidil-transferase, que catalisa a hidrlise da ligao entre a cadeia polipeptdica e o RNAt que est no stio P. Assim, o polipeptdeo liberado, ocorrendo o mesmo com o RNAt. As subunidades do ribossomo separam-se e o RNAm se dissocia. Nesta etapa, os procariotos e eucariotos diferem quanto aos fatores de liberao. Os procariotos tm os fatores RF-1, que reconhece UAA ou UAG, e RF-2, que reconhece UAA ou UGA, enquanto os eucariotos possuem um nico fator de liberao, e Rf, que reconhece os trs cdons de terminao.
Polissomos Durante a sntese proteica, os ribossomos se deslocam ao longo do RNAm na direo 5 3 e deixam para trs a extremidade 5, ou algum stio em que outro ribossomo possa se instalar. Desse modo, vrios ribossomos funcionais
podem estar associados a uma mesma fita de RNAm, caracterizando o polissomo ou polirribossomo. A quantidade de ribossomos associados depende da extenso do RNAm. Uma molcula de RNAm, com 500 nucleotdeos, por exemplo, pode apresentar at 5 ribossomos sintetizando protenas simultaneamente.
Bloqueadores da sntese proteica At 1940, no havia um tratamento eficaz para vrios tipos de doenas infecciosas. Mas, a partir daquela data, foram descobertas vrias substncias com capacidade de inibir o crescimento de bactrias. Essas substncias so os antibiticos e muitos deles tm uma ao especfica sobre bactrias, bloqueando, na sua maioria, etapas da sntese proteica, como ocorre com a estreptomicina, cloranfenicol, tetraciclina e eritomicina. Outros, como a puromicina, que atuam tanto em procariotos como eucariotos, e a cicloeximida, que atua s em eucariotos, no tm aplicao clnica, mas foram importantes em experimentos que levaram compreenso do mecanismo da sntese proteica. No caso da puromicina, por ter uma estrutura muito semelhante a um aminoacil-RNAt, pode ocupar o stio A do ribossomo durante a sntese proteicas, chegando a se ligar ao peptidil-RNAt que est no stio P, formando peptidil-puromicina, que ento liberado do ribossomo sem que este sofra translocao ao longo do RNAm. Esse resultado demonstrou que a translocao s ocorre depois da ligao do aminoacil-RNAt ao peptidil que est no stio P. Um exemplo clinicamente importante de bloqueio da sntese proteica ocorre na doena denominada difteria, que causada por uma bactria, Corynebacterium diphteria, que libera uma toxina (protena) codificada por um bacterifago. Somente aquelas linhagens de C. diphteria lisognicas para o bacterifago causam essa grave doena. A toxina liga-se ao gangliosdeo GM1 na superfcie de membrana plasmtica das clulas humanas. A toxina clivada em dois fragmentos, denominados A e B. O fragmento B se insere na membrana plasmtica e parece formar um poro por onde o fragmento A passa para as clulas das pessoas infectadas. Dentro da clula, o fragmento A catalisa uma reao de ADP-ribosilao do fator de elongao (eEF-2; uma translocase), inativando-o de forma irreversvel. Isso impede a translocao do ribossomo e, consequentemente, a transferncia do peptidil-RNAt do stio A para o stio P, interrompendo a sntese proteica. Um nico fragmento A capaz de bloquear 500.000 molculas de eEF-2 em cada clula, levando a pessoa morte. A difteria uma doena que causa leses, principalmente, na poro superior do trato respiratrio, levando a um espessamento da mucosa nessa regio e facilitando o crescimento das bactrias. O espalhamento da toxina
pelo corpo, pelos vasos sanguneos e linfticos, pode resultar em disfuno neuronal e do miocrdio.
RETCULO ENDOPLASMTICO
O retculo endoplasmtico (RE) formado por um sistema de membranas interconectadas na forma de tubos ramificados, s vezes na forma de cisternas, que delimitam uma cavidade mais conhecida como luz. Podem-se distinguir dois tipos de retculo. O retculo endoplasmtico rugoso (RER) ou granular apresenta ribossomos associados e uma estrutura na forma de cisternas. Clulas com intensa sntese proteicas, como as clulas acinosas do pncreas, possuem RER bastante desenvolvido. Na ausncia de ribossomos, o retculo endoplasmtico denominado liso (REL) ou agranular, formando estruturas predominantemente tubulares. Clulas com retculo liso abundante esto relacionadas sntese de hormnios esteroides, como as clulas de Leydig nos testculos; degradao de glicognio, como os hepatcitos; ou a funes especficas, como o controle do clcio citoplasmtico nas clulas musculares. Nestas ltimas, o RE recebe a denominao especfica de retculo sarcoplasmtico. Retculos rugosos e lisos podem estar presentes numa mesma clula, formando uma estrutura contnua. A associao temporria dos ribossomos s membranas do RE determinada pelo estado fisiolgico da clula, ou seja, reas de REL podem ser substitudas por RER no caso de respostas celulares que envolvem intensa sntese proteica. O inverso tambm pode ocorrer. Havendo a necessidade de eliminao de resduos de fenobarbital (um anestsico que pode se acumular, chegando a nveis txicos para o organismo), reas de RER dos hepatcitos so substitudas por REL, com capacidade de destoxificao, Essa capacidade de converso, assim como a sua observao em clulas vivas, faz do RE uma organela bastante dinmica. Uma caracterstica estrutural do RE a continuidade com o envoltrio nuclear. No entanto, no existe nenhuma ligao direta das membranas do RE com a membrana plasmtica. O RE encontrado na maioria das clulas eucariticas, nas quais ocupa, em mdia, 10% do volume celular, correspondendo a mais da metade do total de membranas presentes em uma clula animal. A quantidade de RE e sua localizao no citoplasma variam de acordo com o tipo e o metabolismo celular. Nos hepatcitos, o RE uma estrutura bastante desenvolvida que aparece disperso por todo o citoplasma. Em clulas
secretoras polarizadas, como as clulas pancreticas, o RER fica restrito preferencialmente poro basal do citoplasma, em geral prximo ao ncleo.
Composio qumica Membranas semelhana das demais biomembranas, as membranas do RE so formadas por uma bicamada lipdica com protenas associadas. Os lipdios presentes correspondem a 30% do seu contedo, sendo principalmente fosfolipdios com cidos graxos de cadeias curtas e insaturadas. O contedo de colesterol e glicolipdios baixo. Os componentes lipdicos esto dispostos assimetricamente nas membranas do RE. Pode-se citar a presena de fosfatidilinositol, fosfatidiletanolamina e fosfatidilserina, preferencialmentena face externa ou citoplasmtica, enquanto fosfatidilcolina e esfingomielina esto presentes em maior quantidade na face interna ou luminal.
Aspectos funcionais O RE est relacionado sntese, modificao e transporte de protenas e lipdios. Esses componentes podem seguir trs destinos diferentes: permanecer no RE, seguir para outras organelas ou ser encaminhado paa o exterior da clula por meio da secreo. Neste ltimo caso, o RE o incio da via biossinttica-secretora da clula. interessante ressaltar que o sistema de canais formado pelo RE delimita uma regio isolada do citosol. Na luz do RE, acontecem reaes bioqumicas especficas, como a formao de pontes dissulfeto, glicosilaes e outras modificaes estruturais de protenas e lipdios. Como foi descrito anteriormente, o RE assume diferentes funes, dependendo do tipo celular e do estado metablico.
Sntese proteica O RER apresenta-se bastante desenvolvido nas clulas com intensa sntese de protenas destinadas secreo. Os ribossomos que aparecem associados ao RE so idnticos aos ribossomos livres no citosol. A associao com o RE transitria e, to logo a leitura do RNAm termine, o ribossomo liberado no citoplasma at ser novamente engajado na sntese proteica. Podem ser encontrados polissomos associados s membranas do RE. A interao de ribossomos com a membrana do RE sugere que a transferncia de protenas
para a luz do RE ocorra durante sua traduo pelos ribossomos, ou seja, cotraducionalmente, enquanto a importao de protenas em outras organelas (como mitocndrias, peroxissomos, cloroplastos e ncleo) ocorre pstraducionalmente. Algumas protenas podem ser transportadas j em sua forma final para p RE. Essa situao ocorre apenas para um pequeno nmero de protenas em mamferos, mas mais frequente em bactrias e em fungos. A seguir, sero comentados aspectos da transferncia co-traducional de protenas para a luz do RE em clulas de mamferos.
A hiptese do sinal A maquinaria bsica para a sntese proteica que ocorre junto ao RE a mesma encontrada no citoplasma. Como ento, em alguns casos, a sntese proteicas direcionada para o RER? Experimentos reaizados na dcada de 1970 demonstraram que, se a sntese de protenas normalmente destinadas ao RE fosse realizada em um sistema in vitro e na ausncia de membranas (microssomos), resultaria em um peptdeo com cerca de 20 aminocidos a mais do que o mesmo peptdeo produzido tambm in vitro, mas na presena dos microssomos. Essa seqncia de 20 aminocidos localiza-se na extremidade N-terminal das protenas destinadas ao RE e, apesar de no ser constante para todas as protenas, composta predominantemente por aminocidos hidrofbicos. Outros experimentos demonstraram que se essas sequncias hidrofbicas N-terminais fossem adicionadas a protenas normalmente produzidas no citoplasma, elas seriam direcionadas para o RE. A partir dessas observaes, foi formulada a hiptese do sinal. A sequncia Nterminal hidrofbica, denominada peptdeo sinal, sistetizada em ribossomos livres no citoplasma e tem a funo de encaminhar a sntese proteicas para o RE. O peptdeo sinal clivado posteriormente e no permanece na forma final da protena.
Transferncia co-traducional To logo o peptdeo sinal traduzido, uma partcula citoplasmtica denominada Partcula de Reconhecimento do Sinal (PRS) liga-se a ele. A PRS uma ribonucleoprotena composta por um RNA 7S (RNA PRS) e 6 subunidades proteicass (PRS 9, PRS 19/14, PRS 54, PRS 68/72). A subunidade de 54 kDa (PRS 54) responsvel tanto pelo reconhecimento e ligao com peptdeo sinal quanto pela interao da PRS com o ribossomo. Ocorre, ento, o direcionamento do complexo peptdeo nascenteribossomo para o RER, onde existe um receptor para a PRS. A ligao do receptor PRS garante tambm o acoplamento do ribossomo e do peptdeo
membrana do RE. Neste momento, a PRS j terminou seu papel no direcionamento da sntese proteica para p RE e, pela hidrlise de GTP, h o desligamento da PRS de seu receptor, permitindo a reciclagem de ambos e a continuidade da sntese proteicas junto membrana do RE. To somente aps a ligao do ribossomo ao RE e desligamento da PRS, a sntese proteica reiniciada. A ligao do ribossomo membrana do RE ocorre por meio de sua ligao com receptores como a p180. Alm disso, o complexo proteico sec61 parece estar diretamente relacionado ancoragem dos ribossomos. Na fase de associao dos ribossomos membrana do RE, ocorre tambm a ligao da cadeia polipeptdica nascente a um poro aquoso de 2nm de dimetro. Esse um poro fisiolgico que s se forma na presena de complexo ribossomopeptdeo nascente na membrana do RE.
COMPLEXO DE GOLGI
Grande parte das protenas e lipdios, sintetizados no retculo endoplasmtico (RE) tem como destino a superfcie celular ou mesmo o meio extracelular. A Via Biossinttica Secretora composta pelo RE, stio de sntese das substncias; pelo complexo de Golgi (CG) e pelas vesculas de transporte. Juntos, eles promovem o processamento, a seleo e o transporte das substncias a serem secretadas da clula. O CG faz parte do sistema citoplasmtico de membranas, sendo composto por sculos achatados independentes, mas com contnua troca de material por vesculas. Geralmente, o CG est localizado em regio prxima ao ncleo. Na maioria das clulas animais, o CG costuma ocupar a regio central. J em clulas polarizadas, como cinos ou clulas do epitlio intestinal, comum que os elementos CG estejam voltados para a face secretora da clula. Clulas especializadas em secreo como aquelas produtors de hormnios e enzimas digestivas, tm grande parte do citoplasma preenchido por uma rede de CG e vesculas de vrios tipos. O CG est presente em quase todas as clulas eucariticas, no sendo encontrado em apenas em algumas clulas muito especializadas, como as hemcias ou os espermatozoides completamente diferenciados. Em relao s clulas vegetais, cada pilha de sculos do CG tambm conhecida pelo termo dictiossomo. Em geral, existe uma relao especial do RE com o CG, que se posiciona entre o RE e a membrana plasmtica. Nas proximidades do CG, no
so encontrados ribossomos, glicognio ou mitocndrias. Alm da participao do CG na seleo e transporte dos produtos de secreo, tambm h a participao dessa organela no processamento de protenas e lipdeos, principalmente por meio de glicosilao, sulfatao e fosforilao.
Composio qumica Membranas As membranas dos diferentes compartimentos do CG apresentam composio e espessura variveis. A espessura das cisternas varia entre 5 e 10 nm, isto , com valores intermedirios entre aqueles observados para o RE e a membrana plasmtica. Os lipdios compreendem de 35 a 40% dos componentes das membranas do CG e esto representados principalmente por fosfolipdios, distribudos assimetricamente na bicamada. As protenas de membrana correspondem a 60-65% da bicamada lipdica, sendo representadas em sua maior parte por enzimas, protenas estruturais e protenas envolvidas na formao e direcionamento de vesculas. Esto presentes principalmente transferases envolvidas nas etapas de processamentos de lipdios, protenas e polissacardeos presentes na luz do CG, podendo ser citadas as glicosiltransferases, sulfotransferases e fosfatases. interessante ressaltar que o contedo enzimtico caracterstico para cada compartimento do CG, uma vez que as reaes bioqumicas acontecem de maneira sequencial em compartimentos especficos desta organela. Por exemplo, a enzima tiamina pirofosfatase, presente na rede Golgi trans, utilizada como marcadora para o CG em anlises morfolgicas e bioqumicas por no ser encontrada em outros compartimentos celulares.
Aspectos funcionais Nos diferentes compartimentos do CG, as protenas e os lipdios provenientes do RE sofrem importantes modificaes estruturais, dentre as quais se destacam glicosilao, sulfatao e fosforilao, O processamento dessas protenas e lipdios, que em alguns casos iniciado ainda no RE, fundamental para que essas molculas desempenhem adequadamente suas funes. O CG tambm um importante stio de reconhecimento e de encaminhamento de compostos. Ele promove o endereamento e transporte de compostos para endossomo tardio, para a membrana plasmtica e tambm para o meio extracelular (via biossinttica secretora) (transporte antergrafo) e
para o RE (no caso de redirecionamento de protenas residentes do RE) (transporte retrgrado). Outra relevante funo desempenhada pelo CG a sntese de hemiceluloses e pectinas, importantes polissacardeos que compem a parede celular vegetal.
As vesculas de transporte A formao de vesculas a partir de um compartimento doador se d atravs do processo de brotamento. Para que isso ocorra, determinada regio da membrana desse compartimento se curva, aproximando-se at se fundir, liberando, assim, uma vescula. Geralmente, a curvatura na membrana imposta pelo agrupamento de protenas especficas, que permanecem como um revestimento externo nas vesculas liberadas. Tais protenas so conhecidas como protenas de cobertura. Alm dessa funo, as protenas de cobertura possibilitam a seleo das substncias a serem transportadas nessas vesculas. Diferentes classes de coberturas vesiculares podem ser reconhecidas ao microscpio eletrnico e cada uma desempenha papeis especficos no transporte vesicular, sendo responsveis por etapas distintas desse transporte. Atualmente, so facilmente reconhecidas a cobertura de clatrina, a cobertura formada por protenas COP I (COat Protein I) e a cobertura de protenas COP II (COat Protein II) .
Lisossomos Os lisossomos foram inicialmente identificados como organelas membranosas que continham as atividades da enzima fosfatase cida e de, pelo menos, mais quatro hidrolases, que apresentavam funcionamento timo em pH cido. Os trabalhos do Francs De Duve contriburam muito para a identificao dos lisosssomos. Aps caracterizar essas atividades enzimticas e propor que elas estivessem contidas no interior de organelas membranosas, De Duve et al. puderam utilizar a microscopia eletrnica na caracterizao morfolgica dos lisossomos. Na verdade, os aspectos estruturais dos lisossomos so bastante variveis. Como o repertorio enzimtico tambm varia em decorrncia de uma grande diversidade funcional, parece mais plausvel definir os lisossomos como uma famlia de organelas com caractersticas bsicas comuns. Os lisossomos so organelas citoplasmticas que acumulam cerca de 40 enzimas hidrolticas, que apresentam uma ampla gama de substratos. A principal funo dessas organelas a digesto intracelular. Esse papel
extremamente importante, pois permite clula eliminar pores envelhecidas ou danificadas do citoplasma, incluindo organelas e molculas, e degradar componentes oriundos da endocitose, sejam eles fragmentos da membrana plasmtica, macromolculas, partculas, outras clulas ou microorganismos.
Estrutura Os lisossomos so estruturas geralmente esfricas e de tamanho extremamente varivel, delimitadas por membrana. A identificao dessas organelas ao microscpio eletrnico depende da localizao de marcadores especficos, como a atividade da fosfatase cida, ou da presena de resduos dos processos de digesto. Os lisossomos acumulam, ao longo do tempo, resduos no digerveis. Estes so eliminados por alguns tipos celulares, como os organismos unicelulares de vida livre, mas dificilmente por alguns tipos celulares dos organismos multicelulares. A observao desses resduos consiste em elemento adicional identificao ultraestrutural dos lisossomos. Os lisossomos apresentam uma cobertura de carboidratos que fica associada fase interna da membrana que os envolve e, aparentemente, responsvel por evitar a digesto da prpria membrana pelas hidrolases que se acumulam no seu interior. Nas clulas vegetais, os lisossomos aparecem em diferentes formas, acoplando funes adicionais digesto intracelular.
Crinofagia As clulas secretoras que, em determinado momento deixam de receber o estimulo para secreo, precisam eliminar os grnulos onde seus produtos so acumulados no citoplasma. Neste caso, acionado o processo denominado crinofagia, no qual os grnulos so digeridos pela via lisossomal. Dessa forma, o material oriundo da via biocinttica encaminhado para o interior dos lisossomos. A clula volta, ento, para um estado basal de atividade, at que um novo estmulo induza novo acmulo de secreo.
Endocitose A endocitose envolve trs mecanismos principais: a pinocitose (quando a clula engloba parte da fase fluida do meio externo), a fargocitose (quando a clula engloba partculas slidas) e aquele que envolve a endocitose mediada por receptores.
A pinocitose um processo continuo de formao de vesculas que trazem gua, pequenas molculas e protenas solveis. Geralmente, as vesculas de pinocitose so pequenas e numerosas. Uma vez dentro das clulas, as vesculas fundem-se entre si e aos lisossomos resultando na digesto do seu contedo. Esse processo permite a ingesto de volumes excessivos de lquido e resulta tambm na ingesto de grandes extenses da membrana plasmtica, que em sua maior parte reciclada. A taxa de pinocitose e de internalizao de membrana plasmtica varia de clula para clula, sendo especialmente alta em clulas como macrfagos. Neste caso, a reciclagem dos segmentos de membrana fundamental para a manuteno de sua extenso e do volume celular. J a fagocitose corresponde internalizao de partculas relativamente maiores. Em organismos unicelulares, a fagocitose corresponde a um processo importante para a obteno de nutrientes. J nos organismos multicelulares, a fagocitose desempenha um papel importante na defesa contra microorganismos; na eliminao de clulas danificadas, envelhecidas ou em processo de morte celular, alm de ser essencial nos processos de remodelao durante a embriognese e na cicatrizao. Dentre as clulas fagocticas, existem os neutrfilos e, principalmente, os macrfagos. Os fagcitos depende da ligao de partculas presentes no meio extracelular a receptores presentes na superfcie das clulas, levando primeiro adeso dessas partculas e, em seguida, projeo de membranas que as envolvem e as internalizam. Neste processo, h uma participao efetiva dos filamentos de actina e a vescula contendo a partcula a ser ingerida denomina-se fagossomo. No caso das clulas do sistema imune referidas antes, a ligao se d com pores de imunoglobulinas que se ligam partcula a ser internalizada. Essas clulas so conhecidas como fagcitos profissionais. J clulas cuja funo primordial no a fagocitose, mas que podem exercer essa atividade em situao especfica, so conhecidas como fagcitos no profissionais ou eventuais. Fibroblastos e clulas epiteliais so exemplos de fagcitos no-profissionais. As hemcias so removidas da corrente circulatria por um processo fagcito que acontece no fgado pelas clulas de Kpfer. A exposio de resduos de galactose na superfcie das hemcias de resduos de galactose na superfcie das hemcias, devido clivagem de acares mais terminais por enzimas circulantes no plasma, permite o reconhecimento dessas clulas por molculas semelhantes s lectinas presentes na superfcie das clulas de Kpfer. A endocitose mediada por receptores permite s clulas a internalizao de molculas pela formao de vesculas de maneira bastante especfica. A
especificidade desse processo depende da existncia de receptores na membrana plasmtica. Esses receptores conhecem as molculas a serem internalizadas, agrupam-se no plano da membrana e formam os coated pits a partir da associao da poro citoplasmtica dos receptores com as molculas de clatrina. A associao com a clatrina permite a invaginao do segmento da membrana e o brotamento da vescula recoberta por clatrina no citoplasma. Uma das caractersticas desse processo a concentrao, em duas etapas, do material a ser internalizado. Primeiramente, ao se ligar aos receptores, as molculas so concentradas junto superfcie da clula, reduzindo o segmento de membrana a ser ingerido. Esse mecanismo utilizado no controle remoo de receptores da superfcie celular. comum que, a partir das vesculas de endocitose, os receptores sejam reciclados para a membrana plasmtica. Porm, em outros casos, eles so destinados digesto entro dos lisossomos, juntamente com os seus ligamentos.
QUESTES
O que clula?
Resp. Clula a unidade bsica da complementaridade entre estrutura e funo.
vida
em
que
existe
uma
Do que uma clula revestida?
Resp. As clulas so revestidas por uma membrana plasmtica, tambm denominada plasmalema, de constituio lipoproteicas.
Descreva a diferena entre as estruturas primrias, secundrias, tercirias e quartenrias das protenas.
Resp. Protenas so as mais abundantes molculas presentes nos organismos vivos exceo da gua, podendo perfazer 50% da matria seca deles. No ser humano, por exemplo, cerca de 15% da massa corprea vem das protenas. Grande parte das protenas formada apenas por aminocidos e, por isso, elas recebem o nome de protenas simples. Entretanto, h protenas denominadas protenas conjugadas, pois possuem outras molculas ou tomos (grupo prosttico), alm dos seus aminocidos formadores. Em decorrncia, medida que novos aminocidos so incorporados molcula de protena que est sendo sintetizada, comear a haver a atrao entre o oxignio e o H de ligaes peptdicas distintas. Essas interaes determinam uma estrutura especial caracterstica. Que denominada estrutura secundria da protena. Assim, estrutura secundria de uma protena a forma que ela toma em decorrncia das pontes de hidrognio entre oxignio e hidrognio ligado ao nitrognio de ligaes peptdicas distintas. A estrutura secundria, resultante das interaes por ligaes de hidrognio especficas dos tomos das ligaes peptdicas, posiciona os grupos R dos diferentes aminocidos, de tal modo que pode provocar novos tipos de interao. Assim, a estrutura tridimensional total de molcula, agora com a somatria dessas interaes, denomina-se estrutura terciria da protena.
Algumas protenas, para adquirirem seu estado funcional, necessitam mais de uma cadeia polipeptdica (com sua estrutura tridimensional definida). Ao estado funcional, que conta com as estruturas tercirias de dois ou mais polipeptdios unidos, geralmente, a um grupo prosttico, d-se o nome de estrutura quaternria da protena. O exemplo mais conhecido de protena que tem estrutura quaternria a hemoglobina, formada por 4 cadeias (duas denominadas a e duas b), cada uma delas envolvendo um grupo heme contendo ferro. 4. CARVALHO, Isabel C. M. Educao ambiental: a formao do sujeito ecolgico. 3. ed. So Paulo: Cortez, 2008. cap. 1, 3 e 5.
Cap. 1 A epistemologia da Educao Ambiental: A crise de um modo de conhecer e a busca de novos modos de compreender.
Uma das questes que atravessam a crise do conhecimento na contemporaneidade, afetando diretamente a educao, diz respeito a como se pode acessar a realidade e compreend-la. Uma dessas vises a que pensa a realidade como algo mediado pela dimenso simblica (cultural), e situa a produo do conhecimento e, consequentemente, a educao dentro de processos compreensivos e interpretativos, produtores de subjetividades e modos de vida. A outra aquela que tende a tomar a realidade como algo passvel de ser captado em si, como um fato que dispensa a mediao da cultura. Esta viso funda, ao mesmo tempo, um mtodo cientfico e um modo de pensar que se firmou como prprio da racionalidade moderna, tambm chamada de razo instrumental.
O paradigma cientfico-moderno, sua crise e as conseqncias para a educao.
Um dos legados do pensamento caracterstico da modernidade foi a construo de um modelo de racionalidade que alcanou sua formulao mais acabada com o filsofo Ren Descartes (1596 a 1650), no sec. XVII, e representou importante ruptura com modelos anteriores, os quais situavam a compreenso do real em instncias
de legitimao externas ao mundo humano. A razo moderna, com Descartes, trouxe a legitimao do conhecimento para dentro do mundo humano. Assim, j no havia uma instncia externa ao operar como fonte de verdade para compreender a vida. A explicao do mundo e a autocompreenso dos seres humanos eram, agora imanentes, ou seja, estavam no mundo e j no dependiam de uma fonte externa. O acesso verdade imanente ao mundo dava-se por intermdio do prprio sujeito humano, o qual, entendido como sujeito da razo, capaz de pensar logicamente (racionalmente), podia chegar a uma compreenso objetiva do real. Essa mudana de posio, que centrava o sujeito humano e a razo sendo aquele considerado sujeito da razo como fonte do conhecimento verdadeiro, inaugurou a chamada revoluo cientfica e fundou a modernidade. Buscando diferenciar-se de modelos anteriores de pensar, que viam a natureza como uma totalidade ou um organismo vivo, marcado pelas relaes de interdependncia dos fenmenos espirituais e materiais, a racionalidade moderna separou rigorosamente o sujeito cognoscitivo e o objeto do conhecimento e afirmou-se como a capacidade de abstrair as qualidades de seu objeto e fazer uma descrio matemtica do real. Para tanto, o encontro do humano com o mundo teve de ser purificado ou ainda desencantado.
A compreenso do mundo teve de isentar-se das paixes, dos afetos, de todo e qualquer tipo de contaminao por sensibilidades, sentidos, propriedades anmicas, cosmolgicas e modos de experienciar o real no correspondente ao modelo da razo
No mtodo cientfico, a separao entre sujeito e objeto desdobrouse em outras polaridades excludentes com as quais aprendemos a
pensar o mundo; natureza/cultura, corpo/mente, sujeito/objeto, razo/emoo. Somos seres de nosso tempo e, por isso, marcados por essa tradio do pensamento ocidental. Tal maneira de ver o mundo, a qual tem sido denominada de paradigma moderno, entrou em crise justamente por no conseguir responder, adequadamente, aos novos problemas tericos e prticos que atravessam a vida contempornea, entre os quais os ambientais. Ao separar radicalmente a natureza da cultura, a cincia sacrificou a diversidade em nome da universalidade do conhecimento, reduzindo os fenmenos culturais s determinaes das leis naturais gerais. Nesse sentido, os saberes da Fsica e da Biologia tiveram grande ascenso e ganharam legitimidade como portadores do conhecimento verdadeiro do real, e explicativo do humano. J as cincias humanas, entre elas a Educao, neste quadro da hegemonia de uma cientificidade objetivista, ocuparam o lugar menos valorizado, devendo espelhar-se na cincia objetiva para um dia alcanarem tal padro de racionalidade e objetividade.
A interdisciplinaridade: um desafio epistmico e metodolgico.
Ateno: (...) A interdisciplinaridade jamais ser uma posio fcil, cmoda ou estvel, pois exige nova maneira de conceber o campo da produo de conhecimento buscada no contexto de uma mentalidade disciplinar.
Trata-se de um combate, ao mesmo tempo externo e interno, no qual reorganizao das reas e das formas de relacionar o conhecimento corresponde reestruturao de nossa prpria maneira de reconhecer e nos posicionar perante o conhecimento, desfazendo-nos dos condicionamentos histricos que nos constituem. A crise ambiental, de certa forma, alimenta questionamentos epistemolgicos e desacomoda os modos j aprendidos de pensar da racionalidade moderna, ao expor a insuficincia dos saberes disciplinares e reivindicar novas aproximaes para que se compreenda a complexidade das inter-relaes na base dos problemas ecolgicos.
A retido disciplinar e os caminhos hbridos da educao ambiental
A despeito da vocao renovadora da EA, nesse choque de paradigmas ou nessa transio paradigmtica -, como alguns autores preferem caracterizar o momento presente, as armadilhas de uma cientificidade normativa e tecnicista seguem influenciando a esfera educativa. No campo mental, essa perspectiva busca reafirmar-se em certo otimismo tecnolgico que v nas tecnologias ambientais e novos mercados verdes a via rgia para a soluo da crise ambiental. A EA seria, ento, equivocadamente tomada como um sucedneo do ensino de cincias, na acepo de uma cincia morta mas ainda bem viva enquanto idia-fora disputando
hegemonia dentro do campo cientfico. E correria o risco de orientar-se predominantemente para a difuso de conhecimentos cientficos e tecnologias ambientais tomados em sua forma ingnua sem a devida problematizao de seus contextos histricos de produo e dois interesses econmicos aos quais respondem, sendo, reafirmados como conhecimentos desinteressados, em si mesmos verdadeiros e eficazes para a crise ambiental.
Conectando a vida e o conhecimento no cotidiano da prtica educativa
No mundo vivido, os aspectos tomados isoladamente pelas disciplinas esto permanentemente relacionados, como a trama de um s tecido. Ao puxar apenas um fio, tratando-o como fato nico e isolado, cada rea especializada do conhecimento no apenas perde a viso do conjunto, como tambm pode esgarar irremediavelmente essa trama em que tudo est imbricado. Com isso, a multiplicidade das camadas de significados que constituem a realidade traduzida em fatos unidimensionais, vistos de somente uma perspectiva. Na escola, organizada sobre a lgica dos saberes disciplinares, o resultado que, por exemplo, o professor de geografia no toca nos aspectos biolgicos da formao de um relevo em estudo; o historiador no considera a influncia dos fatores geogrficos na compreenso do declnio de uma civilizao; o professor de biologia no recupera os processos histricos e sociais que interagem na formao de um ecossistema natural, e assim por diante.
A interdisciplinaridade como atitude: a abertura ao dilogo entre saberes e ao trabalho em equipe.
Os problemas ambientais ultrapassam a especializao do saber. Para confirmar essa assero, basta pensar em alguns dos mais conhecidos, como:
a) o aquecimento global e os desequilbrio climticos; b) a poluio dos rios e mananciais, que tem diminudo a oferta de gua potvel e gerado graves conseqncias para a sade, como no caso da contaminao de rios e represas; c) os organismos geneticamente modificados (transgnicos), cujos riscos para o meio ambiente e para a sade humana ainda esto longe de ser compreendidos plenamente. Contudo, para intervir nos riscos ambientais ou para gerir o ambiente de modo que tais riscos sejam evitados, preciso compreender os complexos processos biolgicos, geogrficos, histricos, econmicos e sociais geradores desses problemas. Por isso, as equipes que estudam as questes ambientais e intervm nelas so, em sua maioria, compostas de profissionais de vrias reas que atuam em conjunto e buscam formas interdisciplinares de cooperao entre si e de compreenso da realidade. Com os outros profissionais da rea, o educador ambiental compartilha o desafio gerado pela complexidade das questes ambientais. Isso implica atitude de investigao atenta, curiosa, aberta observao das mltiplas inter-relaes e dimenses da realidade e muita disponibilidade e capacidade para o trabalho em equipe. Significa construir um conhecimento dialgico, ouvir os diferentes saberes, tanto os cientficos como os outros saberes sociais (locais, tradicionais, das geraes, artsticos, poticos, etc.); diagnosticar as situaes presentes, mas no perder a dimenso da historicidade, ou seja, dar valor histria e memria que se inscreve no ambiente e o constitui, simultaneamente, como paisagem natural e cultural.
A educao Ambiental no debate das idias: elementos para uma EA crtica
A EA tem uma proposta tica de longo alcance que pretende reposicionar o ser humano no mundo, convocando-o a reconhecer a alteridade da natureza e a integridade e o direito existncia no utilitria do ambiente. Essa proposta tica tem repercusses, quando se trata de eleger os princpios pedaggicos da EA. Ao construir-se como prtica educativa, a EA posiciona-se na confluncia do campo ambiental e as tradies educativas, as quais vo influir na formao de diferentes orientaes pedaggicas no mbito da EA ou, produzir diferentes educaes ambientais. A intensificao do dilogo com os movimentos sociais tem ampliado a interface entre a esfera educacional e os acontecimentos socioculturais. E isso tem ocorrido no apenas com relao ao ambiente, mas tambm no caso de outras educaes de fronteira, como a educao para a paz, a educao de gnero, a educao para os direitos humanos, etc. O que essas novas educaes tm em comum o fato de tratarem as questes emergentes da vida social em uma perspectiva interdisciplinar, convocando diferentes saberes e reas de conhecimento para compreend-las.
Superando uma viso ingnua de Educao Ambiental
Apesar de atualmente todos concordarem que preciso fazer algo a respeito da crise ambiental, h muitas divergncias e disputas entre diferentes pontos de vista sobre o que fazer, sobre como gerir as questes ambientais, sobre que interesses devem prevalecer na complexa negociao entre os diversos grupos sociais, envolvendo seus projetos e vises de mundo, sobre as
necessidades do presente e as expectativas de futuro que podemos construir em conjunto. Educao ambiental tem sido a expresso cada vez mais utilizada nos textos das polticas e programas de educao e de meio ambiente, bem como nos projetos comunitrios de extenso, de gesto e de ao. O uso cada vez mais corrente e generalizado da denominao Educao Ambiental pode contribuir para uma apreenso ingnua da idia contida nela, como se fosse uma reunio de palavras com poder de abrir as portas para um amplo e extensivo campo de consenso. A expresso Educao Ambiental passou a ser usada como termo genrico para algo que se aproximaria de tudo o que pudesse ser acolhido sob o guarda-chuva das boas prticas ambientais ou ainda dos bons comportamentos ambientais.
Elementos para uma Educao Ambiental crtica
A construo de uma EA crtica implica a explicitao de algumas posies terico-metodolgicas. A primeira delas diz respeito viso de educao como um processo de humanizao socialmente situado. Nessa definio, a prtica educativa processo que tem como horizonte formar o sujeito humano enquanto ser social e historicamente situado. Segundo tal perspectiva, a educao no se reduz a uma interveno centrada no indivduo, tomado como unidade atomizada e solta no mundo. A formao do indivduo s faz sentido se pensada em relao com o mundo em que ele vive e pelo qual responsvel. Na EA esta tomada de posio de responsabilidade pelo mundo em que vivemos, incluindo a a responsabilidade com os outros e com o ambiente. O projeto poltico-pedaggico de uma EA crtica poderia ser sintetizado na inteno de contribuir para uma mudana de valores e atitudes, formando um sujeito ecolgico capaz de
identificar e problematizar as questes socioambientais e agir sobre elas.
Ultrapassando a fronteira entre a educao formal e a noformal
A EA tem sido uma fora potencializadora para construir pontes e aproximar a educao formal da no-formal. Muitos trabalhos nessa rea passam-se justamente na fronteira do formal e do no-formal, integrando a escola e as comunidades do entorno. Esses trabalhos geralmente incluem aes que envolvem alunos dentro e fora da escola, chegando a propor novos contedos escolares ou orientaes curriculares. Fecha-se, assim, um crculo virtuoso formado pela aprendizagem escolar e social desenvolvida nas comunidades.
Atitude, comportamento e ao poltica: elementos para pensar a formao ecolgica
A formao de uma atitude ecolgica pode ser considerada um dos objetivos mais perseguidos e reafirmados pela EA crtica. Essa atitude poderia ser definida, em seu sentido mais amplo, como a adoo de um sistema de crenas, valores e sensibilidades ticas e estticas orientando segundo os ideais de vida de um sujeito ecolgico. As atitudes orientam as decises e os posicionamentos dos sujeitos no mundo. Nesse sentido, quando falamos em atitude, devemos diferenci-la da noo de comportamento. Atitudes so predisposies para que um indivduo se comporte de tal ou qual maneira, e assim podem ser preditivas de comportamento. Contudo, no h determinao do tipo causa e efeito que nos permita traar correspondncia direta entre a formao de atitudes e o plano dos comportamentos. Estes ltimos so as
aes observveis, efetivamente realizadas, e podem estar ou no de acordo com as atitude do sujeito
5. DEAN, W. A ferro e fogo: a histria e a devastao da Mata Atlntica brasileira, So Paulo: Companhia das Letras, 1996.
Cap. 1 A Revoluo Da Floresta. Estamos voando, a sete mil metros de altitude, rumo ao sul do subcontinente brasileiro na direo da cidade do Rio de Janeiro. Comparadas com a impressionante produtividade, abundncia e variedade das florestas maravilhosas para contar. Os relatos sobre essas florestas famintas dizem respeito resistncia de pioneiros desesperados, no ao estabelecimento de grandes santurios da vida. Elas so to resistentes e descomplicadas que toda a devastao imposta pelos homens no impede de se reconstiturem, a menos que sejam cobertas com asfalto. O mesmo no se pode esperar em relao s florestas tropicais, que podem quase certamente ser destrudas, mas talvez nunca se restabeleam nos lugares de onde foram eliminadas. Da a tragdia. A destruio dessas florestas irreversvel no mbito de qualquer escala temporal humana.Quando a floresta tropical destruda, a perda em termos de diversidade, complexidade e originalidade no apenas maior que a de outros ecossistemas: incalculvel. A histria de destruio das florestas da Amrica do Sul, contudo, um caso at certo ponto acessvel a nossa curiosidade. A histria chegou tarde a praticamente todos os outros encontros entre o homem e a vida selvagem. Para o homem, a coexistncia com a floresta tropical sempre foi problemtica. Antagonismo no , em nenhum momento, parte necessria da relao, mas a limpeza da floresta no mero ditame do preconceito ou orgulho cultural ou de arranjos polticos e sociais imprevidentes.O avano da espcie humana funda-se na destruio de florestas, uma vez que ela est mal equipada para que se possa habit-la. Na costa leste da Amrica do Sul, estendia-se outrora uma imensa floresta ou, mais precisamente, um complexo de tipos de florestas, em geral latifoliadas, pluviais e de tropicais e subtropicais. Entre oito e 28 de latitude sul, interiorizava-se a cerca de cem quilmetros da costa no norte e alargavase a mais de quinhentos quilmetros no sul. No total, a floresta cobria cerca de 1 milho de quilmetros quadrados. Esse complexo tem sido chamado de Mata
Atlntica brasileira, associado outra muito maior, a Floresta Amaznica,mas distinto dela. Em conjunto, as duas florestas formavam uma zona biogeogrfica diferente e mais rica em espcie que as outras florestas tropicais do planeta,situadas na frica e no Sudeste Asitico. A Mata Atlntica era, em si mesma, de uma diversidade extraordinria, levando-se em conta seu tamanho relativamente modesto. A floresta ocorreu ao longo dessa costa devido a relevo, regimes de vento e correntes ocenicas. Dado o grande mbito de latitude ao longo do qual se espalha a Mata Atlntica, ocorreu outro gradiente de temperatura e insolao. Mudanas sutis podem, assim, ser observadas na composio da floresta entre seus extremos norte e sul. Na verdade,essas formaes se interpenetraram. No sul, a floresta subtropical acompanha o litoral martimo mais quente e penetra nas baixadas do rio Paran. O solo foi menos determinante que a chuva e a temperatura no estabelecimento da Mata Atlntica. Exceto pelas faixas litorneas de dunas, seus solos tiveram origens granticas, baslticas e gnissicas antigas, altamente intemperizados e,consequentemente,de baixa fertilidade. A Mata Atlntica uma cobaia dessa ordem. Graas a seu tamanho avantajado, sua complexidade e variabilidade,biogegrafos e ecologistas tatearam em busca de designaes comuns para seus componentes,mas seu carter distinto indiscutvel. Esta histria da Mata Atlntica no uma histria natural; ou seja, no uma explicao das criaturas da floresta e das relaes que estas mantm entre si. ,antes, um estudo da relao entre a floresta e o homem. Podemos visitar a floresta tropical e at especializarmo-nos na extrao das mil e uma raridades que ela oferece, mas no moramos nela, exceto em desespero. O morador humano brasileiro da floresta vive em suas margens, perto de cursos de gua ou campos. difcil dizer se correto referir-se Mata Atlntica no tempo presente. Certa extenso de cobertura florestal ainda existe na regio e, embora a maior parte seja identificvel como secundria, algumas pequenas faixas podem jamais ter sido derrubadas ou queimadas pelo homem. Mesmo para os moradores da floresta difcil decidir sobre a identidade de determinada rvore, e empregam nomes que incluem diversas espcies e at diversos gneros. rvores da Mata Atlntica que se elevam a 35 metros ou mais, em suas reas de crescimento mais luxuriante, podem ter mais de cem anos, embora algumas, de circunferncia muito grande, possam ser muito mais velhas, com idade de at mil anos. No se sabe quanto tempo pode levar o processo de recomposio. A floresta secundria, do tipo que ocorre quando se abandona uma clareira que foi queimada para agricultura,em vinte ou trinta anos, pode atingir exuberncia prxima da floresta intocada.Uma nica copa de rvore pode abrigar mil espcies de insetos e a Mata Atlntica como um todo pode ter abrigado um
milho delas,das quais apenas pequena percentagem foi,ou ser um dia, batizada pelos cientistas. As plantas e animais da Mata Atlntica tambm eram raros no sentido de que muitos deles no seriam encontrados em nenhuma outra floresta. Dentre suas espcies arbreas, mais da metade era endmica.Outros 8% eram compartilhadas com a Floresta Amaznica. Diversos locais na Mata Atlntica tm sido postulados como centros de endemismo de pssaros,mamferos, rpteis e insetos,contendo numerosas espcies que no haviam conseguido expandir sua rea de ocorrncia alm de suas fronteiras ou que ali haviam encontrado um refgio quando sua rea de ocorrncia anterior se tornou restrita.Certos beija-flores, sanhaos, marsupiais,preguias, sagis, pererecas,patiobas e inmeros outros seres esto limitados Mata Atlntica. A especializao implica maior eficincia, e a Mata Atlntica foi realmente eficiente na captao de energia solar, absoro de nutrientes do solo, da gua da chuva e atmosfera e na reciclagem e intercmbio de recursos.O resultado foi uma biomassa vegetal que,em alguns lugares,pode ter chegado a seiscentas toneladas por hectare e a uma capacidade de gerar talvez cinquenta toneladas de biomassa por ano. Quando e como surgiu a Mata Atlntica H mais de 400 milhes de anos,plantas vasculares fotossintetizadoras e multicelulres comearam a aderir s margens dos continentes. Logo esses organismos cobriram as costas rochosas,desaceleraram o fluxo das guas,formaram solos e avanaram para o interior. Formaram-se radculas absorventes de nutrientes, estames se achataram em folculos que captavam a luz solar e sementes substituram esporos como agentes de disperso.Por 50 milhes de anos,as formas de vida da Mata Atlntica evoluram localmente,sem sofrer transtornos geolgicos adicionais. O ltimo perodo glacial alcanou seu pice de 25 a 18 mil anos trs; h no menos de 12 mil anos, as geleiras sul-americanas se retiraram para seus santurios atuais no topo dos picos mais elevados dos Andes. Nesse instante do tempo geolgico, a Mata Atlntica mais uma vez se expandia vigorosamente,para ocupar um vasto imprio, 3500 quilmetros ao longo da costa sul-americana, e acima e por sobre a escarpa costeira, na medida em que as chuvas o permitiam. Dessa forma, a moderna Mata Atlntica evoluiu e por fim passou a ocupar suas fronteiras histricas. Cap. 2 A Primeira Leva De Invasores Humanos. A retomada da expanso da Mata Atlntica coincidiu com sua descoberta pelos homens, caadores que subitamente invadiram as plancies sul-americanas h
talvez 13 mil anos. Assim comeou um longo perodo de interao, com resultados muito difceis de avaliar as atuais apreciaes no passam de conjeturas e possivelmente jamais deixaro de s-lo. Para esses caadores migratrios, a fronteira mais remota era a floresta costeira, que abordavam pelo lado continental, interno. Penetravam distncias considerveis na floresta ao longo de riachos, buscando corredeiras onde era fcil apanhar peixes. Afora essas incurses, provvel que o interior da floresta fosse pouco promissor como lugar de moradia,porque os animais ali eram mais raros e difceis de caar. Os caadores-coletores no desapareceram nas regies montanhosas. Levantamentos arqueolgicos recentes em diversos stios de barragens hidreltricas descobriram seus acampamentos, todos ao longo de altos trechos de cursos d gua,normalmente a uma distncia de algumas centenas de metros das correntes, sugerindo talvez o quanto eram, outrora, extensas as florestas de galeria. A julgar por relatos do sculo XIX, possvel que antigos caadores-coletores utilizassem fogo para tocar sua presa e para eliminar a vegetao lenhosa que crescia na plancie, substituindo-a por folhagens tenras para atrair herbvoros esparsos. Essas especulaes so antigas, porque geralmente faltam evidncias. Geraes de observadores tm se perguntado, porm, porque faltavam rvores em reas to grandes do continente na poca em que os europeus o invadiram, j que as rvores podem facilmente ser plantadas em muitas dessas regies onde florescem.O pampa,a plancie gramada que se estendia da Bahia Blanca,na Argentina,por 2300 quilmetros ao norte at a atual cidade de So Paulo, a maior e mais intrigante dessas anomalias. Mesmo que a Mata Atlntica fosse at certo grau modificada pela invaso de caadores-coletores, no poderia ter sido transformada to extensamente como o foram as florestas dos continentes ocupados h muito mais tempo pelo Homo sapiens.Afinal,a Amrica do Sul foi o ltimo continente descoberto pelos homens e sua residncia de quatrocentas geraes foi breve,comparada com as 1600 geraes de manejadores de fogo na Austrlia e as 4 mil geraes na frica. Com o declnio dos grandes animais de caa, alguns dos homens que ainda exploravam a atividade, mudaram-se para a margem das baixadas do continente:para eles,eram os domnios mais distantes de suas expedies. evidente, no entanto, que esses povos tinham pouca necessidade de explorar a floresta. Provavelmente coletavam seus frutos e remdios silvestres; afora isso,viviam de costas para ela, distantes de seus vizinhos do planalto.
Durante a maior parte da era dos sambaquis, as populaes montanhesas na regio da Mata Atlntica persistiram na caa e na coleta.Apesar disso, possvel que seus usos do recurso florestal estivessem se intensificando. Milho foi encontrado em um stio em uma rea de floresta de galeria no atual Estado de Minas Gerais, e datava de 3900 anos atrs.Assim,a agricultura foi empreendida vrios milnios aps sua adoo nos Andes e na mesoAmrica,sugerindo uma longa fase durante a qual os recursos coletados eram suficientes para uma populao humana modesta e itinerante. As culturas mais antigas dos primeiros grupos agricultores, alm do milho,eram nativas das terras baixas da Amrica do Sul, principalmente a mandioca uma raiz, produto bsico em suas diversas variedades e outras razes do gnero ao qual pertence o inhame,e ainda a abbora,o abacaxi e o amendoim. A adoo da agricultura transformou radicalmente a relao dos homens com a floresta. O que havia sido um recurso residual, produto inferior para os caadores-coletores, queimado por descuido ou acidente quando se tocava ou atraa a caa, agora se tornava seu principal habitat.Descobriram que os solos do cerrado eram demasiado arenosos, secos, cidos e saturados de alumnio para cultivar. A agricultura era muito mais vivel nos solos da floresta.Desde o comeo, a agricultura na regio da Mata Atlntica de fato, em todas as reas de baixada do continente exigiu o sacrifcio da floresta. A tcnica era extremamente simples:perto do fim da estao seca, a macega de uma faixa de floresta um hectare mais ou menos era cortada e deixada secar, e,por meio de machados de pedra,retirava-se um anel da casca dos troncos das rvores maiores.Ento,um pouco antes da chegada das chuvas,a rea era queimada,fazendo com que a enorme quantidade de nutrientes na biomassa da floresta casse sobre a terra na forma de cinzas.A floresta, que nunca antes havia sido queimada,no s ficava maravilhosamente frtil, mas tambm livre das sementes de plantas invasoras e, dessa forma, pouca capina era necessria. O abandono da faixa cultivada era provocado, mais provavelmente, pela invaso de ervas daninhas e pragas.As ervas podiam ser desenraizadas mo, mas no havia defesa contra as pragas,entre as quais a mais terrvel era a formiga-cortadeira,mais conhecida como sava. A floresta secundria, ou sucessria,que os indgenas chamam de capoeira terra anteriormente plantada -, que acabava recuperando os campos abandonados,era similar em estrutura e composio ao crescimento que recuperava naturalmente as aberturas no dossel provocadas pelas quedas de rvores.O rebrotar das rvores corrigia o solo ao elevar os nutrientes do subsolo e dispers-los pela queda das folhas e pela proteo dos processos do solo contra a insolao e chuva diretas.
A agricultura itinerante poupava muito trabalho, mas no era de todo simples.A agricultura em si era uma tarefa perigosa e problemtica.Perceber seu momento oportuno exigia um sexto sentido,para que no fosse feita com muita ou pouca antecedncia em relao s chuvas.No podia ser intensa demais para no queimar a camada rasa e frtil do lixo florestal e seus organismos.A agricultura itinerante no implicava a perda de habilidades j adquiridas porque esses agricultores tinham que continuar a caar e a pescar. A agricultura de derrubada e queimada era extraordinariamente redutiva. Quase tudo que estivesse vivo no interior das faixas queimadas era reduzido a cinzas e apenas as cinzas eram aproveitadas.Talvez os primeiros agricultores lamentassem esse desperdcio inicial.Sem dvida,havia episdios de queima que escapavam ao controle,como poderia ter ocasionalmente ocorrido em anos de seca no lado mais mido e de barlavento da Mata Atlntica e,mais comumente, na floresta mais seca do interior. A agricultura pode, portanto,ter reduzido a complexidade e a biomassa em reas considerveis da Mata Atlntica durante os mais de mil anos em que foi praticada antes da chegada dos europeus. O aparecimento da agricultura, ao longo do litoral, parece ter sido resultante de uma migrao de diversos povos das montanhas.Certamente foram atrados pela mariscagem e pelos recursos piscosos dos esturios, mas j eram agricultores consumados. Esses primeiros grupos de agricultores foram varridos da maior parte do litoral, numa extenso que chegou quase ao rio Amazonas, por um outro grupo,cultural e linguisticamente aparentado aos guaranis das bacias dos rios Paraguai e Paran.Esse grupo foi o tupi,que se aproximou da regio da Mata Atlntica comeando por volta do ano 400,talvez na direo da famlia guarani, embora isto no esteja solidamente esclarecido.Mais de mil stios tupis foram localizados no Brasil, todos ao longo do litoral florestado,at quinhentos quilmetros do litoral martimo. Foram principalmente os tupis que os europeus encontraram em suas primeiras investidas ao longo da costa. De fato,este grupo e seus parentes guaranis,os mais numerosos e poderosos habitantes das baixadas, mantiveram, por aproximadamente trs sculos,intenso intercmbio cultural com seus invasores. A base principal da agricultura tupi nas baixadas era a mandioca.O milho, rico em protenas, que poderia impor-lhes maior dependncia da agricultura mas que tambm teria demandado mais nutrientes dos solos da floresta,era empregado apenas como matria-prima alternativa no preparo de uma bebida fermentada que apreciavam muito.
Embora diversificadas e sazonalmente abundantes, essas fontes de protena provavelmente representavam o fator crtico no abastecimento alimentar.Isto quer dizer que possvel que os tupis tenham sentido a escassez de caa e peixe mais cedo que a de seus produtos agrcolas bsicos.Esse desequilbrio talvez tenha salvo a floresta de ser reduzida a uma formao inteiramente secundria. Os tupis no submetiam seus vizinhos escravido e tributos,o que poderia ter estimulado o uso mais intensivo da terra. provvel que nem o pudessem, porque a escala continental do territrio de batalhas sempre permitia aos derrotados escapar escravido pela retirada. O trao cultural mais impressionante dos tupis era a antropofagia. captura de um inimigo em batalha, seguia-se um elaborado e sdico ritual de execuo, muitas vezes na presena de aldeias convidadas.O falecido era ento assado na grelha e distribudo entre os presentes. A questo relevante porque existe a hiptese de que a ingesto de carne humana era uma fonte necessria de protenas para os tupis. Pode-se deduzir das crnicas do sculo XVI que cada aldeia tupi abrigava, em mdia,seiscentas pessoas e controlava pouco menos de setenta quilmetros quadrados do interior, sugerindo uma populao de cerca de nove pessoas por quilmetro quadrado.Com essa densidade, os tupis da baixada teriam provocado, no curso de cinco sculos, uma degradao da Mata Atlntica consideravelmente maior que seus vizinhos do planalto.Supondo novamente o desmate de 0,2 hectare de floresta primria por pessoas/ano,todo o domnio tupi teria estado sujeito a queimada em um lapso de apenas 55 anos. As presses tupis sobre a floresta no resultavam totalmente de impulsos utilitrio-racionais de otimizar a produo e a mo-de-obra ou de melhorar a segurana militar.Havia tambm os fatores imponderveis das animosidades pessoais e angstia existencial que dividiam as aldeias ou provocavam seu abandono. Embora as prticas agrcolas tupis no paream ter sido mais intensivas que as dos povos da baixada que eles desalojaram, sua habilidade na preservao de alimentos pode ter sido um pouco maior.Isso representava uma vantagem militar especial, porque os suprimentos portteis de comida aumentavam a mobilidade e possibilitavam a montagem de expedies maiores por perodos mais longos. Na guerra na floresta, os que estavam na defensiva eram forados a abandonar suas safras e, em ltima instncia, retornar caa e coleta para sobreviver.Em tais circunstncias,os tupis podem bem ter reduzido as presses sobre a floresta ao longo das suas fronteiras nas montanhas.
A hiptese de que a Mata Atlntica foi muito modificada pelos tupis, ao longo da plancie costeira, durante sua ocupao milenar, circunstancial,mas ajudaria a explicar por que os europeus tinham to pouco a dizer sobre a floresta logo que chegaram. Os caadores que irromperam nas savanas sul-americanas, h cerca de 12 mil anos, participaram do grande ato final de ampliao territorial humana.Os homens que brandiam fogo e armas haviam finalmente alcanado uma distribuio continental cosmopolita.Uma vez exaurida a megafauna que os atrara, assumiram postos ao longo da franja florestal, onde o encontro de dois ecossistemas estimulava plantas de crescimento rpido e atraa caa.A floresta era sua antagonista e o fogo era o meio para penetr-la e frear seu avano secular. Agricultura itinerante e guerra interminvel intensificavam a presso humana sobre a floresta. Os cerrados eram inadequados lavoura;desta forma,o novo regime de subsistncia implicava o desvio da populao de um bioma para outro.Ainda que a itinerncia exigisse apenas pequenos insumos de terra e ainda que a populao que ela sustentava fosse limitada,em ltima instncia, pelo suprimento de recursos no-agrcolas,durante longo tempo a lavoura foi capaz de reduzir grande parte da floresta a formaes secundrias.Parece improvvel que alguma parte das baixadas da Mata Atlntica localizadas em stios adequados lavoura tenha escapado de ser derrubada pelo menos uma vez durante essa fase de desenvolvimento cultural. Os grupos que obtiveram controle da Mata Atlntica, ao fim de alguns anos de ocupao humana, demonstraram seu talento para reverter a histria da vegetao.A produtividade superior do solo perturbado os havia atrado quando ainda eram caadores-coletores;a agricultura foi uma racionalizao e uma intensificao do ciclo de perturbao.A derrubada da floresta para fins de assentamento, agricultura, comunicao e guerra ampliou em muito a rea de perturbao. A regio da Mata Atlntica, ento, apresenta duas, ou talvez trs, categorias de fronteira:aquela entre a floresta e o cerrado ,com temperatura gradualmente mais quente e precipitao pluviomtrica maior favorecendo a floresta,que desintoxicava o solo medida que avanava e abrigava os microclimas adequados para sua posterior consolidao e desenvolvimento; aquela da primeira leva de homens que confrontavam a floresta ao longo de sua orla do interior do continente e, consistentemente, se aliavam com o cerrado; e aquela entre os tupis e seus adversrios caadores-coletores.
Cap. 3
A Segunda Leva De Invasores Humanos. Um dos primeiros atos dos marinheiros portugueses que, a 22 de abril de 1500, alcanaram a costa sobrecarregada de floresta do continente sul-americano nos 17 graus de latitude sul, foi derrubar uma rvore. Do tronco desse sacrifcio ao machado de ao, confeccionaram uma cruz rstica para eles,o smbolo da salvao da humanidade.Uma missa foi ento celebrada aos ps dessa cruz,durante a qual, para satisfao dos portugueses,os indgenas ali aglomerados imitaram sua postura ajoelhada com as mos em prece, embora no imitassem suas expresses devotas. Os indgenas que, inocentemente, se irmanaram com eles naquela praia,no faziam ideia, tal como as rvores s suas costas,da destruio que essa invaso causaria. Esse evento memorvel da histria da humanidade o fim de milnios de separao entre os dois maiores contingentes de populao da espcie foi tambm o mais trgico. Incapazes de compreender intelectualmente a magnitude de sua descoberta,os portugueses tropearam em um meio continente,movidos por cobia e virtude, sem se deixarem levar por compaixo ou mesmo por curiosidade.A Mata Atlntica os deixava impassveis ou atnitos.Por diversas vezes penetraram-na,e traziam apenas relatos delirantes sobre esmeraldas e ouro.Produziram tamanha devastao entre seus irmos que, no prazo de um sculo,quase todos aqueles com quem haviam se deparado estavam mortos e suas sociedades em runas.Esse foi o comeo, a fundao do povoamento, da colonizao e do imprio, de uma civilizao transferida e imposta. A expedio se retardou bucolicamente na baa de Porto Seguro. Os capites tiraram sua folga sombra das rvores.Os tripulantes abateram rvores novas para lenha e derrubaram palmeiras para extrair palmito.As viagens pela frica de certo modo haviam acostumado os descobridores a este ambiente novo, tropical. Ao longo da praia, e nas colinas prximas,seu reconhecimento nada lhes relevava alm de matas,compostas de rvores enormes e imponentes, cheias de muitos tipos de pssaros.Os indgenas lhes ofereceram cocares emplumados ,no como meros smbolos de estima,mas porque os consideravam dotados de virtudes mgicas.Cabral os aceitou afavelmente,bem como os arcos e flechas, pssaros canoros e macacos. Assim,pacificamente,comeou a segunda invaso da Mata Atlntica. Os recursos da Coroa portuguesa eram extremamente limitados e na poca se destinavam quase totalmente a empresa do Oriente, que parecia muito mais promissora. Desta forma, D. Manuel entregou a explorao da nova colnia a um grupo de comerciantes,que deviam despachar pelo menos seis navios por ano para extrair pau-brasil e o que mais pudessem encontrar. Nisso foram diligentes:consta que, nos primeiros anos, coletaram cerca de 1200
toneladas/ano.O comrcio era maravilhosamente lucrativo, j que o rei havia providencialmente fechado o mercado madeira corante asitica. Em 1588, 4700 toneladas de pau-brasil passaram pela aduana portuguesa,talvez metade do verdadeiro volume.O trfico francs clandestino de madeiras corantes era to bem estruturado quanto o dos portugueses ou at melhor um cronista de meados de 1550 relatou haver observado 100 mil pedaos de tronco estocados na colnia francesa do Rio de Janeiro.Havia ainda o contrabando intermitente feito por navios espanhis e ingleses.Em conjunto, todos esses negociantes podem ter provocado a extrao de 12 mil toneladas por ano. Em 1605, a Coroa portuguesa,alarmada com os relatrios sobre a explorao de pau-brasil de que,com o corte indiscriminado e a estocagem,as madeiras viro a acabar e perder de todo - , passou a controlar o corte e criou a funo de guardas florestais.A penalidade para a extrao ilegal era a morte.Em 1607,o pau-brasil voltou a ser um monoplio,cujos concessionrios eram autorizados a importar apenas seiscentas toneladas por ano um convite continuidade do contrabando.O mercado,talvez, j estivesse saturado.Segundo um comissrio real enviado para investigar a situao, no se tratava, contudo,de que a rvore estivesse em perigo de extino comercial.Considerando que um volume mdio anual de 8 mil toneladas, no sculo XVI, implicava corte, carreto, estocagem e transporte em chatas de cerca de 320 mil sees de 25 quilos cada,vrios milhares de indgenas teriam sido empregados na explorao.Calcula-se que essa tonelagem exigiu a derrubada de aproximadamente dois milhes de rvores durante o primeiro sculo do trfico. Muitas vezes, as viagens de volta Europa levavam tambm escravos nativos, A prtica foi registrada pela primeira vez em 1511,quando o navio Bretoa, com carga de madeira em Cabo Frio, incluram em seu carregamento 36 escravos, dez homens e 26 mulheres, ainda que os proprietrios houvessem proibido esse trfico tripulao. Fazia tempo que os portugueses comerciavam escravos na costa africana, fornecendo mo-de-obra para suas colnias insulares e para as plantaes no Algarves e sul da Espanha. Os nativos trazidos para Lisboa provavelmente no se destinavam a ser escravos no campo, mas eram vistos como curiosidades, exotismo, e serviram para exibio ou venda a nobres, como macacos ou papagaios, embora fossem apreciados tambm como objetos sexuais.Os tupis, evidentemente, forneciam abundante fonte dessa mercadoriae os portugueses poderiam congratular -se por realizar um gesto humanitrio salvando esses cativos da churrasqueira. A presteza com que os tupis se engajaram no escambo com os europeus foi motivada, em grande parte, pelo desejo de poupar trabalho, expandir sua base de subsistncia e evitar alguns dos perigos da floresta. Um grupo caingangue
residente no Paran, que havia recebido ferramentas de ao apenas na metade do sculo XX, lembrava-se de que no mais tinha de escalar rvores, outrora uma atividade muito frequente, para apanhar larvas e mel. Muitos dos que caam das rvores morriam agora eles simplesmente derrubavam as rvores. O mais importante, pelo menos para a Mata Atlntica, a evidncia de que os tupis no eram conservacionistas no sentido de poupar os recursos naturais para as geraes vindouras. provvel que a ausncia de preocupao no fosse descuido, mas resultasse da certeza razovel sobre a adequao de seus recursos e sua capacidade de defend-los contra os competidores. Infelizmente, seus principais competidores logo seriam os europeus,cujos apetites insaciveis eram muito mais destrutivos que qualquer uma das armas que empunhavam. Os tupis tambm se prontificavam a capturar animais vivos e peles para seus hspedes insaciveis. Alm da madeira corante, o manifesto de carga do Bretoa, em 1511, registra 23 periquitos,dezesseis felinos,dezenove macacos e quinze papagaios; o de 1532, do navio Plrine, 3 mil peles de leopardos etc., trezentos macacos e seiscentos papagaios. difcil imaginar o custo dessas mercadorias exticas para os tupis. Alm disso,fica-se a imaginar qual seria o impacto sobre a Mata Atlntica do extermnio contnuo de milhares de seus maiores predadores. Os europeus gostavam de papagaios, por serem uma novidade e devido a sua plumagem exuberante, alm de sua capacidade de imitar a fala humana. Na verdade, de acordo com um mito,eram transfiguraes dos anjos decados que acompanharam Lcifer em sua queda.Os nativos comungavam desse fascnio: cuidavam das aves e trocavam-nas em funo de suas penas, com as quais se enfeitavam para ocasies sagradas. Ainda que o pau-brasil abastecesse um comrcio muito modesto, comparado aos dos portugueses no oceano ndico, trouxe tal notoriedade recmdescoberta Terra de Vera Cruz a atitude do rei D. Manuel de rebatiz-la de Santa Cruz foi casual que, por toda Europa, ela logo passou a ser chamada Brasil. No nome aplicado aos habitantes da terra era, e continua a ser, brasileiro; da, todos os brasileiros por extenso, parecem estar envolvidos no ofcio, ou negcio, do corte de madeira corante, uma designao peculiar e pungente, agora que as madeiras corantes h muito desapareceram do comrcio e a prpria rvore se tornou to rara em seu hbitat nativo, sobrevivendo, talvez, mais na forma de espcimes plantados patrioticamente em praas e parques pblicos. O trfico de pau-brasil era mais intenso rumo ao norte, ao longo da costa, saindo de Porto Seguro, porque as rvores nativas do sul eram menos apreciadas. A convenincia da baa da Guanabara, apesar disso,estimulava a
derrubadas no local, tal como o faziam a distncia e o isolamento de Cabo Frio, que atraa madeireiros franceses clandestinos.Em 1534, a Coroa, considerando a dificuldade de eliminar do litoral os concorrentes estrangeiros, decretou que todas as rvores de pau-brasil eram propriedade real e que sua derrubada estava sujeita concesso, medidas que continuaram em vigor por mais de trs sculos. Desejoso de acreditar nesses relatos, o rei enviou,em 1531,uma enorme armada sob o comando de Martim Afonso de Souza para estabelecer uma fortificao em algum ponto prximo ao extremo sul da terra portuguesa, o mais prximo dos propalados domnios das riquezas andinas. O destino de Martim Afonso de Souza estava alguns quilmetros ao norte, o porto dos Escravos, que j havia sido um ponto de parada constante de navios que iam e vinham da Europa. Devido a isso,talvez,os tupis chamavam o local de temiuri, sua palavra para provises. Martim Afonso de Souza rebatizou-o de So Vicente, em homenagem ao santo padroeiro de Lisboa.Dentro de poucos anos,fundou-se um segundo porto,chamado Santos,prximo entrada da baa,e instalaram-se duas outras vilas no topo da escarpadura,Santo Antnio da Borda do Campo e So Paulo.Essa apressada extenso de um posto avanado precrio no planalto,aparentemente,foi resultado de penetrar as regies aurferas o mais depressa possvel. Uma segunda cabea de ponte da invaso portuguesa da Mata Atlntica foi estabelecida apenas trs anos depois da de So Vicente. Ficava na baa do Esprito Santo, um pouco abaixo dos 20 de latitude do sul. Em 1534, o rei portugus, impaciente diante do fracasso de seus exploradores em trazer riquezas, decidiu reduzir o custo da colonizao, conferindo direitos de propriedade a alguns de seus cortesos que desejassem assumir as despesas de assentamento e defesa. O Brasil foi fatiado em faixas costeiras paralelas, as capitanias,e foram emitidos ttulos concedendo aos donatrios,como eram intitulados os proprietrios cortesos, poderes que se consideravam suficientes para a tarefa.Foi enviado um governador real em 1549 para estabelecer uma capital em Salvador,na baa de Todos os Santos,fortalecer as defesas da colnia e supervisionar os esforos dos donatrios. A invaso europia do Novo Mundo diferiu da primeira invaso pelo fato de que essa segunda leva j havia adotado a agricultura. Suas fileiras, portanto, contavam com uma srie considervel de espcies domesticadas que,com vantagens,poderiam trazer consigo. Eram capazes tambm, at certo ponto, de avaliar os recursos biticos dos agricultores nativos que encontraram, fosse para comercializ-los ou cultiv-los em sua terra natal.
A outra caracterstica crtica da segunda invaso era que os europeus no estavam meramente migrando para um novo ambiente, mas pretendiam tambm manter contato com o antigo. Desse ponto de vista, a colnia de explorao e a colnia de assentamento que os portugueses, desde o incio, distinguiam como conquista e colnia no eram muito distintas entre si. O intento portugus de conquistar e transformar esse territrio, e nele no sucumbir ou admitir sua rendio s culturas nativas, evidencia-se no seu interesse limitado pela sua vegetao e vida animal. Reconheceram e relataram diferenas grosseiras entre o ambiente tropical e o do conhecido Atlntico Norte. Sua atitude inicial, em relao a essa costa distante, foi fria e at desdenhosa.O encontro de Porto Seguro havia includo trocas de alimentos.Foi o nico momento de estranheza desse interldio idlico,embora tenha sido revelador,pois nenhum dos grupos apreciou as respectivas culinrias. Os invasores portugueses perceberam, contudo, a importncia de efetuar transferncias para o Brasil a partir das regies de floresta tropical do Velho Mundo e estavam, entre todos os europeus, na melhor posio para assim agir. Algumas das espcies domesticadas semitropicais do Velho Mundo que enviaram ao Brasil laranja amarga, limo, gergelim e arroz, por exemplo j haviam sido aclimatadas em Portugal.Mas outras transferncias eram de origem tropical,africana e asitica inhame,banana,coco,gengibre e quiabo,por exemplo que os portugueses j haviam levado para suas colnias de almmar nas ilhas de So Tom,Madeira e Cabo Verde. Com o assentamento em So Vicente, iniciou-se de fato a conquista bitica do Brasil. Embora o reconhecimento anterior tivesse introduzido espontaneamente uns poucos animais e plantas exticos domesticados, Martim Afonso de Souza parece ter deliberado trazer uma variedade de espcies domesticadas europeizadas como experincia daquilo que a terra poderia produzir. No planalto paulista, descobriu-se um habitat frio adequado aos produtos mediterrneos: marmelo,figo,damasco,pssego,pra,tmara e rom;hortalias como repolhos,cebolas,pepinos,meles,colza,endro,rosas (para a gua medicinal de rosas); e essencial prtica da F uvas e trigo. De todos os produtos coloniais isto , aqueles plantados para render um excedente exportvel para a metrpole o mais valioso e vivel era a cana-deacar. Cultivada h sculos na ndia e plantada em todo o Mediterrneo,foi um importante produto de exportao dos portugueses para os mercados do norte da Europa; provavelmente fora trazido para Temiuru ainda antes da expedio de Martim Afonso de Souza. Essas espcies domesticadas, trazidas para o Brasil, foram plantadas em campos queimados, empregando-se quase as mesmas tcnicas dos tupis e
valendo-se da surpreendente fertilidade inicial da biomassa florestal reduzida a cinzas. Para a Mata Atlntica, os perigos eram imensos, porque uma sociedade baseada na mo-de-obra compulsria no levava em conta o ambiente. Os que comandavam o trfico eram pessoas em trnsito e imprevidentes: De quantos l vieram,nenhum tem amor a esta terra.[...] porque todos querem fazer em seu proveito,ainda que seja custa da terra ,porque esperam de se r ir.Os plantadores de cana no viam na floresta nada alm de um obstculo realizao de suas ambies. Aqueles que cada vez mais vinham ocupar as margens da economia de plantao posseiros do serto eram impelidos a explorar a floresta de maneira descuidada e imprevidente. A conservao dos recursos naturais iria mostrar-se irrelevante em uma sociedade na qual a conservao da vida humana era irrelevante. Era propsito dos jesutas converterem os indgenas no apenas ao cristianismo, mas passividade e dependncia caractersticas de um campesinato portugus que aceitasse a tutela perptua e todas as indignidades que a caracterizavam chicoteamento, prstimos pessoais, submisso sexual e trabalho nos campos. Para alcanar essa meta, impunha-se pr um fim explorao itinerante e extensiva da floresta pelos indgenas. Os missionrios ou seus agentes empreendiam ento as entradas a partir de So Paulo,Rio de Janeiro e Esprito Santo.Eram numerosas e paralelas aos saltosdos caadores de escravos,abrangendo quase toda a regio da Mata Atlntica,e juntavam at dois ou trs mil indgenas de uma vez. Consta que uma ofensiva de seis anos sobre o planalto paulista, iniciada em 1590 sob o comando de Jernimo Leilo, destruiu trezentas aldeias, matando ou capturando seus habitantes, que chegavam a 30 mil. Talvez seja um relato exagerado uma densidade de 0,4 habitante por quilmetro quadrado nas regies montanhosas implicaria o desmatamento de 75 mil km -, embora cifra no muito inferior seja totalmente possvel, porque as entradas muitas vezes retornavam aps seis meses na floresta com mil ou 2 mil cativos enfileirados. Embora o trfico escravo e as agruras das plantaes trouxessem a morte aos tupis e seus vizinhos, sua destruio pelas doenas infecciosas do Velho Mundo foi ainda maior. Na regio da Mata Atlntica, os saltosdos traficantes de escravos e das entradas dos jesutas intensificaram a exposio da populao nativa s infeces. As aldeias atuavam como foco de surtos epidmicos.Em 1576,por exemplo,os padres no Esprito Santo relataram que 75 dos 160 nativos que eles haviam atrado para sua aldeia haviam deixado este mundo para viver no outro,o feliz.
Ao longo da costa, de So Vicente e Cabo Frio, onda aps onda de doenas devastou os tupis; em 1600, estavam reduzidos a uns quatro ou cinco mil, um declnio assustador de 95% em um sculo. bem possvel que a floresta do planalto tenha passado por escala similar de despovoao. Antes da iniciativa da apropriao, o monarca portugus aplicou nova colnia seu conceito de propriedade da natureza. No ato de dividir o Brasil em capitanias em 1532, a Coroa se presumia como a possuidora legtima de tudo sobre o continente sul-americano a leste da linha de demarcao, por direito de conquista,ainda que seu controle efetivo na poca se estendesse a um nico enclave, com um raio de uns poucos quilmetros, em So Vicente.Estava implcito neste conceito todo o mundo bitico que a habitava e o substrato do qual ela dependia.Porque reservava para si,alm das rvores de pau-brasil,a propriedade das minas de metais e pedras preciosas,e submetia a julgamento especial pleitos privados quanto a cursos d gua e stios ribeirinhos para a instalao de moinhos d gua,utilizados para moer cana. Os tupis consideravam as florestas como pertencentes aos espritos e animais que as habitavam, ou pelo menos como pertencentes tanto queles seres como a si mesmos, ainda que em algum nvel suas guerras fossem certamente um esforo pela posse exclusiva dos recursos naturais. Em 1600, o relacionamento humano com Mata Atlntica tinha se transformado, principalmente porque a maioria de seus habitantes humanos originais havia desaparecido e porque o nmero de invasores era insuficiente para substitulos. Ao longo da costa,aps um sculo de constantes guerras,escravizao e epidemias,apenas sete dos donatrios haviam conseguido instalar algum tipo de assentamento. Diversos assentamentos j estavam decadentes. Em conjunto, nas capitanias do Sudeste apenas quinze engenhos estavam operando: Santos e Rio de Janeiro,com seis engenhos cada um, e o Esprito Santo, com trs. A populao do Brasil, sob controle portugus, em 1600, pode ter chegado a menos de 65 mil. Menos de 10 mil desses residentes eram europeus ou mestios. Quanto populao nativa sobrevivente, que habitava a rea da Mata Atlntica fora dos limites portugueses, pode-se apenas especular; possvel que tivesse praticamente desaparecido em um raio de trezentos quilmetros das vilas litorneas,em decorrncia das entradas e saltos,da disseminao de doenas infecciosas e da fuga de sua populao para reas mais interioranas. Um sculo depois de os portugueses terem inicialmente reivindicado seu direito, o controle da costa ainda no estava garantido, mesmo aps a expulso dos franceses do Rio de Janeiro. A luta pelo litoral norte, estendendose at o Amazonas, havia apenas comeado,e a usurpao portuguesa ao sul de Canania enredaria a colnia na verdade, os estados sucessores
independentes em guerras intermitentes por mais de um sculo e meio. As violentas rivalidades coloniais do primeiro sculo foram, portanto, apenas um aperitivo das crescentes ameaas ao reino da natureza. O declnio catastrfico da populao nativa que se seguiu invaso europia do sculo XVI e a insignificncia demogrfica da imigrao portuguesa concederam Mata Atlntica um alvio, aps 10 mil anos de caa e coleta e mil anos de lavoura itinerante. A floresta que estava afastada de portos adequados ou carente de pau-brasil foi deixada em paz. Teve incio um perodo de recomposio da vegetao e da fauna. Isto pode ser verificado em alguns lugares. A vila de So Paulo,fundada em uma colina no meio de uma plancie ampla,aberta,em 1587 possua mais bosques que outra coisa,e os citadinos receberam ordens para cortar as rvores no interior e ao longo de suas paliadas. A Mata Atlntica ainda era quase totalmente desconhecida ou jamais observada. Uma imensa realidade natural, inteiramente vista dos invasores,estava ainda, h mais de um sculo de distncia de ser descoberta. Cap. 4 Alheamento: Despovoada, A Floresta Renasce. A Mata Atlntica, tendo a sua populao drasticamente reduzida em grande parte dos seus domnios, passou a uma fase em que diminuram as necessidades e os anseios humanos. No curso do sculo XVII, porm, foi se tornando tambm uma realidade cada vez mais afastada do entendimento humano.Um ecossistema pode ser visto como um reservatrio de informaes,as geneticamente programadas e, ao mesmo tempo,as acumuladas por suas espcies,relevantes a sua sobrevivncia e reproduo em seu interior.Os homens da Mata Atlntica,como todas as suas outras criaturas,haviam armazenado,durante 12 mil anos,seus prprios estoques de informao.Uma vez que os recursos e experincias de cada aldeia diferiam dos de seus vizinhos,milhares de espcies da Mata Atlntica tinham sido catalogadas na memria de seus habitantes humanos. A primeira ou as duas primeiras geraes de invasores portugueses haviam dependido totalmente dos conhecimentos indgenas sobre a Mata Atlntica. Os tupis caavam, plantavam e cozinhavam para os expedicionrios, alm de curar suas doenas. A gerao mestia gerada pelos degredados era culturalmente indgena, falante do tupi, conhecedora da floresta e dos meios de explor-la. Os filhos da primeira gerao de unies entre portugueses e tupis no aceitavam o status de camponeses e muito menos o de escravos. Resistiam ao batismo e eram indiferentes ao estigma de ilegitimidade. Andavam nus e no hesitavam em insultar os padres quando se irritavam com suas exigncias.
Os habitantes portugueses haviam passado a ser quase como os ndios, porque,sendo cristos,viviam moda de gentios.O resultado foi uma queda maior na autoridade moral da igreja:seu esforo em impor a monogamia sociedade portuguesa apenas havia sido bem-sucedido at o ponto em que as relaes abertas eram passveis de punio por tribunais leigos. Mas a Mata Atlntica aceitava qualquer portugus que fosse intrpido o bastante para pular a cerca e ignorar as injunes da igreja no tocante a esta e a muitas outras questes. No demorou muito para que os brancos casados com ndias e sua prole mestia fossem arrebanhados e submetidos a uma empresa colonial que tinha pouco uso para o seu conhecimento da Mata Atlntica ou para sua ao potencial na integrao de tcnicas europias e indgenas. Rio de Janeiro e So Paulo continuaram a receber fluxos bem pequenos mas significativos de migrantes.Entre os migrantes,havia umas poucas mulheres,deliberadamente importadas de forma a evitar a absoro gentica dos brancos.Tornou-se possvel, ento, uma sociedade de castas, na medida em que os homens brancos mais ambiciosos e dominantes,astutos o bastante para se darem ao trabalho de granjear favores aos olhos das autoridades portuguesas, buscavam parceiras brancas para o casamento. Entre as medidas institudas pela Coroa, para delinear uma sociedade de castas, estavam aquelas que segregavam residncias neoeuropeias e nativas e definia a cidade como domiclio prprio de brancos.Em 1583,o conselho municipal de So Paulo proibiu que os brancos visitassem as aldeias para beber e danar segundo seu costume. Esta foi uma primeira tentativa, na Mata Atlntica, de definir a cidade como sinnimo de civilizao, dotando-a dos instrumentos de comando e relegando o que era nativo e brbaro,e devidamente reprimido, s aldeias. Os habitantes das aldeias estavam sujeitos ao tributo de trabalho, aplicado at o ponto de esgot-los. Os mais novos desses recrutados para a civilizao continuavam a sofrer a calamidade de epidemias peridicas. Em So Paulo, a aldeia era principalmente um meio de manter mo-de-obra temporria ao alcance da mo;no Rio de Janeiro e Esprito Santo,onde os escravos africanos eram mais facilmente obtidos para tal fim, as aldeias, no mais das vezes,serviam para estender a fronteira. Outra medida eficaz no estabelecimento de uma sociedade de castas foi o modo pelo qual as sesmarias eram concedidas. A abundncia de terra desocupada, em consequncia dos assaltos e epidemias, possibilitava a qualquer pessoa livre ocupar uma terra devoluta. Em Portugal, a poltica real de concesso de terras fora concebida para estimular a produo camponesa necessria ao abastecimento das vilas; no
Brasil, curvou-se ao propsito de encorajar a busca de ouro e pedras preciosas ou, no caso de no haver descobertas, a produo e exportao de acar.Surgia,assim,uma classe dominante para quem a propriedade da terra era uma forma de privilgio,conquistado por meio da prepotncia local e confirmado pela conivncia real. As primeiras concesses de terra foram modestas, compatveis com uma escala europia de propriedade, talvez devido limitao do domnio colonial e dificuldade de defend-lo das levas de indgenas empenhados em restabelecer suas terras. No obstante,cesses iniciais de cerca de cem hectares logo deram lugar a doaes muito maiores,caracterizando o que em Portugal seriam consideradas vastas propriedades.Tais donatrios eram homens com numerosos escravos e uma enorme ambio a que a Coroa estava disposta, ou era obrigada, a tentar satisfazer. De vez em quando, havia concesses ainda maiores e os mais ricos e poderosos dos notveis locais brigavam com os donatrios para adquirir propriedades do tamanho de principados.O mais arrogante de todos os notveis que procuraram se apossarda Mata Atlntica, contudo, foi Salvador Correia de S,cujo tio havia capitaneado o extermnio da colnia francesa na baa da Guanabara. Designado governador, coroou uma srie de faanhas militares e comerciais com o confisco, em nome do rei, de um grupo de sesmarias sucessoras da fracassada capitania de So Tom, que ele conferiu, em 1652, a sim mesmo e a seus filhos. Infelizmente, para a tranqilidade da colnia e seu governo, concesses de terra eram normalmente atribudas em condies de extrema impreciso e confuso.O rei, os donatrios, governadores e os conselhos municipais no tinham o menor empenho em coordenar suas doaes, de sorte que o mesmo territrio poderia muito bem possuir quatro donos aparentemente legais.As sesmarias nunca especificavam claramente os limites e, s vezes, nem mesmo as localizaes.Embora as concesses contivessem clusulas, prevendo demarcao a ser efetuada pelo donatrio,elas, com frequncia, eram omitidas ou adulteradas por fraude e incompetncia. Um dos principais motivos para o tamanho espantoso das sesmarias era a tcnica adotada para a explorao da floresta e seus solos.Adaptada do regime de derrubada e queimada praticado pelos indgenas,inclua a aceitao do milho e da mandioca como culturas de primeira necessidade.Uma vez que os solos recm-queimados da floresta eram imensamente frteis e o regime eliminava a necessidade de cultivar e, por isso,o arado foi abandonado.Com isso, a economia de mo-de-obra era enorme,verdadeira liberao para o colono portugus que no possua escravos. Os invasores viabilizaram a lavoura mais intensiva, pelo menos no curto prazo, introduzindo, desde o princpio, animais domsticos do domnio bitico
eurasitico. Porcos, galinhas, ovelhas, cabras e, especialmente, gado bovino propiciaram suprimento de carne com reduzido acrscimo de mo-de-obra.Os porcos,contudo,eram preferidos pelos pequenos agricultores por serem fceis de criar e conduzir aos mercados das vilas. O gado bovino foi introduzido inicialmente em So Vicente, talvez com a expedio de Martim Afonso de Souza em 1532,trazido das ilhas do Cabo Verde,onde a variedade portuguesa,criada solta e pesando de 200 a 300 quilmetros, j se aclimatara.Parte desse gado galgou o paredo costeiro at So Paulo,parte foi levada de navio para o Rio de Janeiro. A insero de gado bovino no regime de derrubada e queimada foi, no entanto, problemtica. Os agricultores que o praticavam no construam cercas, mas suas clareiras eram efmeras demais. Na prtica nativa, as tentadoras faixas de mandioca atraam caa selvagem, mas os animais domsticos eram intrusos incmodos. Os pastos se formavam logo depois dos limites das vilas. Concedeu-se uma plancie aos jesutas, chamada Santa Cruz,a oeste do Rio de Janeiro. Drenaram o terreno e colocaram umas 20 mil cabeas de gado, em um nico e amplo pasto. A pecuria bovina na regio do Rio de Janeiro expandiu-se para leste, ao longo da costa, onde se encontrariam faixas de campos gramados at a foz do rio Paraba do Sul. No Sudeste, raramente se utilizavam cavalos e animais de carga nos primeiros dois sculos de colonizao. As expedies em busca de escravos e o transporte de mercadorias, mesmo de mercadorias de pouco valor por unidade de peso, eram realizados a p, onde fosse impraticvel o transporte fluvial.Os cavalos eram encontrados principalmente nas vilas; a primeira meno a jumentos data somente de 1635 .O desperdcio de energia humana era extraordinrio, quase incompreensvel, principalmente considerando-se o pequeno esforo exigido pelo cultivo ou criao de animais. Enquanto o interior da floresta permaneceu em grande parte intacto e as clareiras relvadas eram dispersas demais e pouco conhecidas, continuaram impraticveis as viagens longas a cavalo. Os aldees teriam achado difcil empregar jumentos na presena das clareiras no cercadas caractersticas do regime de derrubada e queimada. A introduo de implementos de ferro possibilitou outro tipo de intensificao do regime de derrubada e queimada. O machado tornava bem mais fcil derrubar floresta primria, e no capoeira, porque reduzia em muito o trabalho de cortar rvores. possvel que a enxada tenha permitido aumentar as faixas cultivadas de um para 2,5 ou cinco hectares por trabalhador.O uso da enxada significou que a invaso inevitvel de ervas daninhas no provocava necessariamente o imediato abandono de um campo.Significava tambm que a mata secundria no seria to desconsiderada devido a sua provvel
infestao por sementes de capim.Um simples utenslio, portanto, a enxada, era potencialmente capaz de eliminar para sempre a Mata Atlntica. A rpida expanso da rea ocupada por neoeuropeus, nos anos de 1600, sugere uma dilapidao impiedosa de recursos florestais. Seus assentamentos estendiam-se por 250 quilmetros ao longo dos cales dos rios Tiet e Paraba do Sul,uma expanso que evidencia o acerto do cronista Gabriel Soares de Souza ao observar,neste caso com bvia aprovao,que tratando em suma da fertilidade da terra,digo que acontece muitas vezes valer mais a novidade de uma fazenda que a propriedade, pelo que os homens se mantm, honradamente, com pouco cabedal. O tamanho habitual da sesmaria sugere uma medida da densidade demogrfica e do ritmo de derrubada da floresta. Cada sesmaria era a morada do donatrio e de um modesto squito, que consistia de seus parentes, dependentes livres e escravos. A consolidao da colnia portuguesa ocorreu durante um perodo de extrema incerteza relativo independncia da metrpole e sobrevivncia de seu imprio. O trono portugus ficou vago em 1578 e foi ocupado, dois anos depois por Felipe II, e tiveram acesso ao comrcio brasileiro do acar, em grande parte financiado por eles mesmos. Mesmo enquanto as coroas de Portugal e Espanha se achavam unidas, mantinha-se uma administrao independente nas colnias portuguesas,de sorte que a linha do Tratado de Tordesilhas continuava a dividir o continente sul-americano. Ao longo da costa, em Recife, Salvador, Esprito Santo e Rio de Janeiro, a cana-de-acar, introduzida quase um sculo antes, ainda no havia sofrido a ao de nenhum dos parasitas ou pestes que co-evoluram com ela no Velho Mundo, exceto por umas poucas ervas daninhas, ratos e outros animais domsticos bovinos, caprinos e sunos. O acar tornou-se a nica atividade econmica de grande vulto que vinculava a regio da Mata Atlntica metrpole, afora o corte do pau-brasil. Pode-se calcular que at 1700 cerca de 150 anos, aps a exportao do acar, alcanar escala comercial os campos de cana teriam eliminado uns mil km da Mata Atlntica, supondo-se um crescimento quase constante e os campos de canacansadossendo abandonados para a agricultura de subsistncia ou pastagens aps uma mdia de quinze anos.A produo de acar tambm consumia floresta na forma de lenha, queimada debaixo dos tonis nos quais o caldo da cana era cristalizado.Cerca de quinze quilos de lenha eram queimados para cada quilo de acar produzido,o que daria a mdia de 210 mil toneladas de matas secundrias e florestas de manguezais de enseada cortadas anualmente para esse fim. Alm disso, as moendas
precisavam de cinza de madeira para purgar o acar de suas impurezas; os manguezais das baas eram os preferidos para esse material. Calculando-se duzentas toneladas de lenha por hectare, as moendas teriam consumido mais 1200 km no curso de 150 anos. O trigo no conseguia amadurecer nas baixadas mais, introduzido no plat semitropical de So Paulo, em dado momento aps 1609, ali floresceu. A nostalgia portuguesa por seu cereal bsico sacralizado na cerimnia da missa , e a incapacidade de Portugal produzir um excedente, geraram um mercado considervel, que se estendia at o nordeste brasileiro. O cultivo da cana-de-acar e do trigo e a criao de gado eram empreendidos com o trabalho escravo. No Sudeste, os escravos na maioria eram nativos, obtidos atravs de uma expanso das caadas de escravos por paulistas.Depois de 1600, com a maioria dos agricultores nativos da regio entre os rios Grande e Paranapanema mortos ou afugentados, os paulistas voltaram sua ateno mais para o sul e sudoeste, para a regio ocupada pelos atuais estados do Paran e Santa Catarina. Na extremidade sudoeste da Mata Atlntica,numa regio chamada Guair, ao longo do rio Paran, concentravamse populaes realmente densas de guaranis e outras tribos. Contra as aldeias de Guair, os paulistas organizaram ondas de assaltos, ou entradas. Suas tropas formavam companhias militares chamadas bandeiras, donde o neologismo patrioteiro bandeirantes, que sugere porta-bandeiras ou pioneiros.Cada expedio era composta de cem ou mais brancos e mamelucos e at mil auxiliares nativos.Seus lderes eram especialistas no trfico de escravos excelentes batedores e caadores que falavam tupi e possuram destreza nas armas e preservavam parte de sua herana indgena para fins menores.Os caadores marchavam por terra ou navegavam descendo o rio Paranapanema ou o Tiet. Cada jornada durava meses e tambm cobrava seu tributo dos prprios caadores. Em 1607,um bandeirante lamentava com petulncia os 240 soldados nativos que havia gasto na floresta na busca de mais nativos para escravizar uma taxa de mortalidade de trs por dia.Em 1640,de uma populao original que pode ter chegado a 150 mil, toda essa crueldade havia capturado talvez uns 60 mil, dos quais cerca da metade pode ter sido vendida para os fazendeiros do acar do Rio de Janeiro. Em So Paulo, muitos dos cativos, expostos a doenas contagiosas, logo adoeciam e morriam.Assim,o retorno de cada bandeira era acompanhado de mais uma epidemia.O despovoamento de Guair foi seguido de caadas ainda mais para oeste, na rea da misso de Itatim, nas cabeceiras do rio Paraguai, e para o sul, contra as misses da regio dos tapes, no atual Rio Grande do Sul.Os indgenas da tribo dos tapes, contudo, estavam armados, e os paulistas, por causa das dificuldades criadas pelo excessivo alongamento de suas linhas de suprimento, foram ali derrotados em 1641.No Sudeste, nesta poca, os
assaltos se limitavam captura de bandos de caadores-coletores, de propores modestas, sem dvida, j em fuga de zona mais litorneas. Mesmo para os contemporneos, parecia bizarro que o rei espanhol permitisse aos colonos brasileiros exterminaram enormes contingentes de nativos, que ele supostamente devia considerar tambm como vassalos, em troca do resultado to passageiro e insignificante de alguns milhares de toneladas de acar. Peties indignadas foram dirigidas Coroa para que, em vez disso, empregasse esses guerreiros na campanha contra os invasores holandeses.De fato, foram recrutados para esse fim,mas no atenderam convocao. Os brancos que no possuam escravos acabavam se casando com nativas e mamelucas e, pouco a pouco, formavam uma massa de mestios deserdados, subordinados aqueles que manejavam o poder social, e vivendo a favor termo regional ironicamente oportuno. Na hierarquia social, esses mestios ocupavam uma posio acima da dos escravos africanos,que logo comearam a tomar seu lugar;seu status de pessoas livres era respeitado, enfim, devido presena desse novo estrato despossudo ao extremo. A introduo de africanos na regio da Mata Atlntica comeou nos anos de 1550. Eram preferidos aos indgenas para o trabalho na plantao devido maior expectativa de vida e ao hbito de trabalho no campo dos homens africanos. Enquanto povos sequestrados e deslocados,era presumvel que fossem menos inclinados a fugir, porque no tm para onde ir. Apenas na metade do segundo sculo de colonizao no Brasil foi que as florestas brasileiras,totalmente exticas e imponentes,tornaram-se um hobby menor dos jesutas, agora em condies de consider-las isoladas de seus temveis, mas em grande parte exterminados,habitantes humanos.Simo de Vasconcellos,que morou na Bahia,em sua crnica escrita em 1663,objetivou mostrar que o Brasil no era inferior aos outros trs quartos do mundo, uma ideia que havia rancorosamente grassado entre os europeus depois de seu entusiasmo inicial por aquilo que haviam fantasiado como um paraso terrestre. Eles haviam se equivocado, por exemplo, ao supor que os climas iam ficando mais secos quanto mais prximo do equador. O Brasil era vioso e desfrutava de influncias favorveis do Sol,da Lua das estrelas,uma abundncia de peixes em suas guas e pssaros em seus cus,e todo tipo de fera terrestre,tal como havia afianado o primeiro captulo do Gnese.Como qualquer outro cronista,Vasconcellos regozijava-se com a eterna primavera do Brasil, que recria os olhos e convida as almas a louvar o Autor da natureza,porque sem dvida excede nessa formosura todas as outras partes do orbe. A descrio da floresta feita por Vasconcellos,no entanto,ocupa apenas umas poucas pginas de seu massudo livro, cujo objetivo principal era descrever a
misso heroica de sua ordem de arrancar o Brasil do poder do inferno, sob cujo domnio havia estado desde o incio do mundo, h uns6 mil anos. Embora o interesse cientfico portugus em seus territrios do Novo Mundo fosse pequeno, a botnica econmica mereceu um pouco mais de ateno em consequncia da perda das colnias asiticas. A sociedade que habitava a Mata Atlntica, apesar de toda destruio e desordem operados pela invaso europia, ainda era, em grande parte, indgena no final do sculo XVII, tanto em termos culturais como genticos. No lado indgena de uma fronteira osmtica, as adoes da cultura europeia foram gradativas no totalmente por uma questo de escolha,aldeias, ou que tinham sido escravos nas lavouras de trigo ,s vezes fugiam e at em massa. Entrementes, elementos culturais nativos,embora reprimidos e desprezados pelos neoeuropeus dominantes, persistiram e invadiram o domnio neoeuropeu.Mesmo aps dois sculos de ocupao, por toda parte os brancos ainda constituam minoria,exceto nas cidades do Recife,Salvador e Rio de Janeiro.Os portugueses atribuam, por analogia,os nomes de plantas e animais europeus familiares a algumas das espcies da Mata Atlntica, mas o resto foi recebido dos indgenas.Dois teros dos nomes comuns das rvores da Mata Atlntica e praticamente de todos os seus animais so de origem tupi-naturais geolgicas, edficas,vegetais e de acidentes geogrficos rios,montanhas,vales e esturios. No fim de um segundo sculo da invaso portuguesa, a empresa colonial instalada na borda da Mata Atlntica havia se intensificado muito. O assalto genocida a povos tribais ampliara-se por uma rea muito maior e ampla faixa de floresta havia sido deixada quase sem ocupantes humanos. Os sobreviventes dessas culturas da floresta serviam agora como subclasse cativa.Os homens,no mbito do setor de controle portugus da Mata Atlntica, podem ter chegado a 300 mil,dos quais talvez um tero era de origem portuguesa.Cerca de 20 mil, possivelmente, moravam em vilas. A densidade rural continuava a ser muito baixa, talvez cinco pessoas por quilmetro quadrado no litoral, e duas por quilmetro quadrado no planalto paulista,numa rea total ocupada por neoeuropeus e seus subordinados de cerca de 65 mil km.A maior parte dessa rea fora subtrada Mata Atlntica, embora parte dela tivesse sido reiteradamente derrubada e queimada pelos agricultores tupis. Mais de um sculo e meio depois da invaso portuguesa, a geografia do interior da colnia ainda era mtica, seus grandes rios concebidos como oriundos de um nico grande lago, em algum ponto nos sops dos Andes. As autoridades coloniais estavam desesperadas para localizar ouro, que salvaria a Coroa Portuguesa dos efeitos desastrosos da perda das colnias asiticas e da perda de grande parte do mercado do acar para o Caribe. Por fim,em 1690,
uma das bandeiras descobriu ouro. Depsitos de aluvio foram encontrados longe, no interior, ao longo da serra do Espinhao, abaixo de um monte que chamaram Itacolomy. A Mata Atlntica ia agora passar por uma invaso ampla e permanente. A maior corrida do ouro da histria estava prestes a comear. Cap. 5 Ouro E Diamantes, Formigas E Gado. As grandes descobertas de ouro e diamante, no Brasil do sculo XVIII, foram as mais importantes ocorridas no Novo Mundo colonial. De 1700 a 1800, 01 milho de quilos de ouro foram oficialmente registrados e talvez outro milho tenha escapado ao fisco real.Cerca de 2,4 milhes de quilates de diamantes foram extrados,segundo registros oficiais e uma quantia adicional desconhecida e incalculvel foi contrabandeada.A populao e os recursos locais fluram para as reas aurferas,que se situavam em pleno serto,ao longo da fronteira interna da Mata Atlntica.Com isso,a colnia passou a depender mais dos cativos africanos e estes,com seus descendentes, passaram a ter participao preponderante na tarefa de interpretar e destruir a Mata Atlntica. Com a descoberta do ouro, o Brasil se tornou muito mais estratgico para Lisboa,como se depreende da deciso de 1720 que o elevou do status de governadoria-real para o de vice-reino. Era inevitvel que uma populao mais numerosa, uma atividade econmica mais intensa,o aumento da vigilncia governamental e a rivalidade europia afetassem a integridade das florestas brasileiras.Quando os garimpeiros finalmente perderam a esperana e adotaram a lavoura e a pecuria, contudo, estenderam seus domnios cada vez mais para o interior da floresta ainda intocada. O sculo XVIII, ento, representou o incio de uma tendncia irreversvel e cumulativa na explorao da Mata Atlntica. Por todo o sculo XVII, houve lavras de ouro espalhadas por diversos pontos ao longo da serra do Mar. Nas lavras, todas de aluvio,o ouro a princpio era facilmente encontrado em leitos de crregos.A onipresena de depsitos pequenos desse tipo, ao longo das escarpas e da face interna da serra do Mar, deu origem a um tipo especial de penetrao de fronteira, feita por exploradores chamados faiscadores ou garimpeiros. A necessidade de auto-suficincia deve ter-lhes reavivado a curiosidade sobre os recursos da floresta, especialmente produtos alimentcios e medicinais. Apesar disso,a garimpagem, que se caracteriza por se concentrar em um nico recurso, no-renovvel e abitico, no era uma ocupao que tendesse a ampliar a apreciao da natureza.
Em poucos anos, diversos riachos muito ricos foram encontrados e em 1713 todos os principais achados j estavam produzindo. Estavam localizados em um arco ao longo da serra do Espinhao,desde a atual cidade de Belo Horizonte at So Joo Del Rei.Outros achados menores dispersos foram feitos em Pernambuco,Sergipe,Bahia e Esprito Santo. A data da primeira descoberta de diamantes incerta. Esses depsitos,encontrados mais para o norte ao longo da mesma serra, concentrados em uma vila hoje chamada Diamantina, foram levados considerao do rei apenas em 1729, poca na qual diversas fortunas privadas provavelmente j se haviam constitudo. No obstante, as autoridades coloniais mantinham-se firmes. Foram institudos governos nas vilas; a minerao ficou mais estreitamente limitada a concesses de arrendamento feitas pela Coroa; e a regio de ouro e diamante, hoje conhecida como Minas Gerais, foi separada da capitania de So Paulo e seu governo sediado em Vila Rica de Ouro Preto. Patrulhas militares, embora no muito eficazes na interdio das operaes de garimpeiros ou pequenos traficantes ou da sada de contrabando, ainda assim bloqueavam a penetrao de novos assentamentos mais a leste da serra do Espinhao para dentro da Mata Atlntica. Foram africanos da Coroa do Ouro que ensinaram a seus proprietrios a batear e se mostraram peritos em localizar minas. Esses trabalhadores alcanavam preos mais elevados porque, acreditava-se, eram especialmente afortunados ou praticantes de uma espcie de bruxaria para encontrar ouro.O ouro tambm era encontrado em sops de montanhas, em certos tipos de argila a um metro ou dois de profundidade e em formaes rochosas friveis. Nesse tipo de terreno,a floresta era queimada grandes parcelas dela,suficientes,em certos lugares,para erradicar a lavoura itinerante s vezes,os riachos eram voltados contra as encostas, tcnica chamada carregar uma montanha em talho aberto, para poder lavar cada torro de solo que pudesse conter ouro ou provocar o desabamento de uma aflorao rochosa promissora. Evidentemente, essa prtica hidrulica perversa exigia enormes quantidades de gua fluindo livremente, de sorte que, no geral, foram tentadas mais imediatamente na face leste da serra do Espinhao, isto , a face florestada do planalto, a barlavento, onde as chuvas eram mais pesadas e os riachos, mais largos, fluam o ano todo. Um sbio local, Jos Vieira Couto,observando tais depredaes em 1799,considerou o termo minas totalmente inadequado: preferia usar o termo local, lavras.Curiosamente, a mesma palavra era aplicada na agricultura, lavrar e lavoura, uma atividade quase igualmente predadora em seus mtodos e concepo.O prprio conceito de trabalho assumia, assim, mais o carter de destruio que o de produo,construo ou conservao.
O efeito desse tipo de minerao foi o de substituir a floresta por charnecas esburacadas. Por todos os lados, tnhamos, sob os olhos, os vestgios aflitivos das lavagens, vastas extenses de terra revolvida e montes de cascalho , relatava o botnico francs Auguste de Saint-Hilaire quando atravessou a estrada ao norte de Ouro Preto, na segunda dcada do sculo XIX. Pode-se estimar aproximadamente a rea devastada pela prospeco de ouro e diamante. Modernos levantamentos na regio sugerem que os mineiros obtinham talvez um grama de ouro a partir de um metro cbico de material contendo ouro e ganga e que esta camada tinha, em mdia,cinquenta centmetros de profundidade. Dessa forma,o volume total de ouro obtido durante o sculo XVII teria revirado 4 mil km da regio da Mata Atlntica. As lavras nessa zona teriam se concentrado ao longo dos leitos dos riachos, causando dano, portanto, e principalmente, s associaes de plantas hidrfilas isto , que buscam umidade. Ao longo dos taludes internos,teriam sido florestas de galeria, em certos lugares, a nica cobertura arbrea local e, em outros, a nica cobertura no-caduca disponvel, por isso uma importante defesa contra a eroso da terra e o assoreamento dos rios. A descoberta de ouro e diamantes aumentou muito a populao neo-afroeuropeia da Mata Atlntica. Em 1800, totalizava cerca de 1800 000 indivduos, tendo se multiplicado seis vezes durante o sculo, um crescimento anual de cerca de 1,8%. Na regio de minerao,metade dos habitantes eram escravos africanos, e mais da metade do restante eram pessoas de origem africana ou mestias de africanos. Entrementes, a rea no limites neoeuropeus no cresceu na mesma proporo.Os neo-afro-europeus podem ter ocupado na poca 90 mil km do segmento sudeste da Mata Atlntica, de Paranagu at Caravelas. No curso do sculo XVIII, as necessidades da populao da regio mineradora em termos de gneros alimentcios, quase totalmente satisfeitas pela lavoura em regime de derrubada e queimada, teriam exigido a destruio mdia de seiscentos quilmetros quadrados de floresta por ano. Grande parte dessa queimada se repetiria na floresta secundria,acessvel s vilas e lavoura itinerante cada vez menos vivel, de sorte que as zonas densamente povoadas passaram a ser abastecidas a partir de distncias considerveis. A itinerncia da lavoura em regime de derrubada e queimada, outrora desespero dos missionrios, tornava-se agora alvo das autoridades civis, que nela viam o principal obstculo coleta de impostos, ao aumento do abastecimento alimentar urbano e, de fato, imposio da autoridade e da ordem social. Em virtude disso, um dos principais objetivos do governador designado para a capitania de So Paulo em 1765, Lus Antnio de Souza Botelho, era impor vida alde a seus habitantes que vagavam em clamorosa falta de fiscalizao.
O governador procurou converter os fazendeiros paulistas ao emprego do arado, uma reforma que incluiria integrar animais de trao e seu estrume ao cultivo dos campos. Os habitantes lhe ofereceram, contudo, a opinio universal de que os solos de So Paulo no eram adequados a arao. A presso sobre a Mata Atlntica teria, em grande parte, diminudo se essa reforma tivesse sido bem-sucedida. Pode-se imaginar, porm, a incredulidade dos horticultores mestios quando lhes era dito que deviam trocar um mtodo de cultivo, que interrompia seu lazer por no mais de quinhentas horas por ano, por outro que os teria sobrecarregado com pelo menos 2 mil horas de labuta e que, alm disso, no era comprovado e talvez no fosse capaz de render colheitas maiores. O governador percebia que a poltica fundiria colonial havia contribudo para a preferncia pelas derrubadas e queimadas. O custo excessivo dos ttulos de terra deixava-os fora do alcance dos lavradores simples e os concentrava nas mos dos diletos da corte. Souza Botelho entendia bem que o mercantilismo e o militarismo portugueses contrariavam sua poltica agrcola. Sua proposta de autorizar fbricas txteis que teriam absorvido o algodo local foi rejeitada por Lisboa.Em vez disso,foi instrudo a arregimentar os parcos recursos da capitania para a guerra no sul.Ele e seus sucessores tributaram vorazmente os pequenos fazendeiros, exigiram mo-de-obra para construir fortes, confiscaram colheitas para alimentar as tropas e apoderaram-se de suas casas para alojar soldados.Seus coletores de impostos exigiam pagamentos em dinheiro,embora seus tesoureiros atrasassem os pagamentos durante anos. Aqueles que fugiam s extorses das autoridades necessariamente se afastavam de contatos com as vilas. Adentravam uma floresta extremamente insegura,para a qual escapavam tambm os fugitivos da justia criminal e para onde outros criminosos haviam sido banidos, continuao de uma prtica judicial que convertia a fronteira em uma espcie de depsito de degredados. Muitos dos expulsos, contudo, eram apenas os desempregados das vilas.O governador de Minas Gerais recrutava compulsoriamente para a milcia homens acusados de vadiagem e os enviava a postos na fronteira, julgando mais razovel que a tropa seja composta de vadios e patifes que de homens trabalhadores, necessrios ao cultivo da terra. Os sertanejos entravam na Mata Atlntica, que se tornara mais luxuriante nos dois sculos em que a populao humana em seu interior havia praticamente desaparecido. Ao longo da fronteira oriental da zona de minerao de ouro at o litoral, uma distncia de mais de duzentos quilmetros,e a nordeste do distrito diamantino, outros quatrocentos quilmetros descendo o rio Jequitinhonha,os agricultores de subsistncia podiam devastar floresta virgem para formar seus campos de milho e mandioca.Todos esses sertanejos, racialmente mesclados,
estavam formando uma cultura distinta das vilas sob controle portugus. A populao das vilas os chamava, desdenhosamente, de caboclos ou caipiras.Expressivamente, so termos tupis pejorativos, com os seguintes e respectivos significados mais provveis: moradores em casa de um homem branco e lenhadores da floresta. A fronteira florestal tambm se tornara menos hospitaleira aos homens. Embora as molstias tropicais do Velho Mundo, transmitidas por mosquito, possam ter sido introduzidas logo aps o primeiro contato,os cronistas do sculo XVI continuavam a louvar os bons aresdo Brasil.A malria,introduzida pelo trfico de escravos,ironicamente estimulava a continuidade deste porque apenas os africanos resistiam ao seu ataque violento.A febre amarela provavelmente estava se tornando endmica tambm entre populaes de primatas, j que os mosquitos infectados se reproduziam nas copas das bromlias do dossel,com influncias desconhecidas sobre a distribuio desses animais e sobre a composio da floresta e a disperso das plantas. Na mesma fronteira difusa penetravam escravos africanos foragidos, muitas vezes reunidos em comunidades independentes, chamadas quilombos, uma palavra da lngua quimbundo que significa unio ou ajuntamento. Esses assentamentos surgiram nas terras altas do Rio de Janeiro logo depois de ali se iniciar o trfico de escravos. Em Minas Gerais,onde a imensido da floresta em torno dos distritos mineradores possibilitava constantes fugas de recmchegados,novos quilombos brotavam por toda parte,muitas vezes ressurgindo nos mesmos locais. Os quilombos suscitavam medo e raiva entre os proprietrios de fazendas e os moradores da cidade, que logo contrataram caadores de escravos.Em meados da dcada de 1860,o capito do matoprofissional organizava expedies de cem ou mais caadores de escravos para destruir quilombos nas regies montanhosas do Rio de Janeiro.Em certa ocasio,os capites negociaram uma recompensa calculada pela intensidade do pnico dos fazendeiros: dois teros do valor dos cativos,excetuando-se apenas os cabeas, que eram enforcados, e as crianas nascidas livres no povoado. A leste do Tiet, ao longo do rio Paran,os caiaps,uma tribo guerreira que pode ter continuado a praticar o canibalismo, juntou-se em contingentes para defender, por um sculo inteiro,aquela margem contra os neoeuropeus.Ali exploravam os recursos das matas ciliares que eram o limite extremo da Mata Atlntica e cultivavam,incorporando sua dieta,bananas,arroz e galinhas.A itinerncia desses grupos agora terminava geralmente na aceitao da reduoem aldeias.Enquanto os portugueses mantiveram a poltica de isolamento da regio do outro a fim de diminuir o contrabando,a selva a leste,rumo ao litoral,permaneceu um porto seguro.
A incorporao de indgenas foi menos violenta ao longo da fronteira cabocla. O conceito de caboclo agora se aplicava a uma mistura cultural e racial mais complexa,com a adio de escravos africanos fugidos,e sua descendncia com mestios e nativos,s vezes separadamente categorizados como cafuzos ou caburs,o primeiro talvez um termo quimbundo que significava defeituoso,o segundo,outro termo tupi referente a uma pequena coruja marrom da floresta. Em alguns lugares prximos s aldeias neoeuropeia, os indgenas podiam ser encontrados empreendendo a lavoura por derrubada e queimada,sinal de que estavam livres da presso de expedies escravizao e de que algumas formas de intercmbio pacfico haviam sido restabelecidas.As vilas recmfundadas por Souza Botelho registravam,entre seus primeiros colonos,numerosos nativos,complacentemente identificados como guaranis. A troca cultural ao longo da fronteira cabocla seguiu percursos curiosos, mas, por certo, foi consequente.Escravos fugidos aprendiam dos nativos e,em troca,ensinavam-lhes prticas mgicas e religiosas,tcnicas de caa e pesca e remdios para picadas de cobra,feridas e doenas.O ritual afro-brasileiro atual substituiu quase totalmente as espcies nativas de plantas por plantas africanas.Esses fragmentos de evidncia sugerem uma reconstituio de alguma pequena frao de conhecimento da floresta que se perdera durante o sculo de caadas genocidas aos escravos. Infelizmente, africanos,caboclo e mesmo muitos dos indgenas eram todos refugiados em terra aliengena e duvida-se que tenham retido intatas as informaes que seus ancestrais acumularam.O portugus continuou a enriquecer-se com nomes tupis para plantas e animais e,nas zonas de minerao,os topnimos indgenas continuavam a ser aplicados.O portugus brasileiro aceitou uma palavra quimbundo at para o alimento mais bsico da colnia (fub).As tcnicas de caada da fronteira podem ser consideradas como inspiradas por crenas indgenas relativas aos espritos guardies da floresta. evidente que a opresso do governo colonial estava provocando seu prprio fracasso.Os pequenos agricultores no assumiriam residncia fixa enquanto a proximidade da autoridade os sujeitasse ao confisco de seus excedentes e de seus filhos. A maravilhosa diversidade da Mata Atlntica inclua uma ordem notvel de criaturas invasoras ansiosas por se apropriarem das colheitas das zonas alteradas que eram as clareiras queimadas dos lavradores. Os ciclos naturais da floresta permitiam a infestao ocasional de animais roedores, por exemplo , que se multiplicavam prodigiosamente quando o bambu frutificava e,quando este se exauria, voltavam-se para as plantaes e milho armazenado.
Infinitamente mais variados eram os insetos, dos quais de longe o mais implacvel e desalentador era a formiga cortadeira,chamada pelos tupis de sava.Na verdade, para os lavradores nativos,essa peste era o nico fator limitante mais significativo:nenhum de seus produtos bsicos estava imune.Em 1587,o cronista Gabriel Soares de Souza foi o primeiro europeu a registrar o espanto dos invasores diante da voracidade dessas formigas. A opinio de Soares de Souza sobre as cortadeiras era tambm um lamento pela inviabilidade de uma colonizao de povoamento: Se elas no foram que o despovoar muita parte da Espanha [Portugal na poca estava unido Coroa espanhola]para irem povoar o Brasil:pois se d nele tudo o que se pode desejar,o que esta maldio impede,de maneira que tira o gosto aos homens de plantarem seno aquilo sem o que no podem viver na terra. O gnero Atta de formiga extremamente bem-sucedido ele se espalhou por todas as regies tropicais e semitropicais do Novo Mundo. Pelo menos cinco de suas espcies habitam na regio da Mata Atlntica.Seu sucesso se deve a uma adaptao extraordinria:as formigas consomem exclusivamente um fungo,que cultivam em buracos subterrneos. Na floresta, ocasionalmente, so vistas filas ondulantes de savas, cada uma carregando um retalho de folha como um guarda-sol em miniatura.Na floresta primria,contudo,as formigas no passam de uma entre inumerveis espcies em competio,e suas populaes so contidas. Infelizmente para os homens, muito poucas plantas que eles domesticaram eram, em algum grau, repelentes sava muito pelo contrrio e centenas de anos de seleo ainda tero de passar para que surjam variedades de plantas resistentes. Entre as criaturas que poderiam ser arregimentadas contra as formigas, poucas so persistentes. Os tamandus, infelizmente, evitam as cortadeiras. Embora agrnomos prticos tenham recorrido a outros insetos,mais notadamente uma outra formiga,a cuiabana,para combat-las,todos relutam em enfrentar os soldados da Atta quando h outras presas ao alcance.O trax da formiga fmea,chamada i ou tanajura,era e considerado uma iguaria,e at um afrodisaco,entre os povos rurais mestios.Lamentavelmente,era desdenhada pelos moradores urbanos neo-europeus e,por isto,no era comercializvel. As medidas defensivas dos homens, at o sculo XX, foram, em grande parte, inteis. As formigas recusavam iscas que poderiam danificar seus jardins de fungos. medida que as reas cultivadas se espalhavam e se intensificava a destruio humana dos habitat de predadores de formigas pssaros, lagartos e rs aumentava a infestao de sava. Em certo sentido,contudo,destruir a sava,na medida em que isso fosse possvel,era contraproducente,porque as
formigas certamente realizavam uma tarefa herclea de melhorar as qualidades mecnicas do solo e transportavam nutrientes. Um fazendeiro afortunado talvez plantasse em local cujas condies, por algum motivo indefinvel,eram desvantajosas para as cortadeiras,mas as nicas caractersticas ambientais desfavorveis para elas eram solo arenoso ou solo baixo,mido.A Atta evitava terra mida porque os lenis freticos elevados impediam-nas de aprofundar os formigueiros. A nica outra manobra de evaso eficaz era ceder velhos campos s cortadeiras e queimar novas faixas de floresta madura. Como observou Soares de Souza, em clareiras novas no h formigas para fazer estragos em nada. Como teria sido a agricultura brasileira, se no tivesse havido sava?Atrairia mais colonos europeus,como suponha Soares de Souza?Teria o ataque Mata Atlntica ocorrido to rapidamente?A lavoura por derrubada e queimada prosseguiria, por ser a tcnica menos trabalhosa, mas os campos limpos teriam tido mais valor como pasto e terra agricultvel, e o incentivo ao uso de tcnicas mais produtivas, que incluem o arado, teria sido muito maior.Em vez disso,a presena das cortadeiras confirmava a transitoriedade inerente ao mtodo de derrubada e queimada e fortalecia o engodo da floresta virgem. Os garimpeiros de ouro e diamante comiam carne de boi. O boi era a fonte preferida de protenas na dieta neo-europia,e o crescimento de um mercado monetrio fez os preos dos bovinos subirem e muitos lavadores dedicados subsistncia passarem para a pecuria. Na pastagem natural e sem cercas, o gado se reproduzia vontade. Aqueles que sobreviviam eram os que escapavam s onas e resistiam s doenas e rigores da plancie sem abrigo.Essas raas curraleira,caracu,Junqueira,crioula no apresentavam nenhuma vantagem a seus predadores humanos,exceto a rusticidade;ou seja,no precisavam de nenhuma interveno humana para manter seus rebanhos.Os fazendeiros no davam forragem e, por isso, a estao seca implicava severa mortalidade do gado, cujos sobreviventes requeriam vrios anos para alcanar o peso do mercado.O nico suplemento alimentar fornecido aos animais era o sal.Este nutriente,escasso nos solos das terras altas,encontrava-se em baixas concentraes nos barreiros leitos secos de lagos e margens de rios.A o gado,em companhia de animais selvagens famintos por sal,raspava com a lngua os galhos viscosos,um efeito,segundo um observador,muito parecido ao produzido pelo arranhar de uma superfcie em busca de ouro. A Coroa considerava o sal uma mercadoria conveniente para gerar receita. Decretou o monoplio da sua importao e venda, taxando-o por peso em estradas com pedgio e encerrando as atividades das salinas concorrentes em Cabo Frio.
O gado era vulnervel a morcegos, vampiros,moscardos e s larvas que invadiam as feridas que aqueles provocavam. Indiretamente, as moscas eram outra causa de invaso da floresta. Para escapar a elas,o gado e os cavalos habitualmente penetravam nas franjas da floresta, fazendo com os cascos suas prprias clareiras e transportando neles sementes de capim. O surgimento de gado em campos gramados e cerrados nativos foi um evento memorvel em termos botnicos. Os primeiros observadores contavam maravilhas sobre os pastos como era luxuriante o crescimento do capim e como o gado engordava rapidamente com ele. Bastava, porm, uma gerao ou duas de pastoreio por uma nica espcie para transformar essas paisagens ednicas. Sem ser tangido, o gado tendia a pastar demais no capim mais palatvel, de sorte que os campos definhavam em plantas raquticas, doentias. Contra essa transformao, os fazendeiros dispunham apenas de uma arma, o fogo, que utilizavam sem misericrdia. A queimada destrua pequenos animais roedores, rpteis,tatus,tamandus,insetos e pssaros que nidificam no cho convertendo as pradarias em um deserto. A queimada reduzia a substncia da planta no comestvel a cinzas, enriquecendo assim, temporariamente, o solo, se logo viesse a chuva. Mas a queimada danifica de modo sutil os solos e o capim. Destri plantas que se disseminam horizontalmente formando esteiras,em favor das que formam touceiras, expondo o solo e provocando eroso. A queimada, alm do mais, elimina nitrognio e as bactrias que participam de sua fixao ao solo, essencial nutrio animal. A deteriorao do pasto nativo o tornava, paradoxalmente, uma mercadoria escassa. Dessa forma, o gado se espalhava sobre terra da floresta que havia sido cultivada mas estava abandonada. Esses pastos eram muitas vezes chamados artificiais, sugerindo que eram plantados com capim selecionado, mas isso nunca ocorreu no sculo XVIII e era raro no sculo XIX. Em algum momento do sculo XVIII, capins africanos apareceram nas invernadas e palhadas reservadas para forragem de cavalos, prximo cidade do Rio de Janeiro. Podem ter sido introduzidos na regio da Mata Atlntica a partir das camas de palha dos navios de escravos. Capim-guin, capim - Par, capim-marmelada e capim-jaragu so espcies que, na frica, ocupam as franjas da floresta alterada. Uma vez chegados e aclimatados, esses capins por vezes foram disseminados deliberadamente.O capim-marmelada foi,por isso,durante algum tempo,chamado capim do irmo Lus em regies de Minas Gerais,em memria de um benfeitor clerical. provvel,contudo,que o prprio gado fosse o agente mais comum de disperso. Os capins foram saudados como
concorrentes mais eficazes das ervas em pastagens manejadas pelo fogo,mas eram de fato substitutos desapontadores. O sistema de pecuria era extraordinariamente improdutivo. Pasto nativo degradado e pasto convertido permitiam apenas um rebanho muito ralo, provavelmente no mais que uma cabea para cada dois a cinco hectares. tendncia, de longo prazo, de pastos e animais a degenerar, criadores de gado reagiram se expandindo cada vez mais para novos pastos, onde no haveria nem ervas nem nada mais que pudesse matar o gado. Assim, a criao de gado tornou-se quase to itinerante quanto a agricultura e, como esta, para ser produtiva degradava ecossistemas primrios. O terceiro sculo da invaso europeia da Mata Atlntica reduziu consideravelmente sua extenso. A minerao, a lavoura e a engorda de gado no sudeste podem ter eliminado, durante o sculo XVIII, outros 30 mil km. To ampla, completa e irreversvel havia sido a eliminao da floresta em Minas Gerais, durante o sculo XVIII, que Karl Friedrich Philipp Von Martius, o mais famoso dos botnicos acolhidos no Brasil nos anos de 1810, foi levado a supor que a regio do ouro e do diamante, a sudoeste de Minas Gerais, e a regio nordeste da cidade de So Paulo nunca haviam tido floresta, mas eram constitudas de campos gramados nativos. Um aspecto notvel dessa expanso que os ganhos para os neoeuropeus, com suas formas extraordinariamente perdulrias de explorao dos recursos naturais, foram to exguos quanto imensos foram os desperdcios. Thomas Lindley, visitando o litoral prximo a Porto Seguro em 1802, ficou assombrado de que em um pas que, com o cultivo e a indstria, chegaria fartura com as bnos excessivas da natureza, a maior parte do povo sobrevive em necessidade e pobreza, enquanto mesmo a minoria restante no conhece os desfrutes que fazem a vida desejvel. Padres de vida baixssimos, em um Brasil rico de bens naturais, espantavam os visitantes europeus, acostumados a obter retornos muito mais elevados de um cardpio medocre de recursos naturais. Naturalmente o objetivo era acumular capital no na colnia, mas na metrpole. Um levantamento de um sculo de melhorias no estoque de capital social de Portugal talvez fornecesse justificativa para o desperdcio de recursos que a Mata Atlntica sofrer, mas isso parece discutvel. Ser que uma proporo significativa da crescente riqueza britnica derivava de suas relaes comerciais nitidamente assimtricas com Portugal, de seu financiamento do trfico colonial de Portugal e de sua consequente extrao de grande parte do ouro cobiado? Se derivava, ento Mawe estava observando as consequncias de uma relao, em vez de um contraste entre duas esferas distintas, e a esterilidade da destruio da floresta pode ser atribuda,
portanto,como muitos o fazem,ao colonialismo e ao capitalismo.No entanto,existem muitas evidncias de que a autoridade real era obedecida apenas quando convinha aos colonos. As autoridades coloniais, na verdade, no raro se mostram mais esclarecidas e progressistas que seus governados em relao explorao da natureza. O que est em questo a efetividade e a eficincia com a qual se empregam recursos escassos, a habilidade que participa de sua transformao e a validade dos usos para os quais so empregados. Uma questo mais complexa a capacidade de considerar o mundo natural como algo mais que um conjunto de utilidades disponveis, um patrimnio inerentemente valioso mesmo quando no transformado e inexplorado. A Mata Atlntica estava comeando sua longa jornada rumo extino. Seus habitantes ainda no tinham nenhum resultado palpvel e mal a haviam descoberto. Cap. 6 A Cincia Descobre A Floresta. Quase trezentos anos se passaram desde que os primeiros europeus colocaram os olhos Mata Atlntica. Uma parte considervel dela havia sido degradada por seus descendentes, cujas demandas tinham sido poucas, mas devastadoras. A floresta continha uma rvore de cujo crtex podia ser extrada uma tintura, era o esconderijo de ouro e diamantes e recobria solos sujeitados a um tipo primitivo de lavoura e, nas etapas finais de sua degradao, a um tipo primitivo de pastagem. Atravs do saber vagamente rememorado dos indgenas, transmitido com dificuldade aos caboclos e africanos fugidos do permetro colonial, algumas espcies da Mata Atlntica haviam adquirido nomes e alguns nomes tinham se associado a usos. O conhecimento indgena que os primeiros invasores haviam, desdenhosamente, ignorado seria, a um custo considervel, recriado e, por ltimo, superado. A Mata Atlntica iria,enfim,ser objeto de curiosidade. Empregar a cincia para alcanar essas metas era colocar a Mata Atlntica diante da perspectiva de mais devastao. Se,por um lado, a explorao intensiva dos recursos da floresta poderia acelerar sua destruio,a explorao cuidadosa e deliberada possibilitaria mais rea preservada e o repouso necessrio para render frutos constantes.O papel contraditrio da cincia e da tecnologia no manejo Mata Atlntica comea aqui, no despertar da compreenso desses servidores civis de que a floresta seria manejada ou destruda. Em 1779, foi fundada em Lisboa a Academia Real de Cincias. Mantinha correspondncia com outras sociedades,estimulou a coleta e observao entre amadores nas colnias e publicava seus relatrios.
A primeira sociedade cientfica brasileira realizou reunies em Salvador, em 1759, sob os auspcios do vice-rei, o conde dos Arcos. Foram planejados estudos sobre a agricultura, a flora,a fauna e os minerais mas, se algum deles foi levado adiante, no deixou vestgios. Em 1772, outro vice-rei na poca, a sede do governo colonial havia sido transferida para o Rio de Janeiro -, o marqus do Lavradio, fundou a Academia Fluviense, cujo objetivo era estudar medicina, cirurgia,botnica e farmcia. Seus diretores principais scios eram membros do corpo militar do vice-rei. Na sesso inaugural,o farmacutico Antnio Ribeiro de Paiva queixava-se de que, em Portugal, o comrcio de ervas estava em poder de comerciantes ignorantes que, frequentemente, erravam na identificao de seus produtos, enganos que ele queria evitar no Brasil. A academia restaurou o antigo jardim botnico jesuta e nomeou coletores, jardineiros e artistas botnicos, mas teve breve existncia.O marqus do Lavradio promoveu a diversificao agrcola e a busca de plantas nativas teis, mas foi desviado de seu intento pela guerra do Sul; quando deixou seu cargo em 1779, seu grupo de cientistas amadores voltou com ele para Portugal. Os portugueses no haviam autorizado quaisquer instituies de ensino superior na colnia alm dos seminrios. Jovens de talento tinham de ir para Portugal em busca de instruo, e l,frequentemente, eram instados a servir ao imprio.Se retornavam ao Brasil, ficavam afastados do incentivo de colegas e impedidos de formar uma nova gerao de estudiosos.De fato,muitos dos projetos cientficos apoiados nesses anos no parecem ter merecido publicao ou intercmbio, permanecendo como segredos de Estado, de concepo to mercantilista quanto s polticas econmicas da Coroa. O mais desastroso, contudo, que as cincias em Portugal eram ideologicamente suspeitas. As concepes polticas e sociais subversivas do Iluminismo estavam entrelaadas com suas propostas cientficas, tanto na cabea dos governantes quanto na dos cientistas. Os cientistas portugueses, por sua vez, estavam habituados a trocar correspondncia com colegas franceses, porque a Frana era o lcus da maioria das inovaes e recursos para seus campos de estudo. Em 1788, o botnico Felix de Avellar Brotero fugiu da Inquisio para Paris. O abade Jos Correa da Serra,nascido no Brasil,foi obrigado a se exilar por duas vezes para escapar a seus rigores:em 1786,foi para Paris,onde se encontrou com Jussieu e Geoffroy Saint-Hilaire no Jardin ds Plantes;de volta a Lisboa em 1795,novamente sob suspeita por causa da ajuda que dera a um botnico francs exilado,fugiu para Londres. A essa altura, Correa da Serra e outros jovens cientistas passaram a ser protegido por um influente corteso, Rodrigo de Souza Coutinho, um polmata de distino intelectual e membro diplomado da Sociedade Lineana de
Londres. Em 1796 , Souza Coutinho tornou-se ministro das colnias e da marinha. Sua estratgia ao lidar com o perigo francs foi acelerar a absoro do conhecimento cientfico estrangeiro, que ele via no s como um meio de galvanizar as colnias atrasadas, mas tambm de garantir sua lealdade, empregando seus bacharis universitrios em projetos que em termos de concepo e exceo abrangiam todo o imprio portugus. Souza Coutinho concedeu tambm fundos do vice-reino ao padre brasileiro Jos Mariano da Conceio Velloso, que passou muitos anos estudando botnica no Rio de Janeiro. Designou Velloso como diretor de uma nova editora,que se especializou em editar e traduzir uma srie de escritos sobre melhoramentos em agricultura tropical coligidos de outras colnias europias. O perodo era propcio para a diversificao das exportaes agrcolas brasileiras porque os concorrentes caribenhos da colnia estavam arruinados pela guerra. Primeiro,a Guerra de Independncia norte-americana,depois as revolues francesa e haitiana e,finalmente,o imperialismo napolenico haviam levado as potncias coloniais a enviar imensas foras expedicionrias para a regio,praticamente extinguindo seu comrcio.Portugal continuou por algum tempo tranqilo margem dessa agitao e o Brasil passou a fornecer quantidades crescentes de acar,algodo,arroz e tabaco para o mercado europeu.Souza Coutinho renunciou em 1803,quando sua poltica de promoo do desenvolvimento econmico do Brasil em detrimento potencial dos interesses peninsulares foi interpretada como convergente demais com a dos britnicos,que estavam passando a considerar Portugal como descartvel. Napoleo convencera a Espanha a lhe permitir invadir Portugal, como represlia por sua omisso em participar de um boicote continental aos produtos britnicos. O pobre Portugal j estava sob bloqueio naval ingls, por haver prometido aos franceses que aplicaria o boicote. O prncipe regente,D. Joo, resolveu seguir a recomendao de Souza Coutinho,de seis anos antes,e viajou para o Brasil protegido pela mesma frota britnica que estava bloqueando sua capital. Mais de 20 mil cortesos, burocratas e comerciantes logo se dirigiram para a corte exilada. Vrios milhares de estrangeiros se juntaram a eles artistas,artesos,oficiais militares,comerciantes e at cientistas.O governo portugus reconstituiu-se no Rio de Janeiro.Rodrigo de Souza Coutinho,logo intitulado conde de Linhares,tornou-se o principal ministro do regente,encarregado das relaes exteriores e da guerra. Por toda parte, investimentos bem pequenos em tcnicas j amplamente difundidas na Europa evidenciavam que poderiam aumentar em muito a produo brasileira. Os cortesos torciam o nariz para queijos e manteiga malfeitos,carne-de-sol e peixe seco produzidos na colnia.Todos poderiam ter
se tornado artigos de exportao mas,em vez disso,sofreram concorrncia estrangeira. Infelizmente, a invaso francesa no s excluiu os cientistas naturais brasileiros do centro de sua rede de informaes, mas tambm enredou os cientistas residentes em Portugal em novas dificuldades. Enquanto isso, a Coroa no hesitou em instalar em sua nova corte poderosos instrumentos de investigao do mundo natural: uma grfica, uma biblioteca, uma escola de medicina, um laboratrio de anlises qumicas, uma ctedra de ensino agrcola (em Salvador) e uma academia militar, cujas funes incluam engenharia civil e minerao. Essas instituies ofereciam emprego a cientistas locais como Jos da Costa Azevedo, professor de zoologia e mineralogia, Leandro do Sacramento, professor de botnica e agricultura, e Francisco Vieira Goulart, diretor do laboratrio qumico. A imprensa comeou a editar,entre outras publicaes teis,um jornal chamado O Patriota,que imprimia regularmente notcias de melhorias agrcolas.Duas importantes instituies cientficas foram criadas:um jornal botnico e um museu de histria natural. Receber e aclimatar plantas tropicais de interesse econmico era uma das incumbncias do Jardim Botnico. Destinava-se a aperfeioar a transferncia de culturas coloniais,uma atividade de importncia crtica que havia sido realizada at ento emprica e esporadicamente. Manuel Arruda da Cmara, naturalista pernambucano, escrevendo em 1810 em favor da instalao de um jardim botnico em sua provncia, apresentava uma crtica ainda mais pertinente da poltica colonial e da concepo botnica anteriores. Quando sangue,lamentava,teria sido poupado se os portugueses tivessem transferido as especiarias asiticas quando entraram pela primeira vez no oceano ndico,em vez de tentar conquistar a regio e defend-la contra seus concorrentes. Arruda da Cmara identificava corretamente a transferncia de culturas tropicais como um empreendimento difcil para os europeus. Ao contrrio de suas transferncias para a Amrica do Norte temperada,no estavam familiarizados com as condies de crescimento das plantas que removeram da sia e da frica e com os ambientes para onde as trouxeram no Brasil. O prncipe regente rapidamente ordenou a invaso de Caiena (atual Guiana Francesa), em represlia pela invaso de seu reino e por receio de que aquela pequena colnia pudesse logo ser reforada. Souza Coutinho ordenou ao capito-general de Belm sua total runa,receando que a Frana retomasse o territrio na prxima conferncia de paz. Felizmente, a ordem cruel de Souza Coutinho encontrou a oposio do comandante da vitoriosa expedio luso-brasileiro-britnica, que foi persuadido
pelo governador francs a poupar La Gabrielle em todo o seu esplendor. O regente D. Joo recebeu agradecido as sementes e plantas enviadas por seu governador civil natural de Minas Gerais, Joo Severiano Maciel da Costa, que tratou de fazer com que o jardineiro francs ficasse em seu posto. O grande golpe seguinte dos portugueses foi fortuito. A maioria das plantas exticas de La Gabrielle procedia do jardim botnico da possesso francesa das ilhas Maurcio. Esse jardim, Le Pamplemousse,havia sido organizado pelo ativo e apropriadamente chamado Pierre Poivre,que havia comandado expedies de coleta pelos quatro cantos do oceano ndico.Um oficial portugus,Luiz dAbreu,capturado pelos franceses e preso nas ilhas Maurcio,conseguiu fugir e roubar algumas sementes antes de deixar a ilha. Usufruindo a fama por sua audcia, Abreu escreveu a um amigo, Rafael Bottado de Almeida, um senador da colnia portuguesa d Macau, para pedir sementes de ch. Estas chegaram em 1812, em um navio de guerra portugus. Germinaram e cresceram mas,porque ningum no Brasil tinha qualquer experincia no cultivo e beneficiamento do ch e ningum parece ter se lembrado do tratado escrito por Avellar Brotero sobre o assunto,D. Joo VI ordenou que se contratassem jardineiros chineses. Um carregamento de sementes de pimenta de Goa, aparentemente destinado a Ajuda, havia sido desviado para a Bahia em 1782. Outros embarques de pimenta-do-reino e canela foram feitos de Goa em 1787,1788 e 1790.O governador de Goa relatou,em 1800,que a canela,que era uma espcie silvestre ali,estava comeando a ser cultivada,sem dvida por insistncia de Souza Coutinho. Juntamente com introdues de plantas oficialmente patrocinadas ou conhecidas pelo governo, havia muitas outras cuja procedncia obscura. Saint-Hilaire encontrou diversas plantas de clima temperado crescendo no planalto paulista:morangos,pssegos,damascos e castanhas.Tanto ele como Johann Emanuel Pohl,que passaram pelo Rio de Janeiro em 1818,comentaram a grande diversidade de plantas ornamentais transferidas da Europa. Essas numerosas transferncias no tiveram nenhum efeito na diversificao de exportaes. Embora repetidas introdues de especiarias no Brasil fracassassem em termos de um mercado exportador, os franceses na Guiana, os holandeses em Java, os ingleses nas ndias Ocidentais e os rabes em Zanzibar foram todos bem-sucedidos no cultivo de canela, cravo e nozmoscada. Os incentivos oficiais eram muitas vezes contraproducentes.Frequentemente garantiam aos produtores a compra de sua produo,mas os pagamentos atrasavam tanto que provocavam sua bancarrota.Os comerciantes ingleses,que financiavam o comrcio entre Portugal e suas colnias,representavam um outro obstculo,porque competiam com as colnias britnicas.
Ao menos por algum tempo o arroz foi nica exceo a esse pssimo desempenho. Introduzido anteriormente no Norte, tornara-se um gnero domstico de primeira necessidade na dieta colonial. Seu sucesso como cultura de exportao pouco teve a ver,contudo,com o patrocnio oficial;ao contrrio,deveu-se em grande parte promoo comercial por um beneficiador do Rio de Janeiro,Manuel Lus Vieira. Exceto pela mangueira, que possui a admirvel vantagem da resistncia s formigas cortadeiras, a fruta-do-conde (talvez uma referncia ao conde de Linhares), a jaca e o chuchu, as frutas e os legumes introduzidos no mercado interno nesses anos foram ignorados pela massa da populao. Encontraram, talvez, pestes ou parasitas, aparentemente invencveis no Brasil da poca, ou, quem sabe, a srie notvel de frutas nativas que os indgenas havia muito tinham trazido ao cultivo parcial tornassem suprfluas as espcies asiticas. Curiosamente, a domesticao, aprimoramento e comercializao de espcies nativas brasileiras no estava entre os objetivos arrolados no decreto que reorganizou o Jardim Botnico do Rio de Janeiro. O cultivo das plantas at ento silvestres era, certamente, um projeto muito mais complexo do que a simples transferncia de plantas exticas j domesticadas. Entre os visitantes europeus, contudo, havia alguns cuja curiosidade e simpatia sobrepujavam o preconceito. Saint-Hilaire, um observador particularmente arguto, salientava que as frutas europias eram o produto de sculos de cultivo, mas que se as suas variedades silvestres fossem comparadas com as frutas brasileiras, estas ltimas seriam indiscutivelmente superiores. Continuava a ser difcil extrair dos povos tribais e colonos da fronteira seu conhecimento sobre os recursos oferecidos pela floresta. Seu saber chegava s cidades e portos do Brasil na forma de uns poucos artefatos e bens de troca,extrados dos esturios e das florestas. As plantas florestais de valor medicinal eram extremamente variadas; de fato, o saber medicinal dos indgenas era o nico aspecto de sua cultura que os brancos da cidade no desdenhavam. As assustadoras doenas dos trpicos os levavam a aceitar,temerosos,os vermfugos,febrfugos,remdios para a pi,doenas venreas e,o mais enganoso deles,antdotos para picadas de cobra.Alm destes,acrescentem-se as curas para queixas mais prosaicas reumatismo,clica,endoparasitas,diarreia,hemorroidas,erupes cutneas,inflamaes,infeces e deficincias alimentares. Apenas um desses remdio da Mata Atlntica, no entanto, tornou-se artigo importante do comrcio exterior. Foi a ipecacuanha ou poaia, cujo rizoma um emtico. Na virada do sculo XIX,o Rio de Janeiro exportava cerca de quatro toneladas dela por ano.A ipecacuanha uma planta comum que constitua parte da cobertura de solo da floresta primria.Os caboclos a coletavam como
uma cultura comercial,arrancando-a,geralmente,quando dava flor,porque era quase impossvel identific-la durante o resto do ano. Outras plantas foram urgentemente procuradas na Mata Atlntica: salsaparrilha, cnhamo e cinchona. A salsaparrilha era considerada um remdio especfico contra a sfilis. Diversas de suas espcies locais passaram a ser comercializadas,mas no tanto como produto de exportao,porque as espcies amazonenses eram muito mais apreciadas.O cnhamo era considerado o melhor material para a manufatura da cordoalha naval e tentativas inteis foram feitas desde os anos de1620 para aclimat-la na Bahia e no Rio de Janeiro.A Mata Atlntica continha muitas plantas que poderiam servir para cordoalha.Na regio do Rio de Janeiro,a guaxima,um cip,e,na Bahia a piaava,uma palmeira,eram apanhadas em estado selvagem,para fazer corda.No entanto,depois que se estabelecia o cultivo do cnhamo,ele era preferido. A cinchona havia muito despertara o interesse oficial. Correa da Serra parece ter enviado,de Londres,sementes dela,ou o que ele imaginou que o fossem.Vandelli formulara a hiptese de que a planta existia no Brasil porque o pas margeava regies onde os espanhis a colhiam. O cacau era colhido na selva em seu vale nativo amazonense quando Alexandre Rodrigues Ferreira constatou essa prtica extremamente trabalhosa. Alm do mais, o cacau,cultivado em seu habitat nativo,era vulnervel a parasitas coevoludos.Sua transferncia e cultivo,oficialmente estimulados,na floresta litornea do sul da Bahia nos anos de 1780 podem ser contabilizados como uma conquista do perodo.Mesmo assim,ainda levaria mais um sculo e meio at se tornar produto de exportao significativa. Apenas dois dos inmeros produtos silvestres da Mata Atlntica chegaram ao ponto de cultivo e exportao no perodo a cochonilha e o indigno. Um oficial da delegao espanhola da fronteira lhe chamara a ateno para um determinado inseto que estava infestando um cacto do gnero opncia ali mesmo,a seus ps.O inseto,observava,era a cochonilha,do qual se extraa facilmente um corante vermelho.O corante,segundo lhe dissera o oficial,era um item comercial importante no Mxico.Aparentemente,a cochonilha nunca havia sido notada pelos tupis;tampouco o conhecimento de seu uso fora difundido pela regio da Mata Atlntica.Em breve,diversos outros relatos sobre a presena da cochonilha foram fitos em Santa Catarina e no Rio Grande do Sul. Essa descoberta foi nica realizao substantiva da academia antes de se dissolver, embora justifique a memria da instituio. A cochonilha tornou-se em breve um artigo comercial, mas depois desapareceu das pautas de exportao por motivos que no so claros. O historiador ingls Robert Southey afirmou que ela passou a ser adulterada e por isso perdeu crdito junto aos importadores.
O ndigo foi o outro recurso nativo domesticado, exportado e, durante certo tempo, mais bem-sucedido que a cochonilha. Esse corante azul,extrado de numerosas espcies nativas da Indigofera,disseminadas tanto no Novo como no Velho Mundo,havia sido notado e utilizado pelos indgenas.Por um sculo e meio,contudo,os neo-europeus no fizeram nenhuma tentativa de explor-lo. Os reiterados experimentos de plantio e beneficiamento do ndigo eram incentivados pela disposio do governo de comprar toda a produo. Em 1779,havia algumas centenas de manufaturas de ndigo no Brasil,a maioria na capitania do Rio de Janeiro,que satisfaziam inteiramente a demanda portuguesa do corante. Velloso discernia a grande vantagem do comrcio de ndigo, comparado a culturas de plantation como a do acar. Embora menos lucrativas,culturas como o ndigo eram mais acessveis a pessoas menos poderosas e eram menos complicadas em seu equipamento,menos dispendiosas em sua operao,[e]muito menos destrutivas das florestas.O colapso do ndigo foi,portanto,mais que um incidente na histria comercial brasileira:foi um sintoma da dominao social que canalizava recursos para uma cultura nica.Esse foi um fracasso de mau agouro para a Mata Atlntica. Interesse central da Coroa, essencial defesa do imprio e ao aumento de seu comrcio, era o suprimento de madeira para embarcaes. Embora a construo de barcos fosse comum por toda a costa brasileira,existe muito pouco registro de navios ocenicos antes do sculo XVIII.Por mais de dois sculos,a maior parte da madeira prpria para navios ocenicos era enviada aos estaleiros reais de Lisboa(assim como a lenha usada nos foges dos palcios reais!) Em meados do sculo XVIII, contudo, havia comeado a se organizar uma grande indstria de construo naval em Salvador, no arsenal e estaleiros reais e em estaleiros privados. Navios de guerra e mercantes construdos na Bahia constituam, no final do sculo, a maior parte da frota sob bandeira portuguesa. Ordens reais no sentido de conservar a madeira naval comearam a ser editadas em 1698,quando foram proibidas sesmarias em reas de reserva dessa matria-prima.Embora poucas sesmarias fossem concedidas
posteriormente,o corte de madeira tornou-se uma indstria privada fortemente organizada.Ainda que formalmente supervisionada por guardas -mores e administradores,estes eram sistematicamente subornados e ludibriados por um pequeno nmero de madeireiros,serradores e tropeiros que efetivamente decidiam quando e onde a madeira seria cortada.
Os governadores executaram diligentemente essas instrues. Realizaram-se levantamentos sobre reservas e meios de transporte de madeira,traaram-se mapas e foram elaboradas tabelas de preos a serem pagos aos contratados ou proprietrios particulares.Parte dos posseiros na rea poderiam ser autorizados a ficar,desde que limitassem suas atividades pesca ou estivessem dispostos a assumir os encargos de guardas-florestais (esta uma medida proposta por ambientalistas de hoje).Prticas incendirias seriam punidas com priso. Foram criadas reservas adicionais.Na capitania de So Paulo,todas as madeiras de lei das florestas entre a linha do mar e a escarpa costeira eram declaradas passveis de expropriao real,e cinco reservas reais foram estabelecidas e demarcadas entre Paranagu e Santos. A inspees e relatrios indicam que no existia mais madeira adequada por muitas lguas nas proximidades das vilas maiores.Grande parte das rvores que serviam para construo naval j havia sido derrubada.Em Iguape,no havia mais rvores apropriadas num raio de sessenta quilmetros da barra. As polticas de Souza Coutinho haviam sido delineadas em parte por Baltasar da Silva Lisboa,um juiz brasileiro que estudara cincias naturais em Coimbra e conhecia de perto as florestas do sul da Bahia. O conhecimento de que as rvores da Mata Atlntica no cresceriam a partir dos tocos,que no se poderia faz-las crescer exceto na floresta e que seu crescimento era to lento que se passariam geraes antes que pudessem ser colhidas,sugeria que,ao contrrio das florestas temperadas do Velho Mundo,o replantio de madeira de lei nativa em arvoredos homogneos no era nem econmica nem ecologicamente vivel e que o suprimento de madeira de navios deveria ser sustentado apenas atravs do cauteloso manejo das florestas existentes,que inclua o corte seletivo e o incentivo regenerao de espcies arbreas de maior valor. Essa realidade no era de nenhum interesse dos madeireiros e seus aliados,que ficaram furiosos com a inesperada contestao de seu modo de produo. No sul da Bahia,essas medidas no ficaram muito tempo em vigor.Os madeireiros simplesmente se recusaram a suprir os estaleiros reais e o monoplio das melhores madeiras de lei foi rapidamente rescindido.Contra as
concepes da burocracia mercantilista,os opositores das polticas de Silva Lisboa levantaram os princpios do liberalismo. Por estranho que possa parecer que Portugal fosse obrigado a importar madeira da Nova Inglaterra,mais estranho ainda que o Rio de Janeiro,que em 1818 comprava a maior parte de sua madeira da distante Porto
Seguro,tambm a comprasse dos portos africanos,norte-americanos e suecos,de onde ela chegava pela metade do preo da madeira cortada em nvel local. No foi s ao longo do litoral que a competio por recursos florestais resultava em escassez de troncos,madeira serrada e at mesmo de lenha.Na regio serrana,os mercados eram mais dispersos,o transporte mais caro e a madeira naval totalmente inacessvel.No havia restries reais e as florestas com frequncia se consumiam em chamas antes que qualquer madeira fosse retirada. Naturalistas nascidos no Brasil podiam ser ainda mais mordazes em sua viso da destruio da floresta.O minrio Jos Vieira Couto,outro aluno de Vandelli,descrevia o agricultor brasileiro com um machado em uma das mos e um tio na outra,praticante de uma agricultura brbara,como algum que olha para duas ou mais lguas de florestas como se elas no fossem nada,e ele mal as reduziu a cinzas e j lana seu olhar ainda mais adiante para levar a destruio a outras partes;no nutre nem afeio nem amor pela terra que cultiva,tendo plena conscincia de que ela provavelmente no ir durar para seus filhos. Nesses textos, a devastao dos caboclos ilimitada;so capazes at de derrubar uma rvore para se pouparem o trabalho de galg-la para apanhar frutos ou desalojar uma colmia de abelhas silvestres.Sua agricultura no considerada til. possvel que a perda de recursos florestais fosse menor do que esses. lamentvel que os naturalistas de formao europia da virada do sculo XIX tenham deixado apenas memrias breves e plidas de sua experincia sob o dossel da Mata Atlntica.Cheios de queixas diante das inconvenincias do caminho,da incompreenso e parvoce dos habitantes,das dificuldades de manterem secos seus espcimes coletados e suas anotaes,raramente
confiavam a seus leitores suas sensaes em meio ao silncio e melancolia dos gigantes da floresta. A Coroa tinha intenes claras ao patrocinar a cincia natural no Brasil:ampliar suas receitas atravs do aumento de exportaes e melhorar as defesas imperiais por meio e da manufatura local de material blico.Fustigada pela turbulncia da guerra e da revoluo,no fez nenhum esforo para substituir seu governo desptico por uma Constituio ou para pr um fim s injustias da sociedade colonial.A escravido foi abolida em Portugal em 1773,mas no havia nenhum movimento para elimin-la no Brasil,agora que este havia se convertido na metrpole. Johann Emanuel Pohl, chegando exausto certa noite a So Joo Del Rei, ficou grato por encontrar hospedagem com uma cama limpa e confortvel, que ele considerou um sinal de civilizao. Quando escancarou sua janela na manh seguinte, contudo, deu de cara com os olhos de uma cabea sangrenta, crivada de moscas, afixada a uma estaca fincada na praa da vila, apenas alguns metros adiante .Devia-se esse espetculo repulsivo a um escravo que assassinava o seu senhor. Atitudes como essa obviamente reservavam para a investigao cientfica um papel tutorial, quando no francamente favorvel ao status quo. Baltasar da Silva Lisboa, em comparao,representava um posio mais direta honesta . Sendo um abolicionista que apoiava a extino do trfego escravo, observava uma conexo lgica entre essa instituio e a falta de respeito e venerao pelos benefcios inestimveis [da natureza], que levavam os brasileiros a destruir cruel e barbaramente suas maravilhas, sem seguir e imitar sua conduta. O brasileiro,observava Lisboa, no era menos ingrato,injusto e destrutivo para com sua prpria espcie no modo como tratava com homens que eram seus escravos,maltratando-os com punies que assombram a toda a natureza. A onda de investigao cientfica e os planos de utilizao racional da Mata Atlntica mal haviam sido contemplados pela ateno
real,quando,repentinamente, seu patrocnio foi retirado. D.Joo VI demorou-se no Rio de Janeiro aps a retirada francesa da pennsula ibrica em 1814,talvez porque receasse novos levantes polticos na Europa.Em 1815,declarou-se o Brasil um reino,igual a Portugal e ao minsculo Algarves,esperando com isto
pressionar os congressos que estavam prestes a redistribuir os despojos de guerra,mas tambm enganando seus sbitos brasileiros de que o Rio de Janeiro continuaria a ser a sede do imprio. Esses vertiginosos eventos polticos,encerrados em 1826,modificaram
consideravelmente a relao dos brasileiros com o patrimnio natural que herdaram.Livres da tutela colonial,aptos a decidir os termos de suas trocas materiais e intelectuais com o resto do mundo,aptos a redefinir suas relaes sociais como lhes fosse adequado,estavam finalmente livres para decidir o que fariam com o que restara da Mata Atlntica.No era de todo surpreendente que abandonassem as tentativas de investig-la,de racionalizar seu uso e conserv-la,optando,ao entusiasmo. Cap. 7 A Floresta Sob O Governo Brasileiro. O novo governo, desvinculado de sua subservincia a um monarca explorador e distante, viu-se abenoado com recursos e oportunidades extraordinrios. A independncia fora conquistada a um custo insignificante em termos de sangue.As classes abastadas em cada capitania haviam aceitado a legitimidade do imperador e sua Constituio,retirando com isto,das massas oprimidas,a oportunidade para um levante.O territrio brasileiro foi assim preservado integralmente,ao contrrio dos vice-reinos do imprio hispnico,que na mesma poca mergulharam em guerras civis cruentas e aparentemente interminveis. No se pode afirmar,todavia,que essa intelectualidade concebesse a nao independente como algo mais que um Estado e uma cidadania assegurados por fronteiras polticas definidas,ocupando um espao geogrfico consensual.A ideia de que poderia ser igualmente uma comunidade identificada com o mundo vivo,a partir da qual seus cidados retirassem perpetuamente seu sustento,era de imediato abafada,quase indiscernvel. A oportunidade propiciada pela independncia,por um Estado tranqilo e pelo esprito reformista dos tempos foi desperdiada.O imprio logo foi assolado por conflitos externos to constantes que parecia que seu destino seria o mesmo das ex-colnias espanholas.D.Pedro se envolveu na sucesso do trono portugus e,consequentemente,foi obrigado a renunciar ao seu. contrrio,por atac-la com redobrado vigor e
No de admirar que,a princpio,da agenda dos notveis que construram o novo Estado constasse predominantemente,na verdade,quase
exclusivamente,o engrandecimento pessoal e faccional,porque conquistar poder era pr-requisito para a realizao de programas substantivos.Tampouco de admirar que seu ressentimento para com a finada autocracia tenha se voltado contra muitas de suas polticas racionais,embora seja impressionante como muitas dessas polticas se frustravam por mera indiferena e incompreenso. O mais importante dos interesses que desejavam promover era o trfico escravo africano. O rei D.Joo VI havia sido obrigada pelos ingleses a restringir a escravido s colnias portuguesas ao sul do equador e a condicionar seu governo extino final do trfico.Julgou conveniente,em princpio,juntar-se s outras potncias europias na oposio escravatura. impossvel calcular a extenso do impacto de um regime de trabalho agrcola escravo sobre a regio de florestas do sudoeste do Brasil;identificar esse impacto,porm,no difcil.O afluxo desse grande contingente em si mesmo requeria reas muito mais vastas para culturas de subsistncia.Esses recrutas permitiram excluir os lavradores nativos itinerantes em regime de subsistncia e os pequenos produtores de safras comerciais do comrcio exportador,que os fazendeiros reservavam para si mesmos. O fato de permitir que essa forma de explorao suprimisse e distorcesse o emprego de habilidades humanas por mais 66 anos,at cessar,por fim,o fluxo de novos escravos e a ltima gerao deles,nascida no Brasil e cada vez mais saturada com a protelao,deixar de pegar na enxada,obrigando com isto a assemblia a aceitar em 1888 o fait accompli, a mais terrvel reflexo sobre as classes governantes da nao. Outro grande interesse dos fazendeiros era livrar-se de toda restrio ao seu monoplio sobre as terras pblicas.O sistema de sesmarias,uma fico legal que legitimara a usurpao do patrimnio da Coroa,mostrara-se veculo conveniente para esse fim em todo o perodo colonial.Durante o sculo XVIII,a Coroa continuara a emitir essas concesses para favorecer ricos e poderosos,com os quais se identificava e contava para povoar a vasta colnia,a produo de bens exportveis e defender suas fronteiras.
Uma causa importante da destruio da Mata Atlntica foi que o governo no dava nenhum valor terra que concedia to gratuitamente.Tendo consumido toda a floresta primria mais promissora em dada sesmaria,um donatrio costumava vend-la por uma ninharia e pedia outra,que normalmente obtinha sem dificuldade. Curiosamente, apenas a maior intensidade e escala distinguiam essas prticas da itinerncia praticada pelas classes inferiores e deplorada pelos
governadores e os prprios donatrios, sempre vidos de explorar uma fora de trabalho estvel. O resultado,segundo o relatrio,era que h na mesma capitania centenas de sesmarias h muitos anos concedidas e que no tm o menor cultivo;e os habitantes continuam sempre a pedir novas terras,sem que haja qualquer necessidade de conced-las Embora incapaz de conceber alternativas para as sesmarias,a Coroa procurava,cada vez mais,impor condies restritivas para sua concesso.O decreto de 1795,provavelmente obra de Rodrigo de Souza Coutinho,exigia a demarcao de todas as sesmarias existentes e futuras. Os proprietrios no tinham o menor interesse em que o Estado fixasse os limites de suas erras e legitimasse seus direitos. Preferiam a incerteza,para melhor invadir terras pblicas.Mas a incerteza levava violncia,em escala que devastava o interior e desafiava a autoridade da Coroa.O fato de que os proprietrios de terra preferissem derramamento de sangue estabilidade de ttulos bem definidos garantidos pelo Estado sugere um sistema poltico ainda mais centrfugo que o feudalismo. A posse dependia da violncia ainda mais do que a sesmaria. O presidente do Rio de Janeiro,em 1840,confessou abertamente na assemblia de sua provncia que sabido que para estabelecer uma posse e mant -la [...] indispensvel a fora.Aquele que carece dela obrigado a ceder a terra a outro que mais forte ou a vend-la a algum que seja capaz de ret-la mediante a mesma fora. O governo imperial nunca realizou um inventrio das terras
pblicas.Ficava,pois,limitado a indagar dos conselhos municipais se existia alguma em sua jurisdio.Ardilosamente,estes sempre notificavam que todas haviam desaparecido.
A incapacidade do imprio de controlar as terras pblicas na verdade,sua inclinao a ser conivente com a expropriao privada sem custo algum para os expropriadores foi uma das maiores causas do rpido desmatamentos.Um cronista de Minas Gerais do sculo XIX reconheceu os terrveis efeitos dessas polticas:Os sesmeiros derrubaram e queimaram a floresta,no beneficiaram os terrenos e,quando lhes faltava espao necessrio para as plantaes,abandonavam as sesmaria ou vendiam-nas por pouco mais de nada e iam requerer nova sesmaria ou apossavam-se de terreno em outro lugar. A aplicao patriarcal da lei de herana foi um fator que estimulou a expanso explosiva da fronteira no sculo XIX.Alida Metcalf demonstrou que os pais habitualmente favoreciam genros e filhos mais novos na concesso de suas propriedades,na tentativa de protelar ao mximo a inevitvel transferncia do controle da famlia e seus recursos para a gerao mais nova. A terceira facilidade pleiteada pelas classes proprietrias de terra era a constante remoo dos indgenas de suas terras e seu recrutamento forado para a fora de trabalho como auxiliares,se no mesmo como escravos.Os monarcas portugueses diversas vezes haviam proibido sua escravizao,e tentaram de novo de modo mais retumbante em um decreto que entrou em vigor na regio da Mata Atlntica em 1758. A chegada da corte portuguesa no levou a uma poltica mais humana.A declarao de guerra do regente aos botocudos,que habitavam a regio serrana entre os distritos de ouro e diamante e o litoral,destinava-se a incentivar a garimpagem na regio,at ento interditada para a colonizao a fim de coibir o contrabando. As aldeias eram estabelecidas para receber os cativos trazidos pelas tropas,fix-los em um lugar para que pudessem ser catequizados e ficar disposio para trabalhar nas terras dos brancos.Essas aldeias eram confiadas no aos missionrios,mas aos mesmos comandantes de armas cujos salrios eram acertados pelo nmero de nativos que matassem ou capturassem. Os indgenas eram capturados por cidados patriotas que organizavam entradas na floresta.Os aprisionados dessa maneira estavam sujeitos escravido pelo prazo,fixao na declarao de guerra,de dez anos;logo depois,esse prazo foi elevado para doze,depois quinze anos.
Surpreendentemente,em 1813,a corte designou para esse sinistro reduto genocida um oficial francs no exlio,que era simptico aos indgenas,Guido Thomaz Marlire.A floresta remota e ameaadora foi a princpio apenas uma oportunidade para demonstrar sua lealdade,mas ele logo se afeioou a ela e ficou mesmo depois que o rei D.Joo regressou a Lisboa. A partir da,a preocupao constante de Marlire,conforme informou a seu superior em 1825,era precaver-se contra colonos brancos que procuravam usurpar terras dos nativos assediando-os com bebidas alcolicas,soltando gado em suas clareiras e expondo-os deliberadamente varola. As cartas de Marlire relatam dzias de crimes praticados pelos prprios funcionrios do governo a compra e venda de crianas,a conivncia de juzes com a grilagem de terras,o rapto de indgena para as milcias e para o trabalho nas estradas e roubo e leses corporais cometidas por seus prprios soldados. A rede de aldeias de Marlire chegava a cerca de duas dzias ao final de sua carreira,estendendo-se do alto Jequitinhonha at as cabeceiras do rio Pomba,uma distncia de quase seiscentos quilmetros,ao longo de uma faixa de uns cem quilmetros de largura.No auge do assentamento,viviam nelas uns 10 mil coroados e botocudos. Embora D.Joo viesse a entender que as rebelies nativas eram reaes aos maus-tratos e que as campanhas de pacificao envolviam assassinatos e crueldades,no via outra maneira de lidar com os indgenas seno intimidando-os e mesmo destruindo-os.Os partidrios da independncia no divisaram medidas significativamente diferentes das do regime colonial. As prescries que emanavam dessa anlise eram inevitavelmente similares s polticas do passado.Expedies,que Jos Bonifcio no se envergonhava de chamar de bandeiras,inspirado nos caadores de escravos do sculo XVII,deveriam ser enviadas para a floresta com intrpretes e padres,para persuadir os indgenas a se fixar em aldeias.As aldeias deveriam estar sob a superviso no de funcionrios militares,mas de missionrios. Foram estas,pois,as linhas grais da poltica adotada pelo imprio nos anos seguintes.As provncias,e bandeiras o ministro foram do organizadas lhes pelos governadores tolerncia das e imprio aconselhava
afabilidade.Aos notveis locais,eram concedidas faixas de terra,onde residiam
grupos tribais,na suposio um tanto hipcrita de que mostrariam aos nativos o benefcio da civilizao. Em 1843,o governo imperial convidou capuchinhos italianos,j presentes no pas,a assumirem a pacificao e assentamento de grupos tribais por todo o Brasil.Acima dos missionrios,estava um rgo governamental,o Servio de Catequese e Civilizao,representando em cada provncia por um diretor.Em So Paulo e no Paran,que foi separado de So Paulo em 1853,algumas colnias militarizadas,destinadas a resistir aos ataques nativos,foram
estabelecidas nos extremos da Mata Atlntica.Destas,a ltima foi assentada em 1888,na foz do rio Iguau. Embora a formao de aldeias prosseguisse at o final do sculo,em meados deste o zelo missionrio das pocas anteriores,por meio do qual como afirmou o administrador das aldeias de So Paulo em 1862 tantas almas foram conquistadas para a religio,tantos homens para a civilizao e tantas mos para o trabalho,no mais brilha[va]na Provncia.As aldeias tornaram -se obsoletas,seu papel perdera valor econmico.A ao predatria dos nativos sobre o gado solto em seus antigos terrenos de caa enfurecia os fazendeiros.O potencial de mo-de-obra desses silvcolas era de reduzido interesse.Os neo-europeus sempre quando os haviam tachado os indgenas de ltimos grupos
preguiosos,mesmo
escravizavam.Estes
sobreviventes eram de utilidade ainda menor que seus predecessores. Por isso,tornara-se comum a prtica de evitar os aborrecidos atrasos inerentes fundao de aldeias simplesmente assassinando nativos encontrados em terra cobiada.Isso podia ser feito com menor risco agora,j que o mercado brasileiro estava adequadamente abastecido de espingardas.Em So
Paulo,esses caadores passaram a ser conhecidos como bugreiros.Os funcionrios das provncias,a despeito de expirada a declarao de guerra aos botocudos,continuavam a autorizar assaltos e,de fato,a comand-los:em uma expedio punitiva,em algum momento depois de 1838,prximo a So Mateus,no litoral do Esprito Santo,o comandante militar do distrito retornou em triunfo com trezentas orelhas. Aqui e acol,alguns invasores procuravam poupar os silvcolas,aps o exemplo de Marlire.Em 1852,Tefilo Otoni,uma figura poltica visionria,caindo em desgraa na corte por haver participado da rebelio liberal de 1842,exilou-se no
vale do rio Mucuri,onde colocou em prtica um plano de acesso ao mar ao norte de Minas Gerais.Organizou uma companhia de transporte hidrovirio e rodovirio e,associado a ela,fundou um povoamento em um dos afluentes do Mucuri,no terreno onde hoje est a cidade que leva seu nome. Otoni pretendia que o Mucuri ficasse livre da escravido; por isso, introduziu imigrantes alemes, suos, belgas e outros, colocando como requisito que trouxessem poupanas em dinheiro e prometendo, em troca, que a companhia os trataria como associados e no como proletrios. Embora frequentemente Otoni fosse enganado por alguns de seus agentes de imigrao na Europa,que deturpavam as vantagens que a companhia oferecia ou enviavam patifes recm-sados da priso,o vale do Mucuri prosperou sob esse regime liberal de igualitarismo e respeito mtuo. Esse assentamento idlico sofreu, ento, vicissitudes fatais. Eclodiram conflitos entre indgenas e neo-europeus que continuaram at 1873,quando padres capuchinhos fundaram uma aldeia,chamada Itambacuri,cerca de quarenta quilmetros ao sul de Filadlfia.Ao contrrio dos jesutas,os capuchinhos,desde o incio,misturaram neo-europeus entre os nativos,encorajando o casamento intertnico.Os colonos,que desejavam se livrar dos capuchinhos,incitaram os indgenas a acreditarem que os missionrios os estavam envenenando.Os ndios se sublevaram em 1893,destruram a misso e voltaram para a floresta. Em So Paulo e Paran,apenas seis aldeias foram fundadas aps a independncia,mais por iniciativa de particulares.A mais duradoura foi a de Joo da Silva Machado,baro de Antonina,que em 1845 assentou em sua fazenda,na atual Itaporanga,estado de So Paulo,centenas de
caingangues,cujas terras haviam sido roubadas,a que se juntaram diversos guaranis. No final do sculo ,portanto,quase todos os ocupantes originais da Mata Atlntica tinham sido afastados ou expulsos.Residncia permanente nas aldeias era o sinal mais seguro de que a converso se efetuara.Embora os jesutas no dessem a seus pupilos permisso para partirem,os capuchinhos ficaram satisfeitos em traz-los gradualmente volta. O desaparecimento de povos nativos da Mata Atlntica,ntimos da floresta e empiricamente familiarizados com suas utilidades,no consternava os neoeuropeus.Os nativos tinham sido
ingratos,inconstantes,desleais,invejosos,segundo
Antnio
de
Souza,um
observador brasileiro sem papas na lngua,como outros do sculo XIX,que escreveu um livro de memrias de suas viagens pelo interior. De fato, admitia ele, seu presente estado de constante misria devia -se circunstncia de que tiramos deles uma grande parte de suas terras e florestas, reduzindo seu suprimento de caa e outros recursos. Em compensao,queixava-se de que os vizinhos dos indgenas haviam adquirido seus hbitos deplorveis,em particular a preguia,que era generalizada no Brasil. A quarta questo importante para os interesses dos proprietrios de terra era livrar-se da legislao florestal imposta desde o incio da colonizao. As proibies reais do corte de pau-brasil e de madeiras de lei adequadas construo naval negavam,em princpio,os direitos de domnio pleno da propriedade da terra,mesmo que essas proibies recassem sobre reas onde eram de aplicao quase impossvel e fossem fceis de burlar.O imprio dotado de Constituio preservou esses monoplios por certo tempo.Os interesses econmicos locais,j agressivos antes de a corte portuguesa deixar o Rio de Janeiro,abrigaram-se sob o novo regime para destru-los. O governo imperial deteve ainda por algum tempo o monoplio de compra e exportao do pau-brasil.A madeira corante representava uma fonte modesta mas valiosa de divisas que eram aplicadas na dvida externa e no custeio do inexperiente servio diplomtico.Infelizmente,era um produto sempre fcil demais de contrabandear a partir de dezenas de pequenos portos ao longo da floresta litornea. Enquanto isso,no interior,o extrativismo propiciava um meio de troca entre os indgenas que continuavam na Mata Atlntica,fugindo ao sedentarismo e sujeio,e os agentes comerciais das vilas e portos. repugnante listar,como um dos produtos extrativos,seres humanos,embora assim fosse,como havia sido desde as bandeiras do sculo XVI.Os botocudos capturavam as crianas de tribos rivais para vend-las como curucas aos neo-europeus,ao que parece a prelos consideravelmente abaixo de seu valor de revenda. As exportaes de ipecacuanha cresceram em volume de cerca de quatro toneladas por ano, no incio do sculo, para cerca de 25 toneladas no final dos anos de 1860. Nesta poca,contudo,caboclos do Rio de Janeiro,onde
arvoredos nativos da planta j haviam desaparecido,deslocaram-se para Minas Gerais,obviamente atrados tanto pelo extrativismo vegetal quanto pela esperana de descobrir ouro longo das trilhas da floresta.Infelizmente para a floresta,abriram estradas que estimularam o afluxo de migrao bem maior de lavradores itinerantes. Indgenas e caboclos eram caadores perseverantes. O naturalista alemo Georg Freyreiss relatou que muitos povos caavam,passando dias na floresta.Procuravam presas para o comrcio e tambm para subsistncia:peles de animais,pssaros vivo,peixes,macacos,flores,penas de pssaros,borboletas e outros insetos.Muniz de Souza esbravejava contra os caadores,a quem chamava a primeira classe dos vadios,mas uma pele de ona valia seis mil ris,o equivalente ao preo de um boi. Um dos primeiros entusiasmos que arrebataram os burgueses europeus foi a coleo de flores tropicais. Fascinados pelas memrias de viagem de cientistas como Humboldt,Von Martius e La Condamine,eram vidos expositores de trofus inanimados do mundo natural objetos como fsseis,pedras,pssaros empalhados e cristais.A regio Planaltina da Mata Atlntica,com seus invernos frios e chuvas sazonais intermitentes,era uma fonte excelente desses tesouros,porque suas espcies resistiriam a sistemas de aquecimento domiciliar incertos e inadequados da Europa setentrional e aos hbitos errticos de rega de jardineiros novatos. Essas incurses no interior da Mata Atlntica,fazendo dela um almoxarifado para os moradores das cidades da costa,e mesmo para os urbanoides sofisticados do mundo que se industrializava,sugerem uma homeostase salutar,uma floresta primria convertida em reserva extrativa,um conceito recentemente transformado em lei na Amaznia. A realidade era a rpida invaso do comrcio coletor pelos caboclos, que consideravam os indgenas apenas como um obstculo ao acesso direto aos recursos da floresta e que tinham apenas aprendido muito bem a lio de que a maximizao imediata dos lucros era alcanada mediante a minimizao dos insumos de mo-de-obra e supresso de qualquer respeito que podiam eventualmente sentir pela floresta que os alimentava. A epfitas desejadas pelos agentes comerciais ingleses,belgas,franceses e alemes se aninhavam nos galhos das rvores mais altas da Mata Atlntica.Eventualmente,todas as
orqudeas
de
uma
dada
localidade
podiam
ser
removidas.Uma
localidade,abandonada pelos garimpeiros de ouro anos antes,refloresceu na metade do sculo pela chegada de um agente comercial estrangeiro que comprava orqudeas at por dois mil ris a unidade.Moradores locais entravam alegremente na floresta com seus machados,abatendo rvores at que as orqudeas se tornaram to raras que no valiam mais o tempo do agente;ento,segundo o botnico lvaro da Silveira,que relatou este caso,o explorador dos caboclos abandonou os exploradores da floresta e a localidade voltou a afundar na apatia. Para os colecionadores europeus, mais importante que a beleza, fragilidade e exotismo era a raridade das epfitas. A raridade,em termos grosseiros,elevava o seu preo. De longe,o mais valioso dos produtos extrativos da Mata Atlntica era sua madeira de lei.Equipamentos de serraria tornaram-se comuns em 1820;em 1838,havia 53 serrarias em So Paulo.Nos anos de 1850,a siderrgica de Monlevade,em Minas Gerais,estava fabricando lminas para serrarias.Havia surgido uma modesta demanda externa de madeiras de lei,vendidas sob os nomes genricos de jacarand,pau-brasil ou palissandra e utilizadas em marcenaria fina. A derrubada comercial de madeiras de lei era realizada em vales dos rios abaixo das corredeiras;na regio da Mata Atlntica,essa era uma rea muito limitada,devido barreira dos penhascos costeiros.Grande parte das madeiras de lei era mais pesada que a gua e,ento,tinham de ser transportadas em balsas junto com madeiras mais leves para traz-las ao porto. No de admirar,portanto,que grupos tribais que haviam sido assentados em aldeias fossem capazes de se integrar no comrcio como
lenhadores.Inteiramente familiarizados com o terreno,eram especializados no reconhecimento das espcies de rvore e estavam dispostos a trabalhar em tarefas ocasionais e sem superviso. Nas terras altas,que no dispunham seno de transporte de mula,a madeira de lei tinha pouco valor para os proprietrios de terra,afora suas necessidades imediatas.Quando uma faixa de floresta tinha de ser queimada,suas rvores raramente eram tiradas de antemo.Tal material til poderia ter sido armazenado para necessidades futuras,mas,ao que parece,armazenar era uma
prtica ainda mais rara,sugerindo que a imprevidncia seria mais justamente atribuda aos neo-europeus. Espantosamente,o Rio de Janeiro importava mogno da Jamaica,pagando consideravelmente mais caro por ele que pelas madeiras nobres
locais.Saldanha da Gama espantou-se de ver o jacarand sendo utilizado na Europa para o mais refinado trabalho de marcenaria. O Brasil independente,liberado da opresso mercantilista,no mais um peo movido por uma monarquia portuguesa avara para defender seu jogo no tabuleiro de xadrez europeu,decidira adotar polticas autnomas que lhe permitissem prosperar acelerando a explorao de seus recursos
naturais,intensificando,em suma,a economia extrativista colonial,mas agora em uma era de livre comrcio e agricultura racionalizada.A Mata Atlntica era um ativo fundamental e a maneira de administr-lo no diferiria da dos tempos passados. Tais eram, frequentemente, os argumentos em favor de estabilizar o curso da economia brasileira. No entanto,Moraes Navarro se perguntava, podero eles alcanar todas essas vantagens sem se privarem de muitas outras,que mediante seu erro esto perdendo,e que seus descendentes no sero capazes,mesmo que o queiram,de reparar?.Essa pergunta foi ignorada depois da independncia poltica.O Estado transferiu para os grandes proprietrios de terra,e para os que tinham recursos para virem a s-lo,o direito de converter a floresta em dinheiro o mais depressa possvel.A Mata Atlntica passaria por uma nova provao quando seus proprietrios de terra se desviassem para uma forma nova e drstica de produo.Abandonando em grande parte a busca de pedras e metais preciosos,tornaram-se novamente agricultores, adotando uma cultura extica de enorme potencial econmico. Cap. 8 O Caf Desaloja A Floresta. As polticas sociais exigidas pelos grandes proprietrios de terra do Brasil independente e imperial garantiam-lhes uma fora de trabalho e os credenciavam a qualquer poro de terra pblica que quisessem chamar de sua. Essas polticas no podiam garantir,no entanto,que o emprego dos recursos assim assegurados gerassem capital;na verdade,esse monoplio autoritrio em grande parte frustraria sua utilizao eficiente.No interior,havia
vastas posses,habitadas por esfomeados e controlada por strapas cuja reserva de capital consistia em bois e porcos e cujas habitaes,vesturio e conhecimento do mundo eram praticamente indiscernveis dos de seus camponeses e escravos. A independncia havia melhorado as condies nas tais transferncias eram possveis,porque banira o jugo do mercantilismo,j retrado quando a corte se transferira para o Brasil.De fato,o governo britnico obrigara o imprio a adotar uma poltica de livre comrcio em troca do reconhecimento.Embora esse tratado caducasse em 1845 e tarifas mais elevadas fossem ento progressivamente impostas,a avidez do governo em estimular as exportaes jamais esmoreceu.O governo imperial desejava ampliar o comrcio porque praticamente toda a sua receita derivava de taxas de importao. Infelizmente para a classe proprietrio e o Estado,a mais cobiada das exportaes,ouro e diamantes,na qual o Brasil tinha longa experincia,no podia restabelecer sua proeminncia.As jazidas remanescentes,muito
exploradas durante a estada do rei no Rio de Janeiro,no davam base a grandes esperanas,pelo menos no nos termos da tecnologia em vigor.O baro Von Eschwege,o mais ativo especialista contratado para ressuscitar o setor,previa a necessidade de mtodos de uso mais intensivo de capital. Os capitalistas ingleses logo ocuparam esta lacuna e constituram uma srie de companhias de minerao de ouro.A maioria delas faliu,mas duas foram bemsucedidas,aplicando as tcnicas de poo fundo dos mineiros de Cornwall aos poucos veios importantes de Caet,e que em haviam Morro sido localizados,em da atual Gongo Belo
Soco,prximo
Velho,perto
Horizonte.Juntas,essas operaes somavam menos de um dcimo daquilo que se obteve durante o pico da corrida do ouro na metade do sculo XVIII.Muitas vilas mineradoras entraram em decadncia e seus moradores vagavam pela floresta para reassumir a lavoura da derrubada e queimada,ainda que com um olho sem dvida voltado para os riachos promissores. Os proprietrios de terra ou o governo tampouco se mostravam capazes de avanar na aclimatao e desenvolvimento das culturas tropicais que tanta ateno mereceram dos botnicos brasileiros e portugueses. O Jardim Botnico do Rio de Janeiro e o museu de histria natural rapidamente foram relegados ao quase abandono.Observadores estrangeiros
notavam,consternados,que de um parque pblico no qual no se desenvolvia mais trabalho botnico algum. O experimento mais inovador e conduzido com mais determinao, que se iniciou no Jardim Botnico no governo do ltimo rei portugus, foi a tentativa de aclimatar o ch. Os plantadores chineses de ch,trazidos de Macau em 1814 para cultivar as plantas e ensinar mtodos de beneficiamento das folhas,continuaram a trabalhar assiduamente no Jardim
Botnico.Permaneceram trabalhando no Jardim Botnico pelo menos por dez anos,alheios e solitrios,sob a suspeita do diretor do Jardim,Leandro do Sacramento,de estarem deliberadamente ocultando as tcnicas de
beneficiamento.Darwin ficou surpreso e desapontado ao descobrir que o ch,que ele viu no Jardim Botnico em 1832,sistematicamente plantado em fileiras retas,era um arbustozinho insignificante.Ach ou que a infuso preparada com as folhas verdes,que gentilmente lhe fora servida por um dos jardineiros, mal possua o aroma prprio ao ch. Sacramento parece ter desistido de tentar extrair dos chineses os segredos do beneficiamento do ch,antes mesmo de se aposentar da diretoria do Jardim Botnico.As plantas,contudo,interessaram pelo menos alguns proprietrios de terra.Ela crescia prodigiosamente at em solos inapropriados.A provncia de So Paulo importou um segundo grupo de chineses supostamente qualificados no cultivo e uma srie de fazendeiros importantes decidiu plantar o ch,inclusive Jos Arouche de Toledo Rondon,que escreveu um ensaio sobre seu cultivo e preparao.Tambm houve plantios experimentais no Jardim Botnico de Ouro Preto e de Santa Tereza,no Rio de
Janeiro.Infelizmente,amostras de ch enviadas Inglaterra em 1837 no foram recebidas favoravelmente.O horticultor belga Jean-Christian Heusser
considerou falhos os mtodos de beneficiamento e props melhorias muito simples.Tais medidas,no entanto,no foram adotados.O Brasil foi obrigado a importar a maior parte do seu ch,e apenas um sculo depois os colonos japoneses imigrantes foram capazes de abastecer o mercado interno. possvel que as dificuldades experimentadas na introduo de novos produtos no mercado decorresse em parte de preferncias imperiais no reconhecidas. evidente que a Companhia Britnica das ndias Orientais,uma empresa ainda monopolista,desejou manter seu controle do mercado,com base
em sua fonte chinesa.Era visvel a rivalidade econmica na reao dos franceses diante das notcias de que o Brasil estava cultivando ch.Em 1838,o botnico D.M.Guillemin foi enviado ao Rio de Janeiro para inspecionar esses esforos e para levar sementes para a Frana,de onde poderiam ser implantadas em suas prprias colnias. A regio da Mata Atlntica experimentou por certo tempo o renascimento da exportao de sua cultura mais tradicional e menos aprimorada, a cana-deacar do Rio de Janeiro haviam entrado em declnio no incio do sculo XVIII com a descoberta do ouro, o que aumentou o custo da reposio de sua fora de trabalho escrava. O acar brasileiro perdera mercados da Europa setentrional quando os holandeses,franceses e ingleses estabeleceram plantaes em suas colnias insulares no final do sculo XVII. A cana-de-acar era cultivada em quase todo povoado neo-europeu na regio da Mata Atlntica porque o mascavo acar no refinado,saturado de melao era um gnero bsico e porque a garapa era a matria-prima para a destilao da aguardente.Centenas,talvez milhares,de pequenos engenhos primitivos atendiam a uma demanda interna consideravelmente maior que a do mercado ultramarino. A maior parte do acar,e praticamente todo o acar,para exportao,era produzida em usinas movidas a boi ou gua engenhos que unificavam a moagem e a cristalizao.As usinas menores eram designadas,de forma pejorativa,engenhocas,s vezes operadas manualmente.Em geral,o seu produto era cachaa e no acar.A cachaa das engenhocas,destilada diretamente da garapa,era considerada superior dos engenhos,destilada do licor que sobrava do beneficiamento. A poltica do governo colonial em relao produo de acar e lcool, a partir de 1681, tinha sido aparentemente conservacionista: nenhuma usina de acar podia ser construda a menos de meia lgua (3,3 quilmetros) de outra, considerando que as reservas florestais de cada usina tinham de ser grandes o bastante para garantir a lenha necessria aos processos de fervura e cristalizao. Objetava-se, provavelmente com justia, que essas medidas eram meros pretextos para manter os privilgios dos proprietrios das usinas existentes e relegar os pequenos produtores de cana a uma situao de dependncia. As medidas eram tambm uma oportunidade para extorses
informais por parte dos funcionrios da Coroa: patente,por exemplo,que o governador do Esprito Santo tirou partido do decreto de 1802 para favorecer seus interesses particulares. O status especial,e contraditrio,desses produtores de acar que eram donos de engenhos fica evidente no epteto que sempre os acompanhou:senhores de engenho.Imagine-se um dispositivo to complexo quanto um engenho como o domnio de um senhor um ttulo que tinha tudo a ver com status social e nada a ver com engenhosidade ou empreendimento.O decreto de Pombal,de 1758,isentando os fazendeiro de acar do Rio de Janeiro de processos por dvida,resumia e legitimava o privilgio feudal de uma elite colonial imersa em um regime que,quanto ao resto,era implacavelmente mercantilista. A ressurreio do acar ao final do sculo XVIII incentivou certos proprietrios de terra a instalar algumas das melhorias que havia muito eram comuns no Caribe.A transio para tcnicas mais eficientes de moagem foi,contudo,muito gradual.Embora a mudana dos cilindros da posio vertical para a horizontal e a adio de um terceiro cilindro tivessem sido introduzidas pelos menos um sculo e meio antes,essas alteraes no tinham sido generalizadas.O primeiro engenho com cilindros movidos a vapor foi construdo em 1816.Da em diante,a converso prosseguiu e,pelo menos na regio de Campos,os engenhos movidos a vapor eram comuns em meados de 1850.O trem jamaicano a disposio ordenada de tanques de fervura graduados por tamanho,mais econmicos em combustvel. Melhorias na moagem no implicaram melhorias no plantio.A cana
taitiana,trazida triunfalmente de Caiena,foi rejeitada pela maioria dos fazendeiros.Embora essa variedade fosse de crescimento mais rpido,mais resistente aos ventos e de rendimento maior,seus talos eram mais finos e muito difceis de moer com cilindros de madeira ou de madeira chapeada com ferro. Por fim, aps vinte ou trinta anos, o fazendeiro perderia o interesse, declararia sua terra cansada e faria a solicitao de outra sesmaria. O cultivo do acar espalhou-se,assim,para alm da plancie de Campos,rumo ao norte e interior do sul do Esprito Santo e rumo a oeste e interior do piemonte florestado;no planalto paulista,mais para o interior,depois de Campinas. As exportaes brasileiras de acar podem ter chegado em mdia a 16 mil toneladas por ano durante todo o sculo XVIII, aumentando talvez para 30 mil
toneladas em 1850. Isso sugere a produo para exportao de cerca de 2,6 milhes de toneladas por um perodo de 150 anos.Qual o tamanho da rea da Mata Atlntica que tinha de ser derrubada para produzir 2,6 milhes de toneladas de acar?O coeficiente extrativo provavelmente no era melhor, em mdia, que 3,5%%, ou 74 milhes de toneladas de cana. A cana-de-acar tambm consumia a floresta na forma de combustvel para os tanques de fervura.Havia muita preocupao,principalmente no
Nordeste,com a enorme demanda dos engenhos por lenha.As exigncias de combustvel foram muito reduzidas pela implantao do trem jamaicano,de cerca de quinze para cinco quilos de lenha por quilo de acar. Os clculos sugerem que o plantio de cana-de-acar at 1850 no destrua tanto a floresta a ponto de ser abandonado em qualquer rea grande,mesmo levando em conta sua grande extenso.Existem,no entanto,alguns registros de abandono de plantaes devido exausto de lenha;casos
isolados,provavelmente. As plancies gramadas de Campos eram excepcionais. Ali,a falta de floresta nativa pode ter representado uma limitao real para a expanso do cultivo da cana-de-acar.A lenha era mandada de So Fidlis,pelo rio Paraba,e de mais longe,provavelmente descendo o rio Muria,em cuja margem,a uns oitenta quilmetros de Campos,existe ainda um local chamado Porto Madeira.Em 1844,um canal de cerca de cem quilmetros foi aberto at Maca,em parte para trazer lenha para os engenhos. O plantio da cana exigia outros produtos da floresta, em quantidade menor, mas no insignificante. A madeira era queimada para produzir cinzas para purificar o acar cristal.O acar continuava a ser embalado em caixas e certas rvores eram preferidas para tal fim porque no lhe conferiam cor ou sabor.Entre elas,estavam o jequitib e a tapinho,supostamente reservada para uso da armada.A cachaa era colocada em barris,preferivelmente de canela (Ocotea spp.).Todas eram rvores da floresta primria. Os produtores brasileiros enfrentavam um mercado internacional de acar que lhes foi favorvel apenas por pouco tempo. Surgiram muitos concorrentes,entre eles os produtores europeus de beterraba,aps as crises de revoluo e guerra que haviam propiciado ao Brasil a oportunidade inicial. Em consequncia, o
cultivo da cana-de-acar no sudeste da Mata Atlntica nos anos de 1850 entrou em declnio. A rea montanhosa, acima da cidade imperial do Rio de Janeiro, havia escapado ao plantio comercial da cana-de-acar, porque possua apenas faixas isoladas de terras de aluvio adequadas cultura. Mas outro produto tropical de plantation, que acenava para os proprietrios de terra nesta zona problemtica e ainda densamente florestada com a perspectiva de ganhos muito maiores,estava atraindo sua ateno. Essa cultura nova,o caf,tornar-seia,nas primeiras dcadas do imprio,a base da economia exportadora do Rio de Janeiro. O caf, ou Coffea arbica, uma pequena rvore da famlia das rubiceas,nativa do sub-bosque da floresta do sudoeste do planalto da Etipia, gerava as sementes carregadas de cafena to apreciadas pelos moradores urbanos da Europa.Seu oportuno surgimento resolveria de modo brilhante a busca de um produto que o novo imprio poderia trocar pelas manufaturas e luxos da Europa. A chegada do caf regio do Rio de Janeiro obscura. Sculos antes,a planta havia sido transferida da Etipia para o Imen,onde passou a ser cultivada comercialmente.Todo o caf, posteriormente introduzido no Sul e Sudeste da sia e no Novo Mundo, derivou de duas variedades do Imen, atualmente conhecidas como tpica e Bourbon.Sementes da variedade tpica podem ter chegado ao Brasil no final dos anos de 1600,talvez via ndia,mas o produto no era ento exportado. Com a formao da Academia Fluviense em 1772, Johan Hopman, um holands exilado que mantinha um horto, comeou a distribuir as sementes dessas rvores e instrues sobre seu cultivo. Os primeiros que as receberam podem ter sido dois padres que possuam fazendas nos subrbios. Em 1779, quando o vice-rei Lavradio entregou seu cargo, o caf ainda era to insignificante que ele o desconsiderou em seu relatrio final, muito embora mencionasse diversas outras culturas at menores. O caf passou a ser o produto das grandes fazendas doadas em sesmarias, enquanto corte portuguesa residia no Rio de Janeiro. Na verdade, o caf foi a salvao da aristocracia colonial. Foi tambm a salvao da corte imperial cambaleante,que,assediada por rebelies regionais e duramente pressionada a pagar pelas burocracias civil e militar necessrias para consolar o Estado, foi
resgatada pelas receitas do caf que afluam para a alfndega do Rio de Janeiro. Na poca, esse comrcio foi totalmente fortuito para o imprio. O imprio, portanto, mimou os fazendeiros do Rio de Janeiro: eles eram seu grupo de interesses primordial e seu esteio financeiro. Por outro lado, as exigncias ecolgicas dessa planta etope colocavam os limites fsicos para a reproduo do sistema de plantation e, portanto, para a estabilidade do imprio. A planta encontrou na provncia do Rio de Janeiro um ambiente adequado, se no ideal, para o seu cultivo. O caf mesial, isto , exige solos que no sejam nem encharcados nem secos. Nas reas altas do Rio de Janeiro, os fundos dos vales eram fracamente drenados e, dessa forma, o plantio tinha de ser em encostas ngremes e desencorajantes os mares de morros ou meias-laranja da paisagem fsica regional. A mata atlntica se estabilizaria, nessas reas, ao longo de milhares de anos de incipiente interveno humana, um solo raso mas moderadamente frtil e um tanto cido. Este material e a biomassa da prpria floresta podiam, por certo tempo, suprir nutrientes essenciais. Era precisamente este o perigo para a Mata Atlntica: acreditava-se que o caf tinha de ser plantado em solo coberto por floresta virgem. O capital e o trabalho eram escassos demais para gastar no plantio em solos menos frteis. O caf uma planta perene leva quatro anos para atingir a maturidade e pode permanecer produtiva por trinta anos e assim se podia imaginar que, uma vez implantado, representaria um regime agrcola de perspectivas estveis e conservadoras. Os fazendeiros no prezavam nem produtividade nem qualidade, apenas a economia de trabalho e capital e, no por acaso, seu prprio esforo administrativo. Esta estratgia, se assim pode ser chamada, gerava um produto de qualidade apenas medocre. Francisco Peixoto Lacerda Werneck, um dos mais importantes da primeira gerao de plantadores de caf do Rio de Janeiro, um homem que sem dvida conhecia bem seus vizinhos e partilhava da maioria de seus preconceitos, no estaria expressando nada alm de suas aspiraes comuns ao parafrase-los assim: O que eu quero muito [caf]; d um preo mais baixo..., mas eu tenho o mesmo lucro lquido, muito embora eu no tenha o mesmo trabalho. Felizmente, os norte-americanos eram menos exigentes que os europeus. medida que a populao e a economia dos
Estados Unidos explodiam mais e mais seus consumidores podiam dar-se ao luxo desse hbito e o imprio brasileiro ficava cada vez mais atado a esse nico mercado. Uma patrulha de reconhecimento foi encarregada de localizar espcies consideradas como padres indicadores dos melhores locais para cafezais. O manual dos agricultores, de Lacerda Werneck, com base em sua prpria experincia, apresenta reflexes sobre as prticas dos fazendeiros mais representativos e aparentemente foi o mais lido, recebendo duas outras edies aps a sua primeira publicao, em 1847. Em outros lugares e climas, o caf era cultivado na sombra, uma prtica que imita seu habitat original e que parece melhorar sua qualidade. No Brasil, em vez de preservar parte do dossel nativo, a floresta inteira era destruda na preparao para o plantio salvo, aqui e acol, um pau-dalho. Essas rvores eram poupadas, porque eram consideradas o mais seguro de todos os padres e, pois, exibidas para um comprador potencial da fazenda como prova da produtividade de seus cafezais. Nos meses frios de maio, junho e julho, turmas de lenhadores itinerantes eram contratados para executar a tarefa da derrubada segundo um sistema chamado picarias. Trabalhando de baixo para cima a partir da base da montanha, brandiam os machados sucessivamente contra cada rvore, talhando at que o tronco, ainda inteiro, gemesse com a iminncia de sua queda. Alguns desses troncos assim derrubados eram retalhados para fazer carvo para o mercado da cidade ou eram arrastados ou cortados no local, a serrote de dois cabos, como material de construo. As posturas municipais normalmente exigiam que os troncos derrubados em estradas ou cursos dgua fossem removidos. Era fundamental uma queimada de intensidade adequada: que no chamuscasse a camada de hmus, mas que no fosse to superficial que no produzisse cinzas suficientes para neutralizar o solo ou que deixasse ilesos os insetos residentes. A vegetao ressecada saltava em labaredas com um rugido e o espocar das varas de bambu, segundo um observador, soava como disparos de espingarda e os troncos rachavam como artilharia. As posturas locais exigiam tambm que, antes de atear fogo, fossem abertos aceiros e alertados os vizinhos. improvvel que, no caso de floresta primria afastada de vilas, tais barreiras, teriam custado um considervel trabalho
adicional, fossem regularmente adotadas, especialmente se o campo no confrontasse com a terra do vizinho. A destruio da floresta era lamentada apenas momentaneamente por parte daqueles que a assistiam; depressa seus sentimentos eram seguidos por outros pragmticos, ou estranhamente pirfilos. Os incndios de muitas clareiras elevavam imensas nuvens cinzentas de fumaa. O vale do Paraba deve ter parecido infernal ao final das estaes secas, com centenas de fogos se espalhando por todos os lados. Na metade do sculo, medida que se acelerava a derrubada da Mata Atlntica para o caf, uma nuvem amarelada pairava sobre a provncia durante esses meses, obscurecendo o sol de dia e apagando as estrelas noite. Tamanha era a quantidade de fumaa que durante os dias, e mesmo meses, o sol fica quase totalmente oculto ou, se o vemos, ele vermelho, quase como se o enxergssemos atravs de vidro enegrecido. Curiosamente, havia gente que na cidade no admitia que as queimadas fossem a causa e chegou-se a debater formalmente, na culta sociedade Vellosiana, por que o Rio de Janeiro experimentava essa assim chamada nvoa seca anual. O terreno assim preparado pela mo purificadora do homem assemelhava-se um pouco a um moderno campo de batalha, enegrecido, fumegante e desolado. Muitas arvores tombadas tinham sido apenas parcialmente incineradas; eram deixadas a apodrecer com seus tocos ainda enraizados e os troncos cados ao longo da linha da encosta. Os cafeicultores, na pressa de instalar seus cafezais o mais rpido possvel, no os dividiam em blocos. Isso teria sido uma perda de tempo, porque no se preocupavam em experimentar sementes de procedncias, tcnicas de plantio ou cultivo diferentes, ou mesmo manter registros de custos e rendimentos por cafezal. A densidade normal de plantio era de 800 a 900 plantas por hectare. Era extraordinariamente baixa, considerando-se a prtica moderna de trs mil a cinco mil. A baixa densidade permitia que as rvores crescessem em largura e altura. Dificultando a colheita, facilitando a invaso de capim. O pior de tudo que isso reduzia o rendimento por hectare. Se o plantio denso tivesse sido a regra, a derrubada da floresta teria sido em grande parte reduzida em sua extenso.
O cultivo no era realizado mediante princpios de conservao do recurso. O capim comeava a surgir nos solos florestais recm expostos dos cafezais novos s quando as plantas chegavam a maturidade, com trs ou quatro anos. As tcnicas usuais de colheita e beneficiamento reduziam a produo e a qualidade. Dividiam-se cotas aos escravos no campo e estes ento pelavam indiscriminadamente os galhos com frutos verdes, maduros e passados. Os mtodos de secagem, descasque e peneiragem dos gros tiveram de ser decifrados nos anos que se seguiram introduo do caf, um processo predominantemente emprico que adaptava implementos simples utilizados no descasque do arroz, moagem do milho e triturao da cana. Os proprietrios das fazendas no dispunham dos recursos para colocar de imediato todas as suas propriedades na produo de caf e, assim, o vale do Paraba se tornou uma colcha de retalhos de cafezais e floresta primria medida que, em primeiro lugar, as encostas voltadas para o norte e, depois, os locais menos favorveis, eram queimados e plantados. O caf chegou um pouco mais tarde ao Esprito Santo, onde as condies do solo e crescimento eram menos favorveis. Assim, o cultivo de caf espalhouse de modo extensivo e uma parcela considervel da regio montanhosa da Mata Atlntica foi transformada em um mar encapelado de ps de caf. O primeiro sculo do cultivo comercial do caf na regio da Mata Atlntica foi tambm o ltimo da escravido. Durante esse perodo, o Brasil produziu cerca de 10 milhes de toneladas de caf, quase todas passando pelos portos do Rio de Janeiro e Santos. Supondo-se que setecentos quilos fossem o rendimento mdio por hectare, e supondo-se que o cafezal mdio fosse economicamente produtivo durante vinte anos, ento foi necessrio desmatar para esse fim 7200 Km de floresta primria, o equivalente a trezentos milhes de toneladas de biomassa florestal consumida em fumaa. Na poca do colapso da escravido, as terras consideradas adequadas para o cultivo de caf estavam quase exauridas no Rio de Janeiro. O quanto seriam biologicamente nicas essas zonas da Mata Atlntica? Infelizmente, no se trata de uma questo que possa ser objeto de investigao retrospectiva, embora seja historicamente verificvel que poucas de suas espcies foram coletadas antes de virarem fumaa.
s vezes, afirma-se que a explorao mais radical de um recurso natural a mais rpida, acompanhada pelo menor dispndio de trabalho ou capital para que o recurso possa ser transformado o mais rapidamente possvel em mais capital, uma herana maior que o recurso intocado a ser transmitido para a prosperidade. A maioria dos recursos obtidos atravs da venda do caf no foi acumulada ou formada como capital para equipar a energia de uma gerao futura, mas gasta em bens na poca apreciados como luxos, exclusivamente importados para consumo das famlias que possuam as propriedades e os trabalhadores. As caractersticas da sociedade ps-colonial sua avidez pelo lucro imediato, concentrao de riqueza, fixao na vigilncia e no controle, empirismo extremo e total desrespeito por aquilo que apenas cem anos depois uns poucos iluminados lembrariam como riqueza natural inestimvel evocam o quanto ftil levantar agora tais objees quando o feito est consumado e nenhum trao da floresta restou sobre os morros secos e amarelados do vale do Paraba. A floresta primria sobrevivente, na regio da Mata Atlntica ou em qualquer lugar do Brasil ou, de fato, no resto do mundo tropical -, onde alguma parte dela encontre solos adequados para se disseminar, continua a ser um enorme tentao para qualquer um que, mediante um ganho, se disponha a administrar espcie humana sua dose diria de cafena. Cap.9 Instrumento da devastao
A queimada da floresta para plantar cafezais foi a principal causa, mas no a nica, do desflorestamento no sculo XIX. O comrcio do caf induziu o crescimento demogrfico, a urbanizao, a industrializao e a implantao de ferrovias. Consequncias indiretas da prosperidade febril baseada numa nica mercadoria de exportao, exerceram presses sobre uma rea mais ampla da Mata Atlntica, dando inicio ao que agora pode ser considerado como danos irreversveis a paisagem antropomorfizadas. A populao humana da regio sudeste da Mata Atlntica multiplicou-se de modo invulgar no sculo XIX. Contando cerca de um milho de pessoas em 1808, atingiu 6,4 milhes em 1890.
O comrcio do caf e de outros produtos de exportao finalmente tornava econmica instalao de meios de transporte mais eficazes que as tropas de mulas. Com a melhoria, a presso sobre domnios antes inacessveis da floresta tambm ficaria maior. A taxa de crescimento demogrfico no sudeste durante o sculo XIX, superou a do sculo XVIII, to rico em ouro e diamantes. Isso em parte foi resultado de um afluxo maior de escravos. A ameaa britnica convertia o trfico em especulao ainda mais febril e lucrativa. Considerando as elevadas taxas de mortalidade imputadas no sculo XIX, principalmente s populaes escravas, esse extraordinrio e contnuo aumento demogrfico deve ser atribudo, principalmente, aos teros das mulheres brasileiras, que heroicamente sobrepujavam o ritmo da morte por malria, tuberculose, enterite, esquistossomose, doenas de Chagas, picadas de cobra, tiroteios e milhares de outros males e perigos. Essa populao ampliada ainda era distribuda de maneira muito irregular. Continuava a existir uma fronteira de assentamento, alm da qual os povos tribais sobreviviam em pequenos bandos e ao longo da qual apenas uma escassa populao de pioneiros fugitivos da escravido, da servido ou da justia e colonos mestios se dispersava pela margem da floresta. Embora a fronteira continuasse a se expandir, as populaes no domnio neoeuropeu tornavam-se nitidamente concentradas: a provncia do Rio de Janeiro, que estava mais ou menos regularmente assentada em 1890, continha ento 32 pessoas por quilometro quadrado. Como poderia a agricultura itinerante alimentar uma populao to densa, da qual uma parte crescente consistia de moradores urbanos e trabalhadores das plantaes que no cultivavam seu prprio alimento? A produtividade por unidade de terra desmatada a partir da floresta primria era extraordinariamente modestos. A agricultura itinerante tradicional produzia excedentes de mercado que no s eram reduzidos como tambm incertos. As perdas de safras em cultivo devidas s formigas cortadeiras muitas vezes eram superadas pelas depredaes de diversos mamferos tatus, cutias e caititus e por pssaros que se alimentam de sementes e frutos entre eles, maitacas, pssaros pretos, periquitos e papagaios.
Quando as chuvas no vinham logo aps a estao da queimada, as cinzas simplesmente se dispersavam e seus nutrientes no se incorporavam ao solo. O plantio era ento quase intil, e os agricultores tinham que recorrer aos alimentos estocados. Um enorme prestigio era associado aos cultivadores de safras de exportao; nenhum, aos cultivadores de safras de consumo interno; uma escala colonial de valores que no foi reordenada pela independncia poltica. Os rendimentos inferiores, tpicos das terras de capoeirinha desmatadas, resultavam na expanso dos campos cultivados, e por isso reduzidos. Surpreendentemente, a adoo do arado no se generalizou nessa etapa. Mesmo em campos onde enormes tocos de arvore h muito havia apodrecido, os trabalhadores rurais insistiam em empunhar a enxada durante muito mais horas e dias que anteriormente e a rea mdia sob cultivo se elevou de talvez um hectare para trs a cinco hectares por trabalhador. Quando aps uma srie de plantios e abandonos, um campo no mais rebrotava como floresta secundria mas gerava apenas cupim, carrios e fetos, era abandonado ao pastoreio. A partida do fazendeiro era menos abandono que debandada. A crise na agricultura itinerante tornava o pequeno proprietrio extremamente vulnervel invaso do gado. Enquanto seus campos estivessem dispersos na floresta, dificultando a penetrao do gado. Estavam a salvo. Mas uma vez que a floresta degenerasse na forma de macega e matas de arbustos, o gado errante podia destruir suas plantaes. Os cupins africanos que haviam imigrado no sculo anterior eram a runa dos fazendeiros, tal o seu vigor e resistncia diante da enxada. No era prtico cercar campos temporrios. Faltava material para cercas: escasseava a madeira resistente podrido que serviria de moures; o clima e os solos no deixavam grandes pedras na superfcie e o arame farpado estava alm das posses dos pequenos proprietrios. Nas vizinhanas do Rio de Janeiro, os campos abandonados eram alvo de grande demanda enquanto pastagem temporrias, gado trazido de Gois, So Paulo, Minas Gerais e Mato Grosso, nos ltimos meses de inverno, tinha de ser engordado, ou pelo menos mantido vivo, para o mercado.
A manuteno da capoeira era crtica para a sobrevivncia de pequenos produtores. Fossem posseiros ou parceiros, precisavam de reservas
substanciais de matas. Normalmente, o fogo da cozinha ficava aceso durante o dia inteiro o feijo cozinhava durante horas -, ferver roupas, aquecer gua para o banho, secar guarnies de cama e mesa e roupas da estao chuvosa e para manter a cozinha aquecida no inverno. Os pequenos produtores necessitavam de lenha para beneficiarem suas safras comerciais. Por fim, as mulheres apanhavam lenha sem permisso, outra fonte de conflitos locais. Em todo caso, mais horas eram gastas a cada dia, caminhando para l e para c entre matagais dispersos, e a capoeira sofria ainda mais presso, extinguindose. Os fazendeiros tinham de abater rvores vivas para material de construo, moures de cerca, caixotes, cabos de enxada e machado, gamelas, cangas, moblias e muitas outras utilidades. O terreno de mata era tambm fonte de cips, utilizados como cordas e para substituir pregos. A cidade era tambm grande consumidora de lenha e carvo. Vendedores de lenha e fabricantes de carvo normalmente compravam os lotes que limpavam, embora s vezes trabalhassem por contrato. Havia tambm um ativo comrcio atacadista de madeira. Sessenta e seis fbricas de chapu de feltro, onze olarias, cinco fbricas de papel e papelo e cinco fbricas de vidro e porcelana necessitavam de processo a quente, alimentado por lenha ou carvo. A madeira de manguezais era a preferida para diversos objetivos, principalmente para alimentar caldeiras; queimava lentamente e no deixava sedimentos de resina. Os manguezais pantanosos se espalhavam numa faixa de 500 metros por trs das praias da baa da Guanabara. Por serem muito mais apreciados, devido ao tanino nas folhas e casca do mangue, os ltimos decretos coloniais haviam proibido sua queima antes da extrao da casca. A introduo de tijolos e telhas cozidos aumentou, consideravelmente, a demanda de lenha. O cimento e o estuque para as residncias eram feitos de cal, que tambm consumia lenha, normalmente de mangue. O mangue era o mais conveniente para o fabrico da cal derivada das conchas de ostras. Algumas reaes ao declnio dos solos da floresta primria podem ser observadas na agricultura do sculo XIX, que envolvia insumos muito maiores
de mo-de-obra: um sucedneo para a lavoura de arao em solos mal drenados de baixadas eram canteiros elevados, diversamente denominados leiras, valas, camalhes ou matumbos este ltimo um termo quimbundo, indicando pelo menos em alguns locais, mais uma contribuio africana explorao do mundo natural. No existem evidncias de que os caboclos deserdados tenham sido conservadores ideais da floresta primria para a qual eram empurrados. Continuavam a cultivar como o faziam no litoral, queimando e plantando durante curtas temporadas. medida que suas invases os levavam mais para o interior, chegavam a formaes florestais mais sujeitas a perodos de secas e dotadas de arvores que se caracterizavam mais como decduas. bem provvel que a queimada anual estivesse se tornando cada vez mais problemtica. Suas queimadas eram espantosamente amplas, porque no havia motivo para conte-las. As repetidas e frequentes queimadas de pastos nativos constituam uma grave ameaa s orlas interiores e mais secas da Mata Atlntica. Pode-se considerar que a difuso da agricultura de plantation, a pecuria e o crescimento de cidades e vilas no suplantaram, mas intensificaram formas tradicionais de explorao da floresta, sujeitando-a a presso suficiente para desestabiliz-la e degrad-la de modo permanente. Durante o primeiro sculo e meio de conquista e povoamento, o transporte por terra era feito a p, em trilhas no mais largas que as pegadas de um homem. Foi apenas no sculo XVIII que as mulas substituram os ndios como transporte de cargas. A corrida do ouro alterou essas circunstncias, fazendo com que as faixas considerveis de campos aurferos abandonados, fossem invadidas com maior intensidade. Os contrabandistas e comerciantes que seguiam as numerosas trilhas ilcitas para o interior desafiavam os esforos do vice-reino no sentido de legaliz-las atravs da instalao de registros. A chegada da corte no acelerou muito a construo de estradas. A doao contnua de sesmarias, o meio tradicional de estimular os particulares a abrir trilhas de mula, resultava, na maioria dos casos, apenas em mais picadas, fazendo assim uma srie de derrubadas na floresta para a abertura das mesmas.
Enquanto os outros produtos da Mata Atlntica tinham baixos preos e eram obtidos principalmente por meios extrativos ou predatrios, a rede de transporte por terra pouco diferia daquela traada pelos povos nativos. O comrcio cafeeiro forneceu por fim, o estimulo necessrio. Uma estrada pavimentada para carruagens foi construda conforme os padres de engenharia francesa entre Petrpolis e juiz de Fora. A estrada entre So Paulo e Santos foi alargada e reconstruda mais duas vezes. Os primeiros barcos a vapor na baa da Guanabara comearam a operar em 1821. A verdadeira revoluo nos transportes e, consequentemente, no relacionamento do homem com o que continuava intocado da Mata Atlntica foi a locomotiva. Sua introduo no Brasil chegou bem tarde no sculo. A primeira linha, terminada em 1856, que ia da orla norte da baa da Guanabara at a base da escarpa abaixo de Petrpolis, era pouco mais que um brinquedo da elite da capital. A ferrovia foi um imenso avano na paisagem desembaraada de quaisquer rastros alm dos ps humanos e casco de gado e montaria. A Mata Atlntica estava, pois, diretamente na trilha do que era tomado como progresso no sculo XIX. As ferrovias fariam suas prprias demandas floresta, porque exigiam grandes quantidades de dormentes, para os quais se preferiam madeira de lei da floresta primria. O impacto da ferrovia sobre a Mata Atlntica foi previsto por Gustavo Schuch de Capanema, um fazendeiro e cientista natural amador, em 1858, bem antes da primeira linha haver escalado o paredo costeiro. Capanema antevia, assim, que as ferrovias promoveriam a perpetuao da agricultura extensiva, itinerante, e acelerariam a destruio da floresta. O que restasse da Mata Atlntica seria no mais mero objeto de exame mas de interesse responsvel, seno de efetivo manejo. Cap.10. Especulao e conservao. O destronamento do idoso e enfermo imperador em novembro de 1889 foi realizado por um motim da caserna, liderado por um general de ideologia incipiente e ambio fulminante. Em meio contenda e rebelio polticas que se seguiram, a elite oportunista erigiu uma repblica ainda mais excludente que o imprio.
Os fazendeiros testemunharam com indiferena a expulso da famlia imperial porque a princesa Izabel, legitima herdeira do trono, assinara, na ausncia de seu pai, em maio de 1888, a Lei urea da abolio final. Para a Mata Atlntica, essa mudana rotineira nas fortunas polticas, com a perpetuao pacfica da ordem social, no deixou de ter consequncias duradouras. A constituio promulgada em 1891 transferia para os estados as terras pblicas anteriormente pertencentes ao governo central. Na regio da Mata Atlntica, elas ainda eram extensas. Diversos estados impunham tributos sobre a terra, no apenas para gerar receita, mas tambm para estimular prticas agrcolas mais intensivas. A repblica dessa forma devorou em poucas dcadas quase toda a terra da Mata Atlntica que restara ao poder pblico. O rpido aumento do valor da terra plantada com caf era um grande motivo de especulao. O especulador investia no plantio apenas um modesto adiantamento, pago a um parceiro, que trazia seus prprios escravos para a propriedade, e ao qual era permitido tambm intercalar plantios de subsistncia e vender a colheita de caf do primeiro ano. A nova regio cafeeira, o oeste paulista comeou a produzir quando o vale do Paraba e a zona da mata mineira entraram em decadncia. Os solos mais valorizados do oeste paulista, dibases to ricas em xidos de ferro que pareciam terra roxa, eram apenas ligeiramente cidos e continham mais nitrognio, potssio e clcio necessrios para a formao dos frutos. Queimar e derrubar amplas faixas de floresta primria tambm tinha o efeito de isolar o caf de potenciais agentes polinizadores. A derrubada e queimada da floresta com o propsito de assentar cafezais, portanto, prosseguiu em So Paulo at o sculo XX, em todo o estado e atravessou a fronteira, entrando no Paran, at consumir totalmente a Mata Atlntica que recobria o que se presumia fossem solos adequados ao caf. No entanto, essa extraordinria expanso das plantaes e o crescimento da populao foram acompanhados de crticas. Isoladas e a princpio
inconsequentes, ouviam-se manifestaes de desalento e desconfiana na elite dirigente desde os primeiros anos de vida independente do pas. Membros dessa sociedade expressavam o temor nascente quanto ao dano ambiental, provocado por mais de um sculo de atividade econmica intensificada e
populao mais densa. A lavoura derrubada e queimada era uma preocupao constante. A eliminao de florestas costeiras, portanto, implica um clima mais seco no interior. No sudeste, ao fim do sculo XIX, tinha-se alcanado a orla interior da Mata Atlntica em diversos locais. Mesmo sem interveno humana, essa j era uma zona de invernos secos mais longos, intensos e menos previsveis. medida que as matas nas proximidades das cidades se retraiam, o mesmo acontecia com os animais de caa. De consequncia mais imediata era o surgimento de pestes e pragas que comeavam a atacar os cafezais do vale do Paraba. Plantas tropicais domesticadas normalmente desfrutam quase de imunidade ao parasitismo durante um perodo considervel aps sua transferncia para um ambiente extico. Na verdade foi esta estratgia, totalmente inconsciente, que possibilitou a difuso mundial do sistema de plantation. No curso do sculo XIX, cientistas e tcnicos brasileiros se impuseram a tarefa de reintegrao com a comunidade intelectual europeia. A transferncia de conhecimento cientfico ocidental foi essencial ao sucesso das instituies brasileiras de pesquisas. A necessria colaborao com colegas estrangeiros distantes e, muitas vezes, indiferentes era um desafio difcil. A cincia brasileira se caracterizou por no beber da fonte de conhecimento sobre o mundo natural dos indgenas ou povos mestios da fronteira. Isso pode parecer uma impressionante e obtusa falta de curiosidade, mas facilmente compreensvel, dado o abismo cultural existente entre eles. Apenas o conhecimento europeu era vlido, no s aos olhos dos europeus autoconfiantes, mas tambm aos dos cientistas brasileiros ainda psiquicamente colonizados, empunhados em obter um apoio slido no seio das classes privilegiadas do imprio. A cincia oficial brasileira conseguiu um pouco mais de solidez aps a Guerra do Paraguai, que terminou em 1870, dando incio a um perodo, semelhante ao perodo reformista do incio dos anos de 1850, de crises de inflao e epidemias urbanas. O Instituto de Agricultura e o Museu Nacional iniciaram em suas reas a publicao das primeiras revistas cientficas que desfrutariam da continuidade necessria para a acumulao e transmisso do conhecimento cientfico.
O mais consequente dos cientistas paulistas da primeira gerao foi Alberto Loefgren, um botnico sueco que chegara com uma expedio de coleta e fora contratado para dirigir as sees de meteorologia e botnica da Comisso Geolgica e Geogrfica. Encantado com a natureza intacta da regio, declinou um convite para regressar a um cargo de pesquisa em sua terra natal, dizendo que, na Europa, tudo que havia para se pesquisar eram as mmias plidas das plantas deslumbrantes que ele pesquisava ao vivo no Brasil. O estado de So Paulo consumia a mdia de 1,5 milho de dormentes por ano na dcada que se seguiu a 1902. Madeiras de lei da floresta eram frequentemente utilizadas para tal fim, mas a madeira do serrado comeava a ser preferida. Em 1904, Antonio Prado, presidente da Cia Ferroviria Paulista, contratou Edmundo Navarro de Andrade para implantar uma reserva florestal. O resultado dessa experincia, foi uma vitria ntida das rvores do gnero Eucalyptus, nativa da Austrlia. Evidentemente, a conservao florestal no poderia ser implementada por estrangeiros, ainda que fossem estrangeiros bem intencionados, ativos, e havia muitos residentes no pas. Teria de surgir uma gerao nativa de cientistas e ativistas. Cap. 11 Nomadismo industrial, Industrial ismo predatrio. A regio da Mata Atlntica, na primeira metade do sculo XX, continuou a experimentar um rpido crescimento da populao humana, e isso continuou a ser a principal causa da acelerada destruio florestal. A grandiosa frente de assalto floresta primitiva a cada ano ganhava centenas de milhares de recrutas. Entre 1900 e 1950, a populao do sudeste, inclusive a regio da floresta latifoliada do Paran, cresceu cerca de sete milhes para 22 milhes. Uma economia regional dinamizada por exportaes estimulava o crescimento da populao. O afluxo de famlias trabalhadoras europeias e japonesas, atradas pelas plantaes de caf e algodo de So Paulo e do Paran, reduziu-se na depresso dos anos 30. A partir de ento, porm, o pauperizado nordeste brasileiro e o superpovoado sudeste inundaram a fora de trabalho. Enquanto a populao do Brasil triplicou entre 1900 e 1950, a de So Paulo quadruplicou e a do Paran cresceu quase seis vezes e meia. A floresta se tornou mais acessvel que nunca aos mercados urbano e mundial.
A usurpao de terras pblicas, nos limites mais distantes da Mata Atlntica, foi, at o final, uma atividade assassina. A terra encharcada de sangue t erra boa, observou um bem sucedido negociante de fazendas. Os cadveres eram principalmente de homens pobres. As cidades do sudeste, entretanto, multiplicavam suas funes comerciais, polticas e financeiras e adquiriam significativa capacidade industrial. provvel que a grande maioria dos habitantes da regio da Mata Atlntica encarasse essas transformaes com complacncia e at satisfao. Sobre suas florestas, os membros da elite brasileira projetavam
constrangedoramente suas ambiguidades relativas sua sociedade e cultura. No faltavam tentativas de substituir a flora e fauna nativas por espcies naturais europeias. A rusticidade dos caboclos talvez fosse a melhor prova de sua fora: abandonados e oprimidos por um governo que se recusava a reconhecer seu direito terra, no obstante, sobreviviam, multiplicavam-se e enfrentavam a floresta que aterrorizava imigrantes e o povo da cidade. O caboclo teimoso era o heri do ensasta Alberto Torres, que argumentava em favor do conservacionismo a partir de perspectiva muito diferente da de cientistas como Alberto Loefgren e Hermann Von Lhering. A maior ateno situao das florestas obrigou as autoridades a fazer levantamentos dos recursos remanescentes do pas. Um mapa florestal, produzido em 1910, por Gonzaga Campos para o Ministrio da Agricultura calculava que os quatro estados do Sudeste ainda continham 500 mil km de florestas, uma cifra sem dvida exagerada que aparentemente inclua formaes secundrias e cerrados. O mercado para o caf foi um assunto de considervel inquietao aps 1900, quando o plantio comeou a sobrepujar o crescimento da demanda mundial. Nos anos 30, o algodo tomou por algum tempo o lugar do caf como principal produto de exportao de uma srie de municpios, e o algodo e o acar continuaram a ser importantes gneros bsicos do mercado interno. Mas a associao do caf com a floresta primria no Sudeste permaneceu como fator decisivo na acelerao do avano da fronteira agrcola. Os arvoredos de floresta primria estavam sujeitos a presses extremas de extrao que no mais se pareciam com as atividades de subsistncia de
povos tribais ou caboclos. O extrativismo para exportao no desapareceu no sculo XX, mas, ao contrrio, ganhou mpeto. A transformao mais impactante nas faixas no queimadas de florestas primrias era, porm, a remoo seletiva de madeira. Na ausncia de estatsticas mais completas, difcil calcular a demanda total de madeira de lei. A escala do comrcio sugerida em registros da Estrada de Ferro Sorocabana que, nos anos 20 e 30, expandiu-se para o extremo noroeste do estado de So Paulo. Todas essas atividades extrativas significavam que lotes sobreviventes da floresta primria estavam se tornando menos intocados, menos complexos e menos densos. O Brasil ingressou na Era Industrial extremamente carente de um de seus prrequisitos: combustveis fsseis de hidrocarbono. As chuvas abundantes da regio da Mata Atlntica e sua topografia acidentada ofereciam enorme potencial hidreltrico, que comeou a ser explorado na virada do sculo. Nenhuma indstria teve maior impacto sobre as reservas de lenha que a siderrgica. Essas indstrias empregavam altos fornos e eram projetadas para serem acionadas por carvo vegetal. A lenha tambm alimentava motores a vapor, que forneciam energia em muitas fbricas, bem como uma pequena frao da energia eltrica. As ferrovias queimavam ainda mais lenha que a indstria siderrgica. Apesar do uso de certa quantidade de carvo importado e do incio da eletrificao nos anos 20, as ferrovias dependiam de lenha para mais da metade de seu suprimento de energia. Quase toda essa lenha vinha de florestas nativas. Alm do mais, os fabricantes de carvo comercial, ao contrrio dos catadores domsticos de lenha, preferiam a floresta primria porque ali se encontravam as rvores de lenha mais densa, que propiciavam os rendimentos mais elevados e o carvo mais rico em carbono. A primeira metade do sculo XX testemunhara o cerco final da Mata Atlntica. A expropriao privada de todo o territrio estava concluda. A civilizao urbana e industrial havia triunfado seus tentculos se espalhavam por toda parte, sua nsia por combustvel, madeira e outros recursos da floresta se estendia por toda a Mata Atlntica. Cap. 12 O imperativo do desenvolvimento.
No alvorecer do ps guerra mundial, uma terrvel ameaa se projetava sobre a Mata Atlntica sobre o que restava dela. Era uma ideia, na verdade, uma obsesso, chamada desenvolvimento econmico: a proposta de que se podia conceber polticas de governo que estimulariam a acumulao de capital e a industrializao e, com isso, um ritmo de crescimento econmico muito mais rpido que qualquer outro experimento na histria. Quase todas as transformaes fsicas e econmicas, dos anos 50 aos anos 70, que poderiam ser chamadas de desenvolvimento estavam confinadas regio da Mata Atlntica. Praticamente, todo investimento de corporaes multinacionais e estatais passou a se concentrar no tringulo industrial formado pelas cidades de So Paulo, Belo Horizonte e Rio de Janeiro. A indstria se expandiu vertiginosamente. O perodo foi politicamente agitado; sucessivos governos ostentavam polticas contraditrias em relao explorao e preservao da floresta,
circunstncias que reduzia ainda mais a possibilidade de que alguma parte da floresta primria pudesse sobreviver. As faixas remanescentes de florestas primrias, relativamente intactas, eram agora, em geral, as menos adequadas, ou inadequadas, para a agricultura do tipo que havia sido tradicionalmente praticado. Outra razo para a acelerada perda da floresta era a inflao crnica. O grande projeto de desenvolvimento econmico implicava a aplicao de uma proporo muito maior do produto nacional para a formao do capital. O desenvolvimento econmico aliviou algumas presses sobre a Mata Atlntica, muito embora tenha imposto diversas outras. As madeiras de lei de espcie variadas, dispersas pelas florestas primrias latifoliadas e, pois, difceis de cortar e transportar, eram exploradas de maneira diferente, embora igualmente descuidada e destrutiva. Alm disso, apenas umas poucas espcies mereciam ser cortadas; as demais eram queimadas in loco. O servio florestal estadual era reiteradamente instrudo a no interferir nas atividades dos madeireiros, que estavam contrabandeando madeira rio Paran abaixo. Migrantes afluam em massa para a regio, em busca de migalhas desse banquete. Era muito mais seguro invadir terra pblica, afinal o estado no contratava pistoleiros.
Os militares interromperam bruscamente o debate sobre a reforma agrria em 1964. Derrubaram o presidente esquerdista, Joo Goulart, e baniram centenas de polticos civis e lderes trabalhistas a quem consideravam subversivos e corruptos. Quaisquer que fossem os motivos dos generais, no decorrer de seus primeiros anos no poder, uma srie de leis e decretos foram aprovados sustentando a promessa de administrao efetiva do patrimnio nacional. Foi esboado um substitutivo para o Cdigo florestal de 1934. E o principal defeito do cdigo, de fato, era que ele nunca havia sido cumprido. Em 1967, foram promulgados os elementos finais no programa dos militares. Um novo cdigo de caa e pesca foi aprovado, autorizando a criao de refgios da vida selvagem e proibindo a exportao da caa. Assim, o governo militar ratificava o Brasil na trilha da conservao, firmemente subordinada meta do desenvolvimento econmico. Lamentavelmente, restava pouco da Mata Atlntica a conservar. Cap. 13 Desenvolvimento insustentvel. Durante a dcada de 70, a escala e velocidade dos projetos de desenvolvimento do governo militar atingiram o clmax que no resultou apenas em crise econmica, mas tambm em uma tempestade conjunta de desastres ambientais. O mais prejudicial de todos os programas de desenvolvimento talvez tenha sido o dos projetos hidreltricos. A topografia acidentada e as chuvas abundantes da regio da Mata Atlntica havia atrado empreendedores, engenheiros civis e fabricantes de equipamentos eltricos para colaborar, logo depois da virada do sculo XX, na construo de usinas hidreltricas nas proximidades de quase todas as cidades do Sudeste. Em 1950, havia 126 usinas hidreltricas na regio da Mata Atlntica. A partir do incio do sculo, milhares de quilmetros quadrados de florestas de galeria e semidecdua haviam sido eliminadas para gerar e transmitir eletricidade. O projeto Itaipu, contudo, finalmente provocou um questionamento severo, que no se limitou a ambientalistas ou especialistas em ecologia. A barragem de Rosana, cuja construo comeou, no incio dos anos 80, no rio Paranapanema, prximo a sua confluncia com o rio Paran, subtraiu mais de trinta km da minguante reserva do Morro do Diabo, a ltima remanescente do
Pontal. A companhia estatal de eletricidade cortou mais floresta do que era necessrio para a elevao das guas. Outras tticas empregadas na luta para liberar a economia do fardo do petrleo importado no deixaram de ter consequncias sobre a Mata Atlntica. O lcool combustvel foi criticado, principalmente, pelo custo de seus subsdios e pelo deslocamento das culturas que produziam alimentos bsicos para as populaes urbanas. Em So Paulo, as autorizaes de corte de madeira diminuram em nmero ao longo dos anos 70, porque os proprietrios de terra estavam ficando sem florestas nativas. A racionalizao da explorao de lenha recebia menos ateno do que se podia esperar. A lenha no era mais a principal fonte de energia no Sudeste industrializado, mas continuava importante. No Rio de Janeiro, a reserva ecolgica de Jacarepagu foi eliminada quando se permitiu que os interesses imobilirios se apropriassem da rea e a subdividissem. A despeito do extremo desprezo com que as preocupaes ambientalistas internacionais eram recebidas pelo governo militar e seus auxiliares tecnocratas, considerou-se prudente erigir uma fachada que pudesse minorar as crticas estrangeiras. Em 1973 foi criada uma secretaria especial do meio ambiente, encarregada de monitorar e controlar a poluio. Os conservacionistas civis eram capazes de exacerbar a permanente suspeita de interesses estrangeiros sustentada por seus patrocinadores de farda, como demonstrou um rgo ambiental oficial: foras supranacionais [...] exercem toda a presso para manter os pases subdesenvolvidos neste estgio. Apenas pequenos trechos adicionais da Mata Atlntica foram colocados sob proteo federal durante os anos 70. Os estados criaram apenas mais alguns parques e reservas equivalentes. A expanso extremamente modesta das reas de proteo foi uma reao absolutamente branda diante da destruio dos ltimos arvoredos primrios da Mata Atlntica durante a dcada de 70. No incio dos anos 80, os programas econmicos do governo, freneticamente alinhavados, passaram repentinamente a desfiar. Os emprstimos que havia contrado no rastro das elevaes do preo do petrleo tornaram-se impagveis quando os Estados Unidos elevaram exorbitantemente suas taxas de juros para suplantar uma espiral inflacionria. O Brasil chegou perto da
bancarrota quando novos capitais pararam de afluir. Isso foi fatal para o governo militar, cuja legitimidade, na poca, apoiava-se totalmente em seu desempenho em termos de desenvolvimento econmico. Cap. 14 Fazendo a lei funcionar. A retirada dos militares foi arrastada e rancorosa. O mandato do ltimo generalpresidente expirava em 1985. O congresso, repleto de membros nomeados, conseguiu rechaar uma campanha popular em favor da eleio direta de um presidente civil. Em vez disso realizou sua prpria eleio. O vencedor morreu quando estava prestes a assumir o cargo e foi sucedido pelo vice presidente eleito, um poltico de limitado respaldo e viso. A insero realista das questes ambientais nesse turbilho de dificuldades econmicas e reconstruo poltica mostrou-se, para dizer no mnimo, desafiadora para aqueles que se dedicavam a misso. A sobrevivncia da Mata Atlntica, assim erigida em preocupao mundial, no deixava de ser uma questo essencialmente local, apenas solucionvel por aqueles que a cercavam e abordavam. O movimento ambientalista estava se tornando, seno um movimento de massa, pelo menos mais poltico. Polticos que haviam zombado do movimento apenas alguns anos antes, agora estavam ansiosos para assumir as causas ambientais, pelo menos no perodo eleitoral. Contando com poucos membros, as organizaes ambientalistas tinham de contar com outras fontes de receita. A maior parte das maiores organizaes ambientalistas obtinha fundos da indstria privada. As organizaes
ambientalistas tambm demonstravam relaes problemticas com o prprio governo e geralmente procuravam no ser influenciadas pelo governo, mas influenci-los. Os ambientalistas que se lanaram no salvamento da Mata Atlntica prontamente saram em defesa de seus habitantes indgenas. A Mata Atlntica certamente continha moradores de classe inferior oprimidos, que estavam cada vez mais sujeitos expulso e violncia. Mas era difcil identific-los como ocupantes tradicionais de reservas. Situao mais grave era a das tribos guaranis que estavam saindo das florestas da serra do mar, no Estado de So Paulo, desde os anos de 1830.
Os planos para a preservao de reas adicionais da Mata Atlntica, concediam aos grupos tradicionais a ocupao de zonas tampo que circundariam zonas nucleares nas quais no se permitiriam quaisquer atividades humanas. O movimento ambientalista foi, direta ou indiretamente, responsvel por uma notvel expanso dos parques e reservas na Mata Atlntica do Sudeste. Em uma nica dcada, de 1981 a 1990, seu nmero dobrou, chegando a 205, e sua rea quadruplicou. No fim de 1993, o SOS Mata Atlntica havia concludo seu levantamento de toda a Mata Atlntica durante o quinqunio de 1985 a 1990. Mostrava que 5330 km haviam se perdido e que, naquele ltimo ano, restavam apenas 83500 km - um pouco mais de 8% da floresta que presumivelmente havia em 1500. Cap. 15 O valor da terra nua. No incio dos anos 90, a Mata Atlntica estava em uma situao crtica. Se aquelas faixas remanescentes identificadas ou supostas primrias iriam sobreviver, teriam de ser tomadas medidas drsticas imediatamente, medidas exatamente opostas s habituais cultura e ao governo brasileiros. No se sabia ao certo quantas espcies da floresta ainda existiam. Ainda mais vaga era a possibilidade de recriar partes da floresta que haviam desaparecido. O que era bastante evidente era que, na medida em que a floresta primria continuava a se converter em terra arvel e em pastagem, reservatrios e rodovias, clubes de campo e favelas, e em que mais e mais terras de propriedade pblica conquistassem novas instncias de proteo legal, as reparties governamentais estavam se tornando a ltima esperana de salvao da floresta. A reduo irresponsvel e perdulria da bacia amaznica a terra nua, por isso, deu margem ao alarme e ao escrnio internacionais. Entre brasileiros que estudaram a histria da Mata Atlntica e contemplaram a presena de seus arvoredos remanescentes, a floresta amaznica provoca especial alarme e pressgio. O ltimo servio que a Mata Atlntica pode prestar, de modo trgico e desesperado, demonstrar todas as terrveis consequncias da destruio de seu imenso vizinho do oeste.
6. GRIFFITHS, A.J. F. et al. Introduo Gentica. 9. ed. Rio de Janeiro: Guanabara Koogan, 2009. cap. 1 a 17, 19.
Cap.1 O ENFOQUE GENTICO A BIOLOGIA
Todos os organismos multicelulares, plantas e animais, sofrem um processo de desenvolvimento que transforma o zigoto na planta ou animal adulto. As novas clulas surgem com o resultado de grandes nmeros de divises celulares, e essas clulas diferenciam-se tanto morfolgica quanto bioquimicamente para atender a uma variedade de funes, tornando-se, por exemplo, clulas sanguneas, clulas nervosa, clulas musculares e clulas secretoras. Por meio dessa diferenciao. Por meio dessa diferenciao, as clulas tornam-se organizadas em tecidos e rgos, cujas formas e funes so caractersticas de cada espcie. Assim, as propriedades biolgicas de um organismo so o produto final de uma sequncia de etapas de desenvolvimento. Diversidade de estrutura. As estruturas celulares que levam a informao sobre o desenvolvimento e a funo devem ser capazes de existir em um imenso nmero de formas diferentes, cada uma especificando um aspecto deferente do organismo complexo. Deve haver informao sobre o desenvolvimento de todos os tipos diferentes de clulas, tecidos, rgos e processos bioqumico que caracterizam a espcie. Capacidades de se replicar. A fuso de um gameta masculino a um gameta feminino produz um nico camundongo, embora esse camundongo venha, na maturidade sexual, a produzir um nmero imenso de gametas que contm a informao necessria para fazer outra gerao de camundongos. Mutabilidade. Todos os membros individuais de uma espcie no so idnticos, mesmo assim, a prole tende a assemelhar aos seus genitores. Todas as espcies que existem, surgiram da evoluo de espcies
ancestrais que se diferenciaram delas em uma variedade de caractersticas. Traduo. No suficiente que exista um mecanismo apenas pra transferir informaes sobre um organismo de uma gerao para outra por meio de um zigoto. Os elementos bsicos do sistema de informao, herdadas hoje em dia, so chamados genes, um termo introduzido em 1909 por Wilhelm Johannsen, que investiga a herana em feijes. A coleo de todos os genes de um organismo chamada de genoma. O campo da Gentica envolve a diversidade, replicao, mutao e traduo de informao nos genes.
A cincia da Gentica comeou com um trabalho do monge austraco Gregor Mendel, que publicou o resultado de seus experimentos em cruzamentos entre linhagens que haviam herdados variaes em ervilhas em 1865. Mendel no os deu os resultados experimentais de cruzamentos controlados, como tambm deduziu a existncia de distintos fatores que levam a informao sobre o desenvolvimento dos genitores para a prole. No comeo do sculo XX, tornou-se aparente a informao que especfica do desenvolvimento dos organismos estava contida nos cromossomos do ncleo celular. A chave para a anlise gentica de uma propriedade biolgica examinar os efeitos das mutaes. Foi descoberto que bombardear organismos com raio X ou trat-lo com substncia qumicas induz um grande nmero de mutaes com efeitos claro sobre a morfologia do organismo. Algumas mutaes correspondem a delees observveis ou duplicao de pequenos trechos do material cromossmico. Pelos resultados dos cruzamentos entre as linhagens portadoras de mutaes diferentes, algumas perguntas importantes podem ser respondidas. O acmulo de evidncias, que comeou nos anos de 1920, levou concluso de que o DNA o material gentico. O poder analtico da investigao gentica aumentou imensamente mais recentemente pelo
conhecimento da estrutura qumica do DNA, de como a informao no DNA convertida em molculas de atividade celular e como sinais bioqumicos das clulas controlam que informao gentica ser lida por clulas em partes diferentes de um organismo em pocas diferentes em sua histria de vida. A anlise gentica, usando organismos com mutaes, um mtodo poderoso para investigar vias bioqumicas, fisiolgicas e desenvolvimentais.
A Base Molecular da Informao Gentica A estrutura dos organismos e seus ativos processos fisiolgicos so baseados, na maior parte , em protenas. A informao gentica para a sntese dessas protenas pela clulas esta contida no DNA, cido desoxirribonuclico. Uma molcula de DNA e feita de dois filamentos enrolados um no outro em uma longa dupla hlice. Cada um dos filamentos consiste em um arcabouo feito em cpias repetidas de um acar, chamado de desoxirribose, e fosfato; de cada grupo acar-fosfato ao longo do arcabouo, projeta-se uma base nucleotdica. Existem quatro tipos diferentes de bases nos nucleotdeos no DNA: adenina (A), timina (T), guanina (G) e citosina (C). Na molcula em dupla hlice, o arcabouo acar-fosfato de cada filamento esta por fora da hlice, enquanto cada base nucleotdica projeta-se para dentro e faz par com o filamento oposto. Devido ao espao ocupado pelas bases nucleotdicas, a adenina sempre pareia com a timina, enquanto a guanina sempre pareia com a citosina. As base que formam pares so ditas complementares. De algum modo, determinada sequncia de A, T, G e C deve ser usada pela clula para criar molculas de protenas com sequncias particulares de aminocidos. Alm disso, alguma parte do DNA deve agir como um sinal para maquinaria da clula de que a traduo de determinado gene em uma sequncia de aminocidos deve ocorrer em algumas clulas, em determinados tecidos e, em certas pocas, do desenvolvimento e vida do organismo.
A transformao da informao na sequncia de nucleotdeos em uma sequncia de aminocidos em uma molcula que ir dobrar-se para fazer uma protena compreende duas etapas: transcrio e traduo. Transcrio: A primeira etapa em transformar a informao no DNA em uma sequncia de aminocidos a transcrio da sequncia de nucleotdeos no DNA em uma molcula correlata, o RNA mensageiro (mRNA), que tambm tem um arcabouo de nucleotdeos como estrutura. O RNA mensageiro composto da molcula de acido ribonucleico (RNA), que uma sequncia de nucleotdeos similar ao DNA exceto por conter ribose em vez de desoxirribose em seu arcabouo e ter nucleotdeo uracil (U) em lugar de timina. Na transcrio a dupla hlice de DNA separada em dois filamentos nicos, e um desses filamentos serve de molde pra a construo de uma sequncia de DNA unifilamentar. O processo de transcrio ocorre no ncleo, assim muito similar ao processo de replicao do DNA porque o filamento de DNA serve como molde para fazer um filamento de mRNA. O mRNA copia da molcula original de DNA chamado de transcrito. O RNA que produzido diretamente da transcrio do DNA pode ento ser alterado para um mRNA final (RNA mensageiro). Essa alterao consiste em remover trechos do transcrito original que no codificam aminocidos. Traduo: a produo de uma cadeia de aminocidos com base em uma sequncia de nucleotdeos no processo de traduo. Com uma sequncia feita apenas de quatro tipos diferentes de nucleotdeos pode gerar uma sequncia de aminocidos composta de 20 tipos diferentes de
aminocidos? A soluo que a sequncia de nucleotdeos no mRNA lida em grupos sucessivos de trs nucleotdeos ao longo da cadeia de mRNA. Cada grupo de trs um cdon.
Do polipropileno protena: importante compreender que a estrutura primria que resulta sntese , a cadeia polipeptdica , no a molcula que nos chamamos de uma protena. Uma protena uma cadeia polipeptdica dobrada. Determinada sequncia de aminocidos, embora confine o
processo de dobramento, no o determina totalmente. De fato, para cada determinada sequncia de aminocidos, existem vrios dobramentos estveis alternativos. Alm disso, a clula pode fazer algumas alteraes qumicas nos aminocidos da molcula. O dobramento final que produz uma protena biologicamente ativa depende das condies na clula que incluem a presena de uma variedade de outras molculas. Essa dependncia tem efeitos prticos. A insulina humana usada pelos diabticos, hoje produzida em culturas de clulas para as quais o gene de insulina humana foi transferido. Quando primeiro produzida desse modo, a insulina no tem atividade fisiolgica, muito embora tenha a sequncia correta de aminocidos. A insulina, biologicamente ativa, foi depois produzida por mudana no mtodo de produo que resultou em uma molcula modificada.
Investigao Gentica A essncia do mtodo gentico como meio para explicar os fenmenos biolgicos uso das diferenas genticas entre os organismos. Dentro das espcies, existem diferenas de ocorrncia natural entre membros individuais das populaes. Por exemplo, cerca de um tero de todos os genes codificantes de protenas em espcie de reproduo sexual apresenta alguma variao entre indivduos nas sequncias de aminocidos que eles codificam. Esse poliformismo gentico uma fonte rica de material para o estudo das bases da variao biolgica entre indivduos.
Gentica direta Uma investigao gentica pode comear com a observao de uma variao detectada na morfologia ou fisiologia, isto , um fentipo variante. Assim, a primeira etapa na gentica direta pesquisar a diferena de gentica que causa uma diferena fenotpica. Tal pesquisa envolve procurar os padres de herana dos descendentes de cruzamentos entre indivduos que tem fentipos diferentes. Quando identificado um gene, um programa
de investigao , ento, desenvolvido para mostrar como a informao contida nesse gene normalmente transformada em eventos celulares e fisiolgicos que contribuem para as caractersticas normais do organismo. Uma forma de tal investigao comea com a variao natural normal no fentipo. A maioria das caractersticas fenotpicas so afetadas por vrias vias fisiolgicas desenvolvimentais que interagem, cada via consistindo em um nmero de etapas. Tais casos requerem vrios genes.
Gentica reversa O programa da gentica reversa comea com mudanas genticas conhecidas e procura as alteraes resultantes no organismo. Uma sequncia normal de DNA, podemos usar sua informao para ler a sequncia de aminocidos na protena que produzida, pois sabemos a correspondncia entre o cdigo do DNA e os aminocidos que so codificados. O DNA pode ento ser alterado de modos especficos que,ou bloqueiam totalmente a produo da protena , ou mudam a protena de tal modo que ela fica com atividade metablica gravemente alterada ou sem atividade metablica.
Metodologias em genticas Muitas metodologias diferentes so usadas para estudar os genes e atividades gnicas, e essas metodologias podem ser resumidas no seguinte modo: Isolamento de mutaes que afetam o processo biolgico em estudo, anlise da prole de cruzamento controlados entre mutantes e individuo tipo selvagem ou outras variantes descontinuas, anlise gentica dos processos qumicos da clula, anlise microscpica e anlise direta do DNA.
Organismos modelos
O uso de organismos-modelos vem do trabalho de Gregor Mendel, que usou cruzamentos entre variedades horticulturais da ervilha de jardim, Pisum Sativum, para estabelecer as regras bsicas da herana. O uso feito por Mendel dessas variedades de ervilhas em estabelecer as leis bsicas da herana instrutivo para a nossa compreenso das foras e fraquezas dos organismos-modelo. As leis de Mendel da herana foram a fundao da gentica, em particular, estabeleceram que o mecanismo de herana era baseado em partculas distintas nos gametas, que vieram juntas com a prole e, ento, se separaram novamente, quando a prole produziu gametas, ao invs de se misturarem como um liquido continuo. Mas Mendel no poderia ter deduzido esse mecanismo se tivesse estudado a variao da altura na maioria das espcies de planta, onde tal variao contnua, pois ela depende de muitas diferenas gnicas. A necessidade de estudar uma ampla gama de caractersticas biolgicas e genticas levou a uma gama de organismos-modelo para cada um dos grupos biolgicos bsicos. Vrus: essas partculas simples no-vivas no tem toda maquinaria metablica. Um vrus infecta uma clula hospedeira e modifica deu aparelho biosintetico para a produo de mais vrus, incluindo a replicao dos genes virais. Os vrus que infectam bactrias, chamados de bacterifagos, so o modelo padro. O principal uso dos vrus foi estudar a estrutura fsica e qumica do DNA e os mecanismos fundamentais de replicao do DNA e mutao. Procariontes: esses organismos vivos unicelulares no tm membrana nuclear nem compartimentos intracelulares. Embora exista uma forma especial de cruzamento e troca gentica entre clulas procariticas, na maior parte de suas vidas elas so essencialmente haploides ao invs de diploides. As clulas de organismos diplides contem dois conjuntos dos completos genes, cada um tendo contribudo por um de seus dois genitores atravs de gametas que se fundem fertilizao. Os organismos haplides tem apenas um conjunto de genes em cada clula.
Eucariontes: todas as outras formas celulares de vida so feitas de uma ou mais clulas com uma membrana nuclear e compartimentos celulares.
Genes, o Ambiente e o organismo. Os genes no podem ditar a estrutura de um organismo por si s. O outro componente crucial na formula o ambiente. O ambiente influencia a ao gnica de muitos modos. Mais discretamente o ambiente fornece matria prima para o processo sinttico controlados pelos genes. Um broto torna-se um carvalho usando, no processo, apenas gua, oxignio, dixido de carbono, alguns materiais inorgnicos do solo e energia as luz. Determinao gentica: praticamente todas as diferenas entre as espcies so determinadas por diferenas entre os genomas. No h ambiente no qual um leo dar origem a um cordeiro. Um broto desenvolvese em um carvalho, enquanto um esporo de musgo, mesmo que ambos cresam lado a lado na mesma floresta. As duas plantas que resultam desses processos desenvolvimentais assemelham-se a seus genitores e diferem uma da outra, muito embora tenham acesso mesma restrita gama de materiais do meio. Determinao ambiental: considere duas gmeas monozigticas (idnticas), os produtos de nico ovcito fertilizado que se dividiu e produziu duas crianas completas com genes idnticos. Suponha que as gmeas nasceram na Inglaterra, mas foram separadas ao nascimento e levadas para pases diferentes. Se uma criada na China, ela ir falar chins, enquanto a irm em Budapeste falara Hngaro. Cada uma absorver os valores culturais de seu ambiente. Embora as gmeas comecem a vida com propriedades genticas idnticas, os ambientes culturais diferentes nos quais vivem produziro diferenas entre elas (e diferenas de seus genitores). Obviamente as diferenas, nesse caso, so devidas ao ambiente e os efeitos genticos no tem importncia na determinao das diferenas. medida que um organismo se transforma, no desenvolvimento de um estgio para o outro, seu genes interagem com seu ambiente em cada
momento de sua histria de vida. A interao de genes e ambiente determina o que os organismos so. Gentipo e Fentipo Um organismo tpico assemelha-se a seus genitores mais do que se assemelha a membros de sua espcie as quais no aparentado. Assim , geralmente falamos como se as prprias caractersticas individuais fossem herdadas.desenvolve-se por meio de longas sequncias de eventos nas historias de vida das pessoas afetadas, e tanto os genes quanto o ambiente tem papeis nessas sequncias. No sentido biolgico , um organismo herda apenas estruturas moleculares do zigoto do qual se desenvolve. Os organismos seus genes, no os produtos finais de suas historias de desenvolvimento.
Cap. 2 HERANA MONOGNICA
Existem vrios tipos de enfoques analticos para a descoberta do gene, mas um dos mais amplamente usados baseado na deteco de padres de herana monognica, o tpico desse captulo. Tais padres de herana podem ser reconhecidos na prole de alguns tipos de reprodues controladas, que os geneticista chamam de cruzamentos. Os componentes centrais desse tipo de anlise so os mutantes, organismos individuais de alguma forma alterada da sua propriedade normal. A forma normal de qualquer propriedade de um organismo chamado de tipo selvagem, que encontrada na natureza. Os padres de herana monognica soa teis para a descoberta do gene no s em gentica experimental de organismos-modelo, mas tambm em gentica aplicada. Um exemplo importante encontrado na gentica humana
As regras de herana monognica foram originalmente elucidadas nos anos 1860 pelo monge Gregor Mendel. A anlise de Mendel o prottipo do enfoque experimental da descoberta monognica ainda usado hoje.
Genes e Cromossomos Um conjunto nico e completo de informao gentica (DNA) de um organismo chamado de seu genoma. Nos eucariontes, a maior parte do DNA de um genoma encontrada no ncleo de cada clula. Esse DNA nuclear dividido em unidades chamadas cromossomos. O conjunto de cromossomos presentes no organismo da mesma espcie tem um nmero caracterstico de cromossomo e aspecto. Em ncleo diploide, os dois membros de um par de cromossomos so chamados de cromossomos homlogos ou, as vezes , apenas homlogos. As sequncias de DNA de membros de um par de homlogos, geralmente so iguais, muito embora pequenas variaes de sequncia em geral estejam presentes, o que a base da variao gentica dentro de uma espcie, o tipo de variao de que nos permite distinguir uma da outra. Como os cromossomos homlogos so idnticos, eles levam os mesmos genes nas mesmas posies relativas. Assim, nos diploides, cada gene esta presente como um par de genes. A cromatina varia em compactao ao longo de um cromossomo. Consequentemente, os corantes que reagem, com o DNA produzem intensidades diferentes de colorao ao longo de um cromossomo. A cromatina densa , geralmente, encontrada ao redor do centrmero, uma parte de um cromossomo em geral visvel como uma constrio. A cromatina densa chamada de heterocromatina, e a menos densa de eucromatina.
Bases cromossmicas A viso de Mendel da segregao igual era que os membros de um par de genes segregam-se igualmente na formao de gametas. Ele no conhecia os
eventos subcelulares que ocorrem quando as clulas de dividem no curso da formao de gametas. Hoje em dia, compreendemos que os pares de genes esto situados em pares de cromossomos, e so os membros de um par de cromossomos que, na verdade, segregam, levando os genes consigo. Os membros de um par de genes so segregados como uma consequncia inevitvel. Quando as clulas se dividem, tambm se dividem o ncleo e seus contedos, os cromossomos. Para entender a segregao gnica, devemos primeiro
compreender e contrastar os dois tipos de divises nucleares que ocorrem nas clulas eucariticas. Quando as clulas somticas se dividem para aumentar seu nmero, a diviso nuclear acompanhante chamada de mitose, um estgio programado de todos os ciclos de diviso celular. A mitose pode ocorrer em clulas diploides e haploides. Antes da meiose, como na mitose, a replicao cromossmica ocorre para formar cromtides-irms, que se tornam visveis na meiose. O centrmero parece no se dividir nesse estgio, enquanto o faz na mitose.
Herana monognica em haploides Vimos que na base celular da lei da segregao igual a segregao de cromossomos na primeira diviso da meiose. Na discusso at agora, h evidncia da segregao igual de alelos nos meicitos tanto de plantas quanto de animais e indireta, com base na observao de que os cruzamentos apresentam as propores apropriadas da prole esperada sob segregao igual. Reconhece que os gametas nesses estudos devem ter surgido de muitos diferentes meicitos. Entretanto, em alguns organismos haploides, tais como vrias espcies de fungo e algas, a segregao igual pode ser observada diretamente dentro de um meicitos individual. O motivo que, nos ciclos desses organismos, os quatro produtos de uma nica meiose so temporariamente mantidos juntos em um tipo de saco.
A natureza dos alelos e seus produtos:
Usamos o conceito de alelos sem defini-los em nvel molecular. Os alelos mutantes podes ser usados para estudar a herana monognica sem a necessidade de compreender sua natureza estrutural ou funcional. Entretanto, ultimamente, como um motivo primrio para usar uma herana monognica para investigar a funo de um gene, devemos analisar a natureza molecular dos alelos tipo selvagem e mutante tanto em nveis estruturais quanto funcionais. A maioria das mutaes altera a sequncia de aminocidos do produto proteico do gene, resultando em funo reduzida ou ausente.
Dominncia e recessividade Com uma compresso de como os genes funcionam atravs de seus produtos proteicos, podemos compreender melhor a dominncia e recessividade. Assim, formalmente, o fentipo que dominante ou recessivo, mas, na prtica, os geneticistas aplicam o termo mais frequentemente aos alelos. Essa definio formal no tem contedo molecular, mas tanto a dominncia quanto a recessividade podem ter explicaes simples a nvel molecular. A recessividade observada nas mutaes nos genes que so funcionalmente haplossuficientes. Embora uma nica clula diploide normalmente tenha duas cpias tipo selvagem de um gene, uma cpia de um gene haplossuficientes fornece produto gentico suficiente em geral, uma protena para efetuar as reaes normais da clula. Em um heterozigoto, a cpia restante codificada pelo alelo + fornece produto proteico para a funo normal. Outros genes so haploinsuficientes. Em tais casos, um alelo mutante nulo ser dominante, porque em um heterozigoto o nico alelo tipo selvagem no pode dar produto suficiente para a funo normal.
Gentica direta
Em geral, o tipo de enfoque para a descoberta de gene que temos seguindo , s vezes, chamando de gentica direta, um enfoque para a compreenso da funo biolgica que comea com mutantes monognicos aleatrios e termina a anlise bioqumica e celular detalhada deles, em geral incluindo a anlise genmica. Gentica Reversa, em resumo, ela comea com uma anlise genmica para identificar um conjunto de genes como candidatos para codificar a propriedade biolgica de interesse, e ento, inclui mutantes direcionados especificamente para esses genes, examinando os fentipos mutantes para ver se eles apresentam a propriedade em estudo.
Padres de herana monognica ligadas ao sexo Os cromossomos que analisamos at agora so autossomos, os cromossomos regulares que formam a maioria do conjunto genmico. Entretanto, muitos animais e plantas tm um par especial de cromossomos associados ao sexo. Os cromossomos sexuais tambm segregam igualmente, mas as propores fenotpicas vistas na prole so, em geral, diferentes das propores autossmicas.
Cromossomos sexuais A maioria dos animais e muitas plantas apresentam dimorfismo sexual; em outras palavras, os indivduos so masculinos ou femininos. Na maioria desses casos, o sexo determinado por um par especial de cromossomos sexuais. Vejamos os humanos como um exemplo. As clulas do corpo humano tem 46 cromossomos: 22 pares de homlogos autossomos mais 2 cromossomos sexuais idnticos chamados de cromossomos X. os homens tem um par no idntico no-idnticos, consistindo em um X e um Y. As mulheres tm um par de cromossomos sexuais idnticos, chamados de cromossomo X. o cromossomo Y consideravelmente menor que o X. assim podemos , se representarmos os cromossomos autossmicos por A, podemos escrever: mulheres= 44A+XX; homens=44A+XY.
Na meiose nas mulheres, os dois cromossomos X do par segregam como os autossomos, e cada ovcito recebe um cromossomo X. Assim, com relao aos cromossomos sexuais, os gametas so apenas de um tipo e a mulher dita como sendo sexo homogamtico. Na meiose nos homens, os cromossomos X eY fazem par por uma regio curta, que garante que o X e Y se separem de modo que h dois tipos de espermatozoides, metade comum X e a outra com um Y, portanto o homem chamado de sexo heterogamtico.
Anlise de Heredogramas As reprodues humanas, como a dos organismos experimentais, fornecem muitos exemplos de herana monognica. Entretanto, os cruzamentos de experimentais controlados no podem ser feitos em humanos, e, assim, os geneticistas tm de recorrer aos registros mdicos na esperana de que as reprodues informativas tenham ocorrido (tais como reproduo monobridas) e possam ser usadas para deduzir herana monognica. Tal levantamento dos registros de reprodues chamado de anlise de heredogramas. Para ver a herana monognica, os padres no heredograma tem que ser interpretado de acordo com a lei de Mendel da segregao igual, mas os humanos geralmente tem poucos filhos, e assim, devido a essa prole pequena, as propores esperadas de 3:1 e 1:1 geralmente no so vistas, a menos que muitos heredogramas similares sejam combinados.
Distrbio autossmicos recessivos O fentipo afetado por um distrbio autossmico recessivo herdado como um alelo recessivo. Assim, o fentipo correspondente no-afetado deve ser herdado como o alelo dominante correspondente.
Distrbio autossmico dominantes
Que padres de heredogramas so esperados de distrbio autossmicos dominantes? Aqui o alelo normal recessivo, e o alelo defeituoso dominante. Pode ser paradoxal que um distrbio raro possa ser dominante, mas lembre que a dominncia e a recessividade so simples propriedades de alelos que atuam nos heterozigotos e no so definidos em relao o quanto eles so comuns na populao.
Distrbios recessivos ligado ao X Talvez o exemplo mais familiar de herana recessiva ligados ao X o daltonismo. As pessoas nessas condies so incapazes de distinguir o vermelho do verde. Os genes para a viso em cores foram caracterizados em nvel molecular. A viso em cores baseada em trs tipos diferentes de clulas-cone na retina, cada uma sensvel aos cumprimentos de onda vermelho, verde ou azul. Os determinantes genticos para as clulas-cone vermelha e verde esto no cromossomo X. As pessoas com daltonismo tm uma mutao em um desse dois genes. Como em qualquer distrbio ligados ao X, existem muito mais homens com o fentipo do que mulheres.
Cap. 2 DISTRIBUIO INDEPENDENTE DOS GENES
Fazer um gentipo verdadeiramente superior, combinando tais alelos em uma linhagem, claramente desejvel. Para obter tal combinao as linhagens mutantes devem ser entrecruzadas duas de cada vez. Podemos reconhecer a distribuio independente e como o princpio da distribuio independente pode ser usado na construo de uma linhagem, tanto na agricultura quanto na pesquisa gentica bsica.
Lei de Mendel da distribuio independente
Em grande parte de seu trabalho original com ervilhas, Mendel analisou os descendentes de linhagem pura que diferiram em duas caractersticas. O seguinte simbolismo geral usado para representar gentipos que incluem dois genes. Se os dois genes esto em cromossomos diferentes os pares de genes so separados por um ponto e vrgula - por exemplo: A/a; B/b. Se esto no mesmo cromossomo, os alelos em um homologo so escritos adjacentemente, sem pontuao,sendo separados dos do outro homologo por uma barra por exemplo: AB/ab ou Ab/ aB. No existe um simbolismo aceito para situaes nas quais no se sabe se os genes esto no mesmo cromossomo ou em cromossomos diferentes.
Sntese de linhagens puras As linhagens puras esto entre as ferramentas essenciais da gentica. Por outro lado, apenas essas linhagens totalmente homozigotas expressaro alelos recessivos, mas a principal necessidade de linhagens puras a manuteno de estoques para pesquisa. Os membros de uma linhagem pura podem ser entrecruzados com o tempo e, assim, agir como uma fonte constante de gentipo para uso em experimentos. Assim, para a maioria dos organismos-modelo, existem centros de estoques internacionais que so repositrios de linhagens puras. Centros similares de estoque fornecem linhagens de planta e animais para uso na agricultura.
Vigor Hbrido Considerando-se a sntese de linhagens puras superiores para a pesquisa e para a agricultura. As linhagens puras so convenientes, pois a propagao do gentipo de ano a ano bastante fcil. Entretanto uma grande proporo de sementes comerciais que os fazendeiros usam, so chamadas de semente hbrida. Curiosamente, em muitos casos nos quais duas linhagens diferentes de planta so unidas em um hibrido F1 . Essa superioridade geral de mltiplos
heterozigotos chamada de vigor hbrido. Os motivos moleculares para o vigor hbrido so praticamente desconhecidos e ainda intensamente debatidos, mas o fenmeno inegvel e tem feito grandes contribuies para a agricultura. Um aspecto de usar hbrido que, a cada estao, as duas linhagens parentais devem ser cultivadas separadamente, e ento, entrecruzadas para fazer a semente hibrida para venda. Esse processo muito mais inconveniente do que manter linhagens puras, que necessitam apenas deixar que as plantas se auto-fecundem; consequentemente, a semente hbrida mais cara do que as de linhagem puras.
Base cromossmica da produo independente Com a segregao igual, a distribuio independente de pares de genes em cromossomos diferentes explicada pelo comportamento dos
cromossomos durante a meiose. Considere um cromossomo, que poderemos chamar de 1; seus dois homlogos podem ser chamados de 1 e 1. Se os cromossomos se alinham no equador, ento 1 pode ir para o norte e 1 para o sul, ou vice-versa. Similarmente, para o cromossomo 2 como 2 homlogos 2 e 2, 2 pode ir para o norte e 2 para o sul, ou vice e versa. Assim, o cromossomo 2 ou 2, dependendo de quais cromossom os foram levados na mesma direo.
Distribuio independente em organismos diploides A base cromossmica da lei da distribuio independente formalmente diagramada, que ilustra como o comportamento separado de dois pares de cromossomos diferentes d a origem a propores mendelianos de 1:1:1:1 dos tipos gamticos esperados pela distribuio independente. A clula hipottica tem quarto cromossomos longos homlogos e um par de cromossomos homlogos pequenos. O gentipo dos meicito A/a; B/b e dois pare de allicos A/a e B/b, so mostrados em dois pares de cromossomos diferentes.
Distribuio independentes em organismos haplides Nos fungos ascomicetos, podemos inspecionar os produtos de um nico meicito para mostrar diretamente a distribuio independente. Vamos usar o fungo filamentoso neurospora crassa para ilustrar esse ponto. O neurospora crassa misturando duas linhagens parentais haploides de tipo reprodutivo oposto. Em um modo similar ao de leveduras, o tipo reprodutivo determinado por dois alelos de um gene , nessa espcie, chamados de MAT-A e MAT-a.
Herana Polignica At agora, nossa anlise enfocou diferenas monognicas, com o uso de fentipos bem contrastantes, tais como ptalas vermelhas versus brancas, sementes lisa versus rugosas. Entretanto uma grande proporo de variao em populaes naturais tem forma de variao continua que tipicamente encontrada em caractersticas que podem ter algum valor mensurvel de dois extremos. Altura, peso e intensidade de cor so exemplos de tais caractersticas mtricas ou quantitativas. Tipicamente quando o valor mtrico dessas caractersticas plotado contra a frequncia de uma populao natural. A curva de distribuio tem forma de um sino devida ao fato de que os valores mdios soa os mais comuns, enquanto os valores extremos so raros.
Genes de Organelas At agora consideramos apenas genes nucleares. Embora o ncleo contenha a maior parte dos genes de um organismo eucaritico, um subgrupo distinto e especializado do genoma encontrado nas
mitocndrias, e, nas plantas, tambm nos cloroplastos. Esse subgrupos so herdados independemente do genoma nuclear, e assim eles consistem um
caso especial de herana independente, s vezes chamada de herana extra nuclear. As mitocndrias e cloroplastos so organelas especializadas situadas no citoplasma. Contm pequenos cromossomos circulares que levam a um subgrupo definido do genoma celular total. Os genes mitocondriais esto envolvidos na tarefa mitocondrial de produo de energia, enquanto os genes de cloroplasto so necessrios para o cloroplasto desempenhar sua funo de fotossntese. Entretanto, nenhuma das organelas
funcionalmente autnoma, pois cada uma baseada em grande parte dos genes nucleares para sua funo. Padres de herana em organelas Os genes de organelas apresentam seu prprio modo especial de herana chamado herana uni-parental: a prole herda genes de organela exclusivamente de um genitor; mas no de outro. Na maioria dos casos esse genitor a me, um padro chamado de herana materna. Porque s a me? A resposta est no fato de os cromossomos de organelas estarem situados no citoplasma e os gametas femininos no contribuem como o citoplasma igualmente para o zigoto. Com relao aos genes nucleares, ambos os genitores contribuem igualmente para o zigoto. Entretanto o ovcito contribui com a maior parte do citoplasma, enquanto o espermatozoide no contribui com praticamente nada. Assim como as organelas esto no citoplasma, o genitor feminino contribui com as organelas ao longo do citoplasma, e essencialmente nada do DNA da organela no zigoto vem do genitor masculino.
Segregao Citoplasmtica Em alguns casos, as clulas contm misturas de organelas mutantes e normais. Essas clulas so chamadas de citohets ou heretoplasmons. Nessas misturas, um tipo de segregao citoplasmtica pode ser detectada, na qual os dois tipos se distribuem em clulas-filhas diferentes O processo baseado na repartio aleatria no curso da diviso celular. As plantas
fornecem um bom exemplo. Muitos casos de folhas brancas so causados por mutaes nos genes de cloroplastos que controlam a fila necessria produo e deposio do pigmento verde clorofila. Como a clorofila necessria para a planta viver; esse tipo de mutao letal, e as plantas de folhas brancas no podem ser obtidas de cruzamentos experimentais. Entretanto, algumas plantas variegadas tendo como manchas verdes quanto brancas. Assim as plantas variegadas fornecem um meio para demonstrar a segregao citoplasmtica.
Mutaes citoplasmticas em humanos Existem mutaes citoplasmticas em humanos. Alguns heredogramas humanos apresentam a transmisso de raros distrbios atravs de mulheres e nunca atravs de homens. Esse padro sugere fortemente a herana citoplasmtica e indica uma mutao no mtDNa como o motivo para o fentipo. A doena MERRF (epilepsia miclnica e fibras vermelhas anfractuadas) um desses fentipos, resultando da mudana de uma nica base no mtDNA. uma doena que afeta os msculos, mas os sintomas tambm incluem distrbios oculares e auditivos. Em alguns casos, as clulas de um paciente contm misturas de cromossomos normais e mutantes, e as propores de cada passada pra a prole podem variar como resultado de segregao citoplasmtica. As propores de uma pessoa tambm podem variar em tecidos deferentes ou com o tempo. O acmulo de alguns tipos de mutaes mitocondriais, com o tempo, tem sido proposto como uma possvel causa de envelhecimento.
Cap. 4 Mapeamento de Cromossomos Eucariticos p Recombinao
A obteno do mapa das posies dos genes nos cromossomos uma tarefa que ocupou milhares de geneticistas nos ltimos 80 anos. Existem vrios motivos:
1. A posio do gene uma informao crucial necessria para construir gentipos complexos necessrios para fins experimentais ou aplicaes comerciais. 2. Conhecer a posio ocupada por um gene fornece um modo de enfocar sua estrutura e funo. A posio de um gene pode ser usada para defini-lo em nvel de DNA. Por sua vez, a sequncia de DNA de um gene tipo selvagem ou seu alelo mutante uma parte necessria para deduzir sua funo subjacente. 3. Os genes presentes e sua posio nos cromossomos so, em geral, ligeiramente diferentes em espcies correlatas.
Diagnsticos de ligao Os mapas de recombinao dos cromossomos so geralmente montados com dois ou trs genes de cada vez, com o uso de um mtodo chamado anlise de ligao. Quando os geneticistas dizem que dois genes so ligados, eles querem dizer que os loci desse genes esto no mesmo cromossomo , e assim , os alelos de qualquer homlogo esto fisicamente unidos, pelo DNA entre eles. O modo pelo qual os primeiros geneticistas deduziram a ligao um meio til de introduzir a maioria das ideias principais e procedimentos na anlise.
Crossing Over A ideia de que os recombinantes so produzidos por um tipo de troca material entre os cromossomos homlogos era tentadora. Mas era necessrio experimentao para testar essa hiptese. Uma primeira etapa era encontrar um caso no qual a troca de partes entre os cromossomos seria visvel ao microscpio. O que podemos dizer sobre mecanismo molecular de troca cromossmica em crossing? A resposta simples que o crossing resulta da quebra e unio de DNA. Duas quebras cromossmicas parentais ocorrem na mesma posio, e ento cada pedao junta-se ao pedao vizinho de outro cromossomo.
Qualquer par de cromossomos homlogos, duas, trs ou quatro cromtides podem tomar partes em eventos de crossing em nico meicito.
Mapeamento por frequncia de recombinao A frequncia de recombinates produzidas pelo crossing over a chave do mapeamento cromossmico. A anlise de ttrades de fungos, mostra que para qualquer dois genes ligados especficos, os crossings ocorrem entre ele em alguns meicitos, mas no em todos. Quanto mais distantes os genes estiverem, mais provavelmente ocorrer um crossing e maior a proporo de produtos recombinantes. Assim a proporo de recombinantes a chave para a distncia que separa dois loci genticas em mapa cromossmico. O mtodo bsico do mapeamento de genes com o uso de frequncias recombinantes foi desenvolvido por um estudante de Morgan. medida que Morgan estudava mais os genes ligados, ele percebeu que a proporo de prole recombinante variava consideravelmente, dependendo de quais genes ligados estavam sendo estudados e ele pensou que tal variao na frequncia de recombinantes podia indicar, de algum modo, as distncias reais que separam os genes nos cromossomos. Morgan atribuiu a qualificao desse processo a um estudante de graduao. Alfred Sturtevant, que tambm se tornou um grande geneticista. Morgan pediu a Sturtevant que tentasse dar alguns sentidos aos dados de crossing over entre genes ligados diferentes. O nvel seguinte de complexidade um cruzamento de um tribrido (tripo heterozigoto) com um testador triplo recessivo. Esse tipo de cruzamento de um tribrido com um testador triplo recessivo. Esse tipo de cruzamento, chamado de cruzamento teste de trs pontos, comumente usado na anlise de ligao. A meta deduzir se os trs genes esto ligados e, se estiverem, deduzir sua origem e suas distncias de mapas entre eles.
Mapeamento com marcadores moleculares
A variao fentipa baseada em alelos uma parte da variao geral entre membros individuais de uma populao. Um exame das sequncias de DNA de indivduos diferentes de uma espcie revela uma variao considervel. As variaes moleculares so to comuns que muitos indivduos so heterozigotos moleculares em vrios loci diferentes no genoma. Embora os marcadores moleculares de mapeamento com o uso dos que so efetivamente cruzamentos testes constituam o tipo mais simples de anlise informativa, muitos cruzamentos envolvendo marcadores moleculares no so cruzamentos testes. Entretanto, como cada alelo molecular, tem sua prpria assinatura, detectvel mesmo em heterozigotos, tais cruzamentos, em geral, so informativos porque permitem a deteco de recombinantes e no recombinantes. Loci de qualquer heterozigotose de DNA podem ser mapeados e usados como marcadores cromossmicos moleculares ou marcos.
Poliformismos de comprimento de sequncia simples Uma das surpresas da anlise genmica molecular que a maioria dos genomas contm uma grande quantidade de DNA repetitivo. Alm disso, existem muitos tipos de DNA repetitivo. Em uma ponta do espectro esto repeties mltiplas adjacentes de curtas sequncias simples de DNA. A origem dessas repeties no est clara, mas a caracterstica que a torna til que, em indivduos diferentes, existem geralmente nmeros diferentes de cpias. Assim, essas repeties so chamadas de poliformismos de comprimento de consequncia simples (SSLP). Elas tambm so chamadas, s vezes, repeties em tendem de nmero varivel ou VNTR.
Mapeamento De Centrmero Com Ttrades Lineares Os centrmeros no so genes, mas sim regies do DNA da qual a reproduo ordenada do organismo vivos dependem absolutamente, e, portanto, so de
grande interesse em gentica. Na maioria dos eucariontes, a anlise de recombinao pode ser usada para mapear loci de centrmeros, pois eles no apresentam heterozigose que lhes permita serem usados como marcadores. Entretanto, nos fungos que produzem ttrades lineares, os centrmeros podem ser mapeados. Usaremos o fungo Neurospora como um exemplo. Lembre-se que o fungo Neurospora (um haploide), as divises meiticas ocorrem ao longo do eixo do asco, e assim cada meicito produz um arranjo linear de oito ascsporos, chamado de octade. Esses oito ascsporos constituem os quatros produtos da meiose (uma ttrade) mais uma meiose pos mittica.
Uso de Valores de Lod para Avaliar a Ligao em Heredogramas Humanos Os humanos tm milhares de fentipos heradados autossomicamente, e o mapeamento dos loci dos genes que causam esses fentipos pode parecer uma simples questo de usar recombinao. Entretanto, o progresso no mapeamento desses loci foi inicialmente lento por vrios motivos. Primeiro, os cruzamentos controlados no podem ser feitos com seres humanos, e o geneticistas tiveram que tentar calcular frequncias de recombinantes em dibridos e ocasionais que foram produzidos ao acaso em reprodues humanas. As reprodues que eram equivalentes a cruzamentos-teste eram extremamente raras. Segundo, os seres humanos em geral produzem apenas um pequeno nmero de prole, dificultando a obteno de dados suficientes para calcular distncias de mapa estatisticamente confiveis.
Computo de Crossings Mltiplos No-vistos Na discusso de cruzamento-teste de trs pontos, algumas cromtides parentais resultam de crossing duplos. Esses crossings, inicialmente, no podiam ser contados na frequncia de recombinantes, desviando os resultados. Essa situao levou a uma noo preocupante que todas as distncias de mapa baseadas em frequncia de recombinantes podem ser subestimativas de distncias fsicas porque os crossing mltiplos no-
detectados podem ter ocorrido, alguns de cujos produtos no seriam recombinantes. Vrios enfoques matemticos criativos foram criados para tornar o problema de crossing mltiplo.
Funo de mapa O enfoque desenvolvido por Haldane foi criar uma funo de mapa, uma frmula que relaciona um valor observado de frequncia recombinante como uma distncia de mapa corrigido para mltiplos crossings. O enfoque funciona relacionando FR ao nmero mdio de crossings, m, que deve ocorrido ter nesse segmento cromossmico por meiose e ento deduzindo que a distncia de mapa esse valor m deve ter produzido. Para encontrarmos a relao de FR com m, devemos primeiro pensar no resultado das varia possibilidades de crossing. Em qualquer regio
cromossmica, devemos esperar meioses com zero, um, dois, trs, quatro ou mais crossings.
O uso de mapas baseados em recombinao em conjunto com Mapas Fsicos Os mapas de recombinao tm sido o principal tpico. Eles mostram os loci de genes para os quais alelos mutantes foram encontrados. As posies desse loci em um mapa so determinados com base na frequncia de recombinantes na meiose. A frequncia de recombinantes considerada proporcional a distancia que separa os dois loci no cromossomo. Assim, a frequncia de recombinantes torna-se a unidade de mapeamento. Tal mapeamento, baseado em recombinao de genes com fentipos mutantes conhecidos, vem sendo feito h quase um sculo. Vimos como os stios de heterozigose molecular tambm podem ser incorporados em tais mapas de recombinao. Como qualquer stio heterozigoto, esses marcadores moleculares so mapeados por recombinao e, ento, usados para navegar por um gene de interesse biolgico. Fazemos a suposio perfeitamente razovel de que um mapa de
recombinao representa o arranjo dos genes nos cromossomos, mas, como j foi dito, esses mapa so construes realmente hipotticas. Em contraste os mapas fsicos so o mais prximo do mapa genmico real que a cincia pode ter.
Cap. 5 A Gentica de Bactrias e seus Vrus
Muito embora a gentica de bactrias tenha tornado possvel a gentica molecular, esta nunca foi a meta das pesquisas. As bactrias so biologicamente importantes por si mesmas. So organismos numerosos em nosso planeta. Contribuem para reciclar nutrientes tais como nitrognio, enxofre e carbono nos ecossistemas. Algumas so agentes de doenas humanas, animais e de plantas. Outras vivem simbioticamente dentro de nossas bocas e intestinos. Alm disso muitos tipos de bactrias so teis para a sntese industrial de uma ampla gama de produtos orgnicos. Assim, o mpeto de disseco gentica das bactrias tem sido o mesmo que de organismos multicelulares, para compreender seu funcionamento biolgico. As bactrias pertencem a uma classe de organismos conhecidos como procariontes, que tambm incluem as algas cianofceas. Uma caracterstica fundamental para definir os procariontes que seu DNA no circundado dentro de um ncleo delimitado por uma membrana. Como outros organismos, as bactrias tm genes compostos de DNA disposto em uma srie de um cromossomo. Entretanto, a organizao de seu material gentico nica sob vrios aspectos. O genoma na maioria das bactrias uma molcula de DNA bifilamentar sob a forma de um crculo fechado. Alm disso, as bactrias na natureza em geral contm elementos extras de DNA chamados de plasmdeos. A maioria dos plasmdeos tambm so crculos de DNA, mas so muito menores que o genoma bacteriano principal.
Trabalhando com microorganismos As bactrias dividem-se rapidamente e ocupam pouco espao; logo, so convenientes para usar como organismos modelo. Elas podem ser cultivadas em meio lquido ou em uma superfcie slida tal como gel de gar, desde que os nutrientes bsicos sejam fornecidos. Cada bactria divide-se assexualmente em 1 2 4 8 16 clulas, e assim por diante, at que os
nutrientes sejam esgotados ou ate que os produtos txicos se acumulem em nveis que param o crescimento da populao. Uma pequena quantidade de cultura lquida pode ser pipetada em uma placa de Petri contendo meio slido de gar e espalhar-se igualmente pela superfcie com um espalhador estril, em um processo chamado de plaqueamento. As clulas dividem-se, mas, como no podem viajar pela superfcie do gel, todas as clulas ficam juntas em um aglomerado. Quando essa massa atinge mais de 107 clulas, ela se torna visvel a olho nu como uma colnia. Cada colnia distinta em uma placa foi derivada de uma nica clula original. Os membros de uma colnia que tem um nico ancestral genticos so conhecidos como um clone celular.
Conjugao Bacteriana Os primeiros estudos de gentica bacteriana revelaram o processo inesperado de conjugao bacteriana. As bactrias possuem processos similares reproduo sexual e
recombinao? A pergunta foi respondida pelo trabalho experimental elegante de Joshua Lederbergue e Edward Tatum, que, em 1946, descobriram o processo tipo sexual no que se tornou o principal modelo para a gentica de bactrias. Eles estavam estudando duas linhagens de E. coli com conjuntos diferentes de mutaes auxotrficas. A linhagem A s cresceria se o meio fosse suplementado com metionina e biotina; a linhagem B so cresceria se fosse suplementada com treonina, leucina e tiamina.
Mapeamento de cromossomos bacterianos Mapeamento cromossmico de ampla escala usando o tempo de entrada Wollman e Jacob perceberam que a construo de mapas de ligao pelos resultados de reproduo interrompida seria fcil usando, como medida de distncia, os tempos nos quais os alelos doadores primeiro apareciam aps a reproduo. As unidades de distncia de mapas nesse caso so minutos. Assim, se b+ comea a entrar na clula F- 10 min aps a+ comea a entrar , ento a+ e b- distam 10 unidades. Como os mapas eucariticos baseados em crossings, esses mapas de ligao foram originalmente puras construes genticas. Na poca em que eles foram originalmente criados, no havia modo de testar sua base fsica. Plasmdeo F que levam fragmento genmicos O fator F em linhagens Hfr geralmente bem estvel em sua posio de insero. Entretanto, ocasionalmente um fator F claramente sai d um cromossomo por uma reverso do processo de recombinao que o inseriu em primeiro lugar. As duas regies homlogas de pareamento de ambos os lados fazem novo pareamento, e ocorre um crossing over pra liberar o plasmdeo F. Entretanto, s vezes a sada no limpa, e o plasmdeo leva com ele uma parte do cromossomo bacteriano. Um plasmdeo F levando o DNA genmico bacteriano chamado de plasmdeo F (F primo).
Plasmdeo R Uma alarmante propriedade das bactrias patognicas, primeiro veio a luz ps estudos em hospitais japoneses no ano 1950. Disenteria bacteriana causada por bactrias do gnero shigella. Essa bactria foi inicialmente sensvel a uma ampla gama de antibiticos que foram usados para controlar a doena. Nos hospitais japoneses, entretanto, shigella isoladas de pacientes com disenteria mostrou-se simultaneamente resistente a muitas dessa drogas, incluindo
penicilina, tetraclicina, sulfanilamida, estreptomicina e cloranfenicol. Essa
resistncia a mltiplas drogas foi herdada como nico pacote gentico, e pode ser transmitido de modo infeccioso, no s para outras linhagens sensveis a shigella, mas tambm para outras espcies de bactrias. Esse talento que se assemelha mobilidade do plasmdeo F de E.coli, extraordinariamente til para as bactrias patognicas porque a resistncia pode se expandir rapidamente por uma populao. Entretanto, suas implicaes para a cincia medica so terrveis porque a doena bacteriana subitamente se torna resistente ao tratamento por uma grande gama de drogas.
Transformao bacteriana Algumas bactrias podem captar fragmento de DNA do meio externo, e tal captao constitui outro modo pelo qual as bactrias podem trocar seus genes. A fonte do DNA podem ser outras clulas da mesma espcie. Em alguns casos, o DNA foi liberado de clulas mortas; em outros casos, o DNA foi secretado de clulas bacterianas vivas. O DNA captado integra-se ao cromossomo receptor. Se o DNA de um gentipo diferente do da receptora, o gentipo da receptora pode tornar-se permanentemente alterado, um processo adequadamente chamado de transformao.
Mapeamento cromossmico usando transformao A transformao pode ser usada para medir o quanto dois genes ligados esto prximos em um cromossomo bacteriano. Quando o DNA extrado para experimentos de transformao, inevitvel alguma quebra de pedaos menores. Se dois genes doadores esto situados muito prximos no cromossomo, h uma boa chance de, eles serem levados no mesmo pedao do DNA transformante. Assim ambos sero captados, causando uma transformao dupla. Infeco de bactrias por fagos A maioria das bactrias so suscetveis ao ataque por bacterifagos. Um fago consiste em um cromossomo de acido nucleico (DNA ou RNA)
circulando por uma capa de molculas de protena. Os tipos de fagos so identificados no por nome de espcies, mas por smbolos. Durante a infeco um fago liga-se a uma bactria e injeta seu material gentico no citoplasma bacteriano, como diagramado. A informao gentica do fago ento comanda a maquinaria da clula bacteriana desligando a sntese dos componentes bacteriano se redirecionando a maquinaria de sntese de componentes da bactria para fazer componentes do fago.as cabeas de fagos recm formadas so individualmente recheadas por replicas do cromossomo fago. Finalmente so feitos muitos descendentes do fago e so liberados quando a parede celular da bactria rompida. Esse processo de abertura chamado de lise. A populao de prole de fagos chamada de lisado de fagos.
Transduo Alguns fagos so capazes de captar genes bacterianos e lev-los de uma bactria para a outra: um processo conhecido como transduo. Assim a transduo unta-ser como a bactria de modos de transferir material genmico entre bactrias, juntamente com a transferncia de cromossomo Hfr, transferncia de plasmdeo F e transformao.
Mapas fsicos e Mapas de ligao comparados Alguns mapas cromossmicos, muito detalhados, de bactrias foram obtidos combinando as tcnicas de mapeamento de reproduo
interrompida, mapeamento de recombinao, transformao de transduo. Hoje em dia, novos marcadores genticos so tipicamente mapeados primeiro em segmento com cerca de 10 a 15 min. de mapa usando a reproduo interrompida. Marcadores adicionais aproximadamente ligados podem ser mapeados em uma anlise escala fina com o uso de cotransduo de P1 ou recombinao.
Cap. 6 Interao Gentica
No nvel molecular, os organismos operam mais ou menos do mesmo modo que uma linha de montagem em uma fbrica, como uma srie de vias qumicas nas quais cada etapa controlada por um produto gnico. Existem milhares de tais vias, de tamanhos diferentes e conectados de vrios modos em redes ou sistemas. Os genes que interagem so membros da mesma via ou de uma via conectada. Para fins de discusso, podemos distinguir vrios tipos importantes de vias, essas vias se superpem vrios graus. Primeiro, existem vias puramente biossintticas, nas quais a molcula essencial produzida em uma srie de interconvees catalisadas por enzimas codificadas por genes. Segundo, existem vias de transduo de sinal que transmitem instrues de um sinal extracelular, como substncia ambiental ou um hormnio de outra parte do corpo. Esse sinal ativa uma protena celular que, por sua vez, ativa outra, e assim por diante em uma cascata de ativao, a qual geralmente termina na ativao de uma protena reguladora que liga e desliga a transcrio de conjunto de genes. Terceiro, existem vias desenvolvimentais, um conjunto diverso de processos, muitos dos quais so controlados por genes, que promovem o crescimento e diferenciao do corpo de um organismo. Quando ns deduzimos tais vias por meio de pesquisas, chegamos perto de compreender o subgrupo de genoma que controla a propriedade que nos interessa. Interao de alelos de um nico gene: variao de dominncia
Existem milhares de modos diferentes de altera a sequncia de um gene, produzindo uma mutao, embora alguns desses alelos mutantes paream em uma populao real. Os mutantes conhecidos de um gene so chamados de alelos mltiplos ou uma srie allica.
Dominncia completa e recessividade O tipo mais simples de dominncia a dominncia total ou completa. Um alelo completamente dominante ser expresso quando apenas uma cpia est presente, como no heterozigoto, enquanto o alelo alternativo ser totalmente recessivo. Na dominncia completa, o homozigoto dominante no pode ser distinto do heterozigoto, isto , em nvel fenotpico, A/A=A/a. a feniceltonuria (PKU), e muitas outras doena humanas monognicas, so totalmente recessivas, enquanto os alelos tipo selvagem so dominantes, embora, nesses casos, o alelo tipo selvagem seja recessivo.
Alelos Letais Recessivos Um alelo que capaz de causar a morte de um organismo chamado de alelo letal. Na caracterizao de um conjunto de alelos mutantes recm descobertos, uma mutao recessiva , s vezes, descoberta como letal. Essa informao potencialmente til, pois mostra que o gene recm descoberto essencial para a operao do organismo. De fato, com o uso da tecnologia do DNA, um alelo mutante nulo de um gene de interesse, agora, pode ser feito intencionalmente, e tornado homozigoto pra ver se letal, e sob quais condies ambientais. Os alelos letais tambm so teis para determinar o estgio desenvolvimental no qual o gene normalmente atua. Nesse caso, os geneticista investigam se a morte ocorreu por alelo mutante letal cedo ou tarde no desenvolvimento de um zigoto. O fentipo associado morte tambm pode ser informativo com relao funo do gene; por exemplo, se certo rgo parece ser anormal, o gene provavelmente se expressa nesse rgo.
Interao gnica em outros tipos de vias A noo de que os genes interagem por meio de vias poderosa e tem aplicao em todos os organismos. A via de arginia de Neutospora um exemplo de uma via sinttica, uma cadeia de converses enzimticas que produz nutrientes essenciais. Podemos ampliar essa ideia nivelmente para outro caso j introduzido, a doena fenilceturia (PKU), que causada por um alelo autossmico recessivo. A Pku resulta da incapacidade de converter fenilalalnina em tirosina. Consequentemente, a fenilalanina acumula-se e , espontaneamente, convertida em um composto txico, o cido fenilperuvico. O gene de PKU parte de uma via sinttica, como a via de arginina de Neuspora.
Interaes gnicas interferentes Como efetuar uma anlise gentica que revela os genes interagem para contribuir com uma propriedade biolgica determinada? Resumidamente, o enfoque o seguinte: 1. Tratar clulas com agentes causadores de mutao, tais como radiao ultravioleta. Esse tratamento produz um grande conjunto de mutantes com uma expresso anormal da propriedade de estudo. Cada mutante verificado pra confirmar a herana monognica. avaliado o efeito fenotpico de cada mutante no desenvolvimento da propriedade. 2. Testar esses mutantes para determinar o nmero total de loci gnicos que tomam parte e quais mutaes so alelos do mesmo gene. 3. Combine as mutaes par a par fazendo cruzamentos para formar mutantes duplos para ver se eles interagem. A interao gnica deduzida do fentipo do mutante duplo, essencialmente qualquer desvio da combinao simples de fentipos monognicos mutantes. Se os alelos mutantes interagem no podemos deduzir que os genes tipo selvagem tambm interagem normalmente.
Penetrncia e Expressividade A anlise de herana monognica, h uma tendncia natural para escolher mutantes que produzem claras propores mendelianas. Em tais casos, podemos usar o fentipo para distinguir os gentipos mutantes e tipo selvagem com quase 100% de certeza. Nesses casos, dizemos que a mutao 100% penetrante no fentipo. Entretanto, muitas mutaes mostram penetrncia incompleta, isto , nem todos os indivduos com o gentipo expressam o fentipo correspondente. Assim, a penetrncia definida como a percentagem de indivduos com determinao alelo que exibem o fentipo associado a esse alelo.
Cap. 7 DNA: Estrutura e Replicao
A histria comea na primeira metade do sculo XX, quando o resultado de vrios experimentos levou os cientista a conclurem que o DNA o material gentico, no outra molcula biolgica tal como um carboidrato, protenas ou lipdeos. O DNA uma molcula simples feita apenas de quatro blocos estruturais (os quatro nucleotdeos). Assim, foi necessrio pesquisar como essa molcula to simples podia ser o mapa para a incrvel diversidade para os organismos na terra. O modelo de dupla hlice proposto por Watson e Crick foi feito com base nos resultados de cientistas anteriores a ele. Eles basearam em descobertas anteriores da composio qumica do DNA e propores de sua base. Alm disso, imagem de difrao de raios X revelaram ao olho treinado que o DNA uma hlice de dimenses precisas. Watson e Crick concluram que o DNA uma dupla hlice composta de duas cromtides de nucleotdeos ligados que se enrolam uma na outra.
A estrutura proposta do material hereditrio imediatamente sugeriu como ele poderia servir como um mapa, e como esse mapa poderia ser transmitido ao longo das geraes. A estrutura do DNA
Mesmo antes da estrutura do DNA ser elucidada, os estudos genticos indicam que o material gentico deve ter trs propriedades principais: 1. Como essencialmente cada clula de um corpo de um organismo tem a mesma constituio gentica, a replicao fiel do material gentico em cada diviso celular crucial. Assim, as caractersticas estruturais do DNA devem permitir uma replicao fiel. 2. Como ele deve codificar uma constelao de protenas expressas por um organismo, o material gentico deve ter contedo informacional. Como a informao codificada no DNA decifrada para produzir protenas. 3. Como as alteraes hereditrias, chamadas mutaes, fornecem a matria prima para a seleo evolutiva, o material gentico deve ser capaz de mudar em raras ocasies. Entretanto, a estrutura do DNA deve ser estvel de modo que os organismos possam se basear em sua informao codificada.
Replicao Semiconservativa O mecanismo de cpia, ao qual Watson e Crick se referiram, chamado de Replicao Semiconservativa. Os arcabouos acar fosfato so
representados por fitas espessas, e a sequncia de pares de bases aleatria. Imaginemos que a dupla hlice anloga a um zper que se abre, comeando em uma ponta. Poderemos ver que essa analogia com o zper valida, deselicoidizao dos dois filamentos ira expor as bases em cada filamento. Cada base exposta tem o potencial para parear os nucleotdeos livres na soluo. Como a estrutura do DNA impe requisitos rgidos de pareamento, cada base exposta ir parear apenas com sua base complementar, A com T e
G com C. assim, cada uma dos filamentos ir agir como um molde, para dirigir para a montagem de bases complementares para reestruturar uma dupla hlice idntica original. Os nucleotdeos recm adicionados so supostos como vindos de um conjunto de nucleotdeos livres que devem estar presentes na clula.
DNA polimerase Um problema confrontado pelos cientistas foi compreender como essas bases so trazidas para a dupla hlice molde. Embora os cientistas suspeitem que as enzimas tenham um papel, essa possibilidade no foi provada at 1959, quando Artur Kornberg isolou o DNA polimerase de E.coli e demonstrou essa atividade enzimtica in vitro. Essa enzima desoxirribonucleotdeos ponta 3 de uma cadeia crescente de nucleotdeos, usando como molde um nico filamento de DNA que foi exposto pela deselicoidizao localizada da dupla hlice. Os substratos para a DNA polimerase so as formas trifosfato de desoxirribonucleotdeos, dATP, dGTP, dCTP e d TTP.
Telmeros e Telomerase: Trmino da Replicao A replicao da molcula linear de DNA em um cromossomo eucaritico ocorre em ambas as direes a partir de vria origens de replicao. Esse processo replica a maioria do DNA cromossmico, mas h um problema inerente em replicar as duas pontas das molculas lineares de DNA, as regies chamadas de Telmeros. A sntese continua do filamento leading pode ocorrer at a ponta do molde. Entretanto, a sntese do filamento descontnuo (lagging) requer primeiro frente do processo; logo, quando o ltimo primer removido, resta uma ponta unifilamentar em uma molcula-filha de DNA. Se o cromossomo filho com essa molcula de DNA fosse replicado novamente, o filamento faltando sequncias na ponta seria uma molcula bifilamentar encurtada aps a replicao. A cada ciclo subsequente de replicao, o Telmero continuaria a se encurtar, at que informaes codificantes essenciais fossem perdidas.
Cap. 8 Transcrio e Processamento
Com a liberao da primeira sequncia de rascunho, foi anunciado um vencedor. Surpreendentemente, o que entrou com a baixa estimativa, 25.947 genes, foi declarado o vencedor. Como pode o Homo Sapiens, com seu crebro complexo e sofisticado sistema imune, ter apenas o dobro de genes que um verme nematelminto e o mesmo nmero e genes que o primeiro genoma de plantas sequenciado, a praga Aradobdopsis Thaliana? A resposta a essa pergunta tem a ver com a marcante descoberta feita no final dos anos 70. Nessa poca, as protenas de organismos superiores foram descobertas como sendo codificadas no DNA, no como trechos contnuos, mas em pedaos. Assim os genes de eucariontes superiores so geralmente compostos de pedaos chamados xons, que codificam partes das protenas, e pedaos chamados ntrons, que separam os xons. Uma cpia de RNA contendo tantos xons quanto ntrons sintetizada de um gene. Uma mquina biolgica (spliceossomo) remove os ntrons que une os xons (recomposio do RNA) para produzir um RNA final que contm a informao contnua e necessria para sintetizar uma protena.
Propriedades do RNA O RNA geralmente uma cadeia de nucleotdeos unifilamentares, no uma dupla hlice como o DNA. Uma consequncia que o RNA mais flexvel e pode formar uma variedade muito maior de formas moleculares tridimensionais complexas do que pode o DNA bifilamentar. Um filamento de RNA pode se dobrar de tal modo que algumas de suas prprias bases podem fazer par com
outras. Tal pareamento intramolecular de bases um determinante importante da forma do RNA. O RNA tem acar ribose em seus nucleotdeos, em vez da desoxirribose encontrada no DNA. Como os nomes sugerem, os dois acares diferem na presena ou ausncia de apenas um tomo de oxignio. O acocar do RNA contm um grupo de hidroxila (OH) ligado ao tomo de carbono 2, enquanto o acar do DNA tem apenas um tomo de hidrognio ligado ao tomo de carbono 2. Os nucleotdeos de RNA contem bases adenina, guanina e citosina, mas a base pirimidnica (U) esta presente no lugar de timina. O RNA como as protenas, mas no so como o DNA, pode catalisar reaes biolgicas. O nome ribozima foi criado para molculas de RNA que funcionam como enzimas proteicas.
Classes de RNA Os RNA podem ser agrupados em duas classes gerais. Uma classe de RNA codifica a informao necessria para fazer cadeias peptdicas (protenas). Chamamos essa classe de RNA mensageiro(mRNA), porque como um
mensageiro, esse RNA servem como intermedirios e passam a informao do DNA para a protena. Chamamos as outras classes de RNA funcionais porque o RNA no codifica a informao para fazer protenas. Em vez disso, o prprio RNA o produto funcional final. Como o RNA uma molcula to verstil, ele pode atender muitos papeis funcionais e participar em uma variedade de processos celulares. RNA mensageiro: as etapas pelas quais um gene influencia o fentipo so chamados de expressa gnica. Para a grande maioria dos genes, o RNA transcrito apenas um intermedirio necessrio para a sntese de uma protena, que o produto final funcional que influencia o fentipo. RNA funcional: medida que sabemos mais sobre os detalhes ntimos da expressa gnica e regulao, torna-se aparente que os RNA funcionais caem
em uma variedade de classes e tem papeis diversos. Novamente, importante enfatizar que os RNA funcionais so ativos como RNA; eles nunca so traduzidos em polipeptdeos. Duas classes de RNA funcionais so encontradas tanto em procariontes quanto eucariontes: RNA transportadores e RNA ribossmicos. RNA transportador (tRNA): so molculas responsveis por levar o aminocido correto para o mRNA no processo de traduo. RNA ribossmico (rRNA): so molculas dos principais componentes ribossomos, que so grandes mquinas macromoleculares que guiam a montagem da cadeia de aminocidos pelo mRNA e rRNA.
Transcrio A primeira etapa na transferncia da informao do gene da protena produzir um filamento de RNA cuja sequncia de bases de um segmento de DNA, s vezes seguido de modificao desse RNA para prepar-lo para seus papeis celulares especficos. Assim, o RNA produzido por um processo que cpia a sequncia de nucleotdeos do DNA. Como esse processo reminiscente da transcrio (cpia) de palavras escritas, a sntese do RNA chamada de transcrio. O DNA dito sendo transcrito em RNA, e o RNA chamado de transcrito.
Estgios da transcrio A sequncia codificante de protena em um gene um segmento relativamente pequeno do DNA inserido em uma molcula de DNA muito longa (o cromossomo). Como seguimento apropriado transcrito em uma molcula unifilamentar de RNA de tamanho e sequncia de nucleotdeos corretos? Como o DNA de um cromossomo uma unidade contnua, a maquinaria transcricional deve ser dirigida para o comeo de um gene para comear a transcrever no local certo, continuar transcrevendo ao longo do gene, e finalmente parar de transcrever na outra ponta. Esses trs estgios distintos da transcrio so chamados de iniciao, alongamento e trmino. Embora
esse processo geral de transcrio seja, marcantemente, similar, em procariontes e eucariontes existem diferenas importantes.
Cap. 9 Protenas e sua Sntese
O uso excessivo de antibiticos em todo o mundo provocou a evoluo de linhagens bacterianas resistentes. Por exemplo, a cada ano mais de 2 milhes de pacientes em hospitais nos USA contraem uma infeco que resistente a antibiticos, e 90.000 morrem como resultado. Como a resistncia
desenvolveu-se to rapidamente? Para responder essa pergunta os cientistas tm enfocado a maquinaria celular que o alvo dos antibiticos. Mais de metade de todos os antibiticos em uso tm o alvo ribossomo bacteriano, o stio da sntese de protenas nos procariontes. Embora os ribossomos dos procariontes e eucariontes sejam muitos similares, ainda existem pequenas diferenas. Devido a essas diferenas, antibiticos so capazes de ter como alvo ribossomos bacterianos, mas deixar intocveis os ribossomos
eucariticos. Usando a cristolografia de raio X, os cientistas tiveram sucesso em visualizar antibiticos ligados ao ribossomo. Por esses estudos, eles determinaram que as mutaes no rRNA bacteriano e/ ou protenas ribossmicas so responsveis pela resistncia antibitica. Com esse conhecimento dos pontos de contato entre alguns antibiticos e o ribossomo, os projetistas de drogas esto tentando criar uma nova gerao de antibiticos que, por exemplo, sejam capazes de se ligar a vrios stios vizinhos. A lgica da tal projeto de droga que a resistncia seria mais durvel porque precisaria da ocorrncia de duas mutaes, o que um evento muito improvvel, mesmo para uma bactria.
Estrutura das Protenas Quando o transcrito primrio foi totalmente processado em uma molcula final mRNA, pode ocorrer a traduo em protena. Antes de considerarmos
como as protenas so feitas, precisamos compreender a estrutura da protena. As protenas so os principais determinantes da forma e funo biolgica dessas molculas influenciam muito a forma, cor, tamanho, comportamento e fisiologia dos organismos. Como os genes funcionam codificando protenas, a compreenso da natureza das protenas essencial pra a compreenso da ao gnica. Uma protena um polmero composto de monmeros chamados de aminocidos. Em outras palavras, uma protena uma cadeia de aminocidos. Como os aminocidos j foram chamados de peptdeos, a cadeia s vezes chamada de polipeptdeo.
O cdigo gentico Se os genes so segmentos de DNA se um filamento de DNA apenas uma fileira de nucleotdeos, deve ditar de algum modo a sequncia de aminocidos nas protenas. A simples lgica nos diz que se um nucleotdeo so letras em um cdigo, ento uma combinao de letras pode formar palavras representando aminocidos diferentes. Primeiro, devemos perguntar como o cdigo lido. Ento, devemos perguntar quantas letras do mRNA constituem uma palavra, o cdon, e que cdon ou cdons representam cada aminocido.
Nmeros de letras no cdon Se uma molcula de mRNA lida de uma ponta para a outra, apenas uma das quatros bases diferentes , A, U, G ou C, pode ser encontrada em cada posio. Assim, se as palavras que codificam aminocidos tivessem uma letra de tamanho, apenas quatro palavras seriam possveis. Esse vocabulrio no pode ser o cdigo gentico, pois precisamos ter uma palavra para cada aminocido comumente encontrado na protenas celulares. Se as palavras tivessem duas letras de tamanho, ento 4x4=16
palavras seriam possveis; por exemplo: AU, CU ou CC. Esse vocabulrio ainda no grande o suficiente. Se as palavras tivessem trs letras de tamanho, seriam possveis 4x4x4=64 palavras. Por exemplo: AUU, GCG e UCG. Esse vocabulrio fornece palavras mais do que suficientes pra descrever aminocidos. Podemos concluir que a palavra cdigo deve se constituir em pelo menos trs nucleotdeos. Entretanto, se todas as palavras so trincas, ento as palavras possveis so em considervel excesso s 20 necessrias para nomear os aminocidos comuns.
tRNA: o Adaptador Aps tornar-se conhecido que uma sequncia de aminocidos de uma protena foi terminada pelos cdons de trinca no mRNA, os cientistas comearam a imaginar como era feita essa determinao. Um modelo inicial, rapidamente descartado como muito simples e improvvel, props que os cdons do mRNA poderiam dobrar-se e formar 20 cavidades distintas que se ligam diretamente a aminocido na ordem correta. Os aminocidos de fato esto ligados a um adaptador. Cada aminocido torna-se ligado a um rRNA especfico, que ento leva esse aminocido para o ribossomo, o complexo molecular que ir ligar o aminocido a um polipeptdeo crescente.
Ribossomos A sntese de protenas ocorre quando o tRNA e as molculas de mRNA se associam aos ribossomos. A tarefa dos tRNA e ribossomo traduzir a sequncia de cdons de nucleotdeos em mRNA em uma sequncia de aminocidos na protena. O replissomo, por exemplo, uma mquina biolgica que pode replicar o DNA com preciso e velocidade. O stio da sntese de protenas, o ribossomo, muito maior e mais complexo do que as mquinas descritas at agora. Sua complexidade devida ao fato de
que ele desempenha vrias tarefas com preciso e velocidade. Por esse motivo, melhor pensar no ribossomo como uma fbrica contendo muitas mquinas que atuam em conjunto. Vejamos como essa mquina organizada para desempenhar suas vrias funes.
Cap. 10 Regulao da Expressa Gnica em Bactrias e seus Vrus
A respeito da simplicidade da forma, as bactrias tm em comum com membros maiores e complexos de outros reinos a tarefa fundamental de regular a expresso de seus genes. Um dos principais motivos que elas so oportunistas nutricionais. Considere como as bactrias obtm muitos compostos importantes, tais como acares, aminocidos e nucleotdeos, necessrios ao metabolismo. As bactrias nadam em um mar de potenciais nutrientes. Elas podem adquirir os compostos que precisam do ambiente, ou produzi-los pela vias enzimticas. A sntese das enzimas necessrias para essas vias gasta energia e recursos da clula; assim, dependendo da escolha, as bactrias obtero compostos do ambiente. Para serem econmicas, elas produziro as enzimas necessrias para elaborar esses compostos apenas quando no h outra opo, ou seja, quando esses compostos no esto disponveis em seu ambiente local. As bactrias desenvolveram sistemas reguladores que acoplam a expresso dos produtos gnicos a sistemas sensores que detectam o composto relevante a um ambiente bacteriano local. A regulao das enzimas que tomam parede no metabolismo de acar so um exemplo. As molculas de acar podem ser oxidadas para produzir energia ou podem ser usadas como blocos estruturais de uma grande gama de compostos orgnicos. Entretanto, existem muitos tipos diferentes de acares que a bactria pode usar, incluindo a lactose, glicose, galactose e xilose. Uma importante protena diferente necessria para permitir que cada um dos acares entre na clula. Alm disso, um conjunto diferente de enzimas necessrio para processar cada um dos acares.
Elas devem ser capazes de reconhecer condies ambientais nas quais elas devem ativar ou reprimir a transcrio de genes relevantes. Elas devem tambm, ser capazes de ligar ou desligar, como um interruptor, a transcrio de cada gene especfico ou grupo de genes.
As bases da Regulao Transcricional Procaritica: interruptores genticos A regulao da transcrio depende regularmente de dois tipos de interao protena-DNA. Ambas ocorrem perto do stio no qual comea a transcrio gnica. Uma dessas interaes DNA-protena determina onde comea a transcrio. O DNA que participa dessa interao o segmento de DNA chamado de promotor, e a protena que se liga a esse sitio a RNA
polimerase. Quando o RNA polimerase se liga ao DNA promotor, transcrio pode comear algumas bases distantes do stio promotor ou ser transcrito. Outro tipo de interao DNA protena decide se a transcrio ativada pelo promotor ocorre. Os segmentos de DNA, perto do promotor, servem como pontos de ligao para protenas reguladoras especficas de sequncias chamadas ativadores e repressores. Nas bactrias, a maioria dos stios de ligao para os repressores so chamados de operadores.
Os genes estruturais lac O metabolismo de lactose requer duas enzimas: (1) uma permease, para transportar lactose para a clula, e (2) -galactosidase, para quebrar a molcula de lactose para produzir glicose e galactose. As estruturas da galactosidase e permease so codificadas por suas sequncias adjacentes, Z e Y, respectivamente. Uma terceira sequncia contgua codifica uma enzima adicional, chamada transacetilase, que no necessria para o metabolismo de lactose. Chamaremos os genes Z,Y, e A de genes estruturais em outras palavras, segmentos codificantes de protenas.
Genes controlados juntos Quando Jacob e Monod induziram a -galactosidase, eles descobriram que tambm induziram a enzima permease, que necessria para o transporte de lactose para a clula. A anlise de mutantes indica que cada enzima era codificada por um gene diferente. A enzima transacelilase tambm era induzida junto com -galactosidase e permease, e tambm mostrada depois sendo codificada por um gene separado. }Portanto, Jacob & Monod puderam identificar trs genes controlados coordenadamente. O mapeamento de recombinao mostrou que os genes Z, Y e A eram proximamente ligados no cromossomo.
Mutaes Polares Algumas das mutaes que foram mapeadas nos genes Z e Y foram vistas sendo polares, isto , afetam genes posteriores ao operon. Por exemplo, as mutaes polares Z resultam em um funcionamento nulo no s de z, mas tambm de Y e A. as mutaes polares de Y tambm afetaram A, mas no Z. essa mutaes polares forma observaes genticas que sugeriram a Jacob & Monod que os trs genes eram transcritos de uma ponta como uma unidade. As mutaes polares resultaram de cdons de fim que faziam com que os ribossomos sassem do transcrito. Isso deixava um trecho nu de mRNA que era degradado, inativando assim os genes posteriores.
Represso Catablica do Operon Lac. Controle Positivo O sistema Lac existente aquele que, por um longo processo evolutivo, foi selecionado para operar de modo timo para a eficincia de emergncia da clula bacteriana. Supostamente, para maximizar a eficincia energtica, duas condies ambientais devem ser satisfeitas para que as enzimas do metabolismo de lactose sejam expressas. Uma condio que uma lactose deve estar presente no ambiente. Essa condio faz sentido, pois seria ineficiente para a clula produzir as
enzimas metablicas de lactose se no houvesse substrato a ser metabolizado. J vimos que o reconhecimento da clula da presena de lactose feito por uma protena repressora.
Controle duplo positivo e negativo Como no sistema lac, o controle da transcrio em bactrias no nem puramente positivo, nem puramente negativo; tanto na regulao positiva quanto a negativa podem controlar perons individuais. A regulao perons de arabinose fornece um exemplo no qual uma nica protena de ligao ao DNA pode agir como um repressor ou um ativador, uma variao com tema geral de regulao transcricional por protenas de ligao ao DNA.
Ciclos de vida bacterifagos: mais reguladores perons complexos O bacterifago um chamado fago temperado que tem dois ciclos de vida alternativos. Quando uma bactria normal infectada por um afago tipo selvagem, pode ocorrer dois resultados possveis: (1) o fago pode se replicar e, eventualmente, lisar a clula (o ciclo ltico) ou (2) o genoma do fago pode se integrar ao cromossomo da bactria como um profago inerte (ciclo lisognico). No estado ltico, a maioria dos 71 genes do fago so expressos em algum ponto, enquanto, no estado lisgenico a maioria dos genes esto inativos.
Anatomia molecular da mudana gentica Para entendermos como a deciso executada em nvel molecular,
vejamos as atividades do repressor e Cro. O operador O R fica entre os dois genes que codificam essas protenas e contem trs stios, O R1, OR2 e OR3, que superpe dois promotores postos PR, que promove a transcrio
de genes lticos, e PRM, que dirige a transcrio do gene cI. Os trs stios operadores so similares, mas no idnticos em sequncia e, embora Cro e o repressor possam, cada um, se ligar a um dos operadores, eles fazem isso com afinidades diferentes: o repressor liga-se a OR1 com maior afinidade, enquanto Cro liga-se a OR3 com a mais alta afinidade. A ocupao do repressor de OR1 bloqueia a transcrio de PR e, assim bloqueia a transcrio dos genes do ciclo ltico. A ocupao de Cro de O R3 bloqueia a transcrio de PRM, que bloqueia a manuteno de transcrio de cI, e, assim , permite a transcrio dos genes para o ciclo ltico. A ocupao dos stios operadores determina, portanto, os padres lticos versus lisognicos da expresso do gene .
Fatores Sigma Alternativos regulam grandes conjuntos de genes Algumas respostas fisiolgicas, tais como a formao de esporos em determinada bactrias Gram-positivas, requerem a expresso coordenada de um grande grupo de genes no ligados por todo o genoma para causar marcantes mudanas fisiolgicas e morfolgicas. O processo de
esporulao no bacillus subtilis foi analisado em grande detalhe nas ltimas dcadas. Sob o estresse, a bactria forma esporos que so muito resistentes ao calor e desidratao.
Cap.11 Regulao da Expresso Gnica em Eucariontes
Examinaremos a regulao gnica nos eucariontes, de muitos modos, nossa viso da regulao gnica ser um estudo de contrastes. Nas bactrias, voc aprendeu como as atividades das mudanas genticas em geral eram governadas por protenas ativadoras ou repressoras nicas, e como o controle de grupos de genes foi obtido por sua organizao em perons ou pela atividade de fatores especficos. As expectativas iniciais eram que a expresso gnica eucaritica seria regulada por meios
similares. Nos eucariontes, entretanto, a maioria dos genes no so encontrados em perons. Alm disso, veremos que as protenas e as sequncias de DNA que participam da regulao gnica eucaritica so mais numerosas. Em geral, muitas protenas de ligao do DNA atuam em uma nica mudana, com muitos interruptores separados por gene, e as sequncias regulatrias desses interruptores quase sempre esto situadas longe dos promotores. Uma importante diferena adicional entre bactrias e eucariontes que o acesso aos promotores gnicos eucariticos restrito pela cromatina. A regulao gnica em eucariontes requer a atividade de grandes complexos de protena que promovem ou restringem o acesso a promotores gnicos pela RNA polimerase.
A protena Gal4 tem domnio de ligao ao DNA e ativao separveis Aps Gal4 estar ligado ao elemento UAS, como induzida a expresso de gene? Um domnio distinto da protena Gal4, domnio da ativao, necessrio para a atividade reguladora. Assim, a protena Gal4 tem pelo menos dois domnios: um para a ligao ao DNA e outro para ativao da transcrio. Uma organizao modular similar foi encontrada como sendo uma caracterstica comum tambm de outros fatores de transcrio de ligao ao DNA. A organizao modular da protena Gal4 foi demonstrada em uma srie de experimentos simples e elegantes. A estratgia foi testar a ligao ao DNA e a ativao gnica de formas mutantes da protena nas quais partes haviam sido deletadas ou fundidas a outras protenas. Desse modo, podia ser determinado se uma parte da protena era necessria para determinada funo. Para fazer esses estudos, os experimentadores precisaram de um meio simples de dosar a expresso enzimas codificadas pelos genes GAl. A expresso dos genes Gal e de outros alvos de fatores de transcrio tipicamente monitorada usando um gene reprter cuja expresso facilmente rastreada. Em geral, o gene reprter o gene lacZ de E.coli, que
pode atuar em substratos cujos os produtos so facilmente avaliados por sua cor brilhante ou fluorescncia. Outro gene reprter comum o gene que codifica a protena fluorescente verde da gua-viva, que, como o nome sugere, facilmente rastreada pela luz que ela emite. A regio codificante de um desses genes reprter e um promotor so colocados posteriormente a um elemento UAS do gene Gal. A expresso reprter ento lida da atividade Gal. nas clulas.
Cromatina dinmica e regulao gnica eucaritica Um segundo mecanismo que influencia a transcrio gnica em
eucariontes modifica a estrutura local da cromatina ao redor de sequncias reguladoras gnicas. Para compreender totalmente como funciona esse mecanismo, precisamos primeiro rever a estrutura da cromatina e, ento, considerar como ela pode mudar e como suas mudanas afetam a expresso gnica. O recrutamento da maquinaria trasncricional por ativadores pode parecer um tanto similar em eucariontes e bactrias, sendo a principal diferena o nmero de protenas que interagem na maquinaria transcricional. De fato, h menos de uma dcada, muitos bilogos interpretavam a regulao eucaritica, simplesmente, como uma verso bioquimicamente mais complicada do que havia sido descoberto em bactrias. Entretanto, essa viso mudou acentuadamente medida que os bilogos consideraram o efeito de organizao do DNA genmico nos eucariontes.
Mecanismo de ao acentuadora O desenvolvimento de um organismo complexo requer que os nveis de transcrio sejam regulados em uma ampla gama. Pense no mecanismo de regulao como sendo um reostato em vez de um interruptor. Nos eucariontes, os nveis de transcrio so finalmente ajustveis agrupando os stios de ligao em acentuadores. Vrios fatores de transcrio diferentes ou varia molculas do mesmo fator de transcrio possa se ligar
a stios adjacentes. A ligao desses fatores de stios que esto separados pela distncia correta leva a um efeito amplificado, ou superaditivo, em ativar a transcrio. Quando um efeito maior que o aditivo, ele chamado de sinergstico.
Imprinting genmico O fenmeno imprinting genmico foi descoberto, h quase 20 anos, em mamferos. No imprinting genmico, alguns genes autossmicos tem padres incomuns de herana. Por exemplo, um alelo igf2 expresso em um camundongo apenas se for herdado do pai do camundongo, um exemplo de imprinting materno porque uma cpia de gene derivada da me inativa. Contrariamente um alelo h19 de camundongo s expresso se for herdado da me. H19 um exemplo de imprinting paterno porque a cpia paterna inativa. A consequncia o imprinting paternal que os genes imprintados so expressos como se fossem a nica cpia do gene presente na clula, muito embora existam duas. Assim, o imprinting um exemplo de herana monoallica. Importante, nenhuma mudana observada nas sequncias de DNA de genes imprintados; isto , o gene idntico pode ser ativo ou inativo na prole, conforme tenha sido herdado da me ou do pai.
Domnios de cromatina e sua herana. At agora, vimos como os genes so ativados em um ambiente de cromatina. Entretanto, como visto no incio deste capitulo, a maioria dos genes eucariticos esto desligados na maior parte do tempo. Vejamos agora essas grandes regies do genoma que so trascricionalmente inativas. Um dos modelos mais teis para compreender os mecanismos que mantm a inatividade dos genes esta relacionado ao controle do tipo reprodutivo em levedura e a mudana do tipo reprodutivo.
Cap.12 O controle gentico do desenvolvimento
Um zigoto transparente medida que se desenvolve, um dos objetos mais fascinantes no mundo dos seres vivos. A contnua mudana de forma que ocorre de uma hora para a outra nos desafia por sua simplicidade. Os padres geomtricos que se apresentam a cada mudana convidam a uma anlise matemtica... Esse aparato um apelo irresistvel aos lados emocional e artstico de nossa natureza. Os principais catalisadores para compreender a formao dos animais de monstros genticos, moscas mutantes com marcantes alteraes de estruturas corpreas. Nos primeiros dias da genticas de Drosophila, mutantes raros surgiram espontaneamente ou como subproduto de outros experimentos com transformaes espetaculares de pares de corpo. Em 1915, Calvin Bridges, ento estudante de Morgan, isolou usam mosca com mutao que fazia com que pequenos halteres das asas da mosca-da-fruta lembrassem as grandes asas anteriores. Ele chamou o mutante de bithorax. A transformao em mutantes bithorax chamada de hometica, porque parte do corpo transformada para se assemelhar outra. Subsequentemente, muitos outros mutantes hometicos foram identificados em Drosophila, tais como o marcante mutante Antennapedia, no qual pernas desenvolvem-se no lugar de antenas.
O enfoque gentico do desenvolvimento Por muitas dcadas, o estudo do desenvolvimento embrionrio envolveu a manipulao fisiolgica dos embries, clulas e tecidos. Foram estabelecidos vrios conceitos principais sobre as propriedades de embries em
desenvolvimento por meio de experimentos nos quais,por exemplo, uma parte de um embrio de anfbio em desenvolvimento para outra parte em um embrio receptor foi mostrado induzindo o tecido circulante para formar uma segundo eixo corpreo completo. Similarmente o transplante da parte superior de um membro de galinha em desenvolvimento para a anterior poderia induzir dedos extras, mas com polaridade revertida com relao aos dedos normais. Essas
regies transplantadas do embrio de anfbios e broto de embrio de galinha foram chamados de organizadores devido a sua marcante habilidade em organizar o desenvolvimento dos tecidos vizinhos. As clulas nos
organizadores foram postuladas produzindo morfgenos, molculas que induzem vrias respostas no tecido vizinho de um modo dependente da concentrao.
Classificao dos genes pela funo desenvovimental Umas das primeiras tarefas aps a execuo de uma triagem gentica para mutaes separar aquelas de interesse. Muitas mutaes so letais quando hemi ou homozigotas porque as clulas no podem sobreviver sem os produtos afetados por essa mutao. As mutaes mais interessantes so as que causam algum pequeno defeito no padro corpreo embrionrio e/ou do adulto. Demonstrou-se til agrupar os genes afetados por mutaes em vrias categorias com base na natureza de seus fentipos mutantes. Muitos genes ferramentais podem ser classificados de acordo com sua funo em controlar a identidade de partes do corpo, a formao de partes do corpo, a formao de tipos de clulas e organizao dos eixos corpreos primrios.
Definio de todas as ferramentas Os genes Hox so talvez os membros mais bem conhecidos das ferramentas, mas so uma pequena famlia em um grupo muito grande de genes necessrios para o desenvolvimento dos nmeros apropriados, formas,
tamanhos e tipos de partes corpreas. Pouco se sabia sobre os restos das ferramentas ate final de 1970 e incio dos anos 1980, quando Christine Nusselein-Volhard e Eric Wieschaus, que trabalhava no Max Planck Institute em Tubingen, Alemanha, encontraram genes necessrios para a formao da organizao segmentar do embrio e larva de Drosophila.
Regulao Espacial da Expresso Gnica no Desenvolvimento
Vimos que os genes ferramentais so expressos em relao coordenadas no embrio. Mas como as coordenadas espaciais do embrio em desenvolvimento funcionam como instrues para genes, para lig-los ou deslig-los. O controle fisiolgico da expresso gnica em bactrias e eucariontes simples finalmente governado por protenas de ligao e sequncias especficas de ligao do DNA agindo em elementos regulatrios de ao por cis. Similarmente, o controle espacial da expresso gnica durante o
desenvolvimento amplamente governado pela interao de fatores de transcrio com elementos regulatrios de ao cis. Entretanto o controle espacial e temporal da regulao gnica no desenvolvimento de um embrio multicelular requer a ao de mais fatores de transcrio em elementos mais numerosos e mais complexos de ao cis.
Regulao Ps-transcricional da expresso gnica no desenvolvimento Embora a regulao transcricional seja um meio importante de restringir a expresso dos produtos gnicos para definir reas durante o desenvolvimento, no um meio exclusivo de fazer isso. A recombinao alternativa do RNA tambm contribui para a regulao gnica, e assim, e a regulao da traduo do mRNA em protenas e microRNA (miRNA). Em cada caso as sequncias regulatrias no RNA so reconhecidas por fatores de recomposio, protenas de ligao ao mRNA ou miRNA, e controlam a estrutura do produto protico, sua quantidade ou localizao onde a protena produzida.
Desenvolvimento e Doena As descobertas das ferramentas para os desenvolvimento de moscas, vertebrados e humanos tambm teve um profundo efeito no estudos das bases genticas da doenas humanas, particularmente defeitos de nascimento e cncer. Um grande nmero de mutaes de genes ferramentais foram identificadas afetando o desenvolvimento humano e a sade. Enfocaremos aqui uns poucos exemplos que ilustram como a compreenso do
funcionamento gnico e regulao em modelos animais foi traduzida em uma melhor compreenso da Biologia humana.
Polidactilia Uma sndrome relativamente comum em humanos o desenvolvimento de dedos extras, parciais ou completos, nas mos e ps. Essa condio, chamada polidactilia, surge em cerca de 5 a 17 de cada 10.000 nativivos. Nos casos mais marcantes, a condio esta presente tanto nas mos quanto nos ps. A polidactilia ocorre amplamente nos vertebrados, em gatos, galinhas,
camundongos e outras espcies.
Holoprosencefalia As mutaes na regio codificante Shh tambm foram identificadas. As alteraes consequentes na protena Shh esto associadas a uma sndrome chamada Holoprosencefalia, na qual a anormalidades ocorrem no tamanho do crebro, na formao do nariz e em outras estruturas da linha media. Essas anomalias parecem ser contrapartes menos graves dos efeitos
desenvolvimentais observados em camundongos mutantes homozigotos Shh. De fato, as crianas afetadas vistas em clnica so heterozigotas. Uma cpia de um gene normal Shh parece ser insuficiente pra o desenvolvimento da linha mdia normal. Os fetos humanos homozigotos para a perda de funo Shh muito provavelmente morreram na gestao com mais defeitos graves.
Cncer como uma doena no desenvolvimento Nos animais de vida longa, como ns mesmos e outros mamferos, o desenvolvimento no termina ao nascimento ou ao final da adolescncia. Os tecidos e vrios tipos celulares esto constantemente sendo renovados. A manuteno de vrias funes orgnicas depende do crescimento controlado e diferenciao de clulas que substituem as que foram descartadas ou, de outro
modo morreram. A manuteno de tecidos e rgos geralmente controlada por vias de sinalizao. As mutaes herdadas ou espontneas nos genes que codificam os componentes dessa vias podem perturbar a organizao tissular e contribuir para a perda de controle da proliferao celular. Como a proliferao celular descontrolada uma caracterstica do cncer, a formao de cnceres pode ser uma consequncia. O cncer, portanto, uma doena do desenvolvimento, um produto do processo normal de desenvolvimento que deu errado.
Cap.13 Genomas e Genmica
A genmica revolucionou como a anlise feita e abriu caminhos para investigaes que no eram concebveis at alguns anos atrs. A maioria das anlises genticas que consideramos at agora empregam um enfoque direto para anlise gentica e processos biolgicos, isto , a anlise comea primeiro tirando mutantes que afetam algum fentipo observvel, e a caracterizao desses mutantes acaba levando ao nvel de gene e funo do DNA, RNA e sequncias proteicas. Em contraste saber as consequncias inteiras de DA do genoma de um organismo permitiu que os geneticistas atuassem em ambas as direes: direto do fentipo ao gene e, ao reverso, do gene ao fentipo. Sem exceo, as sequncias genmica revelam muitos genes que no eram detectados pela anlise mutacional clssica. Usando a chamada gentica reversa, os geneticistas hoje podem estudar sistematicamente os papeis de tais genes antes no-identificveis. Alm disso, a falta de estudo gentico clssico no mais um impedimento para a investigao dos organismos. As fronteiras da anlise experimental esto indo alm do nmero modesto de organismos modelos longamente explorados. Revoluo genmica Aps o desenvolvimento da tecnologia do DNA recombinante nos anos 70, as pesquisas de laboratrios tipicamente fizeram a clonagem e o
sequnciamento de um gene de cada vez, ento, apenas aps ter encontrado
primeiro algo interessante sobre esse gene a partir de uma anlise mutacional clssica. As etapas consistem no procedimento de ter um mapa gentico clssico de um lcus para isolar o DNA codificante de um gene (clonagem) para determinar
Mapa de sequncia genmica Quando as pessoas encontram um novo territrio, uma de suas primeiras atividades criar um mapa. Essa prtica foi verdadeira para exploradores, gegrafos, oceangrafos e astrnomos, e igualmente verdadeira para geneticistas. Os geneticistas usam muitos mapas para explorar o terreno de um genoma. Os exemplos so mapas de ligao baseados nos padres de herana de genes alelos e mapas citogenticos baseados na localizao de caractersticas microscopicamente visveis, tais como pontos de quebra de rearranjos. O mapa de maior resoluo a sequncia completa do DNA do genoma, isto , a sequncia completa de nucleotdeos A, T, C e G de cada dupla hlice no genoma. Como fazer um mapa completo da sequncia de um genoma um trabalho imenso, nunca visto antes na Biologia, devem ser usadas novas estratgicas, todas baseadas na automao.
Bioinformtica A sequncia genmica um cdigo gentico altamente criptografado contendo a informao para construir e manter um organismo funcional. O estudo do contedo de informao dos genomas chamado de Bioinformtica. Estamos longe de ler essa informao do comeo ao fim do modo como poderamos ler um livro. Muito embora saibamos que trinca codificam os aminocidos nos segmentos codificantes de protenas, na grande parte da informao contida em um genoma no decifrvel por mera inspeo.
Estrutura do genoma humano Ao descrevermos a estrutura geral do genoma humano, devemos primeiro examinar sua estrutura repetida. Uma frao considervel do genoma, cerca de 45%, repetitiva. Grande parte desses DNA repetitivo composto de cpias de elementos de transposio. De fato, mesmo na cpia nica do DNA restante, uma frao tem sequncias sugerindo que podem descender de antigos elementos de transposio que so imveis e acumularam mutaes aleatrias, fazendo com que divirjam em sequncia dos elementos de transposio ancestrais. Assim, grande parte do genoma humano parece composto de caronas genticos.
Genmica funcional e gentica reversa: Os geneticistas tm estudado e expresso e interaes de produtos gnicos nas ltimas dcadas. Entretanto esses estudos foram em pequena escala, considerando apenas um gene ou alguns genes de cada vez. Como o advento da genmica, temos uma oportunidade de expandir esses estudos a um nvel global, usando enfoques de genoma total para estudar a maioria de todos os produtos gnicos sistemtica e simultaneamente. Esse enfoque global para o estudo da funo, expresso e interao dos produtos gnicos chamado de genmica funcional.
Cap. 14 O genoma Dinmico elementos de transposio.
O sequenciamento do DNAa de genomas de uma variedade de micrbios, plantas e animais indica que elementos de transposio existem em praticamente todos os organismos. Surpreendentemente, eles so, de longe, o maior componente do genoma humano, correspondendo a quase 50% de nossos cromossomos. A despeito
de sua abundncia, o papel gentico normal desses elementos conhecidos com certeza.
no so
Em seu estudo, os cientistas so capazes de explorar habilidade de elementos de transposio de se inserirem em novos locais no genoma. Os elementos de transposio construdos no tubo de ensaio so instrumentos valiosos, tanto em procariontes quanto em eucariontes, para o mapeamento gentico, criao de mutantes, clonagem de genes, e mesmo para a produo de organismos transgnicos.
Elementos de Transposio em Procariontes A descoberta gentica dos elementos de transposio levou a muitas dvidas sobre com que tais elementos poderiam parecer no que concerne a sequncia de DNA e como eles eram capazes de mover-se de um local para o outro no genoma. Embora os elementos de transposio tenham sido primeiro descobertos em milho, os primeiros elementos eucariticos a serem caracterizados
molecularmente foram isolados de leveduras mutantes e genes de drosophila. Os elementos transponveis eucariticos enquadram-se em duas classes: classe 1 - de retrotransposons, e DNA classe 2 - de transposons. A primeira classe a ser isolada, os retroposons no eram como elementos IS procariticos e transposons. PRTICA Os elementos genticos tornaram sua presena conhecida quando foram transpostos para um gene ou em stios de quebra cromossmica ou rearranjo. Aps o DNA de elementos de transposio ser isolados de mutaes instveis, os cientistas puderam usar esse DNA como sondas moleculares para determinar se havia mais cpias correlatas no genoma. Em todos os casos, pelo menos vrias cpias do elemento sempre estavam presentes no genoma e, em alguns casos, at muitas centenas.
Cap. 15 Mutao, Reparo e Recombinao
Uma mudana de DNA dentro de um gene (mutao de ponto) geralmente envolve uma ou algumas bases. As substituies de um s par de bases pode criar cdons de sentido trocado ou sem sentido (termino de transcrio). Uma purina substituda por outra purina (ou mais pirimida substituda por outra pirimida) uma transio. Uma purina substituda por pirimida (ou vice e versa) uma transverso. A adio ou deleo de um par de bases (indels) produz mudanas de matriz de leitura. Alguns genes humanos que contem repeties de trinucleotideos, especialmente as que so expressas no tecido neural, tornam-se mutadas pele expanso dessas repeties e podem, assim, causar doenas. A formao de repeties de monocidos dentro dos polipeptdios codificados por esse genes responsvel pelos fentipos mutantes. As mutaes podem ocorrer espontaneamente como subproduto de processos celulares normais, tais como replicao do DNA ou metabolismo, ou podem ser induzidas por radiao mutagnica ou substncias qumicas. Os mutgenos geralmente resultam em um tipo especfico de mudana devido a sua especificidade qumica. Por exemplo, alguns produzem exclusivamente transies G. C A. T; outros, exclusivamente mudanas de matriz de
leitura. Embora as mutaes sejam necessrias para gerar diversidade, muitas mutaes esto associadas a doenas genticas herdadas, tais como xeroderma pigmentoso. Alm disso, as mutaes que ocorrem nas clulas somticas so a fonte de muitos cnceres humanos. Muitas vias biolgicas desenvolveram-se para corrigir o amplo espectro de mutaes espontneas e induzidas. Algumas vias, tais como o reparo por exciso de base e nucleotdeo e o reparo de mau pareamento, usam a informao inerente
complementaridade de base para executar reparo livre de erro. Outras vias que usam polimerase de bypass para corrigir bases danificadas podem induzir erros na sequncia de DNA.
A correo de quebras bilifilamentares particularmente importante porque essas leses podem levar a rearranjos cromossmicos desestabilizantes. A juno de pontas no homlogas uma via que liga pontas quebradas, de modo que a forquilha da replicao parada no resulta da morte celular. Nas clulas em replicao, as quebras bilifilamentares podem ser reparadas de um modo livre de erro pela via de helicoidao de filamento dependente de sntese, que usa a cromtide-irm para reparar a quebra. Centenas de quebras bifilamentares programadas iniciam o crossing mictico no-irms como outras quebras bifilamentares, as quebras micticas devem ser processadas rpida e eficientemente para evitar serias consequncias, tais como morte celular e cncer. Ainda esta sendo estudado como esse reparo feito. Um modelo atual, informado pela segregao de caractersticas na formao de seporos de fungos, cita o uso de cromtides no-irms de cromossomos homlogos como molde para reparar as quebras. De acordo com esse modelo, a maioria das quebras leva a produtos no-crossing e so provavelmente resolvidas por um mecanismo tipo SDSA. Entretanto, as quebras crticas para a formao de quiasmas levam a crossing seguindo a formao e resoluo das junes duplas de Holliday.
Cap. 16 Alteraes Cromossmicas em Larga Escala
A poliploidia uma condio anormal na qual h um nmero maior do que de conjuntos cromossmicos. Os poliploides tais como os triploides (3n) e os tretraploides (4n) so comuns entre plantas, e so representados mesmo entre animais. Os organismos com o nmero mpar de conjunto de cromossomos so estreis porque nem todos os cromossomos tm um parceiro na meiose. Os cromossomos no pareados ligam se aleatoriamente aos plos da clula na meiose, levando a conjuntos desbalanceados dos cromossomos nos gametas resultantes. Tais gametas desbalanceados no produzem prole vivel. Nos poliplides como nmero par de conjuntos, cada cromossomo tem um parceiro potencial de pareamento e, portanto, pode produzir gametas balanceados e
uma prole. A poliploidia pode resulta em um organismo de maiores dimenses; essa descoberta permitiu avanos importantes na horticultura e na criao de cultivos. Nas plantas, os alopoliploides (poliplides formados de combinaes de conjuntos cromossmicos de espcies diferentes) podem ser feitos cruzando duas espcies relacionadas e ento, duplicando os cromossomos da prole com o uso de colchicina ou pela fuso de clulas somticas. Essas tcnicas tm aplicaes potenciais nos cruzamentos de cultivos porque os alopoliploides combinam as caractersticas de duas espcies parentais. Quando acidentes celulares mudam partes de conjuntos cromossmicos, resultam os aneuplodes. A aneuploidia em si em geral, resulta em um gentipo desbalanceado com um fentipo anormal. Os exemplos de aneuploides incluem monossomicos (2n-1) e trissmicos (2n +1). As sndromes de Down (trissomia do 21), de klinelter (XXY) e de Turner (XO) so exemplos bem documentados de condies aneuplodes em humanos. O nvel espontneo de aneuploidia em humanos bem alto, e contribui com uma grande proporo de distrbios geneticamente baseados em populaes humanas. O fentipo de um organismo aneuploidia depende muito do cromossomo particularmente afetado. Em alguns casos, tais como ma trissomia do 21 humana, h uma constelao altamente caracterstica de fentipos associados. A maioria dos casos de aneuploidia resulta de ma segregao cromossmica acidental na meiose (no-disjuno). O erro espontneo e pode ocorrer em qualquer meicito na primeira e segunda diviso. Em humanos, um efeito de idade materna est associado no-disjuno do cromossomo 21, resultando em uma maior incidncia de sndrome de Down nos filhos de mes com mais idade. A outra categoria geral de mutaes cromossmicas compreende rearranjos estruturais, que incluem delees, duplicaes, inverses e translocaes. Essas mudanas resultam ou de quebra de reunio incorreta, ou de crossing over entre elementos repetitivos (recombinao homloga no-allica). Os rearranjos cromossmicos so uma causa importante de problemas de sade em populaes humanas, e so teis em construir linhagens especiais de
organismos para gentica experimental e aplicada. Em organismos com um conjunto cromossmico normal mais um conjunto rearranjado (rearranjos heterozigoto), existem estruturas incomumente pareadas na meiose que resultam de forte afinidade de pareamento de regies cromossmicas homlogas. Por exemplo, as inverses heterozigotas apresentam alas, e translocaes recproca mostram estruturas em forma de cruz. As segregaes dessa estrutura resultam em produtos micticos anormais nicos para o rearranjo. Uma deleo a perda de uma seo do cromossomo, seja devido a quebra cromossmica seguida de uma perda do segmento intercalar, seja devido segregao em translocaes heterozigticas ou inverses. Se a regio removida em uma deleo essencial para a vida, a deleo homozigota letal. As delees heterozigotas podem ser letais devido ao desequilbrio cromossmico ou porque revelam alelos deletrios recessivos, ou podem ser no-letais. Quando uma deleo em um homlogo permite a expresso fenotpica de alelos recessivos no outro, o desmascaramento dos alelos recessivos chamado de pseudodominncia. As duplicaes geralmente so produzidas por outros rearranjos ou por crossing over aberrante. Elas tambm desequilibram o material gentico, produzindo um efeito fenotpico deletrio ou a morte do organismo. Entretanto, as duplicaes podem ser uma fonte de novo material para a evoluo porque pode ser mantida em uma cpia, deixando a outra cpia livre para desenvolver novas funes. Uma inverso um giro de 180 de uma parte de um cromossomo. No estado homozigoto, as inverses podem causar pouco problema para um organismo. Menos que a heterocromatina cause um efeito de posio ou que uma das quebras perturbe um gene. Por outro lado, as inverses heterozigotas apresenta, alas de inverso na meiose, e o crossing over dentro das alas resulta em produtos inviveis. Os produtos de crossing de inverses pericntricas, que envolvem o centrmero, diferem das inverses
paracntricas, que no envolvem, mas ambas apresentam reduzida frequncia
de recombinantes na regio afetada e, geralmente, resultam em reduzida fertilidade. Uma translocao move um segmento cromossmico para outra posio no genoma. Um exemplo simples uma translocao recproca, na qual partes dos cromossomos no homlogos trocam de posies. No estado heterozigoto, as translocaes produzem duplicaes e delees meiticas, que podem levar a zigotos desbalanceados. Podem produzir novas ligaes genticas por translocaoes. A segregao aleatria de centrmeros em, uma translocao heterozigota resulta 50% de produtos micticos desbalanceados e, assim, 50% de esterilidade (semi-esterelidade).
Cap.17 Genticas de populaes
O estudo das mudanas dentro de uma populao, ou gentica de populaes, est relacionado a mudanas herdveis nas populaes ou organismos de processos individuais subjacentes e desenvolvimento. A gentica de
populaes o estudo da variao herdada e sua modificao no tempo e no espao. A variao herdada identificvel dentro das populaes pode ser estudada examinando-se as diferenas em sequncias de aminocidos especficos de protenas, ou mesmo pelo exame, mais recente, de diferenas nas sequncias de nucleotdeos dentro do DNA. Esses tipos de observaes revelaram que h um considervel poliformismo em muitos loci dentro de uma populao. Uma medida dessa variao a quantidade de heterozigose em uma populao. Tipicamente, uma populao polifrmica para 25 a 33% do seus genes codificantes de protenas, e um individuo heterozigoto em cerca de 10% de tais loci. Quaisquer dois humanos diferem em cerca de trs milhes de nucleotdeos. Os estudos populacionais mostraram que, em geral, as diferenas genticas entre indivduos dentro das raas humanas so muito maiores que as diferenas mdias entre as raas. A fonte principal de toda a variao a mutao. Entretanto, dentro de uma populao, a frequncia quantitativa de gentipos especficos pode ser mudada
por recombinao, imigrao de genes, eventos mutacionais continuados e pelo acaso. Uma propriedade da segregao mendeliana que as reprodues aleatrias resultem em uma distribuio de equilbrio gentipos aps uma gerao. Entretanto, se houver endogamia, a variao gentica dentro de uma populao convertida em diferenas entre populaes, tornando cada populao separada homozigota para um alelo escolhido aleatoriamente. Por outro lado, para a maioria das populaes, atingido um balano entre endogamia, mutao de um alelo para autor e imigrao. Um alelo pode aumentar ou diminuir de frequncia dentro de uma populao pela seleo natural de gentipos com probabilidades mais altas de sobrevida e reproduo. Em muitos casos, tais mudanas levam homozigose em determinado lcus. Por outro lado, o heterozigoto pode ser mais adaptvel que ambos os homozigotos, levando a um poliformismo balanceado. Em geral, a variao gentica o resultado da interao de foras. Por exemplo, um mutante deletrio pode nunca ser totalmente eliminado de uma populao, porque a mutao continuara a reintroduzi-lo na populao. A migrao tambm pode reintroduzir alelos que foram eliminados por seleo natural. A menos que alelos alternativos sejam intermedirios em frequncia, a seleo (especialmente contra recessivos) muito lenta, precisando de muitas geraes. Em muitas populaes, especialmente as de pequenos tamanhos , novas mutaes podem tornar-se estabelecidas, muito embora no sejam favorecidas por seleo natural, ou podem ser eliminadas, muito embora
sejam favorecidas, simplesmente por um processo de deriva gentica aleatria.
Cap. 19 Gentica Evolutiva
A teoria darwiniana da evoluo explica as mudanas que ocorrem em populaes de organismos como resultado de mudanas nas frequncias relativas de variantes diferentes na populao. Se no houver variao, dentro de uma espcie para alguma caracterstica, no pode haver evoluo. Alm disso, essa variao deve ser influenciada por diferenas genticas. Se as diferenas no forem herdveis, elas no podem evoluir, pois a reproduo diferencial de variantes diferentes no pode ser levada por geraes de linhagens. Assim, todas as reconstrues evolutivas hipotticas dependem criticamente se as caractersticas em questo so, de fato, herdveis. Os
processos que do origem variao dentro da populao so causalmente independentes dos processos que so responsveis pela reproduo diferencial dos vrios tipos. essa independncia que significante quando se diz que as mutaes so aleatrias. O processo de mutao fornece uma variao no-dirigida, enquanto o processo de seleo natural seleciona essa variao, aumentando frequncia dessas variantes que, por acaso, so mais capazes de sobreviver e se reproduzir. Muitas so colhidas, mas poucas so selecionadas. A divergncia evolutiva das populaes no espao e no tempo no apenas uma consequncia da seleo natural. A seleo natural no um processo globalmente otimizado que encontra os melhores organismos para determinado ambiente. Em vez disso, ela encontra um conjunto de boas solues alternativas para problemas adaptativos, e o resultado particular da evoluo seletiva em determinado caso sujeito a eventos histricos casuais. Fatores aleatrios tais como deriva gentica e ocorrncia casual ou perda de novas mutaes, podem resultar em resultados radicalmente diferentes de um processo evolutivo mesmo quando a fora da seleo natural a mesma. A metfora, geralmente empregada, que h um panorama adaptativo de combinaes genticas, e que a seleo natural leva a populao a um pico nesse panorama, mas apenas para um dos vrios picos locais alternativos.
Nem toda a evoluo impulsionada pelas foras de seleo natural. Se a diferena seletiva entre duas variantes genticas for suficientemente pequena, menos que o recproco do tamanho ta populao, pode haver uma substituio de um alelo por outro puramente por deriva gentica. Uma grande parte da evoluo molecular parece ser a substituio de uma sequncia de protena por outra de funo equivalente. A evidncia dessa evoluo neutra que o nmero de diferenas de aminocidos entre duas espcies diferentes em alguma molcula- por exemplo, a hemoglobina diretamente proporcional ao nmero de geraes desde sua divergncia de um ancestral comum no passado evolutivo. Tal relgio molecular com uma taxa constante de mudana no seria esperado se a seleo das diferenas fosse dependente de mudanas particulares no ambiente. Alm disso, seria esperado que o relgio andasse mais rpido para as protenas tais como fibrinopeptideos, nas quais a composio de aminocidos no critica para a funo, essa diferena na velocidade do relgio de fato observada. Assim, no podemos supor sem evidncia que as mudanas evolutivas so o resultado de seleo natural adaptativa. Tanta evoluo de sequncias efetivamente neutra que no h uma relao simples entre a quantidade de mudanas na sequncia de DNA de um gene e a quantidade de mudana, se alguma, na funo da protena codificada. Algumas funes de protenas podem mudar por uma nica substituio de aminocido, enquanto outras precisam de um conjunto de substituies. Uma restrio importante na evoluo das sequncias codificantes o efeito pleiotrpicos das mutaes. Se uma protena tem mltiplas funes em tecidos diferentes, as mutaes nas sequncias codificantes podem afetar rodas as funes e ter consequncias negativas para adaptabilidade. Os potenciais efeitos pleiotrpicos das mutaes codificantes podem ser evitados por mutaes em elementos regulatrios no-codificantes que podem mudar ou parte do corpo e no em outras. A evoluo da sequncia regulatria de ao cis central para a evoluo de caractersticas morfolgica e a expresso dos genes ferramentais que controlam o desenvolvimento. Novas funes geralmente surgem pela evoluo de genes duplicados. Esse novo DNA pode surgir pela duplicao do genoma inteiro (poliploidia) seguida
de uma lenta divergncia evolutiva do conjunto cromossmico extra. O que tem sido uma frequente ocorrncia em plantas. Uma alternativa a duplicao de genes isolados seguida de seleo da diferenciao. Outra fonte de DNA, recentemente descoberta, a entrada no genoma de DNA de organismos totalmente no-relacionados por infeco seguida de interao do DNA exgeno no genoma nuclear; ou pela formao de organelas celulares extranucleares com seus prprios genomas. As mitocndrias e os cloroplastos em organismos superiores surgiram por essa via. A grande diversidade de formas de vida diferentes que existem uma consequncia das historias evolutivas independentes de populaes
separadas. Para populaes diferentes divergirem uma da outra, elas no devem trocar genes; assim, a evoluo independente de grandes nmeros de espcies diferentes requer que essas espcies sejam reprodutivamente isoladas umas das outras. De fato, definimos uma espcie como uma populao de organismos que trocam gene entre si e que so
reprodutivamente isolados de outras populaes. Os mecanismos de isolamento reprodutivo podem ser pr ou pos zigticos. Os mecanismos de isolamento pr zigticos so os que impedem a unio de gametas de duas espcies. Esses mecanismos podem ser de incompatibilidade comportamental dos machos e fmeas de espcies diferentes, diferenas na poca ou local de sua atividade sexual, diferenas anatmicas que tornam impossvel
mecanicamente a reproduo, ou incompatibilidade fisiolgica dos prprios gametas. Mecanismos de isolamento pos zigtico incluem a incapacidade de os embries hbridos se desenvolverem ate a vida adulta, a esterilidade dos adultos hbridos e a degradao das geraes posteriores de gentipos recombinantes. Na maior parte, as diferenas genticas responsveis pelo isolamento entre espcies proximamente relacionadas espalham-se por todos os cromossomos, embora em espcies com determinao cromossmica sexual possa haver uma concentrao de genes de incompatibilidade no cromossomo sexual. No geral, a evoluo gentica um processo histrico que est sujeito a contingncias histricas e ao acaso, mas restrita pela necessidade dos
organismos de sobreviver e se reproduzir em um mundo em constante mudana. Atividades
1. Uma protena um polmero composto de monmeros chamados de aminocidos. Em outras palavras: a. Uma protena um cis b. Uma protena um gene c. Uma protena um gameta d. Uma protena uma estrutura do genoma e. Uma protena uma cadeia de aminocidos e a cadeia s vezes chamada de polipeptdeo.
2. Podemos dizer que a Bioinformtica : a. O estudo do contedo de informao dos genomas. b. A sequncia genmica. c. A informao de um organismo funcional. d. A sequncia de gametas. e. Estudo das diferenas genticas.
3. Qual alternativa a correta: 1. Se o DNA de um gentipo diferente do da receptora, o gentipo da receptora pode tornar-se permanentemente alterado, um processo
adequadamente chamado de transformao. 2.A teoria darwiniana da evoluo explica as mudanas que ocorrem em populaes de organismos como resultado de mudanas nas frequncias relativas de variantes diferentes na populao. 3.A respeito da simplicidade da forma, as bactrias no tm em comum com membros maiores e complexos de outros reinos a tarefa fundamental de regular a expresso de seus genes.
a. As afirmaes 1 e 2 esto corretas. b. As afirmaes 2 e 3 esto corretas. c. As afirmaes 1, 2 e 3 esto corretas. d. A afirmao 1 esta correta. e. Todas as afirmativas esto corretas.
4. Existem quatro tipos diferentes de bases nos nucleotdeos no DNA: a. b. c. d. e. Adenina (A), timina (T), guanina (G) e citosina (C ) Adenina (A), guanina (G), citositina (C) e timinamida (T) Adenilina (A), guarnitina (G), cis (C) e trinamina (T) Adenina (A), timina (T), guarnina (G ) e citosina (C) Adenina (A), timina (T), guanina (G) e cistosina (C)
5. Vrus: essas partculas simples no-vivas no tm toda maquinaria metablica. Baseado nessa afirmao pode se dizer que: a. Um vrus infecta uma clula hospedeira e modifica seu aparelho biossinttico para a produo de mais vrus, incluindo a replicao dos genes virais. b. Um vrus infecta apenas animais, que contenham clulas hospedeiras. c. Um vrus pode produzir mais vrus se o ser vivo no tiver aparelho biossinttico. d. Um vrus pode apenas infectar humanos, levando-o a bito. e. Um vrus pode ser considerado uma clula.
Gabarito: 1 e /2 a / 3 c/ 4 a / 5b
7. HICKMAN JR., Cleveland P.; ROBERTS, L. S.; LARSON, Allan. Princpios Integrados de Zoologia. 11. ed. Rio de Janeiro: Guanabara Koogan, 2004. ORIGEM E QUMICA DA VIDA
De acordo com o big-bang, o Sol e os planetas foram formados h, aproximadamente, 4,6 bilhes de anos, a partir de uma nuvem esfrica de poeira csmica e gases, a qual colapsou formando um disco rotatrio. Oparin e Haldane propuseram que a forma de vida mais simples apareceu pela combinao de pequenas molculas em molculas orgnicas mais complexas. Molculas auto-replicveis eram produzidas e formaram o conjunto de microrganismos vivos. A evoluo qumica no ambiente pr-bitico produziu compostos orgnicos simples, os quais formaram blocos de construo das clulas vivas. Os carboidratos so compostos de carbono, hidrognio e oxignio. Participam no protoplasto com funo estrutural e como fonte de energia qumica. So sintetizados pelas plantas verdes a partir do gs carbnico e da gua, com auxlio da luz solar (fotossntese). O mais importante carboidrato a glicose. Exemplos de carboidratos: acares, amidos, celulose. So divididos em: monossacardeos (acares simples: 4, 5 ou 6 carbonos), dissacardeos (acares duplos: ligao entre dois acares simples), polissacardeos (acares complexos: muitas molculas de acares simples unidas em polmeros). Exemplos de polmeros: amido (forma pela qual a maioria das plantas armazena o acar), glicognio (forma mais importante de estocar acares nos animais). Os lipdeos so compostos de molculas de polaridade baixa, sendo insolveis na gua. Os trs principais grupos so: -gorduras neutras: principal combustvel dos animais. No tm funo estrutural nas clulas. Incluem os triglicerdeos. - fosfolipdeos: componentes importantes dos tecidos. - esteroides: so alcois complexos. Ex: colesterol, vitamina D, hormnios sexuais.
Protenas so molculas compostas de aminocidos. Os aminocidos esto unidos por ligaes peptdicas, onde o grupo carboxila de um aminocido se liga ao grupo amina de outro, por ligao covalente e eliminao de gua. As protenas podem funcionar como enzimas (catalisadores biolgicos). Os cidos nuclicos so substncias complexas e fundamentais, que armazenam ordens para sntese de enzimas e outras protenas. Dois tipos: cido desoxirribonucleico (DNA) e cido ribonucleico (RNA), ambos formados por nucleotdeos. Na dcada de 1860, experimentos de Pasteur convenceram cientistas de que os organismos no se originam da matria inorgnica. Mais de meio sculo depois, Oparin e Haldane proporcionaram uma explicao para a pergunta como um ancestral comum a todas as formas vivas poderia ter surgido de matria no viva h mais de 4 bilhes de anos?. A origem da vida aconteceu aps um longo perodo de evoluo molecular abiognica, quando molculas orgnicas acumularam-se lentamente em uma sopa primordial. A atmosfera da Terra era redutora, com pouco ou nenhum oxignio livre presente. Radiao ultravioleta, descargas eltricas de relmpagos ou energia de chamins hidrotermais poderiam ter provido energia para a formao inicial das molculas orgnicas. Miller e Urey mostraram a plausibilidade da hiptese Oparin-Haldane atravs de experimentos simples. A concentrao de reagentes necessria para a sntese inicial de molculas orgnicas possivelmente foi propiciada por superfcies midas, partculas de calcrio, pirita ou outras condies. Provavelmente, o RNA foi a primeira molcula realizadora de funes codificantes da informao. Quando sistemas autoreplicantes se estabeleceram, a evoluo provavelmente aumentou a sua diversidade e complexidade, por seleo natural. A apario da vida na Terra no ocorreria sem gua, e depende tambm da qumica do carbono. Os hetertrofos precisam obter seus suprimentos de alimento do ambiente. Supe-se que os primeiros organismos foram hetertrofos, e no teriam sido obrigados sntese do prprio alimento, pois havia estoque de nutrientes na sopa pr-bitica. A evoluo posterior produziu organismos auttrofos, que sintetizam seu alimento a partir de fontes inorgnicas usando a luz ou outra fonte de energia. A autotrofia evoluiu para a forma
fotossinttica, onde tomos de hidrognio (obtidos da gua) reagem com dixido de carbono (retirado da atmosfera) para gerar acares (fonte de alimento) e oxignio molecular (liberado para a atmosfera). O oxignio produzido pela fotossntese acumulou-se na atmosfera. Quando atingiu aproximadamente 1% de sua concentrao atual, o oznio comeou a se acumular e absorver radiao ultravioleta. Com a acumulao de oxignio atmosfrico apareceu o metabolismo (aerbico) oxidante. A maioria das formas de vida tornaram-se dependentes deste. Atualmente, quase todo o oxignio produzido por cianobactrias, algas eucariticas e plantas. As bactrias so chamadas procariotos, as quais contm uma nica e longa molcula de DNA, localizado no nucleoide. No apresentam organelas com membranas e seu DNA no est associado a histonas. Compreendem duas linhas de descendncia distinta: Eubacteria e Archae. Diferem principalmente em relao ao metabolismo e ausncia de cido murmico (nas Archae). Os eucariotos apresentam clulas com ncleo circundado por membrana, contendo cromossomos compostos de cromatina. Geralmente so maiores que os procariotos e contm muito mais DNA. Incluem animais, fungos, plantas e protozorios. Mitocndrias e plastdeos tm seu prprio DNA (alm do DNA do ncleo da clula) Todos os tecidos e rgos so compostos por clulas. Protoplasma: termo introduzido por J. Purkinje para descrever o contedo celular. O interior de uma clula est composto por vrias organelas celulares, com funes especficas. A principal diferena entre clulas procariticas e eucariticas a ausncia de ncleo circundado por membrana nos procariotos e presentes nos eucariotos. Tanto procariotos quanto eucariotos tem DNA, usam o mesmo cdigo gentico e sintetizam protenas.
Componentes das clulas eucariticas
Esto
envolvidas
numa
membrana
plasmtica,
qual
tem
permeabilidade seletiva (modelo mosaico fluido). O ncleo ovoide, envolvido
por duas membranas, e contm cromatina (carrega a informao gentica), complexo de RNA, histonas e no-histnicas. Os nuclolos levam cpias mltiplas da informao do DNA para sintetizar o RNA ribossmico. A regio fora do ncleo chama-se citoplasma, onde existem vrias organelas. Modelo mosaico fluido: uma bicamada de fosfolipdios, com suas extremidades hidrossolveis orientadas para fora, e lipossolveis para dentro da membrana. A bicamada lquida, permitindo flexibilidade membrana. Glicoprotenas (protenas ligadas a carboidratos) so essenciais. O retculo endoplasmtico (RE) um complexo de membranas que separa alguns produtos celulares da maquinaria sintetizadora que os produz. As membranas do RE podem ser cobertas por ribossomos, conhecidas como RE rugoso, e quando os ribossomos esto ausentes podem ser chamados RE liso (funo de sntese de lipdeos e fosfolipdeos). O complexo de Golgi composto por uma pilha de vesculas membranosas que armazenam e modificam produtos proteicos. Lisossomos: envolvidos na quebra de material estranho. As mitocndrias so envolvidas por membrana dupla. Nela produzido o ATP (trifosfato de adenosina), a molcula de transferncia de energia mais importante. A mitocndria autorreplica-se. O citoesqueleto fornece suporte, mantm a forma e permite locomoo e translocao de organelas na clula. Microfilamentos so responsveis pela habilidade da contrao celular. So feitos de actina. Microtbulos so compostos de tubulina, tendo papel no movimento dos cromossomos para as clulas filhas na diviso celular, sendo importantes tambm na organizao e transporte celular. So gerados pelo centrossomo, os quais no so envolvidos por membranas, e no seu interior encontram-se os centrolos, tambm compostos de microtbulos. Clulas de plantas no tm centrolos.
Superfcies celulares
Clios ou flagelos varrem os materiais para longe das clulas. Os flagelos so o meio de locomoo para clulas reprodutivas masculinas na maioria dos animais e muitas plantas.
Muitas clulas se movem por movimento ameboide, atravs de pseudpodes. Juno oclusiva: membranas de duas clulas podem fundir-se. Esse tipo de juno previne a passagem de molculas entre as clulas. Junes comunicantes: funcionam como estrutura de comunicao intercelular, formando minsculos canais entre as clulas. Microvilos: pequenas projees tubulares, que aumentam a superfcie de absoro. A membrana plasmtica muito importante para manter a integridade celular. Age como um porteiro, selecionando a entrada e sada de substncias envolvidas no metabolismo celular. Modos pelos quais uma substncia pode atravessar a membrana celular: difuso, sistema de transporte mediado e endocitose. Difuso: movimento de partculas de uma rea de concentrao alta para uma rea de concentrao baixa. Se uma clula cercada por membrana imersa numa soluo com concentrao mais alta de molculas do que o fluido interno da clulas, formase um gradiente de concentrao. A gua flui pela membrana da soluo mais diluda para a mais concentrada. As molculas movem-se no gradiente de concentrao pela membrana onde as molculas de gua esto mais concentradas para a rea onde esto menos concentradas, conhecida como osmose.
Transporte por mediao As permeases formam uma passagem pequena pela membrana, permitindo que a molcula do soluto cruze a bicamada fosfolipdica. Difuso facilitada: uma permease auxilia a difuso da molcula atravs da membrana. O transporte ativo prov energia ao sistema transportador para mover molculas em direo oposta a um gradiente de concentrao. Envolve consumo de energia, pois os materiais so bombeados contra um gradiente de concentrao. Endocitose a ingesto de material pelas clulas.
Exocitose: a membrana de uma vescula funde-se com a membrana plasmtica para expulsar seus contedos. Ocorre para remover resduos no digeridos, secretar substncias.
MITOSE E DIVISO CELULAR
Mitose o processo de diviso nuclear para formar clulas somticas. Distribui os cromossomos e seu contedo de DNA para a continuidade das geraes celulares. o nico mecanismo para transferncia de informao gentica do progenitor prognie em animais com reproduo assexuada. Estgios da mitose: prfase: os cromossomos replicados condensam-se em corpos reconhecveis. Forma-se um fuso entre os centrossomos enquanto estes se separam em plos opostos. Metfase: as cromtides irms movem-se para o centro da clula. Anfase: centrmeros dividem-se e cada uma das metades puxada para os centrossomos pelas fibras cinetocricas. Telfase: cromossomos posicionam-se no local do ncleo de cada nova clula. A membrana nuclear reaparece e ocorre a citocinese (diviso do citoplasma). A replicao do DNA ocorre durante a interfase, na fase S. O perodo S precedido pelo perodo G1 (fase preparatria) e sucedido pelo G2. Na G1 so sintetizados RNA de transferncia, ribossomos, RNA mensageiro e enzimas. Existe ainda a fase G0, onde h inatividade (ex: neurnios, pois no se dividem).
METABOLISMO CELULAR
Leis da termodinmica
A primeira lei afirma que a energia no pode ser destruda, embora possa mudar de forma. A segunda lei estabelece que a estrutura dos sistemas tende desorganizao total, ou aumento de entropia, conforma energia dissipada. A energia solar capturada pela fotossntese como energia qumica
passada atravs das cadeias alimentares, onde usada para biossntese transporte ativo e movimento, antes de ser dissipada como calor. Enzimas so protenas que aceleram muito as taxas de reaes qumicas. Uma enzima liga seu reagente (substrato) a um centro ativo. As clulas usam energia estocada nas ligaes qumicas dos combustveis orgnicos degradando-os atravs de passos controlados por enzimas. Esta energia transferida para o ATP. A glicose uma importante fonte de energia para as clulas. Na respirao, a glicose, com seus carbonos, quebrada em duas molculas de cido pirvico com trs carbonos. O cido pirvico descarboxilado para formar acetil-coenzima A com dois carbonos, um intermedirio estratgico conduzido ao ciclo de Krebs. A acetil-coenzima A pode ser derivada tambm da quebra de gordura. No ciclo de Krebs, a acetil-coenzima A oxidada a dixido de carbono em vrias reaes, fornecendo eltrons energizados, passados a molculas receptoras de eltrons (NAD e FAD). No estgio final, os eltrons energizados so passados ao longo de uma cadeia transportadora de eltrons localizados nas membranas internas da mitocndria. Um total lquido de 36 molculas de ATP pode ser gerado de uma molcula de glicose. Na ausncia de oxignio (gliclise anaerbica), a glicose degradada a duas molculas com trs carbonos de cido lctico, fornecendo duas molculas de ATP. Embora seja muito menos eficiente do que a respirao, a gliclise anaerbica supre a energia essncia para a contrao muscular quando um gasto excessivo de energia ultrapassa a capacidade do sistema de fornecimento de oxignio de um animal, alm de ser a nica fonte de gerao de energia para microrganismos que vivem em ambientes onde no h oxignio livre.
PRINCPIOS DE GENTICA
Cada gerao passa para a seguinte instrues necessrias para dar continuidade vida, sendo o gene a unidade da herana. Gregor Johann Mendel foi o primeiro a formular os princpios bsicos da hereditariedade. As observaes de Mendel baseavam-se em ervilhas de
jardins, pois eram linhagens puras, selecionadas cuidadosamente. Outro motivo para Mendel trabalhar com ervilhas, ocorria por apresentarem autofertilizao, mas tambm capazes de fertilizao cruzada. Ele trabalhava com caracteres nicos: plantas altas e plantas ans, e sementes lisas e rugosas. Mendel fez cruzamentos entre plantas com essas caractersticas, removendo o estame de uma flor, para impedir a autofertilizao, e coloca o plen da flor de uma planta com uma caracterstica contrastante em seu estigma. A polinizao das flores experimentais por insetos e vento era impedida. Quando a flor experimental produzia sementes, ele anotava o tipo de plantas produzidas atravs da semente plantada. Em seguida, cruzava esses hbridos entre si. Cada clula do corpo possui dois cromossomos, onde os membros de cada um destes pares so chamados homlogos. A meiose consiste em duas divises do ncleo, onde os cromossomos dividem-se apenas uma vez. Assim, os gametas maduros tm nmero haploide de cromossomos. Quando os gametas se unem na fertilizao, forma-se o zigoto (em humanos: diploide). Formas alternativas de genes para um mesmo carter so chamados alelos. A localizao de um gene em um cromossomo o loco. Para determinao do sexo, foi descrito o tipo XX-XO, onde as fmeas apresentam dois cromossomos X, e os machos apenas um (O ausncia de cromossomo). Em aves, mariposas, borboletas e alguns peixes, o macho tem 2 cromossomos X (chamados ZZ) e a fmea, um X e um Y (chamados ZW). Nem todos os animais dioicos tm o sexo determinado por cromossomos. Em alguns, o sexo determinado por fatores como a temperatura (ex: em tartarugas e alguns lagartos) e comportamento (alguns peixes). Os genes determinam as caractersticas de um organismo, e so herdados. Os variantes allicos podem ser dominantes, recessivos ou intermedirios. O alelo recessivo no gentipo heterozigoto no se expressa no fentipo e requer a condio homozigtica para se manifestar. Primeira Lei de Mendel (lei da segregao): em um cruzamento monobrido envolvendo um alelo dominante e um recessivo (onde os pais so
homozigotos), a gerao F1 vai ser toda heterozigota. Na gerao F2 os gentipos seguiro a proporo 1:2: 1 e os fentipos 3:1. Segunda Lei de Mendel (lei da segregao independente): os heterozigotos homozigotos. apresentam Os fentipos intermedirios (onde os entre genes os fentipos duas
cruzamentos
dibridos
para
caractersticas situam-se em dois pares separados de cromossomos homlogos) mostram propores fenotpicas de 9:3: 3:1, com caracteres dominantes e recessivos. Pleiotropia: muitos genes tm mais de um efeito nos fentipos. Epistasia: um alelo em um loco pode mascarar ou impedir a expresso de um alelo de outro loco que atua sob o mesmo carter. Existem genes polignicos, onde ao invs de originarem fentipos alternativos discretos, os caracteres mostram uma variao contnua entre dois extremos, conhecida como herana quantitativa. Ex: grau de pigmentao em acasalamento entre raas humanas negra e branca. A herana de alguns caracteres depende do sexo do progenitor e do descendente (herana ligada ao sexo). Ex: hemofilia e daltonismo, ambos determinados atravs de sua presena no cromossomo X. Expressam-se fenotipicamente quando ambos os genes para o defeito esto presentes na me, ou quando apenas um gene para o efeito est presente no pai. O crossing-over uma maneira de trocar genes entre cromossomos homlogos, aumentando a quantidade de recombinao gentica. Desvios estruturais e numricos que afetam muitos genes de uma s vez so chamados aberraes cromossmicas. So chamadas de euploidias mudanas no nmero de cromossomos, quando h adio ou deleo de conjuntos de cromossomos e aneuploidias quando um nico cromossomo adicionado ou subtrado. O tipo mais comum de euploidia a poliploidia. A aneuploidia normalmente causada pela no-separao dos cromossomos. Isso resulta em um gameta possuindo n 1 nmero de cromossomos, e outro com n + 1. Se o gameta com n 1 fertilizado por um gameta normal o resultado um animal monossmico. Se o gameta com n + 1 fertilizado com um gameta normal, ocorre uma trissomia.
O conceito de gene foi cunhado por W. Johannsen em 1909, referindo aos fatores hereditrios de Mendel. Eles tm o cdigo para produtos essenciais. So unidades de informao molecular que podem manter suas identidades por vrias geraes e permitem que suas particularidades sejam copiadas. De forma mais abrangente um gene pode ser definido como uma sequncia de cidos nucleicos, que codifica um polipeptdeo funcional ou uma sequncia de RNA. As clulas contm dois tipos de cidos nuclicos: DNA (material gentico) e RNA (atua na sntese protica). Ambos so polmeros, constitudos por nucleotdeos. Cada nucleotdeo contm um acar, uma base nitrogenada e um grupamento fosfato. O acar uma pentose, que no DNA uma desoxirribose e no RNA uma ribose. As bases nitrogenadas dos nucleotdeos so: pirimidinas (nico anel com cinco membros) ou purinas (dois anis ligados). As purinas no DNA e no RNA so adenina e guanina. As pirimidinas no DNA so a timina e a citosina, e no RNA a uracila e citosina. O eixo da molcula de DNA construdo por cido fosfrico e desoxirribose e as bases nitrogenadas ligam-se a este eixo. O DNA composto de duas cadeias, ligadas transversalmente por pontes de hidrognio especficas, entre as bases purnicas e pirimidnicas. O pareamento das bases : adenina com timina (AT) e guanina com citosina (GC). O resultado uma estrutura em forma de escada, torcida em dupla hlice. As fitas so antiparalelas. Em relao estrutura, o RNA semelhante ao DNA, exceto por ter apenas uma cadeia de polinucleotdeos, ter ribose no lugar da desoxirribose e uracila no lugar da timina. Existem trs tipos de RNA (mensageiro transportador e ribossmico). Replicao: quando uma clula se divide, a estrutura do DNA deve ser copiada para as clulas filhas. As duas fitas da dupla hlice se separam, servindo como molde para sintetizar uma fita complementar. A DNA polimerase monta uma nova fita. Transcrio: as trincas codificadas pelo DNA so transcritas na forma de RNA mensageiro (RNAm), onde a uracila substitui a timina. O RNA de transferncia, ribossmico e mensageiro so transcritos diretamente do DNA.
necessria a enzima RNA-polimerase. Apenas uma das fitas do DNA utilizada como molde para a sntese de RNA, pois apenas uma contm o cdon AUG, o qual inicia a mensagem. Muitos genes so descontnuos, interrompidos por sequncias de bases que no codificam o produto final. O RNAm transcrito a partir deles precisa ser editado, antes da traduo. Os segmentos intermedirios so conhecidos como ntrons e os que codificam a parte do RNA que madura, chama-se xons. As sequncias de bases em alguns ntrons so complementares a outras sequncias de bases do mesmo ntron. Sendo assim, o ntron pode dobrar-se de maneira que as sequncias podem se parear. Traduo: ocorre nos ribossomos. O RNA ribossmico composto por uma subunidade pequena e uma grande. As molculas de RNAm se ligam aos ribossomos, formando um complexo RNA mensageiro-ribossomo. O RNAm se liga a diversos ribossomos de uma vez, formando o polirribossomo, o que permite que diversas molculas do mesmo tipo de protena sejam sintetizadas de uma nica vez. Para a montagem das protenas necessrio o RNA de transferncia (RNAt). As molculas de RNAt capturam aminocidos livres do citoplasma e os carregam at o polissomo, onde so montados, formando uma protena. Cada RNAt acompanhado por uma RNAt-sintetase especfica, enzima essa necessria para escolher e ligar o aminocido correto no final de cada RNAt, processo chamado de carregamento. Os cdons so lidos e as protenas so montadas ao longo do RNAm na direo da extremidade 5 para a extremidade 3. DNA recombinante As endonucleases de restrio cortam a dupla hlice do DNA em locais especficos. Muitas cortam os filamentos de DNA e deixam extremidades adesivas. Quando esses fragmentos de DNA so misturados com outros, que foram clivados pela mesma endonuclease, eles tendem a se ligar, sendo fixados em sua nova posio pela DNA-ligase. Se os filamentos de DNA ligados forem de duas fontes diferentes, o produto o DNA recombinante. Por exemplo: um gene de um mamfero pode ser introduzido em clulas
bacterianas, atravs de plasmdeo, que ento se multiplicam e expressam o gene. Reao em cadeia da polimerase (PCR) uma tcnica utilizada para obter grandes quantidades de DNA a partir de amostras muito pequenas. No PCR, dois primers so sintetizados. Uma quantidade de cada primer adicionado na amostra de DNA do organismo, e a mistura aquecida para separar a dupla hlice. A mistura resfriada, tendo uma maior probabilidade de cada filamento do gene de interesse se ligar a um primer. A DNA polimerase adicionada, com os quatro desoxirribonucleotdeos trifosfatos. A mistura reaquecida e esfriada novamente, permitindo que mais primers se liguem a filamentos originais e copiados. A cada ciclo da sntese do DNA, o nmero de cpias do gene duplicado, e como cada ciclo pode durar menos que 5 minutos, o nmero de cpias de um gene pode aumentar de um milho em menos de duas horas.
Mutaes: so mudanas fisicoqumicas nos genes, que resulta na alterao da sequncia de bases do DNA. Uma mutao pode resultar da substituio de um cdon (ex: anemia falciforme), envolver deleo ou insero de bases, onde a traduo do RNAm vai ser deslocada, gerando aminocidos incorretos.
PROCESSO REPRODUTIVO
Reproduo assexuada: produo de indivduos sem envolver gametas, onde todos os descendentes tm o mesmo gentipo e so chamados de clones. Ocorre em bactrias, eucariotos unicelulares e em muitos filos de invertebrados. Tipos: fisso binria: o corpo do progenitor divide-se por mitose em duas partes, as quais crescem e se tornam indivduos similares ao progenitor. Fisso mltipla: ocorre a diviso do ncleo, repetidamente, antes da diviso do citoplasma. Brotamento: o novo indivduo surge como um broto, crescendo a partir do progenitor e posteriormente se separa. Ex: em cnidrios. Gemulao: formao de um novo indivduo a partir de um agregado de clulas
envolvidos por uma cpsula. Ocorre em muitas esponjas. Fragmentao: um animal multicelular fragmenta-se em duas ou mais partes e cada fragmento capaz de tornar-se um indivduo completo.
Reproduo sexuada: produo de indivduos atravs de gametas. Tipos: 1. Bissexuada: envolve dois indivduos separados. Indivduos dioicos: tm sexos separados. Indivduos monoicos: tm rgos reprodutivos tanto de macho quanto de fmea. vulos so produzidos pelas fmeas, sendo grandes, sem mobilidade e produzidos em nmeros pequenos.
Espermatozoides so produzidos pelos machos, sendo pequenos, mveis e produzidos em grandes quantidades. 2. Hermafroditismo e 3.partenognese: envolvem apenas um indivduo. No hermafroditismo, animais possuem tanto rgos reprodutores femininos quanto masculinos. Exemplo de hermafroditas: muitos invertebrados ssseis, subterrneos ou endoparasitas e alguns peixes. J a partenognese o desenvolvimento de um embrio a partir de um vulo no fertilizado, ou num vulo onde os ncleos feminino e masculino deixam de se unir aps a fertilizao. Partenognese ameitica: no ocorre meiose, o ovo formado por diviso celular mittica. Partenognese meitica: ovo haploide formado por meiose. Nos vertebrados, as clulas germinativas primordiais surgem no endodema do saco vitelino, migrando para as gnodas. Nos mamferos uma gnoda vai se tornar um testculo devido aos sinais masculinizantes do cromossomo Y. As estruturas reprodutoras da fmea desenvolvem-se na ausncia de sinais do cromossomo Y. Chama-se gametognese as transformaes que resultam na formao de gametas maduros. Nos testculos, denomina-se espermatognese e nos ovrios oognese. Espermatognese: as paredes dos tubos seminferos contm clulas germinativas em diferenciao, as quais se desenvolvem em contato com clulas de sustentao, as quais fornecem nutrio. A camada mais externa apresenta espermatognias. Cada espermatognia torna-se um espermatcito primrio, o qual sofre uma diviso meitica, originando os espermatcitos secundrios. Cada espermatcito secundrio sofre a segunda diviso meitica,
e aps as duas etapas da meiose, cada um origina quatro espermtides (n). As espermtides transformam-se em espermatozoides. Oognese: as oognias (clulas germinativas iniciais) multiplicam-se por mitose e formam ocitos primrios. Os cromossomos de cada ocito primrio pareiam-se. O citoplasma divide-se. O ocito secundrio, uma das clulas filhas, tem tamanho maior, a outra muito pequena e chama-se primeiro corpsculo polar. Na segunda diviso meitica, o ocito secundrio divide-se em uma otide e um pequeno corpsculo polar. A otide desenvolvese em vulo. Os corpsculos no so funcionais, e desintegram-se. Animais ovparos: depositam seus ovos no meio externo. A fertilizao pode ser interna ou externa (fertilizado pelo macho). Ocorre na grande maioria dos invertebrados e muitos vertebrados. Os animais ovovivparos retm os vulos em seu corpo. Ocorre em muitos invertebrados, peixes, rpteis. Nos
animais vivparos os ovos desenvolvem-se no oviduto ou no tero. Ocorre principalmente em mamferos e peixes elasmobrnquios. Os invertebrados que transferem os espermatozoides dos machos para as fmeas, com fertilizao interna, necessitam de rgos e canais para facilitar essa funo. J os sistemas reprodutores de invertebrados que liberam os gametas na gua, so um pouco mais do que centros produtores de gametas. Reproduo nos vertebrados Em machos de peixes e anfbios o canal que sai dos rins tambm serve como duto espermtico. Os canais abrem-se em cloaca, uma cmara nica onde os canais intestinal, reprodutor e excretor convergem. A maioria dos mamferos placentrios no possui cloaca. Sistema reprodutor dutos masculino: eferentes, inclui testculos (produo de
espermatozoides),
duto
deferente
(duto
ejaculatrio),
glndulas acessrias e pnis (algumas aves e rpteis, e em todos os mamferos). Sistema reprodutor feminino: inclui os ovrios (contendo milhares de vulos dentro de folculos), tubos uterinos (transporte dos vulos), tero e vagina.
A natureza cclica da reproduo tornou necessria a evoluo de mecanismos hormonais que controlam a produo das clulas germinativas, sinalizam o momento para acasalar e preparam canais e glndulas para que os vulos sejam fecundados. Os centros neurossecretores do encfalo secretam o hormnio liberador da gonadotrofina (GnRH), que estimula as clulas endcrinas da hipfise anterior a liberar o hormnio folculo-estimulante (FSH) e o hormnio luteinizante (LH), os quais estimulam as gnodas. Os estrgenos e a progesterona nas fmeas, e a testosterona e a diidrotestosterona (DHT) nos machos, controlam o crescimento das estruturas sexuais acessrias e das caractersticas sexuais secundrias. O estrognio induz a proliferao inicial do endomtrio uterino. A liberao de GnRH e LH na metade do ciclo induz a ovulao e faz com que o corpo lteo secrete a progesterona, que completa a preparao do tero para a implantao. Se um vulo fertilizado, a gravidez mantida por hormnios produzidos pela placenta e pela me. A gonadotrofina corinica humana (hCG) mantm a secreo de progesterona e estrognio do corpo lteo, enquanto a placenta cresce e passa a secretar estrognio, progesterona, hCG e lactognio placentrio humano (hPL). As glndulas mamrias sero estimuladas em preparao para a lactao pelo estrognio, progesterona e hPL. O nascimento ocorre devido diminuio nos nveis de progesterona e aumento dos nveis de estrognio. Nascimentos mltiplos em mamferos podem ser resultados da diviso de um zigoto (produzindo gmeos idnticos, monozigticos), ou de zigotos separados (gmeos no-identicos ou fraternos).
PRINCPIOS DO DESENVOLVIMENTO
Fecundao a unio dos gametas feminino e masculino, para formar o zigoto. Funes: recombina genes maternos e paternos, ativa o vulo para iniciar o desenvolvimento. Porm, nem sempre o contato do espermatozoide ou genoma paterno so fatores essenciais para a ativao do vulo. Na oognese, o vulo prepara-se para a fecundao e incio do desenvolvimento. O espermatozoide elimina seu citoplasma e condensa seu
ncleo no menor tamanho possvel, e o vulo aumenta de tamanho, acumulando reservas de vitelo. Aps o reinicio da meiose, o vulo elimina o excesso de material cromossmico sob forma de corpsculos polares. Aps atividade de sntese, o ocito est altamente estruturado, possuindo reservar importantes para manter as necessidades nutricionais do embrio e direcionar seu desenvolvimento pela clivagem. A maioria dos invertebrados e vrios peixes marinhos liberam seus gametas no oceano. Para aumentar a probabilidade de contato com um espermatozoide, os vulos de vrias espcies marinhas liberam um fator quimiotxico, que atrai o espermatozoide. No local de contato entre o espermatozoide e o envoltrio vitelino do vulo surge um cone de fecundao, onde penetrar a cabea do espermatozoide. Deve ser evitada a polispermia (penetrao de mais de um espermatozoide), pois prejudica o desenvolvimento. O espermatozoide perde seu flagelo aps a fuso das membranas do espermatozoide e do vulo, e ocorre o rompimento de seu envoltrio nuclear, permitindo a expanso da cromatina. O ncleo expandido do espermatozoide (proncleo) dirige-se para dentro do vulo para unir-se ao proncleo feminino. A fecundao origina vrias modificaes importantes no citoplasma do zigoto. Aps, o zigoto inicia a clivagem. O embrio divide-se vrias vezes, transformando a massa citoplasmtica em blastmeros. H apenas subdiviso da massa, no h crescimento nessa fase. A clivagem afetada pela quantidade e distribuio de vitelo e por genes. Em relao distribuio de vitelo: isolcitos: ovos com pouco vitelo, distribudos uniformemente. A clivagem ento holoblstica, com a linha de clivagem estendendo-se por todo o ovo. mesolcitos: ovos com quantidade moderada de vitelo. Ex: ovos de anfbios. A clivagem tambm holoblstica. telolcitos: ovos com grande quantidade de vitelo. A clivagem meroblstica (clivagem parcial). Ex: ovos de aves, rpteis, muitos
peixes,
alguns
anfbios,
moluscos
cefalpodes
mamferos
monotremados. centrolcitos: ovos com grande massa de vitelo localizada na regio central. A clivagem do citoplasma limita-se a uma camada superficial do citoplasma livre de vitelo, o interior permanece no-clivado. Ex: insetos e artrpodes. A quantidade de vitelo tambm afeta o modo de desenvolvimento dos embries. Desenvolvimento indireto: quando o estgio de larva se interpe entre o embrio e o adulto. Ex: invertebrados aquticos, diversos anfbios. Desenvolvimento direto: existe uma ligao placentria com a me, onde os embries so nutridos durante a gestao. Ex: mamferos, aves e rpteis. A clivagem resulta em um aglomerado de clulas, chamado de blstula. Na gastrulao, as clulas da superfcie do embrio movem-se para dentro formando folhetos germinativos (endoderma, ectoderma, mesoderma) e o plano corporal do embrio. O incio do desenvolvimento governado pelos produtos do genoma materno, pois o crtex do ovo contm determinantes citoplasmticos, depositados na oognese, que guiam o desenvolvimento atravs da clivagem. Com a aproximao da gastrulaao, o controle vai passando da me para o embrio. Durante o desenvolvimento certas pginas do genoma de cada clula expressam-se, e o restante encontra-se desligado. Genes homeobox codificam protenas reguladoras, que contm regies conservadas que se ligam ao DNA, denominadas homeodomnios. Os amniotas desenvolveram membranas extra-embrionrias durante sua vida embrionria. So elas: mnion (protege o embrio de choques mecnicos e adeses), alantoide (serve como depsito de rejeitos
metablicos), crion (envolve o resto do sistema embrionrio) e saco vitelino. Nos mamferos, os embries so nutridos pela placenta. Na gravidez ela torna-se um rgo nutritivo, endcrino e regulatrio para o embrio. Os folhetos germinativos formados na gastrulao diferenciam-se em tecidos e rgos. O ectoderma origina a pele e o sistema nervoso. O endoderma origina o canal alimentar, faringe, pulmes e algumas glndulas. O mesoderma forma o sistema muscular, esqueltico, circulatrio e excretor.
PLANOS CORPREOS ANIMAIS
A simetria animal refere-se correspondncia em tamanho e forma das partes de lados opostos de um plano mediano. A simetria esfrica significa que qualquer plano que passa atravs do centro do organismo divide seu corpo em metades equivalentes. Esse tipo de simetria encontrado em algumas formar unicelulares, sendo rara entre os animais. A simetria radial aplicada s formas que podem ser divididas em metades semelhantes por mais de dois planos que passam atravs do eixo longitudinal do organismo. A simetria radial ocorre, por exemplo, em esponjas, guas-vivas, ourios-do-mar. Uma forma variante da simetria radial a simetria birradial, na qual s dois planos que passam atravs do eixo longitudinal produzem metades equivalentes. A simetria bilateral se aplica aos animais que podem ser divididos em duas pores equivalentes ao longo de um plano sagital (as metades direita e esquerda).
CLASSIFICAO E FILOGENIA DOS ANIMAIS
A sistemtica animal apresentam trs principais objetivos: identificar todas as espcies de animais, avaliar os relacionamentos evolutivos entre as espcies animais e agrupar as espcies em uma hierarquia de grupos taxonmicos, que evidenciam relacionamentos evolutivos. Txons so organizados visando aumentar a inclusividade como: espcie, gnero, famlia, ordem, classe, filo e reino. Os nomes das espcies so bonimiais, onde o primeiro nome designa o gnero ao qual a espcie pertence (deve ser escrito com a letra inicial maiscula), seguida pelo epteto de espcie (letra minscula), ambos escritos em itlicos.
PROTOZORIOS
Apresentam apenas uma clula eucaritica, mas so organismos completos funcionalmente. Precisam de umidade e vivem em habitat marinhos ou de gua doce, solo, ou lugares com matria orgnica em decomposio, plantas ou animais. Na maioria das vezes so microscpicos. Muitos protozorios apresentam organelas (mitocndria, retculo
endoplasmtico, corpo de Golgi, vesculas). Movimentam-se por clios e flagelos e pseudpodes. Os clios, em muitas espcies, auxiliam na alimentao, reproduo, excreo e osmorregulao. Existem vrias formas de pesudpodes: lobpodes, filpodes, reticulpodes, axpodes. Apresentam osmorregulao. feita por difuso. Apresentam todos os tipos de nutrio: autotrfica (produo do prprio alimento atravs da fotossntese), heterotrfica (dependem de outros animais ou plantas), saprozoica (uso dos nutrientes dissolvidos no ambiente ao redor), holozoica (por fagocitose) Reproduzem-se assexuadamente por fisso, brotamento e cisto. E sexuadamente por conjugao (h troca de ncleos gamticos entre os organismos) ou singamia (fecundao de um gameta por outro). Filo Sarcomastigophora: inclui protozorios que se movem por flagelos (Mastigophora) e que se movem por pesudpodes (Sarcodina), mas essas caractersticas no so mutuamente exclusivas. Filo Apicomplexa: todos os apicomplexa so endoparasitas. vacolos, que provavelmente participam na
A excreo de resduos metablicos quase totalmente
Apresentam complexo apical. Organelas locomotoras so menos evidentes. Pseudpodes ocorrem em alguns estgios intracelulares, e os gametas de algumas espcies so flagelados. O ciclo de vida geralmente inclui tanto reproduo assexuada quanto sexuada. Filo Ciliophora: so estruturalmente complexos e diversamente especializados. A maioria de vida livre, mas alguns so comensais ou parasitas. Todos possuem clios, que batem de modo coordenado e rtmico. Os ciliados so sempre multinucleares, possuindo no mnimo um macroncleo (responsvel pelas funes de desenvolvimento e metablicas, manuteno
das caractersticas visveis) e um microncleo (participa na reproduo sexual e origina os macroncleos). A maioria dos ciliados holozoica e possui um citstoma (boca). Um exemplo conhecido de ciliado o paramcio, o qual
apresenta forma de chinelo, sendo geralmente abundante em lagos e riachos com pouca correnteza e que contenham plantas aquticas e matria orgnica em decomposio.
MESOZOA E PARAZOA
FILO MESOZOA Representam um nvel extremamente simples de organizao. Todos os mesozorios vivem como parasitas em invertebrados marinhos. Possui duas classes: Os Rhombozoa vivem em rins de cefalpodes bentnicos. So alongados. Os Orthonectida parasitam uma grande variedade de invertebrados.
FILO PLACOZOA Contm uma nica espcie: Trichoplax adhaerens. Apresenta corpo achatado, sem simetria, rgos, sistemas muscular ou nervoso. Deslizam sobre o alimento, liberando enzimas digestivas e ento, absorvem o produto.
FILO PORFERA So as esponjas. Elas possuem um grande nmero de poros minsculos e canais. So ssseis e dependem de correntes de gua passando pelo seu sistema de canal para trazer comida e oxignio, e para levar para o exterior os dejetos. No possuem nenhum rgo ou tecidos verdadeiros. No apresentam sistema nervoso ou rgos dos sentidos desenvolvidos. Variam muito de tamanho, ocorrendo desde alguns milmetros e outras podendo chegar a mais de 2 m de dimetro. Muitas espcies so coloridas. A maioria das esponjas so marinhas, mas existem tambm espcies de gua doce. Seus embries so livre-natantes, porm os adultos so sempre fixos. O padro de crescimento depende da forma do substrato, direo e velocidade das correntes de gua, disponibilidade de espao e condies ambientais.
Muitos animais vivem como comensais ou parasitas dentro ou sobre as esponjas. Porm, muitas esponjas tambm crescem sobre animais viventes As aberturas corpreas chamam-se stios, e os maiores de sculos. Essas aberturas so conectadas por sistema de canais, alguns formados por clulas flageladas (coancitos). Os coancitos mantm a gua em movimento, capturam e fagocitam as partculas alimentares trazidas pela gua. Tipos de sistemas de canais nas esponjas: asconoides (espongioceles flageladas): organizao mais simples. So pequenas e tubulares. A gua entra por poros drmicos microscpicos em uma grande cavidade, chamada espongiocele, a qual forrada por coancitos. siconoides (canais flagelados): possuem corpo tubular e sculo nico. A parede contm canais radiais, forrados por coancitos que
desembocam na espongiocele. Esta formada por clulas epiteliais. A gua entra atravs de stios drmicos nos canais inalantes, indo para os canais radiais pelas prospilas. Nos canais radiais a comida ingerida pelos coancitos. leuconoides (cmaras flageladas): a forma mais complexa.
Associaes de cmaras flageladas recebem gua dos canais inalantes e descartam nos canais exalantes, que conduzem ao sculo. As clulas das esponjas esto organizadas em meso-hilo (matriz gelatinosa). No meso-hilo so encontradas vrias clulas ameboides, fibrilas e elementos esquelticos. Clulas do tipo pinaccitos so do tipo epitelial. So um pouco contrteis. Clulas do tipo coancitos forram os canais e as cmaras flageladas. So ovoides, com uma extremidade no meso-hilo e outra exposta. Os arquecitos so clulas ameboides que se deslocam no meso-hilo. Podem fagocitar partculas e receber partculas para digesto. Todas as atividades das esponjas dependem da corrente de gua fluindo pelo seu corpo. A digesto inteiramente intracelular (ocorre dentro das clulas), onde os coancitos transmitem as partculas de alimento para os arquecitos para a digesto. No existem rgos respiratrios. Apresentam reproduo assexuada e sexuada. A reproduo assexuada ocorre por formao de brotos e pela regenerao. Brotos externos podem se
destacar da esponja parental e flutuar para formar novas esponjas, ou podem permanecer junto da esponja parental e formarem colnias. No caso dos brotos internos (ou gmulas), os arquecitos se juntam no meso-hilo e so envoltos por uma camada de espongina. Quando a esponja parental morre, as gmulas sobrevivem e ficam dormentes. Aps, as clulas das gmulas saem pela micrpila e se desenvolvem em novas esponjas. Em relao reproduo sexuada, a maioria das esponjas monoica (clulas sexuais masculinas e femininas num nico indivduo). Os
espermatozoides surgem da transformao dos coancitos. A maioria das esponjas vivpara, onde aps a fertilizao o zigoto retido e recebe nutrientes da esponja parental, at que liberada uma larva ciliada. Os espermatozoides so liberados na gua por um indivduo e so capturados pelo sistema de canal de outro indivduo. Os coancitos desse ltimo indivduo fagocitam o espermatozoide e o levam at os ocitos. Outras esponjas so ovparas, onde os ocitos e os espermatozoides so expelidos na gua. As esponjas tm uma tremenda habilidade para reparar suas injrias e restaurar as partes perdidas, conhecido como regenerao.
ANIMAIS RADIAIS (Filo Cnidaria; Filo Ctenophora)
FILO CNIDARIA Assim chamado devido presena de cnidcitos, clulas que podem conter organelas urticantes (nematocistos). Ocorrem em todos os habitats marinhos e alguns em gua doce. No h nenhuma espcie terrestre. So principalmente de locomoo lenta, porm so predadores bastante eficientes. Os cindrios, s vezes, vivem em simbiose com outros animais. Apresentam dimorfismo: todas as formas de cnidrios esto ajustadas a um entre dois tipos morfolgicos: um plipo (sendentrio, sssil) e uma medusa (forma de gua-viva, livre-natante). Os plipos geralmente
apresentem corpo tubular com uma boca cercada por tentculos em uma extremidade. Podem viver isolados, ou em colnias. As medusas tm seus
corpos na forma de guarda-chuva ou sino, com simetria tetrmera (partes do corpo organizadas em quartetos). Os nematocistos so produzidos por um cnidoblasto e so enovelados dentro de uma cpsula. Quando disparados, alguns tipos de nematocistos penetram na presa e injetam veneno. Apresentam rede nervosa com sinapses simtricas e assimtricas, alguns com rgos sensoriais. Classe Hydrozoa: a maioria dos hidrozorios colonial e marinha. O ciclo de vida tpico inclui um plipo assexuado e uma fase de medusa sexuada. A maioria dos hidrozorios marinhos est na forma de uma colnia ramificada com muitos plipos. As medusas de hidrozorios podem ser livre-natantes ou podem permanecer fixas s suas colnias. Classe Scyphozoa: os cifozorios so guas-vivas tpicas nas quais o medusoide a forma corprea dominante. Classe Cubozoa: os cubozorios so predominantemente medusoides e incluem as perigosas vespas-do-mar. Classe Anthozoa: os antozorios, ou animais-flor, so todos marinhos e polipoides, no h nenhuma fase medusoide. As ordens maiores possuem as anmonas-do-mar (solitrias, sem esqueleto) e os corais ptreos (coloniais, exoesqueleto calcreo).
FILO CTENOPHORA Os Ctenophora so birradiais e nadam atravs de oito fileiras de pentes. Apresentam coloblastos, com os quais capturam pequenas presas. Apenas uma espcie apresenta nematocistos. Os corpos so frgeis e transparentes, mas so facilmente vistos noite, quando emitem luz.
ANIMAIS BILATERAIS ACELOMADOS (Filo Platyhelminthes; Filo Nemertea; Filo Gnathostomulida)
So
considerados
trs
grupos:
Platyhelminthes,
Nemertea
Gnathostomulida. Apresentam apenas um espao interno, a cavidade digestiva. So considerados animais bilaterais acelomados, pois no possuem celoma ou pseudocele. Apresentam sistema excretor.
FILO PLATYHELMINTHES Variam em tamanho, existindo animais desde um milmetro, at muitos metros de comprimento (ex: tnias). Apresentam corpo achatado, na forma de folha, ou longo, em forma de fita. Incluem formar livres (classe Tuberlaria somente) e parasitrias. A superfcie do corpo dos platelmintos turbelrios normalmente um epitlio celular, contendo clulas mucosas e rabditos que funcionam juntos na locomoo. Os membros de todas as outras classes de platelmintos esto cobertos por um tegumento sincicial (ncleos no so separados pela interveno de membranas celulares) aciliado. A digesto extracelular e intracelular na maioria. Os cstodes precisam absorver nutrientes pr-digeridos atravs do seu tegumento, pois no apresentam sistema digestivo. A osmorregulao ocorre atravs dos protonefrdeos de clulas-flama, sendo que a eliminao de restos metablicos e a respirao acontecem atravs da parede do corpo. Com exceo de Acoela (que possuem sistema difuso), apresentam sistema nervoso do tipo escada-de-corda, com neurnios de associao, motores e sensoriais. A maioria hermafrodita e a reproduo assexuada ocorre em alguns grupos. A classe Turbellaria apresenta a maioria dos seus membros sendo carnvora e livre-natantes. Os tremtodes digneros tm um molusco como hospedeiro
intermedirio e quase sempre um vertebrado como hospedeiro definitivo. Os Digenea incluem vrios parasitas importantes de humanos e animais domsticos. Os cstodes (tnias) geralmente possuem um esclex (rgo de apreenso) em sua extremidade anterior, seguido por uma longa cadeia de progltides, cada uma das quais possui um conjunto de rgos reprodutivos de ambos os sexos. Os cstodes, quando adulto, vivem no trato digestivo de vertebrados, e as larvas encapsuladas so transmitidas pelas fezes. FILO NEMERTEA
Seu nome refere-se pontaria infalvel da probscide. Os membros dos Nemertea so normalmente menores que 20 cm de extenso e apresentam um sistema digestivo completo com um nus e sistema circulatrio verdadeiro. So de vida livre, capturando suas presas com probscide eversvel (se aloja na rincocele). Apresentam sistema vascular sangneo com dois ou trs troncos longitudinais.
FILO GNATHOSTOMULIDA Os Gnathostomulida so um filo com animais vermiformes marinhos, com 0,5 a 1 mm de comprimento, que vivem entre gros de areia e silte. No tm nus.
ANIMAIS PSEUDOCELOMADOS (Filos: Rotifera, Gastrotricha, Kinorhyncha, Loricifera, Priapulida, Nematoda, Acanthocephala e Entoprocta)
Apresentam uma pseudocele, derivada da blastocele embrionria. Consiste em um espao entre o intestino e os componentes mesodrmicos e ectodrmicos da parede do corpo e no revestida por peritnio. Nove grupos pertencem aos pseudocelomados: Rotifera, Gastrotricha, Kinorhyncha, Nematomorpha, Loricifera, Priapulida, Acanthocephala e
Entoprocta. Antigamente, os cinco primeiros eram considerados como classes em um filo chamado Aschelminthes. Porm suas relaes filogenticas diferem tanto, que cada um est um filo diferente. A maioria dos animais pseudocelomados pequena. Todos tm uma parede de corpo de epiderme, uma derme e msculos que envolvem a pseudocele. O trato digestivo completo (exceto em Acanthocephala). O filo Rotifera composto por organismos pequenos, principalmente de gua doce, com uma coroa de clios (de onde deriva o nome, pois parece uma roda girando ao bater), a qual cria correntes de gua para trazer alimentos at a boca. A boca abre-se em uma faringe muscular (mastax), equipada com mandbulas. So dioicos, e os machos normalmente so menores que as fmeas.
Os Gastrotricha, Kinorhyncha e Loricifera so pseudocelomados minsculos. Os Gastrotricha movem-se por clios ou glndulas adesivas e os Kinorhyncha ancoram-se, puxando-se pelos espinhos de sua cabea. Os Loricfera podem recolher seus corpos para dentro da lorica. Os Priapulida so vermes marinhos encontrados principalmente em guas mais frias dos hemisfrios. A maioria predadora e escavadora. Apresentam uma probscide eversvel, usada para explorar o ambiente e capturar alimento. Os sexos so separados. Os maiores e mais importantes deste grupo de filos so os nemtodes. So cilndricos, afilando-se nas extremidades e cobertos por cutcula dura. Os msculos de sua parede do corpo so longitudinais e, para funcionarem bem para a locomoo, esta conformao tem de incluir um volume de fluido na pseudocele a uma alta presso hidrosttica. A maioria dos nemtodes dioica e h quatro fases juvenis, separadas entre si por uma muda da cutcula. Quase todos os animais invertebrados e vertebrados e muitas plantas tm parasitas nemtodes. Outros nemtodes so de vida livre no solo e em habitat aquticos. Algumas graves doenas em humanos e animais so causadas por nemtodes, como por exemplo o oxiro, a lombriga (Ascaris lumbricoides), os vermes da ancilostomose. Os Nematomorpha so vermes que possuem fases juvenis parasitas de artrpodes, seguidas por um adulto de vida livre, aqutico e que no se alimenta. Os Acanthocephala so parasitas do intestino de vertebrados quando adultos, e suas fases juvenis desenvolvem-se em artrpodes. Apresentam uma probscide anterior invaginvel com espinhos, os quais fincam na parede intestinal de seu hospedeiro. Como no possuem um trato digestivo, absorvem os nutrientes atravs do tegumento. Os Entropocta so animais aquticos, pequenos e ssseis. Apresentam uma coroa de tentculos ciliados que circulam a boca e o nus.
MOLUSCOS (Filo Mollusca)
O nome Mollusca indica uma de suas principais caractersticas: corpo mole. Este grupo inclui: qutons, lesmas, caracis, borboletas-do-mar, mexilhes, ostras, lulas, polvos, nutilos e escafpodes. So encontrados numa ampla variedade de habitats. A maioria marinha. A maioria dos moluscos apresenta cabea bem desenvolvida, a qual portadora da boca e alguns rgos sensoriais. No interior da boca situa-se a rdula (rgo linguiforme raspador). A rdula apresenta dentes diminutos e est presente em todos os moluscos, exceto nos bivalves e na maioria dos solenogastres. O p dos moluscos est adaptado para locomoo, fixao a um substrato ou combinao de funes. Geralmente uma estrutura ventral, onde ondas de contrao muscular promovem a locomoo por rastejamento. O manto uma bainha de epiderme que se estende a partir da massa visceral e pende de cada lado do corpo, protegendo as partes moles e criando um espao chamado cavidade do manto (geralmente abriga brnquias). A superfcie exposta do manto tambm auxilia nas trocas gasosas. Alm disso, produtos dos sistemas digestivo, excretor e reprodutor so lanados para dentro da cavidade do manto. Para se protegerem, muitos moluscos podem recolher a cabea ou o p para dentro da cavidade do manto. Quando presente, a concha nos moluscos secretada pelo manto. A concha geralmente constituda por trs camadas: peristraco (camada mais externa), primstica (camada mediana), nacarada (camada mais interna). H um sistema circulatrio aberto. A maior parte dos cefalpodes apresenta um sistema circulatrio fechado. O trato digestivo complexo. A
maioria dos moluscos tem um par de rins (metanefrdeos). A maioria dioica, embora alguns sejam hermafroditas. A primitiva larva dos moluscos a trocfora, e muitos moluscos marinhos tm uma larva mais derivada, a vliger. As classes Caudofoveata e Solenogastres constituem um grupo de moluscos vermiformes desprovidos de concha. A classe Scapho poda (dentesde-elefante) apresenta uma concha tubular, aberta em ambas as
extremidades, e o manto envolve completamente o corpo. A classe Monoplacophora um grupo marinho univalve, exibindo pseudometamerismo.
Os Polycophora (qutons) so organismos marinhos comuns, com concha constituda por uma srie de oito placas. So sedentrios. Gastrpoda a maior classe de moluscos. Inclui: caracis, lapas, lesmas terrestres e marinhas, bzios, litorinas, lebres-do-mar e borboletas-domar. Sua histria evolutiva inclui a toro da extremidade posterior para anterior, de tal modo que o nus e a cabea situam-se na mesma extremidade; e o enrolamento, uma elongao da massa visceral, seguida do seu enrolamento em espiral. Essa toro levou a um problema de autopoluio, o que resulta na liberao de excretas sobre a cabea e diante das brnquias. Esse problema solucionado de vrias maneiras entre os gastrpodes. Um exemplo de soluo a captao de gua para dentro da cavidade do manto por um dos lados e escoamento pelo outro. Outra soluo um certo grau de destoro (opistobrnquios). Alguns tambm apresentam converso da cavidade do manto em um pulmo. Os Bivalvia so marinhos e de gua doce. Apresentam concha dividida em duas valvas unidas por um ligamento dorsal, trazidas uma de encontro outra, fechando-se por um msculo adutor. A maioria dos bivalves drena e filtra a gua atravs de brnquias por ao ciliar. Os Bivalvia incluem os: mexilhes, vieiras, ostras e teredos. Os membros da classe Cephalopoda so todos predadores e muitos podem nadar rapidamente. Incluem as lulas, polvos, nutilos e sibas. Apresentam tentculos, os quais capturam as presas por secrees adesivas ou atravs de ventosas. Nadam expelindo gua vigorosamente da cavidade do manto, fazendo-a passar atravs de um funil.
VERMES SEGMENTADOS (Filo Annelida)
O filo Annelida constitudo pelos vermes segmentados. Os mais familiares so as minhocas terrestres e de gua doce, e as sanguessugas. Os aneldeos so celomados verdadeiros. Apresentam o corpo dividido em anis (segmentos), metamerizado. Com exceo das sanguessugas, a grande maioria apresenta cerdas. Cerdas curtas
auxiliam na ancoragem durante a locomoo. Cerdas longas auxiliam na natao de formas aquticas. Os poliquetos so aneldeos marinhos. Os oligoquetos e sanguessugas ocorrem principalmente no solo e em guas continentais. Um aneldeo tpico apresenta um protostmio, corpo segmentado e uma poro terminal onde se situa o nus (pigdio). Exceto nas sanguessugas, o celoma dos aneldeos preenchido por lquido, funcionando como um esqueleto hidrosttico, muito eficaz para movimentos precisos de escavao e natao. Os Polychaeta apresentam uma regio ceflica bem diferenciada, portadora de rgos sensoriais especializados. Em cada somito apresentam muitas cerdas implantadas em parapdios. Os parapdios incluem vrias adaptaes, incluindo a especializao para nadar, respirar, rastejar, promover o fluxo dgua no interior da galeria e auxiliar na tomada do alimento. Alg uns poliquetos so predadores, possuindo uma faringe eversvel como mandbulas. Outros raramente deixam galerias onde vivem. So dioicos, apresentando um sistema reprodutor primitivo. No apresentam clitelo, a fecundao externa e a larva do tipo trocfora. A classe Oligochaeta inclui as minhocas e formas dulciaqucolas. Diferente dos Polychaeta, os Oligochaeta apresentam um nmero reduzido de cerdas por segmento e no possuem parapdios. O sistema circulatrio fechado, e o principal rgo propulsor o vaso sangneo dorsal. As
minhocas apresentam o sistema nervoso tpico dos aneldeos: gnglios cerebroides dorsais conectados a um cordo nervoso ventral duplo com gnglios segmentares, que se estende por todo comprimento do corpo. So hermafroditas, com fecundao cruzada. As sanguessugas (Hirudnea) so dulciaqucolas, porm, poucas so marinhas ou terrestres. Alimentam-se de fludos. Muitas so predadoras, algumas so parasitas temporrias e outras parasitas permanentes. As hermafroditas apresentam fecundao cruzada, com formao de casulo pelo clitelo.
MANDIBULADOS AQUTICOS (Filo Arthropoda, Subfilo Crustacea)
Os
crustceos
apresentam
dois
pares
de
antenas,
principal
caracterstica que os difere dos outros artrpodes. Muitos apresentam carrapaa. Apresentam na cabea, alm das antenas, um par de mandbulas e dois pares de maxilas, seguindo-se um par de apndices em cada segmento do corpo (somito). Em alguns crustceos, nem todos os somitos apresentam apndices. Em muitos crustceos a cutcula dorsal da cabea pode estender-se posteriormente e ao redor das laterais do animal cobrir ou fundir-se com somitos torcicos e abdominais. Esta cobertura denominada carapaa. Os hbitos alimentares variam muito em Crustacea, havendo
predadores, detritvoros, comedores de partculas em suspenso e parasitas. A respirao ocorre pela superfcie do corpo ou brnquias. A excreo ocorre atravs de glndulas antenais ou maxilares. A circulao ocorre atravs de um sistema aberto formado por seios (hemocele), e um corao tubular dorsal o principal rgo bombeador. Na classe Brachiopoda, quatro ordens so reconhecidas: Anostraca (sem carapaa. Ex: artmia), Notostraca (carapaa forma uma grande concha dorsal), Conchostraca (carapaa bivalve) e Cladocera (carapaa envolve o corpo, exceto na regio ceflica. Ex: pulgas-dgua). A maioria dos braquipodes so dulccolas. A reproduo dos Brachiopoda muito interessante: no vero produzem s fmeas partenogenticas, aumentando rapidamente a populao. Ocorrendo condies desfavorveis, alguns machos e ovos fertilizveis so produzidos. Os ovos fertilizados so altamente resistentes ao frio e dessecao. Entre os representantes da classe Maxillopoda,os membros da subclasse Copepoda no apresentam carapaa nem apndices abdominais. So abundantes e esto entre os mais importantes consumidores primrios em muitos ecossistemas dulccolas e marinhos. Muitos so parasitas. A maioria dos membros da subclasse Cirripedia (cracas) so ssseis quando adultos, secretam uma concha calcrea e filtram alimento atravs de seus apndices torcicos. A maior classe dos crustceos a Malacostraca, sendo suas ordens mais importantes: Isopoda (tatuzinhos-de-quintal, Lgia), Amphipoda (ex:
salto-de-praia), Euphausiacea (krill) e Decapoda (lagostins, lagostas, siris, caranguejos, camares). Os ispodes e Amphiopoda no apresentam carapaa. Os eufasiceos so parte importante do plncton ocenico. Os Decapoda apresentam cinco pares de patas ambulatrias.
8. KORMONDY, E. J.; BROWN, D. E. Ecologia humana. So Paulo: Atheneu, 2002. PARTE 1 EVOLUO: O ANTES E O AGORA
INTRODUO Pode parecer estranho iniciar um livro sobre ecologia humana com uma discusso sobre evoluo. Entretanto, para adquirir algum conhecimento sobre as formas nas quais as diferentes populaes humanas se adaptaram aos seus diversos ambientes, importante entender como ocorrem as adaptaes. Mas tambm precisamos entender como populaes humanas to diferentes como os inuits do plo norte e os tuareg do deserto do Saara apareceram. Assim, necessitamos de uma ideia de variao intra-especfica e tambm da origem e evoluo das espcies.
AS PRINCIPAIS TEORIAS EVOLUTIVAS Para explicar o fenmeno da mudana constante dos organismos ao longo do tempo, alguns cientistas propuseram vrios esquemas e mecanismos racionais, embora altamente especulativos. Devido s suas influncias no passado e presente, apenas duas das mais importantes ideias so aqui rapidamente discutidas: a herana de caracteres adquiridos e a evoluo por seleo natural.
A herana de caracteres adquiridos Significa dizer que, quando o ambiente se torna muito diferente, produz, ao longo do tempo, modificaes correspondentes na forma e organizao dos organismos. Para o caso do aparecimento de novas caractersticas, devidas ao uso, em organismos, Lamarck se utilizava do caso dos pssaros que vivem em
ambientes aquticos e que teriam separado seus dgitos a fim de movimentarem-se melhor na gua. Antlopes, herbvoros ancestrais esticavam seus pescoos, lnguas e pernas a fim de obterem a maior quantidade de folhas possvel, consequentemente alongando essas estruturas. As pores corpreas adquiridas, maiores, eram ento sucessivamente passadas s geraes subsequentes, as quais seguiam o mesmo processo, chegando s conhecidas girafas de pescoos, lnguas e pernas alongadas.
Evoluo pela Seleo Natural Assim como indivduos de uma dada espcie nascem em maior nmero que aqueles que podem possivelmente sobreviver; e como, consequentemente, h uma constante luta pela sobrevivncia, segue que qualquer ser, se variante, mesmo que de forma sutil e benfica para si prprio, sob as complexas e, s vezes, variveis condies da vida, ter uma melhor chance de sobrevivncia e, assim, ser naturalmente selecionado. A partir do forte princpio da herana, qualquer variedade selecionada tender a propagar sua nova e modificada forma.
A ORIGEM DAS ESPCIES Isolamento Reprodutivo Para continuar com a origem estabelecer o que uma espcie . uma espcie uma populao produzem descendentes frteis e populaes. das espcies, precisamos inicialmente Para organismos de reproduo sexuada, na qual os indivduos se reproduzem, no tm sucesso reprodutivo com outras
A impossibilidade de uma populao de se reproduzir com outras conhecida como isolamento reprodutivo e pode ocorrer de diversas maneiras. Pode haver diferenas quanto estrutura e funo dos rgos reprodutivos (por exemplo, diferenas do arranjo das pores florais que permitem a polinizao por uma dada espcie de inseto em um caso e no em outro), as clulas sexuais podem ser incompatveis (por exemplo, os ovos liberam uma substncia qumica que atrai o esperma de uma populao e repele o de outras), pode haver diferenas quanto ao perodo no qual a reproduo ocorre (primavera para uma populao e vero para outra) ou pode haver diferenas comportamentais (por exemplo, a corte masculina atraindo fmeas da prpria populao e no de outras).
Embora a maioria dos cientistas acredite que o isolamento geogrfico o principal mecanismo de especiao, h um crescente nmero de evidncias de que novas espcies so formadas bem no meio da populao parental. Seleo direcional (algumas vezes chamadas de seleo diversificadora) resulta em mudanas mais ou menos regulares das respostas s alteraes do meio ambiente ou at mesmo de um novo ambiente. A seleo sexual ocorre quando h diferenas na aparncia e/ou comportamento entre os dois sexos, um fenmeno especialmente dramtico em muitos pssaros e mamferos.
GRADUALISMO E INTERMITNCIA NA EVOLUO O processo que descrevemos sugere, em geral, que tanto a origem quanto a subsequente evoluo das espcies ocorre vagarosamente. Gradualismo o termo mais bem aplicado a estas mudanas que resultam dos vrios mecanismos de isolamento e seleo que j discutimos. Sequncias fsseis de alguns grupos demonstram este fenmeno de modificao gradual ao longo do tempo. Entretanto, o gradualismo falha ao explicar o substancial nmero de casos no registro fssil nos quais novas espcies so morfologicamente distintas em seu primeiro momento de apario, bem como nos casos de ausncia de formas intermedirias em muitos grupos de animais. De acordo com a teoria do equilbrio intermitente, a maioria das mudanas morfolgicas ocorre durante a especiao em processos tais quais o efeito fundador e a seleo direcional. Isto , ento, seguido por um perodo de seleo estabilizadora. Por exemplo, uma vez que apareceu, o casco do cavalo no demonstrou praticamente qualquer variao morfolgica em 500 milhes de anos, a rvore ginkgo no demonstra nenhuma alterao h 200 milhes de anos e o opossum, h 70 milhes de anos. Estes organismos so exemplos de grupos em xtase evolutivo, ou em equilbrio. Pode parecer que o acmulo de evidncias crescentemente apoia o conceito do equilbrio pontuado como o principal modo de evoluo. Entretanto, isso no destri o conceito de gradualismo, mas sim fornece uma explicao alternativa para alguns eventos evolutivos. O gradualismo evolutivo realmente ocorre e foi muito bem documentado em muitas espcies; em outras, grandes saltos parecem ter ocorrido em perodo breves de tempo. De qualquer forma, a seleo natural opera em ambos os modelos rpida ou gradualmente.
ADAPTAO A adaptao pode ser definida a partir de uma perspectiva evolutiva, como o desenvolvimento ou melhora de caractersticas estruturais, comportamentais e/ou fisiolgicas que aumentam as chances de sobrevivncia e de reproduo em um dado ambiente.
A ORIGEM DOS SERES HUMANOS A teoria da savanizao, para explicar a origem dos homindeos, vem sendo seriamente questionada nos ltimos anos.
Origens Humanas Humanos so classificados como primatas, um grupo que apareceu aproximadamente 65 milhes de anos AP, mais precisamente durante o Paleoceno (datado entre 65 milhes e 54 milhes AP, simultaneamente a uma grande irradiao adaptativa dos mamferos placentrios). Evidncias fsseis indicam que os primeiros primatas apareceram a partir de insetvoros pequenos, terrestres, noturnos e adaptados vida sobre as rvores. Estes se tornaram os primatas primitivos, que deram origem ao que hoje conhecemos como prosmios, grupo no qual esto includos os lmures, os lerises e, possivelmente, os trsios. Durante o Oligoceno (de 38 milhes a 25 milhes de anos AP), os primatas passaram por mais uma radiao adaptativa que levou, finalmente, aos trs principais ramos de primatas antropoides geralmente reconhecidos: os macacos do novo mundo (Ceboides), os macacos do velho mundo (Cercopitecoides) e os smios/humanos (Hominoides). E neste ltimo que a nossa ateno estar concentrada. Os primeiros primatas superiores definitivamente aceitos datam de cerca de 35 milhes de anos AP (no Oligoceno), quando tanto os platirrinos do Novo Mundo (nariz achatado, com as narinas apontando para as laterais) quanto os catarrinos (nariz estreito, com as narinas apontadas para baixo) so encontrados. O Aegyptopithecus (smio do Egito), um representante dos catarrinos do Oligoceno, possua o padro da cspide do molar de um smio (cinco cspides em vez das quatro encontradas em macacos do Velho Mundo), a estrutura quadrpede bsica, um rabo externo de um macaco e o tamanho cerebral de um lmur. O Aegyptopithecus um candidato posio de o mais antigo dos catarhines.
Os Primeiros Homindeos. A linhagem hominoide divergiu em algum momento entre 5 milhes e 8 milhes de anos AP, formando os pongdeos, grupo que inclui os gorilas e chipanzs modernos e seus ancestrais, e os homindeos, humanos modernos e ancestrais. Outros smios hominoides, como o gibo e o orangotango divergiram anteriormente. Os homindeos caracterizam-se pela locomoo bpede, dieta onvora e, posteriormente, pela capacidade craniana aumentada e organizao social, todas as trs consideradas como adaptaes a vida nos campos abertos das savanas em vez das florestas protegidas, como seus precedentes. Australopithecus afarensis, o mais antigo homindeo conhecido, formado por ossos fossilizados apelidados de lucy por seu descobridor, Donald Johans on, datado em 3,1 milhes de anos, tinha apenas 1,1 metro de altura, pesava cerca de 30 kg e possua uma mandbula pronunciada e o crebro semelhante ao de um chimpanz. Lucy, com vinte e cinco anos, tratava-se claramente de um homindeo quando era considerado seu andar ereto sobre pernas que podiam ser completamente estendidas. Ardipithecus ramidus, em 1992-1993, remanescentes fsseis da mais e remota espcie de homindeos, possivelmente a raiz dessas espcies, forma descobertos na Etipia e so conhecidos agora como Ardipithecus ramidus. Australopithecus anamensis, engordando a lista dos primeiros homindeos, datado em 3,9 milhes de anos, um excelente candidatos entre A. Ramidus e o A. Afarensis. Ecotonos e os primeiros homindeos. Estes primeiros australopitecneos devem ter vivido nas reas de transio entre florestas e savanas. Tais transies entre dois (ou mais) tipos de ambiente so conhecidos como ecotonos. geralmente aceita a ideia de que a vida neste ecotono floresta savana contribuiu para o desenvolvimento da bipedia, dos hbitos alimentares onvoros e da encefalizao. Australopithecus africanos, A linhagem dos africanos significativa, porque alguns cientistas acreditam que ela deu origem ao maior dos passos na evoluo humana, o Homo habilis, entre 2 milhes e 2,5 milhes de anos AP, e, talvez, h outras linhagens, as quais incluiriam Homo rudolfensis e at o Homo erectus. Homo habilis, originou-se do Australopithecus h cerca de 2,5 milhes de anos AP e se extinguiu 1 milho de anos depois, ele era muito semelhante ao seus ancestrais mais simiescos, exceto por ser mais alto e possuindo um crnio maior, parece ter feito suas ferramentas lascando as pedras a fim de dar-lhes um formato desejado.
Embora tenha existido durante apenas meio milho de ano, H. Habilis possivelmente deu origem a uma nova espcie, o Homo erectus, da qual os primeiros esqueletos datam de aproximadamente 1,8 milhes de anos AP. Homo erectus, medindo, em mdia, 1,7 metros de altura, reteve o crnio simiesco (testa achatada, toro supra-orbital pronunciado) e mandbula (sem queixo) de seus predecessores, mas seu crnio havia aumentado em relao ao crescimento total do corpo, levando a um maior desenvolvimento cerebral, o que forneceu a plasticidade necessria para que se adaptasse aos diferentes ambientes e para sua ampla disperso. Homo sapiens, so divididos em dois grupos: arcaicos e modernos. Ambas as formas possuem crebro grande, em mdia 1.350 cm (s vezes, alguns indivduos primitivos superavam esta mdia). Formas arcaicas nas quais est inclusa a subespcie Homo sapiens neanderthalensis, so caracterizados por crnios longos e baixos e face robusta, com toro supra-orbital moderado e mandbulas grandes. O Homo sapiens moderno caracteriza-se por um crnio mais arredondado e alto, presena de queixo, dentes e mandbulas relativamente pequenas e uma face mais grcil (menos robusta) que a das formas arcaicas. E foram as formas modernas de Homo sapiens que tiveram um extraordinrio impacto sobre o seu meio ambiente. A elas tem sido atribudo um papel importante na extino de muitas das espcies de mamferos no final do Pleistoceno, bem como nas maiores modificaes de paisagem iniciada pelo desenvolvimento e intensificao da agricultura e de outras atividades.
PARTE 2 CONCEITOS BSICOS EM ECOLOGIA
INTRODUO De uma derivao do grego, ecologia literalmente significa o estudo da casa. Mas, de forma mais ampla, esta palavra se refere ao estudo cientfico do ambiente, incluindo seus componentes orgnicos e inorgnicos. Na medida em que seus praticantes descrevem, classificam, criam hipteses e atestam, a ecologia molda-se aos padres cientficos. Entretanto, graas ao seu envolvimento com a geologia, a fsica, a qumica e a matemtica, suas fronteiras disciplinares so permeveis; juntas, estas caractersticas fazem da ecologia um alvo para diferentes canais de questionamento cientfico, desde a abordagem reducionista, como o estudo das espcies individualmente, at a aproximao holstica, que investiga a totalidade dos ambientes no planeta Terra.
DEFININDO ECOLOGIA A ecologia tem recebido interpretaes diferentes. Charles Elton (1927), um eclogo britnico, a definiu como histria natural cientfica, preocupada com a sociologia e ecologia dos animais. O eclogo vegetal americano Frederick Clements (1905), considerou-a a cincia da comunidade, e o eclogo alemo Karl Friederichs (1958), como a cincia do ambiente. Estes, de certa forma, diferentes focos tem sido abrangidos pelo conceito desenvolvido pelo eclogo americano Eugene Odum, que definiu ecologia como o estudo das estruturas e funes da natureza e depois, como o estudo da estrutura e funes dos ecossistemas. Sendo esta definio operante e atual.
ECOSSISTEMA Ecossistemas podem ser muito grandes, como uma floresta ou um deserto, ou muito mais circunscrito, como um aqurio ou um tubo de ensaio. Determinar as fronteiras de um ecossistema , de certa forma, arbitrrio, mas no aleatrio.
SISTEMAS Consiste de dois ou mais componentes que interagem e que so cercados por um meio ambiente com o qual podem ou no interagir.
Componentes dos ecossistemas No caso dos sistemas ecolgicos, os dois principais componentes so o biticos (vivos) e o abiticos (no vivos).
A ORGANIZAO DOS ECOSSISTEMAS
Produtores, Consumidores e Decompositores Produtores so o primeiro passo de um movimento de energia atravs de um ecossistema. este grupo de organismos, normalmente verdes e clorofilados, que capturam a energia radiante do sol, na verdade produtores no produzem energia: eles convertem ou a transmitem de uma forma, a radiante, a outra, qumica.
Um auttrofo (literalmente, auto-alimentador) um organismo que captura energia e, subsequentemente, a utiliza para sintetizar molculas que servem aos requerimentos nutricionais de seu prprio crescimento e metabolismo. Em contraste, um hetertrofo (literalmente, alimentado por outro) um organismo cujas necessidades nutricionais so atingidas atravs da ingesto de outros organismos: hetertrofos so tambm conhecidos como consumidores. Consumidores so organismos que derivam sua nutrio dos produtores: ou seja, so hetertrofos. As correlaes auttrofo-hetertrofo, ou produtor-consumidor primrio e consumidor secundrio, resultam em um movimento sequencial de energia conhecido como a cadeia alimentar. Decompositores, basicamente bactrias e fungos, so hetertrofos que derivam sua nutrio a partir de produtores (a maioria dos fungos) e consumidores (principalmente as bactrias). Em vez de ingerir seu alimento, como no caos dos outros hetertrofos, os decompositores liberam enzimas de seu corpo capazes de digerir os tecidos animais e vegetais, em seguida, esses produtos processados so absorvidos.
NICHOS E HABTATS
Nicho O fluxo energtico e a ciclagem de nutrientes no ocorrem em um conjunto abstrato dos produtores, consumidores e decompositores. Estes termos que cada espcie possui um papel particular, seno nico, em um dado ecossistema. A soma total destes atributos ecolgicos um nicho do organismo, seu papel nos processos ecolgicos o que ele faz, onde est e o que se faz a ele. Algumas espcies so uma fonte energtica apenas para outras certas espcies, enquanto outras atendem a um maior e mais variado nmero de componentes.
Habitat Qualquer lugar que seja seu nicho, a espcie vive e excuta suas funes em algum lugar. Este lugar seu habitat. O habitat de um rato canguru o deserto, o de um urso polar o rtico; uma dimenso do nicho do rato canguru a de um herbvoro, enquanto a de um urso polar a de um carnvoro.
Espcie, Populaes e Comunidades Como foi observado, uma espcie uma populao de indivduos que so e permanecem distintos porque normalmente no se reproduzem com outros indivduos, ou seja, outra espcie. A populao da espcie pode estar amplamente distribuda como no caso dos humanos. Resumidamente, uma comunidade ecolgica compreende seus componentes biticos (plantas, animais e micrbios), de um dado ecossistema ou de uma subunidade dele.
PARTE 3 ABORDAGENS EM ECOLOGIA HUMANA
A BASE ANTROPOLGICA A antropologia pode ser definida como o estudo da diversidade biolgica e comportamental humana, tanto geogrfica como temporalmente. Diversidade, ou variao, em populaes um importante aspecto da evoluo, assim, a evoluo torna-se um dos principais pontos da antropologia. Os antroplogos estudam toda a variabilidade humana normal (e tambm as patologias) sobre a Terra tanto no presente quanto no passado. Neste ltimo caso, a antropologia pode ser definida como o estudo da histria natural da espcie humana.
Antropologia Ecolgica A Antropologia Ecolgica estuda a ecologia de populaes humanas ao redor do mundo, no passado e no presente. Seu objetivo entender como a ecologia destas populaes influencia, direta ou indiretamente, as caractersticas culturais associadas a cada grupo.
DETERMINISMO AMBIENTAL
Abordagens Pioneiras A ideia de que caractersticas ambientais tm um efeito sobre a presena de traos culturais especficos chamada de determinismo ambiental. Esta provavelmente a mais antiga das abordagens ecologia humana, j que ela claramente engloba a viso das relaes humanos-ambiente de Hipcrates.
reas Culturais reas culturais referem-se s regies do mundo dentro das quais as condies ambientais e as culturas das populaes humanas so similares. Exemplos de reas culturais so a Polinsia e, tradicionalmente, as regies das grandes plancies americanas.
Limitaes do Determinismo Uma grande desvantagem do determinismo ambiental a maneira simplista de conectar as caractersticas ambientais com os traos culturais. O ambiente, de certa forma, visto como precedendo a cultura e, assim, capaz de determin la. Muitos deterministas eram mais sofisticados em suas abordagens, notando certas complexidades nas relaes ambiente/cultura. POSSIBILISMO AMBIENTAL
O ambiente era importante apenas ao limitar as possibilidades de uma cultura por isso o termo possibilismo ambiental. Para os possibilistas, culturas em uma mesma rea cultural eram similares devido s conexes histricas, no por influncias ambientais simplistas.
Similaridades entre o Determinismo Ambiental e o Possibilismo Ambiental No possibilismo, as culturas escolhem entrem as alternativas, com o ambiente determinado a variao das alternativas. Se as alternativas se limitam a uma nica opo, o possibilismo reduzido ao determinismo. De acordo com os possibilistas, a cultura toma o papel principal de determinar as mudanas culturais posteriores na maioria dos ambientes, mas a causa da diversidade cultural comea com o ambiente e essencialmente um processo de mo nica.
ECOLOGIA CULTURAL
A essncia de sua noo de ecologia cultural que o meio de subsistncia de um grupo (as principais maneiras de se obter alimentos) est diretamente relacionada com os outros aspectos de sua cultura. Ele se referia
subsistncia de um grupo, incluindo sua economia bsica e tecnologia, como o ncleo cultural. O ncleo cultural influencia a organizao cultural e social de um grupo.
Ecologia Sociolgica A ecologia sociolgica baseada primeiramente no uso de uma analogia entre os ambientes naturais e aqueles feitos humanos. Por exemplo, a partir dos trabalhos de Park, a sociedade urbana tem sido analisada com base em uma analogia ao ecossistema.
Abordagens Psicolgicas A psicologia ecolgica tenta prever os comportamentos baseada em observaes cuidadosas, geralmente quantitativas, do ambiente no qual o comportamento ocorre. Em outras palavras, o objetivo entender as correlaes entre os tipos de unidades ambientais e os comportamentos comuns, com a noo de que pelo menos algumas dessas unidades ambientais tm grande poder de coero sobre os comportamentos que ocorrem dentro delas.
Abordagens da Arquitetura e do Planejamento Urbano Certas abordagens da arquitetura e do planejamento aplicam algumas das ideias geradas pela sociologia e psicologia no estudo da ecologia humana, particularmente a noo de que os ambientes fsicos artificiais podem ser muito influentes na determinao dos processos sociais e dos comportamentos individuais dentro deles.
ABORDAGENS BIOLGICAS E EVOLUTIVAS ECOLOGIA HUMANA
A teoria evolucionria de Darwin levou a uma nova perspectiva sobre as relaes dos humanos com seu ambiente. A noo de adaptao ao ambiente tornou-se um conceito importante no estudo da evoluo humana, agrupando abordagens ecolgicas e evolutivas. No final dos anos 40, eclogos humanos comearam a integrar estas abordagens ao estudo das culturas humanas.
O Paradigma da Nova Ecologia O paradigma da nova ecologia representa uma tentativa dos antroplogos culturais de reintegrar as anlises das adaptaes culturais com os estudos gerais da ecologia. Problemas ambientais. Um tema central do paradigma da nova ecologia a ateno aos problemas ambientais, seus efeitos sobre as populaes humanas e as respostas das pessoas a eles. Abordagem ecossistmica. A Abordagem ecossistmica da antropologia ecolgica baseada em uma simples premissa: as populaes humanas devem ser vistas num contexto total dos ecossistemas nas quais vivem. A Biologia de Populaes Humanas O principal objetivo o de entender a adaptabilidade humana tanto populacional quanto individual, estimulando a integrao das pesquisas em gentica, demografia, crescimento e desenvolvimento, epidemiologia, fisiologia ambiental, aquisio de recursos e nutrio, bem como os processos comportamentais e culturais que se relacionam aos problemas ambientais. Assim, a Biologia de populaes humanas, como o paradigma da nova ecologia, integra as abordagens socioculturais e biolgicas da ecologia humana.
PARTE 4 ECOLOGIA DE POPULAES
INTRODUO At mesmo a partir de suas observaes mais triviais voc, provavelmente, j sabe que h modificaes em vrias populaes de animais e plantas em seu ambiente. Alguns tipos de plantas somente florescem na primavera e outras, no outono: rvores decduas perdem suas folhas no outono e as recuperam na primavera: moscas varejeiras esto por toda parte praticamente ao longo de todo o ano, mas formigas e abelhas incomodam principalmente durante os meses de vero. Talvez voc tambm tenha notado que, em alguns anos, pestes de animais como mosquitos so mais abundantes, flores no crescem to bem, a menos pssaros cantando e menos peixes nos rios. Em um sentido muito geral, todas essas mudanas so manifestaes da dinmica do crescimento e da regulao das populaes.
CRESIMENTO POPULACIONAL
Potencial Bitico e Resistncia Ambiental Inerente a caractersticas de populao, sejam moscas varejeiras, elefantes, pardais ou humanos, a capacidade de reproduo de uma dada taxa. Isso geralmente conhecido como potencial bitico de uma espcie. Em oposio ao potencial bitico encontra-se a totalidade das forcas ambientais, biticas e abiticas, que impedem que o potencial mximo seja alcanado. Essas forcas so conhecidas como resistncia ambiental e so responsveis pela manuteno dos nmeros populacionais. Ambos os termos foram introduzidos pelo eclogo americano Royal Capman (1928). Ocasionalmente, essas forcas falham, o que resulta em exploses populacionais tais como as pragas bblicas do deserto e outras mais recentes. Primeiro, consideraremos o potencial bitico das populaes, um resultado da interao da taxa de nascimento, ou natalidade, e da taxa de mortes, ou mortandade. Mais sobre o Potencial Bitico O potencial reprodutivo de uma espcie no , entretanto, uma constante. Seu valor diferente sob diferentes condies ambientais, abiticas ou biticas. Fatores abiticos que influenciam o potencial reprodutivo incluem a temperatura, a umidade e a disponibilidade de nutrientes; fatores biticos incluem o nmero de indivduos da populao presente em uma dada rea, ou seja, a densidade da populao, a idade dos indivduos, a idade ativa, o final da idade reprodutora e o perodo da vida. Taxa intrnseca do Crescimento Natural medido sob condies timas, potencial bitico de uma populao. Taxa Reprodutiva de Rede. Uma dimenso adicional do potencial reprodutivo, ou seja, o nmero de descendentes fmeas que substitui cada uma das fmeas da gerao anterior.
MORTALIDADE E SOBREVIVNCIA
Voc j deve saber que o custo de um seguro de vida aumenta conforme a idade do segurado, j que as probabilidades de morte aumentam com esse fator. Para determinar a provvel expectativa de vida de um comprador de seguros e, portanto, o custo da aplice o agente consulta uma tabela de riscos, acidente etc. esta tabela estatstica apresenta a probabilidade de sobrevivncia de certa idade baseada em fatores como idade, sexo e, s vezes, raa.
ESTRUTURA ETRIA Implcita com resultado dos diferentes padres de sobrevivncia a diferena estrutura etria de uma populao, as propores relativas entre jovens, meia idade e indivduos mais velhos de uma populao dependendo do tipo de padro de sobrevivncia caracterstica de seu grupo.
Principais Faixas Etrias Como foi proposto por Bodenheimer (1958), trs faixas etrias so conhecidas em populaes: a pr-reprodutiva ou em expanso, a reprodutiva ou estvel e a ps-reprodutiva ou decrescente. Populao e expanso: a taxa de nascimento alta e o crescimento populacional pode ser exponencial. Populao estvel: quando a taxa intrnseca de nascimento natural se aproxima de 0 e a taxa reprodutiva de rede adquire valores prximos a 1, a taxa de crescimento desacelerada e, ento, se estabiliza. Populao decrescente: quando a taxa de nascimento radicalmente reduzida.
REGULAO POPULACIONAL J demonstramos brevemente que dois grandes fatores do padro de crescimento de uma populao, a natalidade e a mortalidade, esto sujeitos as influencias ambientais biticas e abiticas.
O Papel dos Fatores Abiticos Nutrientes. Embora no seja sempre o caso, estudos envolvendo um bom nmero de espcies demonstraram que a quantidade e a disponibilidade de nutrientes particulares podem ter um papel crtico sobre a regulao da distribuio e do crescimento de uma populao. Fatores Fsicos: caracterizados por gradaes tambm tm um papel significativo sobre a regulao populacional de diferentes espcies (seca, enchentes, alimentos, clima).
O Papel dos Fatores Biticos
Interaes entre espcies envolvem um bom nmero de relaes diferentes, todas podendo ser includas dentro do termo simbiose, o qual, a partir de suas origens gregas, significa simplesmente vivendo junto. Em alguns casos, as duas populaes simbiticas no afetam uma a outra - neutralismo; na maioria dos casos h um grande ou pequeno impacto. A competio pode envolver uma inibio mtua ou um efeito indireto em situaes nas quais um recurso comum (comida) escasso. Predao. Em diversos aspectos, as relaes hospedeiro-parasita e presapredador so similares: o parasita e o predador so beneficiados, enquanto o hospedeiro e a presa, respectivamente, so adversamente afetados em maior ou menor grau. Competio interespecfica. Se duas populaes de diferentes espcimes requerem um mesmo recurso (nutrientes, espao, luz ou umidade), que potencialmente limitado ou que assim se tornam, elas so ditas competidoras entre si.
PARTE 5 A POPULAO HUMANA
CRESCIMENTO POPULACIONAL GLOBAL
Padro de Crescimento O crescimento total da populao humana at agora tem sido exponencial: de fato, ele se parece assustadoramente com uma curva em forma de J. O tempo de duplicao indica o nmero de anos de que a populao necessita para que dobre em tamanho se as taxas de crescimento presentes forem mantidas. Crescimento Populacional Regional. Enquanto a imagem global do crescimento populacional humano possui algumas caractersticas claras, aquelas das populaes regionais ou locais so bem diferentes. Estrutura etria regional. Essas grandes diferenas com relao aos padres das curvas de crescimento so refletidas, como esperado, em suas estruturas etrias. Caracteristicamente, como voc pode observar cerca de 40% da populao encontra-se com menos de 15 anos de idade nos pases em desenvolvimento. Idade mediana. Embora as pirmides de populao mostrem a forma geral da estrutura etria, elas no provem taxas ou medidas que possam ser
comparadas, ao longo do tempo, com outras populaes. Entretanto, a idade mediana, a idade exata na qual metade jovem e metade velha, fornece tal medida. Em geral, sociedades em desenvolvimento possuem populaes em crescimento rpido com baixas idades medianas, enquanto sociedades desenvolvidas possuem populaes com baixo crescimento ou estabilizadas com idades medianas altas.
NATALIDADE E MORTALIDADE Com o cenrio do crescimento populacional mundial do passado e projees futuras, podemos avaliar as duas forcas por trs das mudanas: a natalidade, ou taxa de nascimentos, e a mortalidade, ou taxas de mortes. Tanto a natalidade quanto a mortalidade so calculadas sobre nmeros por mil indivduos por ano: assim, a natalidade o nmero de nascimentos por mil pessoas por ano e a mortalidade o nmero de mortes por mil indivduos por ano.
Mudanas na Expectativa de Vida Expectativa de vida o tempo de vida estimado no momento do nascimento. A expectativa de vida mundial, por exemplo, aumentou de 61 anos em 1980 para 64 em 1990.
Mortalidade Global As taxas de mortalidade infantil em 1980 variavam largamente entre os pases desenvolvidos e em desenvolvimento.
A Natalidade em uma Escala Global A discrepncia do declnio da natalidade e da mortalidade entre os pases e rgios desenvolvidos e em desenvolvimento foi notada. Enquanto a mortalidade semelhante para reas desenvolvidas e em desenvolvimento, cerca de 10, a natalidade chega a ser o dobro, prxima a 30, em rgios de desenvolvimento e 15 para aquelas j desenvolvidas.
IMPLICAES DO CRESCIMENTO POPULACIONAL
Deveria ser intuitivamente evidente que a exploso de crescimento das populaes humanas carrega consigo uma srie de implicaes culturais e ambientais. Mais pessoas criam a necessidade de mais moradias. Mais escolas e mais cuidados, bem como mais comida e uma melhor distribuio destes recursos, mais gua; h mais lixo, a maior necessidade de combustvel mais, mais e mais. Mais pessoas em um dado espao podem levar a um descontrole poltico e social e a emigrao. Nesta poca de interconexes globais, tais complicaes em alguns locais causam problemas em outros.
ALIMENTANDO A POPULAO HUMANA
A Expanso da Fome e da Desnutrio O banco mundial estimou que mais de 700 milhes de pessoas, cerca de 13% da populao mundial, sofre com a falta de alimentos para uma vida ativa e saudvel, com as maiores concentraes na sia e na frica subsaariana. A desnutrio resulta da falta de comida, uma deficincia de um ou mais nutrientes essenciais (protena, clcio e vitamina C) ou de doenas genticas ou ambientais que interferem na digesto, absoro ou metabolismo. A capacidade da Terra. A quantidade de terras areveis e cultivveis no mundo chega a quase metade da superfcie, excluindo as pores cobertas de gelo da Antrtica e da Groelndia. Menos da metade de toda terra arevel e mais da metade da terra cultivvel esto atualmente em uso.
DESERTIFICAO
A expanso da desertificao Em um contexto geolgico, a desertificao, processo de formao e expanso de desertos, tem sido resultado de alteraes climticas; em um contexto contemporneo, a desertificao tem origens antrpicas. A histria geolgica responsvel pela presena dos grandes desertos do mundo: os hiperrios, os ridos e os semi-ridos. Atividades agrcolas humanas so responsveis pelo deserto de sal que hoje existem em regies onde um dia florescia o vale dos rios Eufrates e Tigres.
Fatores causais da desertificao
Dentre os povos criadores, o pastoreiro a principal causa da desertificao, particularmente em situaes nas quais as secas prolongadas. O desmatamento de florestas para obteno de combustvel, removendo a proteo natural contra ventos a fonte de uns do solo, outro fator, o qual extenuado pela necessidade das populaes pelo produto. O cultivo e a irrigao, o pasto em excesso, o desmatamento, a minerao, a recreao e a urbanizao so todos fatores envolvidos na desertificao.
PARTE 6 O ESTRESSE E A FISIOLOGIA AMBIENTAL
O grupo humano deve ser capaz de perceber e avaliar fatores estressores, bem como as respostas. Precisam tambm equilibrar os custos e os benefcios de curto e longo prazo. Uma das tarefas do estudioso da ecologia humana comparar um tipo qualquer de estratgia que seria tima segundo as previses de modelos tericos de mdio alcance com aquilo que o grupo na realidade faz. De fato, muitos modelos ecolgicos tm como seu propsito a identificao da melhor, ou tima, estratgia para um indivduo ou populao. Essas previses tem pouco significado para o mundo real se os organismos na verdade no se comportarem de uma maneira tima ou pelo menos prxima da tima. Os estudiosos da ecologia humana tm minimizado estes problemas ao focalizarem muitos de seus estudos em populaes sob estresses ambientais intensivos, situao em que sobra pouco espao para respostas adaptativas deficientes. Estudos ecolgicos de como os organismos adaptam-se ao estresse adquiriram uma importncia mais do que apenas acadmica. A alta taxa de espcies em extino a subsequente perda na biodiversidade global em parte uma decorrncia de mudanas ambientais causadas pela humanidade. A taxa dessa perda pode acelerar-se devido s crescentes alteraes introduzidas pelas atividades humanas, algumas atuando numa escala global. As ideias para conservao de espcies devem fundamentar-se em conceitos slidos de resistncia ao estresse biolgico, alm de outras consideraes.
PARTE 7 ADAPTAO HUMANA AO FRIO E AO CALOR
INTRODUO
Cada espcie possui uma capacidade de tolerncia a temperatura ambiental, tanto as temperaturas quentes quanto as frias requerem processos adaptativos e apresentam limites, os quais limitam a capacidade funcional dos indivduos. Estudos sobre os efeitos de condies macroambientais de calor e frio sobre humanos ilustram as similaridades e diferenas entre espcies humanas e outros animais quanto ao seu processo de adaptabilidade.
Troca de Calor Conduo se refere transferncia de calor entre dois objetos slidos em contato fsico. Conveco se refere troca de calor entre um objeto e um fluido (gs ou lquido), mais de uma vez devida a colises musculares. A radiao se refere a troca de energia eletromagntica entre objetos ao alcance um do outro. A evaporao se refere a perda de um objeto devido a converso de um liquido em um gs em sua superfcie.
MACROAMBIENTES FRIOS
Os humanos se adaptam a muitos tipos de macroambientes frios. Estes incluem condies noturnas de frio nos habitat tropicais e subtropicais ridos, noites frias em grandes altitudes e pocas frias encontradas no rtico e em muitas latitudes temperadas. Estas condies apresentam alguns perigos para humanos no protegidos, representando, porm, mais um desconforto do que ameaa a vida para os povos contemporneos. Ambientes tropicais tambm possuem perodos de chuvas e ventos frios; entretanto, geralmente no so ameaas srias para os humanos modernos. Habitaes Habitaes, ou abrigos em geral, so uma outra grande adaptao cultural ao frio dos macroambientes. Como as roupas, o fator crtico a proteo, a qual minimiza a perda de calor pela radiao e conveco. O lcool e Outros Agentes
Os humanos tambm so conhecidos pela ingesto de lcool ou de outros agentes farmacolgicos em sua ao contra o frio. O lcool fornece benefcios em curto prazo no frio: metabolizado rapidamente, aumentando a gerao de calor pelo corpo. A longo prazo, entretanto, ele prejudicial pois estimula a dilatao das veias superficiais. A vasodilatao leva a um crescimento da taxa de perda de calor do corpo, diminuindo efetivamente a proteo biolgica contra o frio. Fogo O beneficio do uso do fogo em um ambiente frio so bvios. Porm, eles tem um custo: combustvel. Alguns ambiente gelados so localizados acima das regies onde a madeira necessitando de combustvel obtido a partir de outras fontes, como as lmpadas alimentadas com leo de foca utilizada pelos inuit e fogueiras de esterco usadas pelos quchuas.
MICROAMBIENTES FRIOS
A maioria dos microambientes humanos podem ser descritas como tropical ou subtropical, apesar das mais diferentes condies macroambientais.
RESPOSTAS BIOLGICAS AO FRIO
H duas formas principais de adaptao biolgica ao estresse provocado pelo frio: o isolamento trmico e o metabolismo. A adaptao do isolamento trmico envolve o tamanho do corpo, sua forma e sua composio, bem como a regulao do fluxo sanguneo. A adaptao metablica envolve o metabolismo basal, bem como a gerao de calor metablico pela atividade muscular, tanto voluntria quanto involuntria.
MACROAMBIENTES QUENTES E SECOS
H dois tipos principais de macroambientes quentes: os quentes e secos e os quentes e midos. Os problemas e as respostas adaptativas so to diferentes nestes tipos de condies que os consideraremos separadamente.
Ambientes quentes e secos incluem os desertos e as savanas, ambos apresentando estas condies ao longo de todo ano, e tambm desertos e campos mais temperados os quais so sazonalmente quentes e secos. A aridez destes ambientes permite altas taxas de ganho de calor a partir da radiao solar devido ao ar seco e limpo, com temperaturas ao meio dia nos desertos atingindo at 57 C. O alto ganho de calor leva tambm ao movimento rpido do ar, aumentando o ganho convectivo de calor.
MICROAMBIENTES QUENTES SECOS
Adaptaes culturais ao calor seco so efetivas ao proverem microambientes mais confortveis durante boa parte do tempo. Todavia, as pessoas so expostas a macroambientes quentes e secos em algumas circunstncias, particularmente quando precisam viajar ou trabalhar ou quando a gua escassa, nestas situaes as pessoas dependem de suas repostas biolgicas.
Respostas Biolgicas Transferncia de calor. Os principais meios de lidar com o calor seco so o aumento da transferncia de calor do interior a superfcie do corpo e dela para o ambiente, bem como perder calor da superfcie por evaporao. Respostas vasculares. O fluxo sanguneo superficial aumentado pela estimulao do sistema nervoso simptico atravs da dilatao da veias superficiais, com as outras veias constritas, permitindo uma manuteno quase constante da presso sangunea. Suor. A principal adaptao humana ao estresse provocado pelo calor a habilidade de suar copiosamente. Aclimatizao. Os humanos que se aclimatizam ao calor aumentam sua taxa de suor mxima e tambm comeam a suar em temperaturas de pele e interior mais baixa.
MACROAMBIENTES QUENTES E UMIDOS
Representados em seu extremo pelas florestas pluviais tropicais, diferem significativamente dos macroambientes quentes e secos em suas caractersticas de estresse provocado pelo calor. Devido a umidade do ar e
nuvens frequentes, o ganho de calor irradiado pelo sol menor durante o dia. A perda de calor radiativo a noite reduzida tambm, resultado de uma variao diurna da temperatura muito menor. reas midas nunca ficam to quentes, nem to frias, quanto as regies ridas.
MICROAMBIENTES QUENTES E UMIDOS
O ar ambiente raramente mais quente que o interior do corpo, adaptaes culturais para modificar o ambiente so relativamente ineficientes. Assim, as pessoas so geralmente expostas ao estresse provocado pelo calor em climas midos, particularmente quando precisam se engajar em atividades musculares.
PARTE 8 ADAPTAO ALTITUDE, ATIVIDADE INTENSA E A OUTROS ESTRESSORES FSICOS
INTRODUO As populaes humanas vem se confrontando com muitos outros fatores estressores fsicos alm dos extremos de temperatura. Este captulo explorar brevemente como as pessoas lidam com alguns deles, incluindo hipxia de grandes altitudes (nveis baixos de oxignio), o estresse auto-induzido por nveis intensos de atividade e os perigos naturais, como tempestades, terremotos, tsunamis (vagalhes) e erupes vulcnicas.
A HIPXIA DE GRANDES ALTITUDES A hipxia em grandes altitudes o resultado de uma baixa presso baromtrica. A presso do ar em qualquer local baseada amplamente na quantidade de ar sobre o mesmo; o peso do ar acima empurra para baixo o ar que est embaixo, criando um aumento de densidade das molculas de ar e, assim, um aumento de presso.
Adaptaes Biolgicas Hipxia Nossas adaptaes biolgicas hipxia envolvem o aumento da eficincia e da efetividade do transporte de oxignio para dentro de nossos corpos e tecidos,
onde ele usado no metabolismo, bem como o ajuste das mudanas corpreas que so efeitos colaterais das adaptaes.
Transferncia de Oxignio para os Pulmes. Imediatamente exposio a condies de hipxia, h um aumento na taxa de ventilao, a quantidade total de ar se movendo para dentro e para fora dos pulmes em um dado perodo de tempo. Isto conseguido tanto pelo aumento da taxa de respirao quanto pela sua fora.
A transferncia de Oxignio dos Pulmes para o Sangue. A troca de oxignio com o sangue ocorre em pequenos sacos de ar (alvolos) nos pulmes. O movimento do oxignio do alvolo para os capilares sanguneos adjacentes ocorre atravs de um processo passivo de difuso. A taxa de difuso baseada em muitos fatores, incluindo a superfcie do alvolo e dos capilares, a espessura dos tecidos que separam o ar alveolar do sangue e a diferena das concentraes de oxignio entre o ar alveolar e o sangue. Um outro fator importante a capacidade de transporte de oxignio do sangue; boa parte desta capacidade se deve hemoglobina.
A circulao do sangue para todos os tecidos. Respostas circulatrias para a hipxia de grandes altitudes so primariamente baseadas em modificaes de distribuio. H um aumento transitrio na taxa cardaca e no dbito cardaco sob exposio aguda, mas que rapidamente decai para os nveis preexistentes. Enquanto o aumento do dbito cardaco acelera a taxa na qual o sangue entrega oxignio aos tecidos, h uma necessidade de demanda crescente de oxignio para o msculo cardaco, bem como um maior esforo de todo o sistema cardiovascular.
A transferncia de sangue para os tecidos. O oxignio transferido do sangue capilar para as clulas dos tecidos por meios passivos e ativos. A difuso passiva influenciada por fatores similares aos discutidos na aquisio de oxignio pelos vasos dos pulmes. A transferncia de oxignio ativamente assistida por uma protena dos tecidos, a mioglobina, a qual tem seus nveis elevados nos msculos de pessoas aclimatizadas s grandes altitudes.
Consequncias Adaptativas ao Estresse Provocados pela Hipxia
H problemas especiais para as pessoas devido s condies hipxicas de ambientes de montanhas altas, especialmente relacionadas exposio inicial. O risco e a severidade dos problemas aumentam com a altitude alcanada e com a taxa de subida. Estes problemas incluem enjo agudo, edemas pulmonares e cerebrais, enjo subagudo em crianas e enjo crnico.
Adaptaes Populacionais. As pessoas se adaptaram aos ambientes de grandes altitudes independentemente em diversas regies do planeta, incluindo as terras altas da Etipia e do Tibete, os Andes e o Himalaia. Os locais do globo onde so encontradas as reas de grande altitude. Dada a ampla distribuio destas regies, no surpreende o fato de que os estudos demonstram algumas diferenas biolgicas entre as populaes quanto forma com a qual os humanos se ajustaram.
NIVEIS DE ATIVIDADE ALTA J que a atividade auto-administrativa e no uma fora externa imposta pelo macroambiente, devemos considerar a adaptabilidade humana aos nveis de altos de atividade de uma forma um pouco diferente daquela com a qual abordamos os estressores descritos anteriormente. A grande atividade ainda geralmente devida as causas ecolgicas. Ela necessria para obter recursos e proteo contra o estressem seja correndo para caar uma presa, construindo um abrigo, plantando arroz ou alguma outra tarefa, agradvel ou no, que as pessoas precisam executar a fim de sobreviver. As pessoas podem tambm se envolver em atividades extenuantes por diverso ou esporte.
Resposta Biolgicas a Altos Nveis de Altitude A resposta individual exposio aguda. Quando as pessoas comeam a se envolver em nveis altos de atividades fsica, elas se confrontam com uma necessidade de aumentar tanto a tolerncia ao calor quanto a eficincia no uso de oxignio. Nveis altos de atividade envolvem o aumento do metabolismo, o que, por sua vez, gera calor e uma necessidade de utilizao mais rpida de oxignio. A resposta inicial ao exerccio envolve o aumento da taxa de ventilao atravs de uma taxa de respirao mais acelerada, o aumento da taxa de circulao atravs de batimentos cardacos mais rpidos e fortes e o direcionamento do sangue preferencialmente para os msculos esquelticos onde o metabolismo est ocorrendo. O sangue redirecionado a partir de
reas crticas, como a pelo. Conforme o exerccio continua e o calor do corpo aumenta, os vasos sanguneos da pele so dilatados, permitindo uma liberao mais eficiente de calor para o ambiente a partir da rea de superfcie. O suor inicia quando a temperatura da pele chega a um nvel crtico.
A forma fsica definida como a habilidade de manter condies homeostticas durante grande atividade, ou de se recuperar rapidamente aps o exerccio, envolve dois processos gerais de adaptao biolgica que j foram discutidas: a tolerncia ao calor e o uso eficiente de oxignio.
A capacidade aerbia. A maior taxa na qual podemos consumir o oxignio para o uso em nosso metabolismo conhecida como a capacidade aerbica; este um importante componente da forma fsica.
Velocidade e fora. A fora muscular a qual definida como a quantidade de fora que um msculo pode fornecer, baseada na rea transversal de um msculo, com msculos mais espessos fornecendo mais fora.
Consequncias adaptativas da capacidade diferenciada de trabalho A importante ecolgica da variabilidade da capacidade de trabalho est na capacidade diferencial de indivduos realizarem tarefas essenciais. As culturas humanas diferem quanto as suas expectativas de trabalho para as pessoas em geral e tambm nas formas como elas so divididas entre os grupos de uma sociedade.
A Unidade Econmica. A unidade econmica primaria difere em diferentes grupos culturais, desde aqueles essencialmente individuais e auto-suficientes queles que envolvem famlias extensas.
O impacto da modernizao. A capacidade de trabalho, baseada nos nveis habituais de atividade, tem sido amplamente afetada pelo processo de modernizao em muitas populaes. Este processo geralmente envolve um decrscimo dos nveis de atividade fsica e, consequentemente, um declnio da fora aerbica.
PERIGOS NATURAIS
As populaes humanas tambm precisam lidar com estressores fsicos que so de difcil previso em termos de tempo, de espao, de local e de intensidade. Estes estressores, geralmente chamados de perigos naturais, incluem as tempestades, os terremotos, os tsunamis e as erupes vulcnicas.
Erupes Vulcnicas. Alguns dos maiores estragos causados pelas erupes vulcnicas so relacionados aos terremotos e tsunamis associados a eles. Entretanto, os vulces tm causado diretamente muito sofrimento. Solos vulcnicos nas encostas dos vulces so muito ricos para propsitos agrcolas, o que leva a um grande aumento populacional nestas regies. Erupes vulcnicas podem causar estragos atravs do movimento rpido de lava, pela poeira e pelo movimento de lama. Mesmo a lava que se move vagarosamente pode destruir de vez as estruturas permanentes.
Consequncias adaptativas dos perigos naturais Como j mencionado, os perigos naturais podem ter efeitos devastadores para as populaes humanas. A tecnologia avanada de hoje permite planejamentos melhores de perigos como ciclones ou outras tempestades, erupes vulcnicas insipientes e tsunamis de longo alcance. A previso de terremotos e de tsunamis gerados localmente ainda problemtica. Ironicamente, apesar da habilidade de prever alguns destes desastres, a quantidade de destruio parece continuar crescendo. Isto pode ser devido presso populacional que obriga as pessoas a viverem em fase de maior risco. Comparados aos outros fatores estressores discutidos at agora, os desastres naturais parecem apresentar os maiores problemas para as populaes humanas. Isto se deve maior previsibilidade dos outros estressores. Assim, se for dada s pessoas a chance de se preparar para problemas como o frio, o calor e a hipxia de grandes altitudes, elas podem sobreviver. Quando um grande fator estressor ambiental no pode ser previsto, ela apresenta um desafio muito maior s capacidades adaptativas humanas. Na verdade, as pessoas consistentemente subestimam a importncia dos riscos mais bem
conhecidos. Este pode ser um meio de fatorar a importncia da previsibilidade na preparao para enfrentar circunstancias estressoras.
PARTE 9 ADAPTAO A FATORES ESTRESSANTES BITICOS: M NUTRIO E DOENAS INFECCIOSAS
Alm dos fatores estressantes fsicos em seu ambiente, os humanos tambm precisam lidar com problemas de base biolgica. Este captulo trata de problemas de cadeia alimentar humana, ou seja, como as pessoas se adaptam s circunstncias nas quais elas ou no conseguem obter alimentos de seus recursos, ou quando organismos (patgenos) tentam us-las como fonte de comida. Estes dois problemas esto, na verdade, inter-relacionados.
M-NUTRIO: O PREO DA FALHA
A m nutrio pode ser definida como qualquer tipo de nutrio desbalanceada, incluindo a ingesto de muita ou pouca comida. O foco ser dado, principalmente, subnutrio, na qual a quantidade tanto de alimentos em geral quanto de nutrientes especficos requeridos por uma dieta so inadequados.
M-NUTRIO PROTICO-CALRICA
Uma das principais formas de m-nutrio causada pela deficincia de contedo energtico da comida, medido em kcal (Kilocalorias), e/ou deficincias proteicas.
Deficincia Proteicas As protenas so necessrias para o crescimento, reproduo e sobrevivncia diria; nenhum organismo pode sobreviver muito tempo sem a habilidade de produzir as protenas necessrias. As protenas precisam ter uma ordem precisa de seus aminocidos para manter sua funo. Se um tipo de aminocido falta, os humanos e outros
organismos no conseguem de maneira correta, produzir suas protenas necessrias; isso forma o menor denominador comum da sntese proteicas a partir de seus aminocidos constituintes.
A Qualidade Proteicas dos Alimentos. Ingerir protenas no o suficiente para evitar a m-nutrio: devem ser ingeridas protenas que contenham a taxa apropriada de aminocidos essenciais para as necessidades humanas. A qualidade proteicas se refere ao grau no qual um dado tipo de protena da dieta combina com a taxa de aminocidos essenciais requerida pelos humanos.
O Cenrio da Deficincia Proteicas. A deficincia proteica uma das principais formas de m-nutrio. Ela particularmente comum em pases em desenvolvimento, especialmente em regies tropicais onde os alimentos comuns tendem a ser ricos em carboidratos e pobres em protenas.
Efeitos Biolgicos da M-Nutrio Proteicas. A maioria dos casos de mnutrio proteicas ocorre quando a ingesto de protenas , de certa forma, menor que os valores requeridos a longo prazo. A m-nutrio severa durante a infncia leva a uma doena de deficincia chamada de kwashiorkor, uma palavra derivada da linguagem Ga da frica e que significa doena que ocorre quando se substitudo da mama por outra criana. Os sintomas da doena incluem a atrofia muscular, a falha do crescimento, as irritaes de pele, os edemas e, em alguns casos, a perda dos pigmentos da pele e do cabelo.
Subnutrio Total: Deficincia de Protenas e de Calorias
Reservas Energticas Biolgicas Humanas. Como uma fonte de energia de curto prazo, as pessoas dependem do glicognio, o amido estocado no fgado e nos msculos. Quando os nveis sanguneos se encontram altos, um hormnio chamado insulina secretado e estimula a glicognese (a sntese de glicognio a partir da glicose, um acar simples). Quando os nveis sanguneos esto baixos, como ocorre na subnutrio, o hormnio glucagon liberado e estimula a gliclise (a quebra do glicognio em glicose). O glicognio utilizado na manuteno de nveis de acar mais ou menos estveis no sangue, independentemente da ingesto de acares na
alimentao. O acar a principal fonte de energia a curto prazo no corpo humano, garantindo um fornecimento constante de energia.
O Ciclo de Cori. Enquanto a gordura a principal das fontes de energia, h problemas quanto ao seu uso, j que os msculos esquelticos e crebro humanos no se utilizam diretamente da gordura como fonte de energia, dependendo, na verdade, da glicose. O ciclo de Cori, um processo adaptativo de conservao de glicose, envolve uma mudana para o metabolismo anaerbico no tecido muscular, no qual a glicose quebrada em lactose em vez de molculas pequenas de dixido de carbono e gua. A quebra de glicose libera energia no msculo.
Efeitos da M-Nutrio Protico-Calrica. Em adultos, a m-nutrio protico-calrica tem muitos efeitos negativos. O efeito mais bvio a grande perda de peso corpreo. Enquanto a maior parte desta perda oriunda da queima de gordura, h uma perda considervel de protenas, particularmente das que se encontram na massa de msculo esqueltico. A capacidade fsica de trabalho tambm decai, porm, somente aps um longo perodo de deficincia protico-calrica. Os efeitos da m-nutrio protico-calrica so muito maiores sobre as crianas, podendo levar a uma doena chamada marasmo.
DEFICINCIAS DE VITAMINAS
Alm dos principais componentes nutricionais dos alimentos carboidratos, gordura e protenas -, a comida contm muitos micronutrientes (componentes necessrios apenas em pequenas quantidades) indispensveis para a sade e a funo humanas. Dentre os micronutrientes esto minerais, como o iodo e o zinco, e fatores alimentares acessrios, as vitaminas.
Deficincia da Vitamina A A vitamina A (retinol) encontrada em vegetais verdes e amarelos, no leite, na manteiga e no queijo; ela tambm pode ser sintetizada no corpo a partir do beta-caroteno, o qual encontrado em muitos alimentos vegetais. armazenada no fgado, mas o mau funcionamento do rgo pode acabar com as reservas, contribuindo para a deficincia.
Deficincia de Tiamina A tiamina, tambm conhecida como vitamina B, funciona no metabolismo dos carboidratos e, dessa forma, a sua deficincia particularmente problemtica quando as dietas contm pouca vitamina e so ricas em carboidratos. Embora a tiamina seja encontrada em quase todos os tipos de alimentos, incluindo a carne de porco, o fgado, os gros e legumes, as deficincias so observadas em populaes que dependem do arroz como base de sua dieta.
Deficincia de Niacina A niacina, tambm conhecida como vitamina B, encontrada em baixas quantidades no milho, e as doenas relacionadas sua deficincia sempre esto ligadas a populaes com dietas baseadas no milho. Entretanto, ela encontrada no fgado, na carne, em gros e nos legumes. O aminocido triptofano pode ser convertido em niacina pelo corpo, mas ele tambm relativamente escasso no milho.
Deficincia de Vitamina C O cido ascrbico funciona como um caminho bioqumico no qual o aminocido prolina convertido em colgeno, O colgeno, por sua vez, funciona como um qumico que se liga s clulas nos tecidos do corpo, notavelmente cartilagem, aos ossos e aos dentes. Assim, a deficincia de vitamina C est associada ao enfraquecimento da capacidade de cicatrizao e ao rompimento de feridas antigas. O resultado a hemorragia (sangramento), geralmente interna, que leva a ferimentos graves de juntas, entre outros sintomas.
Deficincia de Vitamina D A vitamina D pode ser obtida a partir da dieta (por exemplo, ovos), mas tambm pode ser sintetizada dos esterides encontrados no alimento. A sntese, que envolve a radiao ultravioleta do sol na reao, ocorre nas camadas superficiais da pele (Cavalli-Sforza,1981). A pele clara favorece a reao, enquanto roupas bloqueiam a luz, inibindo-a. Em regies de radiao solar intensa, a reao produz vitamina D suficiente mesmo quando a cor da pele bem escura. Na verdade, alguns antroplogos sugeriram que a pele clara evoluiu, em parte, como uma adaptao sntese de quantidades adequadas de vitamina D em povos que habitavam as latitudes norte com doses pequenas de radiao solar, enquanto a pele mais escura foi e vista
como uma adaptao para filtrar a radiao ultravioleta e evitar a hipervitaminose D (muita vitamina D) (Loomis, 1967). A vitamina D necessria para a absoro do clcio no intestino e para seu uso subsequente na formao de ossos e dentes. A doena da deficincia na infncia associada vitamina D conhecida como raquitismo. Ela caracterizada pela curvatura dos ossos, pelo atraso da erupo dos dentes (em crianas) e pelo desenvolvimento precrio do esmalte dentrio (Adair, 1987). Osteomalcia o termo para a mesma doena quando manifestada em adultos.
DEFICINCIAS MINERAIS
Deficincia de Ferro Embora o ferro ocorra naturalmente em ovos, carnes, legumes, gros e vegetais verdes, a deficincia de ferro foi chamada de o mais prevalecente problema nutricional dos dias atuais (Scrimshaw, 1991, p.46). Estimativas para os pases em desenvolvimento vo desde 67% das crianas e 33% das mulheres em fase reprodutiva com algum tipo de deficincia de ferro. Porm, a deficincia mais encontrada em pases subdesenvolvidos, particularmente entre as mulheres. A maioria do ferro no corpo utilizada na protena hemoglobina, a qual carrega oxignio e gs carbnico no sangue. Uma deficincia severa de ferro pode levar anemia, conforme a produo de hemcias se torna defectiva devido falta de hemoglobina. A anemia provoca muitos efeitos danosos s pessoas, desde a diminuio da capacidade de trabalho at o empobrecimento da performance intelectual.
Deficincia de Iodo O iodo um elemento qumico importante dos hormnios da tireoide e a deficincia deste mineral pode levar a uma reduo do trabalho da glndula. A glndula tireoide geralmente incha na tentativa de aumentar a produo dos hormnios. Esta situao chamada de bcio. Em casos extremos, o bcio desfigura o indivduo e pode at mesmo causar presso sobre a traqueia caso fique muito grande.
Deficincia de Clcio
As sndromes de deficincia de clcio esto geralmente relacionadas aos problemas causados pela deficincia da vitamina D, j que ela importante na utilizao do clcio pelo corpo. O mineral importante para a formao dos ossos e dos dentes, durante o desenvolvimento e para sua manuteno. O clcio tambm responsvel pelo bom funcionamento do sistema nervoso. Este mineral encontra-se em altas concentraes em itens da dieta como derivados do leite e vegetais verdes como brcolis. A deficincia de clcio pode levar ao crescimento vagaroso dos ossos e osteoporose. A osteoporose torna os ossos muito susceptveis fratura.
ADAPTAO E M-NUTRIO
Muitas das respostas humanas m-nutrio podem no ser adaptativas. Respostas comportamentais podem no ser apropriadas e podem, at mesmo, aumentar a severidade da m-nutrio. Mudanas biolgicas associadas a ela podem simplesmente ser respostas fisiolgicas ausncia ou falta de alguns nutrientes necessrios. Entretanto, parece haver alguns mecanismos preparados para algumas das deficincias nutricionais que servem para atrasar as consequncias mais devastadoras.
Alimentos versus Nutrientes Em todas as populaes humanas, uma diferena pode ser encontrada entre alimento e nutrientes. Alimento um conceito culturalmente definido, incluindo todas as substncias consideradas comestveis. Nutrientes so compostos qumicos necessrios ao corpo e incluem carboidratos, lipdios, protenas, vitaminas e minerais.
Adaptao Biolgica M-Nutrio Problemas nutricionais no podem ser vistos isoladamente. Eles afetam a habilidade de uma populao para lidar com muitos outros estressores, bem como sua habilidade para obter recursos/ podem, por exemplo, diminuir a capacidade de trabalho. A m-nutrio, portanto, afeta todas as outras reas da ecologia da populao. Em nenhum outro domnio este impacto mais fortemente sentido do que na habilidade das pessoas de resistirem a doenas infecciosas.
DOENAS INFECCIOSAS COMO ESTRESSORES BITICOS
Uma Nota sobre Taxas de Doenas Taxas de doenas podem significar muitas coisas para um epidemiologista. Duas das principais formas de determinar o quo comum uma doena em uma populao se do a partir da medio do nmero de fatalidades, devidas doenas, dividido pelo tamanho da populao, o que chamado de taxa de mortalidade da doena em relao ao tamanho da populao, o que chamado de taxa de morbidez. H duas formas comuns de medir a taxa de morbidez: atravs da prevalncia de uma doena ou atravs de sua incidncia. A taxa de prevalncia de uma doena o nmero de casos de uma doena em um dado momento do tempo. A taxa de incidncia de uma doena definida como o nmero de novos casos que aparecem durante um dado perodo de tempo.
Relaes Hospedeiro/Parasita Os parasitas e seus hospedeiros tendem a coevoluir em uma coexistncia pacfica em muitos casos. Ou seja, parece haver uma seleo evolutiva tanto em favor dos hospedeiros que no morrem ou que no ficam muito doentes, fazendo com que o sucesso reprodutivo diminua, quanto em favor dos parasitas que no matam seu hospedeiro (o ganso de ouro). H alguma evidncia de que esta coevoluo de hospedeiros e parasitas resulta em menos virulncia. A variabilidade gentica da populao de hospedeiros, particularmente dos genes relacionados ao sistema imune, pode ser muito importante ao permitir que a populao de hospedeiros rapidamente se adapte ao novo agente da doena. A seleo natural contra a virulncia de parasitas ocorrer somente se houver uma vantagem aos parasitas relacionadas manuteno da vida do hospedeiro. Para parasitas que podem facilmente mudar de hospedeiro, talvez no haja nenhuma modificao acerca de sua virulncia ao longo do tempo.
DOENAS TRANSMITIDAS POR VETORES
Vetores so os veculos pelos quais os parasitas so transferidos de um hospedeiro infectado para um susceptvel. Eles geralmente so artrpodes, um
grande grupo biolgico que inclui insetos, aranhas e camares, dentre outros. Vetores podem ser considerados em dois grupos: agentes biolgicos ou mecnicos. Como um agente mecnico, o vetor simplesmente transfere os parasitas atravs de contato externo (por exemplo, moscas carregam germes em suas patas quando pousam em excrementos, ento, pousam na comida, onde os germes so depositados). Como um agente biolgico, o prprio vetor se torna infectado (com ou sem sintomas). Em alguns casos, o parasita obrigado a passar parte de seu ciclo de vida em um vetor especfico. Em geral, doenas transmitidas por vetores podem ser facilmente transmitidas mesmo quando o hospedeiro j est doente e, dessa maneira, estas doenas tendem a se manter virulentas mesmo aps longa associao com humanos. Doenas transmitidas por vetores tm, sem dvida, afetado a humanidade ao longo de toda a evoluo dos homindeos, e algumas delas tiveram um impacto muito grande sobre a ecologia e a Biologia humanas. Dois exemplos de doenas carregadas por vetores sero discutidos brevemente aqui: a malria e a oncocercose. Estes exemplos foram escolhidos para dar uma ideia de como as doenas transmitidas por vetores podem afetar a ecologia humana.
Malria A malria tem sido, provavelmente, o maior assassino de humanos dos ltimos milnios. H cerca de 300 milhes a 500 milhes de casos de malria em qualquer momento do tempo e, aproximadamente 2,7 milhes de pessoas morrem desta doena a cada ano (Nussenzweig e Long, 1994). Ela causada por diversas espcies de protozorios do gnero Plasmodium, particularmente o P. vivax, o P. malarie e o P. falciparum.
Protetores Biolgicos Contra a Malria. Os humanos desenvolveram algumas adaptaes biolgicas especiais contra a malria que envolvem variantes genticas da hemoglobina. O caso mais bem conhecido de resistncia gentica malria o caractere falsiforme da clula. A clula falsiforme representa uma pequena mutao do gene que determina a molcula da hemoglobina. Esta mutao faz com que uma molcula de hemoglobina tenda a aderir a outra, particularmente quando a molcula se encontra na forma desoxigenada (ou seja, aps o descarregamento de oxignio da hemoglobina para os tecidos) (Edelstein, 1986). O complexo da molcula de hemoglobina pode formar grandes estruturas fibrosas dentro das hemcias, com estas estruturas algumas vezes distendendo as clulas em uma forma falsiforme caracterstica.
Oncocercose A oncocercose uma doena filarial (um tipo de verme parasita) transmitida pela mosca preta (vetor). O parasita desta doena o Onchocerca volvulus e o vetor, Simulium damnosum. Embora esta doena no seja geralmente fatal, os parasitas se movem pela corrente sangunea at o olho, onde frequentemente causam cegueira. J que cada fmea capaz de botar cerca de 2.000 microfilarias ao dia e pode residir no corpo humano por dcadas (Desowitz, 1981), as vtimas podem sofrer uma sobrecarga parastica. As moscas pretas tipicamente se reproduzem em reas de gua altamente oxigenada, sendo as rochas constantemente banhadas seu stio de preferncia. Devido a essa caracterstica, a oncocercose , muitas vezes, chamada de cegueira do rio.
DOENAS TRANSMITIDAS ATRAVS DO CONTATO DIRETO
Doenas de contato direto so geralmente transmitidas pela rota dos perdigotos, quando tossidas, espirros ou contato simples com a respirao das vtimas podem servir para passar os agentes causadores da doena. Muitas das doenas transmitidas por esta rota so especficas dos humanos, embora este no seja sempre o caso. Muitas doenas de contato direto apareceram em associao ao surgimento da agricultura intensiva e resultante alta densidade humana em regies diferentes do planeta (McKeown, 1988). Estas doenas so transmitidas para outras populaes humanas quando o contato feito e elas podem se mostrar devastadoras para novas populaes hospedeiras.
Epidemias em Populaes Virgens Quando uma doena introduzida pela primeira vez, nenhum indivduo possui imunidade prvia e, assim, todos se encontram susceptveis infeco. Estas epidemias, chamadas de epidemias em solo virgem, podem ser calamitosas, com estimativas de at 90% de mortalidade em populaes expostas a diversas novas doenas aps o contato com povos de culturas cosmopolitas e urbanas, onde muitas doenas de contato direto so endmicas. Estimativas altas para a mortalidade depois do contato com determinados tipos de doenas (sendo a varola e o sarampo citados como os principais assassinos).
DOENAS TRANSMITIDAS PELA VIA SANITRIA
Doenas transmitidas por rotas sanitrias tm sido problemas de pessoas pobres, j que elas se dispersam atravs da ingesto de alimentos e de gua contaminados. Um dos principais meios de transmisso de tais doenas a rota fecal-oral, na qual os parasitas so passados de uma pessoa a outra atravs da contaminao da gua e dos alimentos. Tal contaminao pode ocorrer atravs de vetores mecnicos, como moscas que pousam em alimentos, ou atravs da mistura de excrementos com a gua que ser bebida, como ocorre quando uma populao depende de uma nica fonte de gua para diversos propsitos.
DOENAS TRANSMITIDAS ATRAVS DE CONTATO NTIMO
Doenas transmitidas atravs do contato ntimo consistem principalmente de doenas sexualmente transmissveis, mas tambm de doenas transmitidas atravs de fluidos corpreos em geral, sejam eles sangue, saliva ou smen, ou atravs do contato direto com a pele. Assim, estas doenas so muito influenciadas pelas morais sexuais, bem como por outros tipos de comportamento relacionados com a troca de fluidos corpreos, tais como o compartilhamento de agulhas no uso de drogas ilcitas, a esterilizao imprpria de equipamentos mdicos ou at mesmo o transplante de rgos do corpo.
Treponematoses Treponematoses consistem de diversas condies patolgicas causadas por espiroquetos (um tipo de bactria) do gnero Treponema. Espcies diferentes de Treponema so acusadas de causar diferentes condies, mas h dvidas se eles so realmente espcies diferentes ou se, na verdade, as doenas so causadas por bactrias variadas (Wood, 1979). Em geral, as treponematoses so doenas de pele, embora possam envolver outros tecidos. Quatro formas principais de treponematoses so a sfilis venrea, a bolba, a pinta e a sfilis no-venrea (ou endmica), algumas das quais so mortais.
AIDS
O agente da AIDS conhecido como vrus da imunodeficincia adquirida (HIV). O HIV um retrovrus, o que significa que sua informao gentica carregada em uma molcula de RNA, em vez de DNA. A informao do RNA copiada na molcula de DNA atravs da utilizao de uma enzima especial chamada de transcriptase reversa, e o DNA resultante incorpora-se ao DNA do hospedeiro. Este DNA, em algum momento, se replica em molculas de RNA que se tornam parte de novos vrus ou decodificam protenas necessrias replicao do HIV (Stine, 1993).
Os Efeitos da AIDS. O HIV infecta clulas do corpo humano que contm certos tipos de protenas, chamadas de CD4 ou de receptor de antgenos T4, em sua superfcie. As principais clulas que possuem este tipo de receptor de membrana so as T4, ou ajudantes T. Estas clulas so um elemento importante do sistema imunolgico, ajudando a reconhecer vrus, fungos e outros parasitas. Infeces longas levam destruio das clulas T4 e a vtima se torna susceptvel a muitos patgenos que estas clulas ajudam a combater. Assim, o indivduo sofre de uma deficincia de seu sistema imunolgico, sucumbindo a uma ou muitas infeces oportunistas.
CONCLUSO
Os dois estressores discutidos foram, e so, grandes foras seletivas sobre a populaes humanas. Os mdicos, felizes, proclamaram a chegada do final das doenas infecciosas, iludidos pelo sucesso dos antibiticos e das vacinas. A evoluo da resistncia aos antibiticos em patgenos humanos e as flutuaes genticas que derrubam nossas tentativas de criar vacinas efetivas causaram a emergncia de velhas doenas no mundo desenvolvido. Os efeitos da m-nutrio e de doenas parasticas que exacerbam os efeitos da subnutrio podem ser especialmente prejudiciais para as crianas. H evidncias abundantes de que a m-nutrio crnica em crianas pode ter efeitos srios sobre o desenvolvimento mental, com as habilidades intelectuais inibidas por toda a vida (Brown e Pollitt, 1996). possvel que tal debilitao mental possa causar dificuldade quanto obteno de recursos quando o indivduo for um adulto, o que cria o cenrio para a m-nutrio da prxima gerao. A fome e a peste continuam sendo dois cavaleiros do apocalipse, cavalgando muito perto de seus outros colegas cavaleiros, a guerra e a morte. Os humanos se adaptaram a estes dois estressores montados tanto comportamental quanto biologicamente, mas se confrontam com adversrios vivos que tambm se
adaptaram aos humanos. uma corrida evolutiva na qual a adaptao pode ser vista como um processo, em vez de um estado perfeito.
PARTE 10 MODERNIZAO, ESTRESSE E DOENAS CRNICAS
A adaptao humana foi apresentada at aqui envolvendo a criao de microambientes mais agradveis e dependendo de respostas biolgicas somente quando o estresse macroambiental no totalmente resolvido atravs dos meios comportamentais. Para o ambiente urbano moderno, com o qual a maioria de ns lida diariamente, esta aproximao adaptabilidade humana inadequada. Na verdade, muitos dos estressores com os quais temos de lidar so nossas prprias criaes. O microambiente urbano possui muitos estressores, desde a poluio, o crime, os engarrafamentos at as provas pelas quais os estudantes tm de passar, que esto muito pouco relacionados ao macroambiente no qual nosso ambiente urbano se localiza. Enquanto estes estressores podem diferir daqueles que foram considerados nos captulos anteriores, eles so, todavia, estressores, e a permanncia da espcie humana a longo prazo depende de nossa habilidade de lidar com estes desafios ambientais auto-induzidos.
PRINCIPAIS DOENAS CRNICAS EM POPULAES MODERNIZADAS
A prevalncia de grandes doenas crnicas aumenta em populaes que mudam de um modo de vida tradicional para um modernizado.
Cncer Cncer o termo utilizado para uma condio na qual h um crescimento incontrolado de clulas do corpo. H muitas formas de cncer, com muitas causas diferentes. Em geral, entretanto, as clulas se tornam cancerosas devido a mutaes que alteram os genes que controlam o crescimento e a reproduo celulares. Essas mutaes podem ligar alguns genes (chamados de oncogenes) ou desligar outros (genes supressores do cncer).
Hipertenso
Causas da Hipertenso. A hipertenso mais um sintoma do que uma doena, definida como a persistncia de alta presso arterial. A hipertenso pode ser causada por doenas, tais como os problemas de deficincia dos rins. Entretanto, para a vasta maioria dos casos de hipertenso, geralmente classificados de primrios, ou essenciais, nenhuma causa especfica conhecida. Em geral, qualquer fator que aumente o volume de sangue ou que diminua o volume interno dos vasos sanguneos, aumentar a presso sangunea. Isso representa um nmero enorme de fatores possveis, desde a ingesto de fluidos e eletrlitos at a transpirao, a temperatura ambiental, a atividade fsica e a ativao do sistema nervoso simptico, dentre muitos outros. A hipertenso est relacionada ao seguinte: idade, sexo (homens apresentam um risco maior), situao socioeconmica (pessoas mais pobres apresentam mais risco), hereditariedade, dieta, peso e quantidade de gordura corprea, estresse e mudanas sociais abruptas.
Doenas do Corao As doenas do corao, em todas as suas formas, so a principal causa de mortes nos Estados Unidos e no mundo desenvolvido como um todo. A principal forma de doena do corao aquela das artrias coronrias, os vasos que fornecem sangue ao msculo cardaco. A doena da artria coronria envolve leses aterosclerticas dos vasos. A aterosclerose se refere existncia de depsitos duros de lipdios, chamados de placas, na parede interior das artrias. Estas placas podem aumentar e causar cogulos em sua superfcie, levando a uma ocluso parcial ou total do vaso sanguneo. Quando a ocluso ocorre nas artrias coronrias, o fornecimento de sangue para uma poro do corao pode ser bloqueado, levando a um ataque do corao.
Diabete Melito A diabete melito uma desordem que envolve a incapacidade do corpo de metabolizar carboidratos de uma maneira normal. Esta incapacidade devida falta de habilidade de produzir e utilizar o hormnio insulina. A insulina produzida por clulas beta no pncreas, sendo a produo estimulada pelo aumento dos nveis de acar no sangue. A insulina liberada na corrente sangunea, por onde transportada para todo o corpo, ligando-se a receptores qumicos localizados na poro externa das membranas das clulas em todos os tecidos. A entrada de acar nas clulas leva ao declnio dos nveis do acar no sangue e ao trmino do estmulo para a produo de insulina. Finalmente, boa parte da insulina circulante retirada do sangue atravs do fgado.
A Modernizao e as Doenas Crnicas As evidncias que acabamos de apresentar mostram somente que as taxas de doenas crnicas so aumentadas em populaes que vivem em ambientes modernizados, mas este fato sozinho no identifica os fatores verdadeiros envolvidos nas causas das doenas. Agora nos voltaremos para alguns dos fatores em condies modernizadas que, possivelmente, apresentam um papel causal sobre o desenvolvimento de doenas crnicas. Especificamente, consideraremos brevemente trs fatores: a poluio, o estresse generalizado e a adiposidade.
POLUIO
Uma definio preliminar de poluio um recurso que est fora de lugar no ambiente, em grande ou pequena quantidade a partir da perspectiva de comunidades ecolgicas especficas. Uma das principais conexes entre populao e doena parece estar na exposio a grandes concentraes de certos produtos qumicos elaborados em processos industriais e altas taxas de certas formas de cncer. Trs das principais formas de poluio so: ar, gua e lixo slido.
ESTRESSE GENERALIZADO E DOENAS CRNICAS
Uma Definio do Estresse Generalizado Segundo esta viso, o estresse generalizado uma varivel cognitiva, quando as percepes do indivduo sobre um dado estmulo determinam se a reao biolgica ao estresse ocorrer (Pearson et al., 1993). Alguns acreditam que o estmulo deva ser visto como uma ameaa de algum tipo difcil de lidar (Lazarus, 1966, 1993) e, assim, cria-se uma demanda por adaptaes. Uma ameaa neste contexto qualquer coisa que possa atrapalhar a homeostase, o funcionamento normal do corpo. O que percebido pelo corpo. O que percebido pelo corpo como estressor para um indivduo no necessariamente o para outro, j que as pessoas diferem quanto ao que visto como ameaa e ao que visto como fcil ou difcil de lidar.
Estresse Generalizado e o Ambiente Urbano
Sempre se assumiu que a vida urbana em ambientes modernizados est associada a grandes quantidades de estresse generalizado. Psiclogos e fisilogos modelaram os estressores urbanos em pesquisas de laboratrio (conforme Frankenhaeuser, 1973; Glas e Singer, 1972) e demonstraram que os problemas relacionados ao ambiente modernizado, desde o barulho at burocracias, levam a respostas biolgicas ao estresse sofrido pelos indivduos. Pesquisas que se concentram diretamente em pessoas e suas vidas dirias normais em condies modernizadas so menos comuns, em parte devido dificuldade de criar projetos que tenham significado cientfico e que ofeream a possibilidade de controle sobre a mirade de fatores que podem levar s respostas ao estresse.
ADIPOSIDADE E DOENAS CRNICAS
Outro risco sade associado aos ambientes modernizados a tendncia de aumento da adiposidade (quantidade de gordura corprea). Na verdade, em muitas populaes modernizadas, uma grande porcentagem de pessoas obesa (muito gorda), embora isto no seja verdade para todas as populaes (Baker, 1984).
CONCLUSO
A evidncia nos persuade de que a modernizao leva ao aumento srio das taxas de doenas crnicas, sendo os aumentos relacionados poluio, ao estresse generalizado e adiposidade. Este aumento das doenas pode refletir a falha das adaptaes humanas devido rapidez das mudanas ambientais representada pela modernizao. Talvez realmente habitemos corpos adaptados vida na idade da pedra, como caadores-coletores ou como agricultores primitivos. Certamente, dado o longo tempo de gerao dos humanos, os poucos milnios de vida em centros urbanos ainda no forneceram tempo suficiente para grandes mudanas genticas. Quando se considera que muitas mudanas da modernizao poder ser medidas em perodos de anos em vez de sculos e milnios, razoavelmente possvel que algumas adaptaes genticas tenham se tornado obsoletas. A reao do lute ou fuja nos ajuda a fugir de um leo, mas ela nos aju da a lidar com o trfego urbano? O aumento do esforo cardiovascular no possui funo quando entramos em um automvel e pode levar ao aumento do risco de desenvolvimento de doenas.
A rapidez da modernizao pode tambm superar a habilidade de uma populao de modificar suas adaptaes comportamentais. Para os indivduos, os comportamentos e valores aprendidos na infncia podem no ter mais significado em uma sociedade modificada quando eles atingem a maturidade. Pode-se imaginar tambm que as adaptaes biolgicas de desenvolvimento podem se tornar perigosas para os indivduos quando seus ambientes se modificam rapidamente. A mudana rpida em qualquer ambiente leva a dificuldades srias para a comunidade biolgica residente. No surpresa que os humanos necessitem lidar com as mudanas ambientais atravs da mudana de seus prprios meios bsicos de adaptao. Com a modernizao, a mudana rpida parece ser a nica constante e, portanto, as habilidades adaptativas humanas podem enfrentar desafios srios no futuro.
PARTE 11 RECURSOS NATURAIS E ECOLOGIA DE COMUNIDADES
Todos os organismos precisam de substncias, chamadas de recursos naturais, obtidas do meio ambiente, a fim de sobreviver e reproduzir-se. Esses recursos naturais incluem alimento, gua e, frequentemente, algum tipo de proteo contra agentes de estresse do meio ambiente, sejam eles fsicos, tais como temperaturas extremas, ou biolgicos, predadores ou parasitas. Pode-se incluir, tambm, entre recursos naturais, os parceiros sexuais, os materiais ou o territrio necessrio para fins de reproduo, abrangendo desde a seduo de um (a) parceiro (a) at a proteo da prole.
EFICINCIA E RISCO NO USO DE RECURSOS
Recursos Naturais Renovveis e No-Renovveis Ao lidar com populaes humanas, precisamos distinguir entre recursos naturais renovveis e no-renovveis. Como est implcito nos prprios termos, um recurso renovvel pode ser reposto (por exemplo, o oxignio), ao passo que o no-renovvel no pode (por exemplo, combustvel fssil). Essa distino um pouco artificial, uma vez que todos os recursos so renovveis, se lhes for concedido tempo suficiente. Combustveis fsseis, em geral vistos como no-renovveis, na verdade so renovveis, caso se queiram esperar os milhes de anos necessrios para criar petrleo novo.
O Papel da Tecnologia e o Uso dos Recursos Naturais
A tecnologia um ponto central quando nos confrontamos com o uso e o abuso dos recursos naturais. A tecnologia atual pode permitir o uso de recursos naturais anteriormente impossveis de serem obtidos. Na verdade, a maior diferena entre os povos tradicionais e os grupos modernizados, no que se refere ao abuso dos recursos naturais, parece ser a escala em que se processam as atividades humanas; populaes modernizadas so de tamanho muito maior e dispem de tecnologia mais complexa, e por essa razo so capazes de causar maiores danos que as populaes tradicionais, isso tanto no passado como no presente.
PRINCIPAIS TIPOS DE PADRES DE SUBSISTNCIA HUMANA
Comida um recurso natural essencial para todas as populaes humanas e tem merecido ateno especial por parte dos estudiosos da ecologia humana, em particular dos eclogos culturais. Os sistemas de subsistncia podem ser divididos em quatro tipos principais: o forrageio, o pastoral ismo, a horticultura e a agricultura intensiva. No interior de cada uma dessas categorias maiores encontram-se diversas atividades de subsistncia, havendo, porm, relacionados ao uso humano dos recursos naturais, alguns pontos que so comuns aos quatro tipos principais.
Forrageio O forrageio refere-se a atividade de subsistncia que dependem de recursos alimentares no cultivados para formar a parte principal do suprimento alimentar. Usualmente, isto implica uma combinao de coleta de plantas no cultivadas com atividades de pesca ou de caa a animais selvagens. At cerca de 12 mil AP, todos os humanos eram caadores-coletores; agora, porm, s uma minscula proporo de humanos pode ainda ser classificada nesse tipo de subsistncia.
Pastoralismo O termo pastoralismo refere-se dependncia do pastoreio de animais domesticados como a principal fonte de recurso alimentar de uma sociedade. Assim como os caadores-coletores, os pastoralistas contemporneos apresentam uma larga diversidade cultural e so encontrados em muitos ambientes diferentes. A exemplo dos caadores-coletores, os pastoralistas
modernos so mais encontrados em ambientes um tanto marginais, onde as colheitas de alimentos no so tidas como certas ou no podem ser produzidas em quantidade suficiente para dar conta das necessidades de recursos naturais do grupo. Esses ambientes incluem pastagens ridas, montanhas altas e algumas regies subrticas.
Horticultura A agricultura de subsistncia com o uso de ferramentas simples e sem o emprego de tcnicas complexas de irrigao e fertilizao denominada horticultura. Embora os eclogos culturais usualmente se refiram horticultura como um dos principais tipos de subsistncia, diferente da agricultura intensiva, em que comumente se faz uso de ferramentas sofisticadas, irrigao e fertilizao, a diferena entre esses dois tipos de subsistncia baseia-se mais em grau do que em uma clara distino qualitativa. H diferenas de ordem geral, no entanto. As prticas horticultoras ocorrem em reas menores de terra do que as da agricultura intensiva em roas, no em campos. Um lote de terra usado para horticultura fica esgotado por longos perodos aps ter sido usado, ao passo que as tcnicas da agricultura intensiva permitem que os perodos de esgotamento e renovao sejam mais curtos ou mesmo inexistentes. Os horticultores em geral trabalham menos arduamente do que o fazem os agricultores intensivos, a menos que estes ltimos possam usar combustveis fsseis para fazer parte do trabalho para eles, mas os horticultores no conseguem produzir a mesma quantidade de comida numa dada rea de terra. Isto no significa que os horticultores sejam preguiosos, ignorantes ou ineficientes. Pelo contrrio, os horticultores tm mantido atividades de agricultura em regies onde as tcnicas intensivas se mostraram desastrosas.
Agricultura Intensiva Agricultura intensiva representa um impacto humano muito maior sobre o meio ambiente natural. Tecnologia e trabalho so usados para criar sistemas ecolgicos artificiais, com plantas e animais domesticados chegando a suplantar a comunidade nativa, e o uso humano de fertilizantes e de sistemas de irrigao suplantando os ciclos nutrientes naturais. Na verdade, em alguns lugares, toda a paisagem foi redesenhada pela populao, com a criao de terraos, elevaes de nveis, construo de diques e outras mudanas significativas para a geografia natural de uma regio.
ECOLOGIA DE COMUNIDADES
Em ecossistemas, as necessidades de recursos naturais formam a base para importantes conexes entre as populaes. Especificamente, a necessidade de organismos em cadeias alimentares num ecossistema dada em uma forma fundamental: comem ou so comidos uns pelos outros. Esses tipos de interaes tambm ligam organismos em comunidades biolgicas. A estrutura e a forma das comunidades biolgicas so estudadas no campo chamado ecologia de comunidades.
A Natureza de uma Comunidade Ecolgica Uma comunidade ecolgica pode ser vista como um conjunto de populaes de espcies que possuem um potencial para interao. Mais precisamente, uma comunidade pode ser definida como uma reunio interativa de espcies ocorrendo no interior de uma rea geogrfica; um conjunto de espcies cujas funes e dinmicas ecolgicas so de alguma maneira interdependentes (Putman, 1994). Essas interaes incluem as de competio aberta e as relaes de alimentao, bem como manifestaes mais sutis, tais como a dependncia de plantas por animais para polinizao e para disperso das sementes, ou de animais por plantas para fornecimento de hbitat (abrigo). Como seria ento de se esperar, as comunidades ecolgicas existem em todos os tamanhos, formas e graus de interao de suas populaes constituintes.
A Forma e a Estrutura das Comunidades Ecolgicas A forma e a estrutura, ou fisionomia, de uma comunidade pode ser descrita em um sem-nmero de maneiras, cada uma avaliando aspectos um tanto diferentes, cada uma tendo certas vantagens e limitaes, e, como se poderia esperar, cada uma tendo seus defensores nos crculos ecolgicos.
Formas de Crescimento. Talvez a mais recente caracterizao tcnica da estrutura de uma comunidade, particularmente quando aplicada a plantas, emprega formas de crescimento comumente reconhecidas, tais como rvores perenes e decduas, arbustos herbceos, ervas e assim por diante.
Formas de Vida. Uma variao na recebendo a denominao de forma sculo XX pelo botnico dinamarqus entre a superfcie do solo e o (meristemtico) da planta.
forma de crescimento de uma planta, de vida, foi desenvolvida no incio do Raunkiaer (1934), com base na relao tecido embrionrio ou regenerativo
Estratificao. Tanto a forma de vida como a forma de crescimento das plantas enfatizam a altura. At numa olhadela casual, dada a uma comunidade de plantas, observamos diferenas nas alturas, ou estratificao vertical, de seus componentes. rvores so em geral mais altas do que arbustos, que so usualmente mais altos que as ervas, e estas ltimas so mais altas do que os musgos e os lquenes. As florestas tropicais caracterizam-se tipicamente por uma estratificao vertical marcante, especialmente em reas onde o toldo protetor do topo das rvores quebrado.
Zoneamento. Mudanas horizontais no meio ambiente fsico se refletem em alteraes nos componentes vegetais e animais das comunidades ecolgicas. Pode ser encontrado em situaes terrestres onde as caractersticas do solo variam, por exemplo, de condies mais midas para condies mais secas.
Disperso Horizontal. A ocupao horizontal do espao, ou disperso, de plantas e animais tambm pode ser usada para descrever a estrutura de uma comunidade ecolgica. Trs so os padres bsicos de disperso: aleatrio, uniforme (ou regular) e agrupado (ou contagioso); dois padres podem ser adicionados pela combinao aleatrio/agrupado e uniforme/agrupado.
Estrutura Trfica. Outra categorizao produtiva da estrutura da comunidade o uso dos nveis trficos (relacionados com a nutrio).
Composio Filogentica. Talvez uma das maneiras mais bvias e fceis de compreender uma comunidade seja pela identificao dos conjuntos de espcies que a constituem. Assim, uma comunidade florestal pode ser descrita por sua filogenia (os relacionamentos de grupos de organismos de acordo com a sua histria evolutiva), como abeto, bordo ou carvalho, e uma comunidade animal como microartrpodes do solo, peixes bentnicos, insetos voadores e assim por diante.
ASPECTOS FUNCIONAIS DAS COMUNIDADES
Por consistirem de organismos vivos, as comunidades no so entidades estticas. Como as espcies que as compem, as comunidades so dinmicas e sofrem mudanas mais ou menos constantes da fisionomia e composio devido a mudanas tanto no meio ambiente como nas caractersticas dos ciclos de vida dos prprios organismos.
Mudanas Sazonais Por causa da mudana na inclinao do eixo da terra em relao ao sol, ao longo de 12 meses, a radiao solar total recebida em uma dada latitude geogrfica varia em diferentes pocas do ano.
VARIABILIDADE TEMPORAL AFETANDO OS RECURSOS NATURAIS
As comunidades mudam ao longo do tempo. Essas mudanas, possivelmente, se devem - a cataclismos raros tais como erupes vulcnicas ou a fatores estressantes menos raros, porm difceis de prever, como incndios nas florestas, furaces ou estressores antropognicos; ou a mudanas peridicas no meio ambiente, tais como mudanas sazonais, assim como modificaes induzidas pela prpria comunidade. Tomadas isoladamente, as comunidades tendem a seguir padres de mudana bem previsveis, chamados de sucesso.
PERTURBAES E SUCESSO
As sries de mudanas em comunidades observadas em ambientes recentemente criados, tais como novas ilhas, so denominadas sucesso primria. Sucesso secundria o termo usado para as mudanas sequenciais que ocorrem quando uma comunidade ecolgica estabelecida perturbada e subsequentemente passa por mudanas em sua estrutura e funo. Se a sequncia secundria segue o padro da primria, depende de uma srie de fatores abiticos ou biticos tanto de agentes da perturbao quanto de componentes da comunidade em re-desenvolvimento. Entre os primeiros esto os incndios, tempestades de gelo, inundaes, secas, ventanias, deslizamento de terras e grandes ondas. Entre os fatores biticos
esto interaes diretas tais como transformao em pasto, caa predatria, competio, infeco (doena) e os efeitos indiretos da modificao causada no hbitat por atividades como a escavao de tocas por vermes da terra e roedores ou o represamento de cursos de gua por castores. Muitas aes humanas tm considerveis consequncias: terraplanagem, construo e poluio.
MUDANAS PERIDICAS EM COMUNIDADES ECOLGICAS
Mudanas Sazonais em Recursos Naturais Muitos organismos, sejam eles migratrios ou no, apresentam padres sazonais de mudanas no uso de recursos naturais. De fato, a estrutura comunitria pode mudar significativamente no curso de um ano. No extremo, os organismos iro abandonas ou reduzir grandemente sua aquisio de recursos naturais em certas pocas do ano, passando a viver dos recursos estocados. Isto pode ser visto em animais que consomem seus estoques biolgicos tal como o tecido adiposo em associao com torpor ou hibernao para reduzir suas necessidades de recursos naturais.
Mudanas Sazonais no Modo de Obter Recursos Naturais bvio que quando a fonte que um organismo possui de um dado recurso natural muda de acordo com a estao do ano, o organismo muitas vezes ter de mudar a maneira pela qual obtm esse recurso. No entanto, mesmo quando o organismo no muda sazonalmente, as caractersticas do recurso ou do habitat podem mudar, e assim animais devem mudar seus comportamentos para obter os recursos necessrios.
Verticalidade dos Recursos Naturais e Ecologia Humana As populaes humanas precisam lidar com a distribuio vertical e tambm com a horizontal de recursos naturais do macroambiente. Esta uma
considerao fundamental para as populaes que vivem nos ecossistemas de florestas tropicais. Exceto em reas recentemente perturbadas, muitos dos recursos naturais comestveis das florestas tropicais, incluindo folhas, frutos e os animais que os comem, situam-se bem acima do cho, no dossel.
Sazonalidade na Ecologia Humana
Migrao Humana. Muitos grupos humanos precisam mover-se para seguir seus recursos naturais. Esse deslocamento encontra-se comumente entre os coletores e os pastoralistas, ao passo que as populaes que fazem colheitas tendem a uma possibilidade menor de migrao. No entanto, a migrao individual ocorre em muitos grupos agricultores, como pode ser visto em trabalhadores migrantes de fazendas nos Estados Unidos e entre pessoas que se engajam na onda sazonal de empregos. Nomadismo um termo reservado para o deslocamento envolvendo grupos sociais inteiros, devido necessidade de recursos naturais. Transumncia outra forma de migrao, encontrada em alguns grupos de pastores, na qual apenas uma parte do grupo social move-se para seguir as manadas, enquanto os outros membros do grupo permanecem em acampamentos permanentes.
Mudana Sazonal nos Recursos Naturais Entre Populaes Humanas. Muitos grupos humanos que so no-migratrios precisam ainda fazer ajustes em relao s mudanas sazonais na disponibilidade de recursos naturais. As mudanas podem ter a ver com variabilidade temporal tanto na quantidade como no tipo de recursos. Para caadores-coletores, isso envolve ajustar-se a mudanas sazonais nos recursos vegetais e animais para a dieta. Muitas dessas mudanas tm relao com os ciclos reprodutivos de plantas e animais, inclusive insetos. Para os agricultores, os ciclos sazonais giram em torno de estaes de plantio e de colheita. Nos pases em desenvolvimento, muitas pessoas nas reas rurais tm de encarar a diminuio de alimentos nos meses imediatamente anteriores colheita, o que provoca perda de peso corporal devido ao uso das reservas biolgicas (Ferro-Luzzi e Branca,1993). As populaes humanas tm muitas estratgias para lidar com a sazonalidade dietria, incluindo a estocagem dos alimentos, relaes de comrcio com outros grupos (Messer, 1989) e um calendrio de plantio que supra o cambiante tempo do incio das colheitas (Huss-Ashmore, 1993). A sazonalidade dos recursos alimentares tambm provoca mudanas sazonais em outras atividades econmicas (Harrison, 1988), levando a diferentes padres de atividade fsica. As mudanas sazonais de atividade podem provocar alteraes na necessidade de alimentos no decorrer de um ano.
CONCLUSO
Os humanos precisam equilibrar entre si a efetividade, a eficincia e o risco na obteno de recursos naturais, mas tm tambm de equilibrar consideraes de curto e de longo prazo. As grandes populaes e a tecnologia complexa dos povos modernos levaram a um enorme aumento no uso dos recursos naturais, com a taxa de uso frequentemente excedendo a taxa de renovao dos recursos no meio ambiente. Por toda a histria e pr-histria da humanidade, o desafio tem sido obter uma quantidade suficiente dos vrios recursos essenciais. Podemos estar alterando o desafio ecolgico para transform-lo em preveno do superconsumo. De modo claro, esto na ordem do dia os apelos para estratgias de uso dos recursos naturais que maximizem os ganhos sustentados. Essa mudana da otimizao de ganhos de curto prazo para ganhos de longo prazo precisa de alteraes que vo para alm da ecologia humana: vai requerer a criao de sistemas econmicos sustentveis (Owen e Chiras, 1995). Mudar para sistemas sustentveis implica alteraes polticas e socioculturais (Bennett, 1976), assim como uma mudana de valores referente ao relacionamento entre humanos e a natureza (Owen e Chiras, 1995), ao menos para a maioria das pessoas. A questo se os humanos podem fazer essas mudanas antes que a degradao a longo prazo de extensas reas da biosfera acontea. Apesar de serem desencorajadoras as observaes de que at mesmo grupos de caadores-coletores tendem a optar por ganhos de curto prazo, as pesquisas de opinio pblica nos anos 90 mostraram que a maioria das pessoas apoia os esforos ambientalistas em geral. As provas do sucesso de programas ambientalistas nos pases desenvolvidos comeam a se acumular, com ar e gua mais puros, assim como taxas rebaixadas de aumento no uso per capita de alguns recursos naturais entre os anos 60 e 90 (Easterbrook, 1995). .assim, o uso humano contemporneo dos recursos naturais se nos apresenta com uma combinao de otimismo e preocupao. Os humanos modernos esto comeando a mudar para uma viso de longo prazo do uso dos recursos naturais, mas devemos imaginar se a mudana pode ocorrer suficientemente rpido para contrabalanar a rapidez de nosso crescimento populacional e a crescente capacidade tecnolgica para explorar o meio ambiente em detrimento de ns mesmos.
PARTE 12 BIOMAS E POPULAES HUMANAS
INTRODUO
H duas maneiras comuns pelas quais os eclogos humanos classificam as populaes humanas. O primeiro mtodo, baseado na ecologia cultural. Os inuit e os mbuti so, ambos, exemplos de forrageadores, porm, muito complicado argumentar quanto s similaridades entre suas adaptaes ecolgicas. Os biomas so os grandes ecossistemas terrestres do mundo que ocorrem nas principais reas regionais ou subcontinentais. Desertos, florestas pluviais e savanas so exemplos de biomas, bem como o so florestas tropicais e decduas. Ao classificarmos as populaes humanas a partir de seus biomas, a ideia de que grupos humanos em um dado bioma necessitam lidar com as mesmas espcies de problemas ecolgicos.
CARACTERSTICAS E ORGANIZAO DE BIOMAS
Embora alguns eclogos reconheam dez ou mais biomas, iremos nos concentrar em seis: a tundra, a floresta boreal ou de conferas, a floresta decdua, os cerrados e savanas, o deserto e a floresta pluvial tropical. Esta sequncia segue, mais ou menos, uma direo ao sul, desde o plo norte at o equador. Em montanhas altas, tais como as Rochosas americanas, os Andes e o Himalaia, as linhas de diviso entre os biomas so de altitude e no de latitude. Os biomas encontrados em uma dada altitude tambm variam em relao s suas latitudes.
O Papel do Clima e do Solo na Distribuio dos Biomas O clima, ou seja, a interao da temperatura e das chuvas que prevalecem por perodos longos de tempo, tem o papel mais significativo sobre a determinao da localizao dos biomas. Como voc bem sabe, a temperatura est associada diretamente latitude, sendo o norte mais frio e o sul, mais quente, no hemisfrio norte, com o reverso ocorrendo no hemisfrio sul. Grandes padres de ventos, os quais esto associados s latitudes, so os responsveis, em larga escala, pela distribuio das precipitaes. O tipo de solo tambm tem um papel principal, mas menos crtico, sobre a distribuio dos biomas, primariamente graas interao com as plantas na ciclagem dos nutrientes.
OS PRINCIPAIS BIOMAS DO MUNDO
Tundra A Ecologia da Tundra rtica. A tundra, que significa plancie pantanosa, fica, em sua maior parte, ao norte da latitude 60 N e constitui cerca de 20 % da Amrica do Norte, incluindo ao redor de 2,5 milhes de Km da Groenlndia e 0,3 milhes de Km do Alasca (Bliss, 1988). Ela parece a cobertura de um sorvete sobre o norte da Europa e da sia. Embora haja diferenas considerveis de clima, cobertura de gelo, solos, tamanho da flora e composio das comunidades vegetais, a tundra rtica pode ser caracterizada pela ausncia de rvores, pela predominncia de plantas ans (de 5-20 cm de altura) e pela camada superior do solo, muito poroso e distinto, como resultado dos congelamentos e drenagens desta terra pobre.
O Bioma da Floresta Boreal
Ecologia do Bioma da Floresta Boreal. O bioma da floresta boreal, ou de conferas, tambm conhecido como taiga e como as grandes florestas do norte. Ela se localiza amplamente entre as latitudes 45N e 57N, mas se estende para o sul em uma poro mais alta da do leste dos Estados Unidos e em outros locais. Ela ocupa uma poro substancial do norte da Europa e cerca de 80% do que antes era conhecido como a Unio das Repblicas Socialistas Soviticas na sia. O clima vai de frio a gelado, como invernos longos e veres curtos, sendo que a mdia de temperatura mensal varia de -10C, no inverno, at 15C, no vero. As precipitaes so maiores que aquelas observadas na tundra e ocorrem, principalmente, no vero; a mdia anual de cerca de 60 cm.
O Bioma das Florestas Temperadas Decduas
Ecologia do Bioma das Florestas Temperadas Decduas. O bioma de florestas decduas o mais afetado pela habitao humana, primariamente porque, dentro de seus limites, ele confinou a maior parte das populaes por milnios, como consequncia do clima mais favorvel. Este bioma ocupa a
maior parte da metade leste dos Estados Unidos e da sia e praticamente toda a Europa. Embora o clima do bioma das florestas decduas varie consideravelmente de norte a sul e de leste a oeste, ele geralmente moderado, com veres frios a quentes, com um perodo definido de inverno, caracterizado pela neve e pelo congelamento no norte, e pelo frio e pela chuva nas pores mais ao sul. A mdia anual de chuvas de 80 cm a 150 cm. As temperaturas mais quentes do vero resultam em uma decomposio mais rpida, assim, menos serrapilheira acumulada no cho da floresta e o solo mais rico em nutrientes do que nos biomas de floresta boreal e rtico.
O Bioma do Deserto
Ecologia do Bioma do Deserto. O deserto tipicamente caracterizado por terras permeadas de arbustos nos quais as plantas encontram-se dispersas com muito solo nu entre elas. O creosote bush, a planta dominante dos desertos do sudoeste dos Estados Unidos, pode se apresentar em intervalos de espao de 5 a 10 metros. Os desertos ocupam cerca de um quinto da superfcie da Terra e so encontrados em todos os continentes. Para comparaes de tamanho, o deserto do Saara tem aproximadamente o tamanho dos Estados Unidos, e o grande deserto de Gobi, na sia, ainda maior. A aridez, em vez da temperatura, descreve melhor o clima dos desertos. A precipitao baixa e errtica, geralmente menor que 25 cm por ano, sendo que a maior parte ocorre nos meses de inverno. Em nveis mais extremos, o Saara recebe menos de 15 cm de chuva ao ano; sua poro central e o deserto ao norte do Chile recebem menos que isso. A partir de uma perspectiva da temperatura, h desertos frios e quentes: os quentes incluem o Saara, o Negev e o deserto arbico do Oriente Mdio, o deserto da Austrlia e os de Mojave e Sonora, na Amrica do Norte. O deserto da Grande Bacia norte-americana e as pores ao norte do deserto de Gobi so exemplos de desertos frios. Em ambos os tipos de desertos, as temperaturas do ar e do solo demonstram diferenas dramticas entre a noite e o dia. A temperatura do ar em um deserto quente pode chegar a 15C noite e a 40C ao meio-dia, enquanto a temperatura da superfcie, no mesmo perodo, pode variar de 0C a 65C.
O Bioma das Florestas Tropicais
A Ecologia do Bioma de Floresta Tropical. As florestas tropicais pluviais ocorrer a 10 ou mais ao norte ou ao sul do equador nas Amricas Central e do Sul, na frica central e oriental, no sudeste asitico, no leste indiano e no nordeste australiano, alm de ilhas ocenicas, geralmente dentro destas mesmas latitudes. Cerca de 40% da massa tropical e subtropical da Terra dominada por florestas abertas e fechadas: destas, 42% so florestas secas, 33% so florestas midas e somente 25% so de florestas tropicais pluviais (Murphy e Lugo, 1986). As florestas tropicais pluviais esto entre os ecossistemas mais antigos. Evidncias fsseis na Malsia e em muitos outros locais sugerem que elas tm existido continuamente por mais de 60 milhes de anos (Richards, 1973). Elas possuem uma considervel significncia ecolgica devido sua influncia sobre o clima, ao balano de carbono e de poluentes atmosfricos e aos seus diversos conjuntos de espcies, que representam grande potencial como novas fontes de alimentos, de fibras e de produtos medicinais e industriais (Jordan, 1985).
CONCLUSO
O uso de biomas como ferramenta de classificao pela ecologia de comunidades se provou til. H generalizaes dentro dos biomas com relao ao clima e s estruturas fundamentais das comunidades. Os biomas tambm so teis para a classificao de desafios ecolgicos impostos s populaes humanas. Entretanto, devemos ser cuidadosos para no transformar a ferramenta em algo heurstico (ou, em portugus claro, no devemos assumir que uma ferramenta classificatria, criada para simplificar um mundo complexo, de alguma forma se tornou uma representao do mundo real). Apesar de acreditarmos que o demnio vive nos detalhes e, portanto, de usarmos exemplos especficos, tambm temos a crena de que o conhecimento requer generalizaes.
PARTE 13 ENERGTICA ECOLGICA
INTRODUO
O fluxo energtico unidirecional em ecossistemas um dos princpios ecolgicos mais fundamentais. A energia proveniente do sol captada pela
clorofila (e alguns outros pigmentos) de algas verdes e de plantas em um processo conhecido por fotossntese. Estes receptores de energia so coletivamente conhecidos como produtores. Parte da energia por eles capturada usada em seu prprio metabolismo, desenvolvimento e reproduo, e parte repassada aos consumidores, diretamente para os herbvoros e, indiretamente, destes para os carnvoros. Os consumidores tambm se utilizam de parte desta energia em seu metabolismo, desenvolvimento e reproduo/ uma poro de sua energia, junto com parte da energia dos produtores, passada aos decompositores (bactrias e fungos). Este fluxo unidirecional de energia reflete-se na cadeia alimentar. Associados ao fluxo energtico, os nutrientes se movem nos ecossistemas, porm, de maneira cclica. Nutrientes como o carbono e o nitrognio so caracterizados por seus ciclos gasosos, enquanto outros, como o fsforo e o enxofre, possuem gases sedimentrias de curta ou longa durao.
RADIAO SOLAR
A radiao solar produz diretamente o aquecimento da Terra, bem como o ponto de partida do fluxo energtico em ecossistemas atravs do processo de fotossntese. Sua qualidade (por exemplo, o comprimento de onda ou sua cor), intensidade e durao so fatores crticos para a atividade biolgica.
Energia Solar O sol , essencialmente, um reator termonuclear que converte o hidrognio em hlio, um processo que libera uma quantidade tremenda de energia radiante na forma de ondas eletromagnticas. Estas emisses variam desde as de alta frequncia, como os raios gama e X (ambos de ondas curtas), at as de baixa frequncia, como as ondas do rdio (longas). Embora a energia se espalhe por este espectro amplo, 99% dela se encontra entre os raios ultravioleta e infravermelho (comprimentos de onda de 0,136 at 4,0 micra); somente cerca da metade desse valor visvel a ns (0,38 a 0,77 micra), a poro mais crtica para a maioria dos seres vivos. Do total energtico liberado pelo sol, somente 0,2 milionsimos dessa quantia atingem a atmosfera da Terra, o resto dissipado no espao. E a poro de
energia que chega na atmosfera do planeta atinge sua superfcie de maneira diferenciada devido a diversos fatores. Primeiro, a rotao da Terra resulta em variaes dirias que chamamos de dia e noite; segundo, devido inclinao da Terra no plano de seu eixo do equador com relao a seu plano orbital., o fluxo de energia varia sazonalmente com a latitude; e, terceiro, mais da metade da energia solar perdida conforme passa pela atmosfera, principalmente devido reflexo causada pelas nuvens e pela poeira e absoro por gases tais como o oznio, o oxignio e o vapor dgua, ou atravs da disperso difusa.
Mudanas de Intensidade e de Qualidade
Intensidade. A temperatura est diretamente relacionada ao grau, ou intensidade, da radiao solar. Quanto maior o grau de radiao, maior a temperatura; assim, mudanas da intensidade da luz do sol afetam diretamente tanto a temperatura quanto a atividade biolgica.
Qualidade. As mudanas mais dramticas da qualidade da luz solar ocorrem quando a luz penetra na gua, tendo, nesse momento, sua intensidade reduzida pela absoro e pela difuso.
Mudanas Dirias. J que uma grande quantidade de calor necessria para aquecer a gua (a quantidade de calor requerida para elevar a temperatura de uma dada substncia em uma quantia determinada tecnicamente conhecida como calor especfico), as mudanas de temperatura dirias so mnimas em ambientes aquticos, tipicamente na ordem de 1C a 2C. Em contraste, as mudanas de temperatura do dia para a noite na terra podem ser bem considerveis. A variao na superfcie do solo vai de 18C s 3 h e 30 min. at 65C s 13h e 30 min., uma mudana de 47C. E, embora a temperatura do ar a 120 cm de altura varie de cerca de 15C a 38C ao longo do dia, a temperatura a 40 cm sob o solo se mantm constante em cerca de 30C.
ENERGIA SOLAR E PRODUTORES
Medindo a Captura de Energia Solar
Nos estudos de fluxo energtico, o eclogo est interessado tanto na taxa quanto na quantidade de fotossntese que ocorre em um dado perodo de tempo em um ecossistema. A massa total de matria orgnica que produzida em tais condies recebe o nome de produo e sua taxa, produtividade. (Obviamente, estes termos tm seus paralelos na economia). A produo e a produtividade dos produtores so conhecidas como produo primria e produtividade primria, respectivamente; os consumidores recebem o prefixo secundrio ou tercirio, herbvoro ou carnvoro. Produtividade Bruta e Lquida A diferena entre a produtividade bruta e a lquida , como j foi notado, uma reflexo da quantidade da energia expendida na automanuteno, no crescimento, na transpirao (a evaporao da gua) e na reproduo.
Fatores que Influenciam a Produo e a Produtividade Como voc poderia esperar, muitos fatores influenciam as taxas e as quantidades de fotossntese em qualquer ecossistema. Em adio disponibilidade dos componentes qumicos bsicos da fotossntese, mudanas de fatores fsicos e biolgicos exercem influncia, assim como as modificaes dirias e sazonais, sobre a luz, a temperatura e a umidade.
FLUXO ENERGTICO DECOMPOSITORES
ATRAVS
DE
CONSUMIDORES
Ecossistemas de Detritos Os ecossistemas autotrficos, tais como o lago do pntano Cedar, dependem diretamente do influxo da radiao solar. Eles so caracterizados por uma dependncia da captura de energia pelos auttrofos fotossintetizantes e, secundariamente, pelo movimento da energia capturada atravs do sistema realizado pelos herbvoros e pelos carnvoros. Um grande nmero de ecossistemas funciona desta forma e numerosos herbvoros, carnvoros e onvoros, incluindo os humanos, so, mais ou menos, completamente dependentes de tais ecossistemas autotrficos.
Decomposio. A decomposio de matria orgnica ocorre em uma variedade de formas, dentre elas a lixiviao (por exemplo, com a gua percorrendo o material orgnico e dissolvendo seu contedo mineral) e a
fragmentao (a pulverizao do material em partculas pequenas). Entretanto, ela ocorre primariamente atravs da atividade de organismos que podem, por sua vez, facilitar tanto a lixiviao quanto a fragmentao. Os agentes primrios dos ltimos estgios da decomposio so os micrbios (principalmente bactrias, em animais, fungos, plantas) atravs dos processos de seu prprio metabolismo. Para atingir suas necessidades metablicas, os decompositores liberam enzimas digestivas em seu ambiente imediato; estas enzimas quebram alimentos complexos em compostos simples que podem ser prontamente absorvidos pelo fungo ou pela bactria.
CADEIAS E REDES ALIMENTARES
Implcito nos diagramas e nas discusses sobre o fluxo energtico, seja ele atravs de herbvoros/carnvoros ou atravs da decomposio de detritos, h uma conexo entre os organismos cuja existncia depende de outros, no nvel trfico prximo mais baixo. Tais ligaes so geralmente nomeadas de cadeias alimentares.
O Tamanho das Cadeias e das Redes Alimentares
Tambm implcito no fluxo energtico atravs de um ecossistema que o nmero de nveis trficos limitado devido diminuio da disponibilidade de energia resultante das ineficincias quanto transferncia de energia de um nvel trfico para o outro. Se este o caso, ento, uma hiptese intuitiva que abrange o tamanho da cadeia alimentar a de que as cadeias so maiores em ecossistemas com maior produtividade primria. Esta hiptese conhecida como a hiptese energtica (Hutchinson, 1959). Baseado em uma reviso dos estudos sobre os ecossistemas pobres e os altamente produtivos, entretanto, Pimm (1982) concluiu que no h evidncias suficientes que sustentem a hiptese da energtica. Eles concluram, entretanto, que as dimenses espaciais de um ecossistema esto relacionadas ao tamanho das cadeias alimentares, sendo que ambientes tridimensionais possuem cadeias maiores do que os bidimensionais. Os ambientes bidimensionais foram considerados essencialmente achatados, como a tundra, os campos, o mar, o fundo de um lago ou um ambiente intermars; ambientes tridimensionais foram considerados slidos, como o dossel de uma floresta ou uma coluna de gua em oceano aberto.
PARTE 14 A ADAPTAO HUMANA E O FLUXO ENERGTICO
Para sobreviver, as populaes humanas precisam invadir os sistemas naturais de fluxo energtico nos ecossistemas onde residem. A energia um dos componentes principais da dieta, mas suas necessidades energticas vo muito alm das preocupaes da dieta. Na verdade, a maioria das atividades humanas esto relacionadas, de alguma maneira, com a energia (Harrison, 1982). Muitos antroplogos, liderados por Leslie White (1959, 1969), sugeriram que a capacidade crescente de aquisio e de utilizao de energia de nossa espcie tem sido um dos principais fatores da mudana de grupos caadorescoletores para as sociedades modernizadas. Estudos sobre o fluxo energtico em grupos humanos examinam como a populao se apropria da energia de que necessita incluindo a comida ou o combustvel a partir do ambiente. Nesse aspecto, os humanos so muito parecidos com os demais organismos vivos, exceto pelo componente combustvel. As medidas de energia em populaes humanas tm trs tipos (Thomas, 1974): produo, perda e consumo. A produo energtica mede quanta energia pode ser obtida por um grupo e equivalente produo secundria medida para outros consumidores nas cadeias alimentares. A perda energtica mede quanta energia o grupo usa, ou gasta, em suas diversas atividades. Ela , geralmente, quantificada atravs da medida do consumo de oxignio, o qual est relacionado com a perda energtica, porque o gasto energtico humano baseado no metabolismo aerbico. O consumo energtico refere-se quantidade de energia que um grupo usa, seja ela atravs da aquisio atravs da dieta ou do consumo de combustveis. Ele baseado, em parte, nos estudos sobre aquisies nutricionais, bem como nas medidas de consumo de combustveis.
MEDINDO A UTILIZAO DE ENERGIA EM POPULAES HUMANAS
J que a energtica humana uma moeda corrente de recursos em uso, os eclogos humanos devotaram muito tempo s medies de como os humanos obtm e usam energia. Eles utilizam diversas tcnicas para medir sua produo e seu consumo.
A Medida da Produo Energtica Humana
A produo energtica geralmente medida para um grupo, em oposio a medidas individuais, j que a maioria das pessoas coopera para obter os recursos energticos. Um grupo pode ser chamado de uma unidade domstica, um grupo de procura ou de outra forma de unidade social. A produo de energia per capita pode ser obtida atravs da mdia de produo do grupo, mas a participao de um indivduo em muitos grupos e exemplos de desigualdade na diviso tornam esses clculos mais difceis. Eclogos humanos devem considerar a variabilidade sazonal da produo energtica em suas medies. Observaes sobre uma populao em um perodo no podem ser extrapoladas para o ano todo. Assim, estudos sobre a energtica humana, convencionalmente, duram um ano inteiro de atividade.
A Medida do Consumo Energtico Humano A medio do consumo energtico em indivduos da populao geralmente se refere ao valor calrico da dieta para todos os seus membros em um dado perodo de tempo e, portanto, s quantidades de alimento consumidas. Para obter valores precisos, todos os alimentos ingeridos por um indivduo em um dado perodo de tempo devem ser pesados separadamente antes do consumo. Ento, o valor calrico de cada item alimentar derivado de uma tabela de nutrientes. Outro mtodo para a determinao do consumo energtico , simplesmente, pesar as pessoas em dois momentos distintos. Uma diferena de peso significa uma diferena entre o consumo energtico e o gasto energtico durante o perodo de tempo entre as medies; assim, uma das medidas pode ser calculada se a outra for conhecida (Himes, 1991). Este mtodo no muito preciso, mas til na identificao de balanos energticos, positivos ou negativos, em populaes humanas.
Eficincia Energtica Para as populaes humanas, a eficincia energtica geralmente calculada como a energia produzida dividida pela energia gasta em um dado perodo de tempo ou em uma determinada atividade. A eficincia uma medida do ganho energtico em rede em uma dada atividade; ou seja, ela mede quanta energia um ser precisa gastar a fim de adquirir algum recurso que contenha energia.
CONCLUSAO: A UTILIDADE DOS ESTUDOS ENERGTICO PARA A ECOLOGIA HUMANA
SOBRE
FLUXO
Os estudos sobre fluxo energtico vo alm das contribuies ao conhecimento sobre o uso de recursos, eles podem fornecer informaes necessrias para entender a fertilidade, os padres de crescimento fsico, a variabilidade do tamanho corpreo adulto, a capacidade de trabalho dos indivduos e os padres de morbidez e de mortalidade de pessoas de diversas populaes (Baker, 1974). Mais amplamente, a energia fundamental para a estrutura e para a troca de informaes (Odum, 1971; Adams 1974). Assim, os estudos sobre o fluxo energtico podem ser extrapolados em consideraes sobre o fluxo de informaes, o que permite o conhecimento de processos culturais que ocorrem dentro de populaes (Odum, 1971). Os estudos sobre fluxo energtico humano vo alm da simples descrio dos caminhos da energia eles so ferramentas para o entendimento da dinmica adaptativa das populaes humanas.
PARTE 15 CICLAGEM DE NUTRIENTES EM ECOSSISTEMAS
INTRODUO
A importncia dos nutrientes e de sua ciclagem atravs dos ecossistemas tem sido discutida inmeras vezes. Agora, apropriado reconhecer, mais detalhadamente, os padres cclicos desses nutrientes, buscar os provveis padres existentes entre eles e, subsequentemente, considerar os efeitos resultantes das interaes inadvertidas e propositais dos humanos com eles. Mas, inicialmente, uma breve introduo sobre a natureza e a importncia dos nutrientes ser apropriada na construo do cenrio necessrio para o conhecimento da importncia dos processos cclicos.
NUTRIENTES
Embora os carboidratos possam ser fotossintetizados a partir do hidrognio, do carbono e do oxignio existentes na gua e no dixido de carbono, os compostos orgnicos mais complexos necessitam de elementos adicionais tanto em quantias considerveis, como no caso do nitrognio e do fsforo, quanto em quantidades mnimas, como no caso do zinco e do molibdnio. Alm disso, a fotossntese e outras reaes metablicas, tanto em plantas
quanto em animais, ocorrem na presena de enzimas que, em si mesmas, contm uma variedade de elementos trao. Aqueles elementos que so necessrios em quantidades relativamente grandes so conhecidos como macronutrientes.
Macronutrientes Os macronutrientes podem ser classificados em dois grupos: (1) aqueles que constituem mais de 1% do peso orgnico seco carbono, oxignio, hidrognio, nitrognio e fsforo; e (2) aqueles que constituem de 0,2 a 1% do peso orgnico seco enxofre, cloro, potssio, sdio, clcio, magnsio, ferro e cobre. Em humanos, h apenas 4 macronutrientes: o hidrognio 63%; o oxignio 25,5%; o carbono 9,5%; e o nitrognio 1,4% (Frieden, 1972).
Micronutrientes Os micronutrientes conhecidos incluem o alumnio, o arsnio, o boro, o brometo, o cromo, o cobalto, o flor, o glio, o iodo, o mangans, o molibdnio, o nquel, o selnio, o silicone, o estrncio, o estanho, o titnio, o vandio e o zinco. Na verdade, alguns destes micronutrientes podem ser considerados macronutrientes em algumas espcies, ao passo que alguns macronutrientes, o sdio e o cloro nas plantas, por exemplo, podem ser micronutrientes para outras espcies.
Interao de Nutrientes A presena ou a ausncia de um nutriente essencial pode afetar adversamente a disponibilidade ou a atividade de outro nutriente.
O Efeito Estufa. O dixido de carbono tem um papel crtico sobre o controle do clima da Terra porque, como um aerossol, ele absorve, reflete e espalha a radiao que chega do sol; entretanto, ele absorve e torna a emitir a radiao de volta para o espao. Este ltimo fenmeno resulta no que conhecido popularmente como Efeito Estufa, uma analogia ao que ocorre em uma estufa de plantas. Entretanto, esta analogia no perfeita. Os vidros de uma estufa e os gases, como o dixido de carbono no efeito estufa, permitem a passagem da luz do sol at a superfcie e mantm o calor do lado de dentro. A diferena que, em uma estufa, todo o calor fica preso; em contraste, os gases do efeito estufa
prendem apenas a radiao infravermelha. Em ambos os casos, as temperaturas da estufa e da Terra aumentam.
Poluio: uma Definio Tornar ou deixar algo sujo; tornar impuro; desrespeitar; profanar- este o significado do verbo poluir de acordo com o dicionrio Webster New Collegiate. A poluio ambiental a alterao desfavorvel de nossos arredores, principalmente como um subproduto das aes do homem, atravs de efeitos diretos ou indiretos de modificaes dos padres energticos, dos nveis de radiao, das constituies fsicas e qumicas e das abundncias dos organismos. Estas mudanas podem afetar o homem diretamente, ou afetar suas reservas de gua e de produtos agrcolas ou de outra natureza, seus objetos pessoais e suas possesses ou suas oportunidades de recreao e de apreciao da natureza.
PARTE 16 POPULAES HUMANAS E OS CICLOS DE NUTRIENTES
INTRODUO
Os humanos, como outros organismos, utilizam materiais dos ciclos naturais de nutrientes e, dessa forma, tornam-se, eles mesmos, parte desses ciclos. As pessoas necessitam praticamente dos mesmos nutrientes requeridos pela maioria dos organismos, embora, em alguns casos, as propores sejam diferentes. Para a maioria dos animais, boa parte dos nutrientes obtida a partir dos alimentos, embora sejam usadas algumas fontes alternativas, tais como a gua oriunda de fontes na superfcie e o sal a partir do mar. No caso dos humanos, os nutrientes so, geralmente, encontrados em fontes no consideradas como alimentos (pelo menos, no tradicionalmente), desde os laboratrios qumicos das grandes corporaes fabricantes de alimentos processados at os campos de petrleo que fornecem os fertilizantes necessrios ao cultivo. H diversos nutrientes que devem ser considerados quando lidamos com a ecologia humana. As necessidades humanas de energia podem ser supridas, atravs de nossos alimentos, pelos carboidratos e pelas gorduras, molculas compostas de trs elementos diferentes: o carbono, o hidrognio e o oxignio. Na verdade, necessitam de fontes de gua, mesmo sendo ela composta por
hidrognio e oxignio, dois elementos constantemente encontrados nas fontes alimentares.
POPULAES HUMANAS E O CICLO HIDROLGICO
O ciclo hidrolgico envolve consideraes globais sobre a precipitao, a evaporao e a drenagem da gua da superfcie da terra at o oceano; e a adio de gua, a longo prazo, biosfera a partir de fontes geolgicas profundas atravs de erupes vulcnicas, bem como fontes extraterrestres, os cometas. Os oceanos de hoje so, em larga escala, o resultado destas ltimas duas fontes agindo ao longo de bilhes de anos e, assim, a vida na Terra depende dos cuspes dos vulces e do suco dos cometas. Os humanos influenciam o ciclo hidrolgico global, principalmente atravs de seus efeitos sobre os padres de drenagem. O fluxo de gua em direo aos oceanos foi acelerado nos locais onde as pessoas construram cidades de concreto e estradas sobre grandes reas de terra. Isso efetivamente reduziu a volta de gua para a superfcie, o que diminuiu a quantidade dela disponvel para o uso. As pessoas tambm tm usado bombas para trazer a gua do subsolo superfcie. Embora isso tenha aumentado a disponibilidade de gua nos ecossistemas de superfcie, em alguns casos levou ao desgaste total de diversos reservatrios subterrneos.
A Economia da Escassez de gua
A gua como Moeda Corrente. Assim como o que ocorre com a energia, quando a gua escassa, ela um fator principal da economia de uma populao. Foi notado que a energia geralmente utilizada como substituta ao dinheiro nas quantificaes das atividades ecolgicas humanas. Em regies onde a gua escassa, ela pode ser utilizada da mesma maneira. Na verdade, como regra geral, qualquer re3curso escasso e limitante em uma sociedade serve como a mais til moeda na determinao de clculos ecolgicos. De forma similar energia, a gua pode ser convertida em outras unidades de interesse para a ecologia.
Para aos pases ridos, a gua pode ser to desejada quanto abundante se quiserem pagar o seu preo. O preo pode ser atravs da compra e do transporte direto a partir de outras naes, ou atravs do deslocamento de icebergs das regies polares. Infelizmente, o custo da gua em energia, em unidade monetria ou em qualquer outro tipo de valor que se queira usar , geralmente, muito alto.
Irrigao A agricultura intensiva requer grandes quantidades de gua por unidade de terra utilizada nas plantaes. Estes requerimentos, inevitavelmente, levam o fazendeiro a fornecer gua sua plantao em quantidades maiores do que as fornecidas pela chuva. As prticas de irrigao resultantes variam extensivamente pelo mundo, tomando a forma de mudana de cursos de rios, ou o fornecimento de gua para os campos a partir de poos artificiais. A irrigao, muitas vezes, implica o manejo, em larga escala, de fontes hdricas, sendo a gua transportada atravs de aquedutos, tubulaes ou canais, por longas distancias. H diversos exemplos de projetos de grandes construes utilizadas para o transporte de gua.
REQUERIMENTOS POR SAL E A ADAPTABILIDADE HUMANA
O sal outro nutriente requerido pelas populaes humanas, bem como por outros organismos. O sal obtido diretamente a partir do sal seco, de salinas em gua potvel e a partir de fontes de alimento. Os problemas de excesso de salinidade podem constituir um grande desafio para os agricultores que se utilizam da irrigao. Requerimentos por sal no influenciaram tanto a ecologia humana como outros nutrientes; porm, em regies especficas, nas quais o composto escasso, ele foi um importante fator econmico.
Densidade Populacional e Recursos Proticos Enquanto as protenas e demais nutrientes necessrios podem ser encontrados em uma dieta vegetariana razoavelmente balanceada, a disponibilidade de protenas de origem animal e de alta qualidade tem sido associada densidade populacional em muitas regies do mundo.
RETORNANDO OS NUTRIENTES PARA AS PLANTAS: POPULAES HUMANAS E O USO DE FERTILIZANTES
A interao humana com os ciclos de nutrientes vai alm da ingesto destes. A vasta maioria das populaes humanas depende da agricultura como a maior fonte de alimento e, portanto, precisam capacitar suas plantas cultivadas a obter os nutrientes necessrios a partir de seus ambientes. Na verdade, os ciclos de nutrientes naturais so, geralmente, interrompidos pelas atividades agrcolas, e os humanos, usualmente, aplicam nutrientes, na forma de fertilizantes, em suas plantaes a fim de obter a produo adequada. H uma grande diversidade tanto na maneira quanto no grau em que as populaes humanas utilizam os fertilizantes em suas plantaes.
A Irrigao como um Transportador de Nutrientes A gua de irrigao geralmente carrega consigo argila contendo nutrientes que podem ser assimilados pelas plantas cultivadas. Em alguns locais, este processo ocorre sem que haja muita interveno humana. Muitas pessoas utilizaram ciclos de enchentes sazonais dos rios tanto para irrigar quanto para fertilizar seus campos. A irrigao atravs de meios artificiais tambm carrega nutrientes para as plantaes. Por exemplo, a gua de irrigao do arroz carrega muitos nutrientes para estas plantas. Tambm cianofceas, que crescem na gua de irrigao aquecida, fixam o nitrognio, adicionando, portanto, nutrientes mesma (Geertz, 1963).
O Uso Humano de Fertilizantes Orgnicos
Fertilizantes orgnicos naturais de origem animal e humana tm sido utilizados por agricultores h milnios. Prticas compostas, utilizando tais fertilizantes orgnicos, foram muito importantes na sustentao de sistemas agrcolas intensivos em muitos lugares por todo o mundo.
CONCLUSO Os humanos sempre foram parte dos ciclos naturais de nutrientes, mas eles adicionaram complexidade a esses ciclos. Alm dos alimentos, os humanos utilizam materiais para suas vestimentas, abrigos, ferramentas e combustveis.
Estes incluem materiais orgnicos e inorgnicos. Em tempos recentes, um novo tipo de material se tornou importante: o sinttico (usualmente derivado de fontes de combustveis fsseis). A maioria deste material renovvel, e o uso prudente permite sua reposio. Alguns materiais no so renovveis e, portanto, precisam ser reutilizados se os humanos desejarem continuar a uslos no futuro (Brown, 1970). Dos materiais a serem reutilizados, alguns so parte dos ciclos naturais de nutrientes, enquanto outros so parte de sistemas de reciclagem unicamente humanos. O uso humano de recursos no-renovveis aumentou dramaticamente desde a revoluo industrial. A minerao de metais e de combustveis fsseis se tornou mais cara conforme recursos mais facilmente obtidos se acabaram. Precisamos fazer escolhas muito srias entre a reciclagem e a explorao de novos recursos. Claramente, a adaptao humana em longo prazo depende de nossa habilidade de realizar escolhas inteligentes quanto ao uso de nossos materiais.
PARTE 17 MODELOS DE OTIMIZAO NA ECOLOGIA HUMANA
A SOCIOBIOLOGIA E OS MODELOS DE SUCESSO REPRODUTIVO
A sociobiologia (algumas vezes chamada de socioecologia) lida com a seleo natural de caracteres sociais comportamentais (Wilson, 1975). Ela se baseia na noo de que o comportamento , em sua maioria, determinado geneticamente (ou, pelo menos, lida com componentes comportamentais sob forte influncia gentica), e que tais comportamentos esto sujeitos seleo evolutiva (Trivers, 1985). Os efeitos dos comportamentos sociais so medidos em termos do sucesso reprodutivo relativo os clculos so realizados atravs da razo custo/benefcio, tanto para o ator (o indivduo que apresenta o comportamento) quanto para o recipiente (o indivduo, da mesma espcie, para quem o comportamento dirigido). H quatro possibilidades: ambos ganham (definido como comportamento cooperativo); o ator ganha, mas h um custo para o recipiente (definido como comportamento cooperativo); o ator ganha, mas h um custo para o recipiente (definido como comportamento egosta); o ator confere um benefcio ao recipiente com algum custo (chamado de comportamento altrusta) e ambos sofrem um custo (definido como comportamento malicioso). Muito do interesse da sociobiologia reside na noo de que os organismos no possuem interesse idnticos em suas interaes. Se a seleo evolutiva determinou seu comportamento, cada um deles deve se comportar de uma
maneira que tender a maximizar seu sucesso reprodutivo; assim, passaro o material gentico, relacionado a tais comportamentos, para seus filhos, que, por sua vez, iro se comportar de maneira similiar. Dessa forma, pode-se conceber uma corrida armamentista, na qual os indivduos iro interagir socialmente a fim de aumentar seu prprio sucesso reprodutivo, mesmo com o prejuzo de outros.
Comportamentos Sociais Estudos pela Sociobiologia
Cooperao. Em casos de comportamento cooperativo, a seleo evolutiva favorece ambos os indivduos ou espcies. Entretanto, os sociobilogos, algumas vezes, consideram vantagens relativas de carter cooperativista a longo prazo, ou seja, eles reconhecem que um indivduo pode ser relativamente mais beneficiado do que outro.
Comportamento Egosta. No comportamento egosta, o ator beneficiado, enquanto h um custo para o recipiente. A seleo natural, operando sobre o ator, pode favorecer atos egostas, mas se espera que sejam selecionados os recipientes que evitem estes custos. Pode-se conceber uma luta evolutiva entre a tendncia a realizar atos egostas e a tendncia de evit-los.
Altrusmos. No comportamento altrusta, o ator tem um custo, enquanto o recipiente ganha a partir da interao. Isso faz pouco sentido em termos evolutivos, a no ser que o conceito de desempenho inclusivo seja invocado. Na verdade, h trs formas nas quais a seleo natural pode favorecer o altrusmo: parentesco (desempenho inclusivo), reciprocidade e parasitismo.
Comportamento Malicioso. No comportamento malicioso, tanto o ator quanto o recipiente sofrem. Pode parecer que a seleo natural agiria contra este tipo de comportamento. Mas ele ocorre algumas vezes e pode estar relacionado ao desempenho inclusivo. Ou seja, voc pode ferir um competidor (sexual ou outro) mais do que voc ferido.
A Sociobiologia Aplicada ao Comportamento Humano Uma das grandes questes acerca da sociobiologia envolve tentativas de aplic-la aos humanos. Enquanto algumas preocupaes tm suas razes no
uso supersimplificado da sociobiologia, sugerindo que todos os comportamentos humanos so inatos (determinados biologicamente) e, portanto, imutveis (por exemplo, Ardrey, 1961; Lorenz, 1963), h tambm consideraes em trabalhos de cientistas notveis, tais como E. O Wilson (Lumdsen e Wilson, 1981). Controvrsias e discordncias ocorreram entre antroplogos, geneticistas e outros cientistas (por exemplo, Sahlins, 1976; Alexandre, 1977) . Uma das questes se refere aos aspectos genticos versus os aspectos ambientais (aprendizado) dos comportamentos. A sociobiologia assume que os comportamentos so determinados (ao menos a maioria deles) pela gentica e, portanto, so susceptveis seleo natural.
ESTRATGIAS DE FORRAGEIO TIMO
Modelos de forrageio timo que foram aplicados s populaes humanas incluem aqueles usados na explicao de escolhas de dieta, de seleo de locais para caa, de decises acerca de quanto tempo gastar no forrageio em determinado local, a relao dos padres humanos de assentamento com os padres de distribuio das caas e explicaes dos padres de diviso de alimentos.
Modelos de Diviso de Alimentos
A diviso de alimentos com parentes prximos facilmente explicada pelos princpios da sociobiologia; quando a diviso ocorre alm dos limites da famlia que a explicao se torna muito mais complicada. Uma explicao concentra-se no uso do comercio, atravs do qual um indivduo negocia um recurso de pouco valor por outro mais valioso (Kaplan, e Hill, 1985). Aqui, o valor depende das percepes e das circunstncias de um indivduo.
PARTE 18 A ECOLOGIA HUMANA E A ECOLOGIA DOS HUMANOS
Os Desafios Enfrentados pela Cincia da Ecologia Humana Os problemas que confrontam os eclogos humanos precisam ser multiplicados diversas vezes para que sejam entendidos os desafios aos quais eles so submetidos. Aqui, um entendimento de todas as incertezas acerca do comportamento humano deve ser adicionado s complexidades das interaes
ambientais entre espcies, antes que seja possvel uma compreenso completa da ecologia humana. Esta compreenso pode ser impossvel de ser atingida.
O Ambientalismo e a Ecologia Humana. Da mesma forma que a confuso entre o ambientalismo e a cincia da ecologia humana comum, o ambientalismo tem substitudo ainda mais a cincia objetiva da ecologia humana.
A Degradao dos Ecossistemas Os humanos, quando se adaptam a um ecossistema, tendem a otimizar para o imediato, em vez de pensar em longo prazo, no qual foi demonstrado que os modelos ecolgicos para o comportamento humano parecem mais preciso quando os ganhos em curto prazo so focalizados. Isso implica que os comportamentos que maximizam a produtividade de recursos, com o efeito da degradao dos ecossistemas, sero adotados pelas populaes humanas.
A Converso de Florestas em Campos. Um dos exemplos mais claros da degradao antropognica a existncia dos desertos verdes do sudoeste asitico. Estas reas vastas apresentam gramneas imperata, em locais onde antes cresciam florestas pluviais, resultado de mau gerenciamento de agricultura de coivara no passado.
Eroso. A eroso outro srio efeito da degradao antropognica dos ecossistemas. Um dos exemplos mais claros vem das ilhas do Pacfico, tais como o leste da Polinsia e o Hava, onde a eroso no somente levou perda de superfcie nas terras altas, mas tambm cobertura das terras baixas com solo depositado.
Ameaas Biodiversidade Um dos mais graves impactos das atividades humanas recentes tem se dado sobre a biodiversidade, o nmero de abundncia de espcies, atravs de extines globais em massa.
Extines Antropognicas no Passado. Acredita-se que as contribuies humanas s extines tenham uma longa histria. Por exemplo, especula-se que a extino em massa ocorrida no final do Pleistoceno, a qual, diferentemente dos demais eventos, foi restrita aos animais terrestres, particularmente aos grandes mamferos, deve-se, principalmente, s atividades de caa de grupos humanos.
Extines Antropognicas no Presente. Os tempos modernos trouxeram consigo uma habilidade aprimorada dos humanos em afetar o ambiente. As atuais extines antropognicas em massa so causadas por diversos fatores, incluindo as superplantaes (como os exemplos de caa desenfreada ocorridos no passado), a destruio ou alterao do habitat fsico, a substituio por espcies introduzidas, a poluio e a hibridizao com outras espcies e subespcies devido s atividades humanas (Wilson, 1992).
VALORES PARA A NATUREZA
O Valor Econmico da Natureza A natureza , fundamentalmente, um sistema de suporte de vida para todos os organismos, um sistema que desprezamos at que haja um grande colapso, como um terremoto, um tornado ou uma enchente. Os produtos e processos naturais que ocorrem nos ecossistemas so os componentes deste sistema de suporte: eles so crticos para a sobrevivncia e, portanto, possuem um valor econmico e em outros termos. Os produtos naturais dos ecossistemas dos quais os humanos dependem incluem gros, vegetais, alimentos marinhos, caa e outros animais, madeira para combustveis, produtos farmacuticos, dentre muitos outros. Como produtos, eles possuem valor econmico.
CONCLUSO
Ningum pode deixar de se impressionar com o sucesso adaptativo de nossa espcie. Desde o incio dos homindeos no ambiente mosaico das florestas e savanas africanas, h quatro milhes de anos, os humanos habitaram praticamente todos os biomas de superfcie da Terra. Fomos realmente frutferos e nos multiplicamos. Os humanos crescentemente direcionaram os
recursos do planeta, mais e mais, para suas prprias necessidades, incluindo o uso de recursos para evitar, ou ao menos para proteger, a si mesmos, dos muitos estressores ambientais pelos quais eles uma vez foram afligidos. A questo : somos agora vtimas de nosso sucesso? Neste, e em outros captulos, vimos que muitos eclogos acreditam que estamos enfrentando problemas ambientais muito srios causados por nossas atividades. O problema que, em praticamente todos os casos, as limitaes de nosso conhecimento sobre a ecologia, especificamente a humana, no nos permitem muita preciso em nossas constataes, quanto menos consertar os erros. Nossas constataes sobre a cincia da ecologia humana e a ecologia dos humanos, portanto, so conectadas. Melhorias na primeira so necessrias para que o mesmo ocorra com a segunda. Nossa maior esperana a de que um dos leitores deste texto ir desenvolver um aumento de interesse no estudo da ecologia humana e melhorar nestes tempos perplexos. Ns, humanos, somos mais bem vistos como uma espcie incrivelmente adaptvel; parte desta adaptao deve ocorrer devido a mudana causadas por nosso prprio comportamento. Vimos que nossa adaptao consiste de ganhos balanceados de efetividade e de eficincia com redues de risco. Os riscos potenciais, delineados aqui, so to grandes que no obrigam a tomar medidas para reduzi-los, mesmo no estando muito certos acerca de sua magnitude. Ainda est para ser visto se poderemos continuar nos adaptando s mudanas que causamos em nosso prprio jogo evolutivo.
QUESTIONRIO
1. Na organizao de ecossistemas, defina e descreva os principais grupos de organismos. Produtores so o primeiro passo de um movimento de energia atravs de um ecossistema. este grupo de organismos, normalmente verdes e clorofilados, que capturam a energia radiante do sol, na verdade produtores no produzem energia: eles convertem ou a transmitem de uma forma, a radiante, a outra, qumica. Um auttrofo (literalmente, auto-alimentador) um organismo que captura energia e, subsequentemente, a utiliza para sintetizar molculas que servem aos requerimentos nutricionais de seu prprio crescimento e
metabolismo. Em contraste, um hetertrofo (literalmente, alimentado por outro) um organismo cujas necessidades nutricionais so atingidas atravs da ingesto de outros organismos: hetertrofos so tambm conhecidos como consumidores. Consumidores so organismos que derivam sua nutrio dos produtores: ou seja, so hetertrofos. As correlaes auttrofo-hetertrofo, ou produtor-consumidor primrioconsumidor secundrio, resultam em um movimento sequencial de energia conhecido como a cadeia alimentar. Decompositores, basicamente bactrias e fungos, so hetertrofos que derivam sua nutrio a partir de produtores (a maioria dos fungos) e consumidores (principalmente as bactrias). Em vez de ingerir seu alimento, como no caos dos outros hetertrofos, os decompositores liberam enzimas de seu corpo capazes de digerir os tecidos animais e vegetais, em seguida, esses produtos processados so absorvidos.
2. Quais so as causas da desertificao? Dentre os povos criadores, o pastoreiro a principal causa da desertificao, particularmente em situaes nas quais as seca prolongadas. O desmatamento de florestas para obteno de combustvel, removendo a proteo natural contra ventos a fonte de uns do solo, outro fator, o qual extenuado pela necessidade das populaes pelo produto. O cultivo e a irrigao, o pasto em excesso, o desmatamento, a minerao, a recreao e a urbanizao so todos fatores envolvidos na desertificao.
3. O que causa a deficincia de Vitamina A? A vitamina A necessria para o funcionamento normal de certas clulas epiteliais do corpo. A deficincia pode causar a cegueira noturna, dei xar a pele seca e escurecer a crnea. Se a deficincia prolongada, a cegueira permanente pode ocorrer.
4. Defina poluio.
Uma definio preliminar de poluio um recurso que est fora de lugar no ambiente, em grande ou pequena quantidade a partir da perspectiva de comunidades ecolgicas especficas. Uma das principais conexes entre populao e doena parece estar na exposio a grandes concentraes de certos produtos qumicos elaborados em processos industriais e altas taxas de certas formas de cncer. Trs das principais formas de poluio so: ar, gua e lixo slido.
5. Qual o principal recurso natural essencial ao homem? Comida um recurso natural essencial para todas as populaes humanas e tem merecido ateno especial por parte dos estudiosos da ecologia humana, em particular dos eclogos culturais.
9. KRASILCHIK, M. Prtica de ensino de Biologia. 4. ed. So Paulo: EDUSP, 2008. Prtica de ensino de Biologia
Tendncia do ensino de Biologia no Brasil
Introduo
No estgio atual do ensino brasileiro, a configurao do currculo escolar dos ensinos mdio e fundamental deve ser objeto de intensos debates, para que a escola possa desempenhar adequadamente seu papel na formao de cidados. Como parte desse processo, a Biologia pode ser uma das disciplinas mais relevantes e merecedoras da ateno dos alunos, ou uma das disciplinas mais insignificantes e pouco atraentes, dependendo do que for ensinado e de como isso for feito.
Admite-se que a formao biolgica contribua para que cada indivduo seja capaz de compreender e aprofundar as explicaes atualizadas de processo e de conceitos biolgicos, a importncia da cincia e da tecnologia na vida moderna, enfim, o interesse pelo mundo dos seres vivos. Esses conhecimentos devem contribuir, tambm, para que o cidado seja capaz de usar o que aprendeu ao tomar decises de interesse individual e coletivo, no contexto de um quadro tico de responsabilidade e respeito que leve em conta o papel do homem na biosfera. O significado da cincia e da tecnologia na sociedade contempornea merece a ateno especial do professor de Biologia, para que se evite tanto posturas de respeito temeroso alienante como uma atitude de desconfiana que atribui aos cientistas muitos dos atuais problemas da humanidade. Um conceito, cada vez mais presente nas discusses dos educadores, o da alfabetizao biolgica, referindo-se a um processo contnuo de construo de conhecimentos necessrio a todos os indivduos que convivem nas sociedades contemporneas. Um dos modelos desse conhecimento admite quatro nveis de alfabetizao biolgica: 1. Nominal quando o estudante reconhece os termos, mas no sabe seu significado biolgico. 2. Funcional quando os termos memorizados so definidos
corretamente, sem os estudantes compreendem seu significado. 3. Estrutura quando em os estudantes prprias so capazes e de explicar em
adequadamente,
suas
palavras,
baseando-se
experincias pessoais, os conceitos biolgicos. 4. Multidimensional quando os estudantes aplicam o conhecimento e as habilidades adquiridas, relacionando-os a conhecimentos de outras reas, para resolver problemas reais.
Espera-se que, ao concluir o ensino mdio, o aluno esteja alfabetizado e, portanto, alm de compreender os conceitos bsicos da disciplina, seja
capaz de pensar independentemente, adquirir e avaliar informaes, aplicando seus conhecimentos na vida diria.
Biologia no ensino fundamental Nas primeiras quatro sries do ensino fundamental, cada classe tem um professor responsvel por todas as reas de conhecimento. Nas quatro ltimas sries, Biologia faz parte da disciplina Cincias, que engloba tambm tpicos de Fsica e Qumica. A anlise das propostas curriculares das vrias unidades federadas indica que apenas um perodo de 12% a 15% do tempo de escolaridade dedicado ao aprendizado de cincias, com uma mdia de trs aulas por semana. Embora cada unidade federada tenha liberdade para a elaborao do currculo, o padro mais comum dos tpicos selecionados no Braslia e durante as quatro primeiras sries o seguinte: Ser humano; Sistemas do corpo humano; rgos dos sentidos; Necessidades vitais; Alimentao fontes de alimento; Seres vivos; classificao - animais e vegetais; Relao entre os seres vivos; Equilbrio ecolgico; Ser humano e ambiente; Modificaes fsicas e biolgicas dos ser humano.
Da 5 8 srie do ensino fundamental, os temas comumente ensinados so os seguintes: Plantas solo e clima agricultura; Distribuio de animais e plantas; Organismos e reaes qumicas; Nutrio, respirao, excreo;
Sistema nervoso - hormnios comportamentos; Produo de alimentos; Vida e energia fotossntese e cadeias alimentares ecossistemas; Reproduo e estrutura celular
Em geral, no se nota preocupao com aspectos importantes, como as relaes que dinamizam o conhecimento, os mtodos e os valores das cincias biolgicas. So apresentados e cobrados conhecimentos factuais, muitas vezes irrelevantes e desconexos em relao outras reas da disciplina cincias e s demais disciplinas do currculo.
Biologia no Ensino Mdio
Valor educativo ou formativo, relacionado com o desenvolvimento do educando; valor cultural, consistindo na contribuio para os grupos sociais; valor prtico, referindo-se aplicao de conhecimentos e objetivos utilitrios. A estruturao do programa de histria natural, ento em vigor nas escolas brasileiras, refletia claramente a grande influncia exercida pelo ensino europeu, tanto por meio de livros que aqui eram usados como pelos professores estrangeiros que vieram trabalhar nas escolas superiores brasileiras, notadamente na Faculdade de Filosofia, Cincias e Letras da Universidade de So Paulo. A tendncia de ensino nesse perodo era de tratar os assuntos considerando os vrios grupos de organismos separadamente e suas relaes filogenticas. As aulas prticas tinham como meta principal ilustrar as aulas tericas. Na dcada de 1960 a situao de modificou por ao de trs grupos de fatores: o progresso da Biologia, a constatao internacional e nacional da importncia do ensino de cincias como fator de desenvolvimento, e Lei de Diretrizes e bases da Educao Nacional de 20 de dezembro de 1961, que
descentralizou as decises curriculares, at ento de responsabilidade da administrao federal. A exploso do conhecimento biolgico provocou uma transformao na tradicional diviso, botnica e zoologia, passando do estudo das diferenas para anlise de fenmenos comuns a todos os seres vivos. Essa anlise feita em todos os nveis de organizao, da molcula comunidade, teve como consequncia incluir nos currculos escolares a um novo e amplo espectro de assuntos, indo da ecologia e gentica de populaes ate a gentica molecular e bioqumica. Na dcada de 1970, o projeto nacional da ditadura militar que estava no poder era o de modernizar e desenvolver os pas. O ensino de Cincias era considerado importante componente para a preparao de um corpo qualificado de trabalhadores, conforme foi estipulado na Lei de Diretrizes e Bases da Educao, promulgada em 1971. Esse perodo foi caracterizado por uma srie de fatores contraditrios, pois, ao mesmo tempo em que o texto legal valorizava as disciplinas cientficas, elas eram profundamente prejudicadas pelo atravancamento do currculo por disciplinas que pretendiam ligar o aluno ao mundo do trabalho (como zootecnia, agricultura, tcnica de laboratrio), sem que os estudantes tivessem base para aproveit-las. A formao bsica foi danificada sem que houvesse um correspondente beneficio para a profissionalizao (Krasilchik, 1987). Nas salas de aula, a crise e a massificao do ensino provocavam um aviltamento das condies de trabalho do professor. A populao escolar passou tambm por profundas mudanas: de uma composio majoritrias de crianas e jovens, predestinados a frequentar cursos universitrios, ela passou a ser formada tambm por jovens trabalhadores, muitos deles alunos de cursos noturnos. Nesse perodo, vrios projetos nacionais de ensino, incluindo uma extensa variedade que ia desde livros para o mero repasse de informaes ate currculos oriundos de estreito relacionamento como a comunidade, foram preparados abrangendo uma ampla gama de concepes sobre o ensino de Biologia (Krasilchik).
Com relao ao incio da dcada de 1990, os programas determinantes de Biologia do Ensino Mdio da escola brasileira esto discriminados a abaixo:
1 srie: A origem da vida Caractersticas dos seres vivos Citologia estrutura Metabolismo Teoria celular histrico Histologia animal e vegetal.
2 srie. Taxionomia Critrios de classificao Morfisiologia animal Morfisiologia vegetal Embriologia Reproduo e desenvolvimento humano Gentica mendeliana Gentica de populaes Evoluo teorias, mecanismos.
3 srie. Ecologia Populaes Relaes ecolgicas Regies ecolgicas O homem e o meio ambiente
Verifica-se assim a manuteno da tendncia descritiva, j identificada em pesquisas anteriores. Isso evidenciado quando se nota que o assunto estrutura celular predomina, aparecendo em 96%, e o estudo do metabolismo celular, em cerca de 70% das propostas curriculares. A diversidade dos seres vivos aparece nas descries Morfisiolgicas dos vrios grupos, sem que se estabeleam relaes entre eles. A anlise diversificada e as tentativas de construir sistemas abrangentes, que so bases essenciais para a compreenso do procedimento cientifico, aparecem raramente. A forma de apresentao dos grupos muito variada. Em alguns casos, trata-se das funes, considerando-se os processos comuns a animais e vegetais. Em outros, mantida a diviso tradicional entre botnica e zoologia, descrevendo-se mais ou menos detalhadamente cada grupo. O conceito de Biologia como cincia s aparece em 50% dos programas. A relao entre cincia, tecnologia e sociedades aparece ainda menos, indicando a falta de anlise das implicaes sociais do desenvolvimento cientifico e tecnolgico, to presente, por sua importncia, nos currculos das disciplinas cientficas. No final dos anos 1990, o Ministrio da Educao produziu e difundiu Parmetros Curriculares Nacionais para o Ensino Fundamental, considerando que o papel das Cincias Naturais o colaborador para a compreenso do mundo e suas transformaes, situando o homem como individuo participativo e parte integrante do Universo (PCN, 2001, p. 15). A divulgao desse documento evidencia um esforo reiterado de implantar um currculo nacional, embora haja ressalvas informando que no um modelo curricular homogneo e impositivo. No mbito de cincias em geral, e Biologia, em particular, pretende-se enfatizar, nos temas comumente includos (ambiente, ser humano e sade), aspectos prticos e do cotidiano dos alunos. Alguns desse tpicos so retomados nos chamados temas transversais que, segundo os PCN, as
disciplinas convencionais no suprem totalmente no sentido de discutir questes sociais e valores para o pleno exerccio da cidadania. Os temas transversais escolhidos foram: tica, pluralidade cultural, meio ambiente, sade, orientao sexual, muitos dos quais so normalmente discutidos pelos professores de cincias. No ensino mdio, os PCN tm o duplo papel de difundir os princpios da reforma curricular e orientar o professor na busca de novas abordagens e metodologias (PCN, 1999, p 13). Os tpicos de Biologia so includos em conjunto denominados rea das Cincias da Natureza, matemtica e suas tecnologias e pretende -se desenvolver competncias e habilidades resumidas na tabela seguinte:
Representao e comunicao: Descrever processos caractersticas do ambiente ou de seres vivos,
observados em microscpios ou a olho nu. Perceber e utilizar os cdigos intrnsecos da Biologia. Apresentar suposies e hipteses acerca dos fenmenos biolgicos
em estudo. Apresentar de forma organizada, o conhecimento biolgico
aprendido, atravs de textos, desenhos, esquemas grficos, tabelas, maquetes, etc. Conhecer diferentes formas de obter informaes (observao,
experimento, leitura de texto e imagem, entrevista), selecionando aquelas pertinentes aos temas de estudo. Expressar dvidas, ideias e concluses acerca dos fenmenos
biolgicos.
Investigao e compreenso
Relacionar fenmenos, fatos, processo e ideias em Biologia, elaborando identificando regularidades e diferenas, construindo
conceitos,
generalizaes. Utilizar critrios cientficos para realizar classificaes de animais,
vegetais etc. Relacionar os diversos contedos conceituais de Biologia (lgica interna)
na compreenso de fenmenos. Estabelecer relaes entre parte e todo de um fenmeno ou processo
biolgico. Selecionar e utilizar metodologias cientficas adequadas para a
resoluo de problemas, fazendo uso, quando for o caso, de tratamento estatstico na anlise de dados coletados. Formular questes, diagnsticos e propor solues para problemas
apresentados, utilizando elementos da Biologia. Utilizar noes e conceitos da Biologia em novas situaes de
aprendizado. Relacionar o conhecimento das diversas disciplinas para o entendimento
de fatos ou processos biolgicos.
Contextualizao sociocultural Reconhecer a Biologia como um fazer humano e, portanto, histrico,
fruto da conjuno de fatores sociais, polticos, econmicos, culturais, religiosos e tecnolgicos. Identificar a interferncia de aspectos msticos e culturais nos
conhecimentos de senso comum relacionado a aspectos biolgicos. Reconhecer o ser humano como agente e paciente de transformaes
intencionais por ele produzidas no sue ambiente. Julgar aes de interveno, identificando aquelas que visam
preservao e a implementao da sade individual, coletiva e do ambiente. Identificar as relaes entre o conhecimento cientifico e o
desenvolvimento tecnolgico, considerando a preservao da vida, as condies de vida e as concepes de desenvolvimento sustentvel.
Na dcada de 1990, documentos oficiais dividem o que se pretende dos alunos em duas categorias difceis de seres definidas e diferenciadas: competncias e habilidades. Embora no haja um sentido consensual para as expresses, considera-se que competncias so, de forma geral, aes da inteligncia, as quais usamos para estabelecer ralaes entre objetos, situaes, fenmenos e pessoas. As habilidades so decorrentes das competncias adquiridas, e confluem par o saber fazer. Essas habilidades se aperfeioam e se articulam por meio das aes desenvolvidas, possibilitando nova reorganizao das competncias. O impacto dos PCN foi relevante no currculo terico laborado por entidades oficiais e autores de livros, entre outros. O reflexo em sala de aula ainda est por se avaliado, embora dados preliminares indiquem os professores criticam tanto a tentativa da homogeneizao, como o seu distanciamento das discusses da elaborao do material de um projeto amplo que envolva todas as aes da escola. Vimos que a Biologia, como apresentada nas escolas de ensino mdio e fundamental, ainda hoje, reflete o momento histrico do grande
desenvolvimento cientifico das dcadas de 1950 e 1960, da esperana depositada na cincia para a soluo dos problemas da humanidade e, paradoxalmente, dos problemas decorrentes do uso da cincia e da tecnologia. No entanto, a partir dos anos de 1970, eclodiram enormes problemas sociais demonstrando que, pelo menos em parte, essas esperanas eram infundadas e produziram mudanas de documentos legais, embora o ensino nas classes pouco tenha mudado. Muitos educadores, diante desse novo quadro, admitem que a Biologia, alm das funes que j desempenha no currculo escolar, deve passar a ter outra, preparando os jovens para enfrentar e resolver problemas, alguns dos quais com ntidos componentes biolgicos, como o aumento da produtividade agrcola, a preservao do ambiente, a violncia, etc. de acordo com essa concepo, os objetivos do ensino de Biologia so: aprender conceitos bsicos, analisar o processo de investigao cientifica e
analisar as implicaes sociais da cincia e da tecnologia para tanto, vrias dimenses devem ser consideradas no tratamento dos vrios tpicos: Ambiental motivando o aluno a analisar o impacto da atividade
humana no meio ambiente e a buscar solues para os problemas decorrentes; Filosfica, cultural e histrica - levando o estudante a compreender o
papel da cincia na evoluo da humanidade e sua relao com religio, a economia, a tecnologia, entre outras.
As vises de mundo relacionadas ao papel da espcie humana no conjunto da biodiversidade e o conceito de evoluo resultaram em modificaes fundamentais na forma de pensar da humanidade e no podem ser desconhecidas dos alunos: Mdica preveno e cura de doenas e a compreenso de conceitos biolgicos bsicos esto estreitamente relacionados. Recentes descobertas em medicina e em gentica humana so excelentes possibilidades de vincular aspectos cientficos vida doas alunos. tica a presena crescente de problemas individuais e sociais e a sua
divulgao pelos meios de comunicao de massa tornam essencial que os alunos possam justificar e defender posies em relao a tpicos como aborto, eutansia, biodiversidade e relaes internacionais, propriedade das descobertas, por exemplo.
Cap. 2 O Aprendizado de Biologia
Comportamentalismo Nas dcadas de 1950 a 1970, as teorias baseadas no condicionamento tiveram grande impacto na educao, principalmente por intermdio da
ideias do condicionamento operante. Essas ideias admitiam que as consequncias agradveis de um evento funcionassem como reforadoras e as desagradveis como aversivas. O comportamento humano seria modelado por procedimentos de controle, recompensa e punio, e revelado por conhecimentos, atitudes e habilidades observveis e mensurveis. O professor planejava suas atividades de forma a obter o controle de aprendizado dos alunos, modificando, eliminando ou introduzindo
comportamentos. Na dcada em que as ideias comportamentalistas de aprendizagem tiveram preponderncia, desenvolveu-se bastante a tecnologia educacional apoiada nas maquinas de ensino foi a nfase na necessidade de estabelecer objetivos educacionais. A tarefa de preparao de objetivos educacionais dos cursos, obrigatria durante a fase de planejamento em certas escolas, foi reduzida a um formalismo sem nenhum resultado efetivo, demonstrando que o prprio sistema desenvolveu um processo homeosttico de ajustamento diante de uma exigncia exagerada e sem significado.
Cognitivismo
Este termo engloba teorias que do nfase aos processos mentais dificilmente observveis, enfatizando a possibilidade de o aluno adquirir e organizar informaes. O conhecimento seria o produto da interao do homem como o seu mundo. Como influncias cognitivas preponderantes no ensino de cincias devem ser mencionados o psiclogo norte-americano Jerome Bruner (1963) e o bilogo que formou escola na Sua, Jean Piaget, devido ao interesse de ambos para que os alunos lidassem com objetos e realizassem experincias para melhor aprender.
A influncia de Bruner resultou do seu trabalho com os autores dos projetos circulares da dcada de 1960, defendendo as ideias de que o estudante um solucionador de problemas e aprende pela descoberta. Foi um dos que valorizaram o desenvolvimento de habilidades intelectuais, colocando o aluno face s chamadas situaes discrepantes, que provocariam a necessidade de soluo. Sua teoria enfatiza o papel da curiosidade e a possibilidade de aceitar vrias formas de resolver as questes propostas. Ao analisar essas solues, o professor deve dar informaes aos estudantes sobre sua apreciao das mesmas, como fonte de motivao. Considerou ainda que cada tpico deve ser estruturado de forma a ser entendido pelos alunos. Admite que toda ideia ou campo de conhecimento pode ser apresentado para que qualquer estudante tenha a possibilidade de entender. A seguir, elaborou a concepo de currculo em espiral, em que as mesmas ideias reapaream com complexidade crescente nos vrios nveis de ensino. importante adquirir e desenvolver certas habilidades de elementos estruturados. Assim, a criana primeiro conhece o mundo pela ao, depois por imagens e representaes simblicas, principalmente pela linguagem. No Brasil a obra de Bruner teve importncia relativa. J os trabalhos de Piaget, tanto aqui, como nos EUA foram muito mais influentes. L os interessados em projetos de ensino de cincias promoveram nos anos 1960 conferencias denominadas Piaget Rediscovered, introduzindo as ideias desse importante cientista no meio educacional americano. Tais ideias so to importantes e significativas que at hoje fundamentam muitas discusses dobre o ensino de cincias. Alm das ideias sobre a psicognese dos conceitos, Piaget revolucionou a metodologia da pesquisa usando, em lugar de grandes amostras, entrevistas e testes com alunos observados individualmente, modalidade do chamado mtodo clnico. Considera tambm o aprendizado um processo ativo em que as estruturas cognitivas resultam da interao dinmica entre o organismo e o ambiente por processos de assimilao, equilibrao e auto-regulao.
Admite ainda que fatores que influenciam no desenvolvimento de estruturas mentais e na maturao do ser humano so as relaes como o ambiente e com o meio social. Por meio de entrevistas e para verificar em que fase se encontrava em relao ao conceito de conservao, foram preparadas provas em que os alunos deveriam realizar certas tarefas como, por exemplo: o professor apresenta duas esferas de massa de modelar e, em seguida, faz, com que uma delas, um cilindro longo e pede ao estudante que identifique aquela que tem mais massa.
Piaget identificou quatro estgios de desenvolvimento: 1. Sensrio-motor desde o nascimento at cerca de 2 anos. Perodo
no-verbal caracterizado por atos reflexos, explorao como os sentidos, imitao e permanncia dos objetos, o que significa que a criana esquecese do objeto este removido de sua presena.
2.
Pr-operatrio dos 3 anos at cerca de 7 ou 8 anos - A criana
egocntrica e nessa fase ocorre o desenvolvimento da linguagem. Nessa fase ela j capaz de representar mentalmente e simbolicamente objetos, mesmo na sua ausncia, e classific-lo com base em uma caracterstica. Desenvolve tambm a capacidade de lidar como novos objetos e eventos.
3.
Concreto-operacional idade 7 a12 anos - Nesta etapa os alunos
ordenam, classificam, estabelecem relaes causais e espaos-temporais. Realizam operaes lgicas com objetos concretos; portanto, a
manipulao muito importante, tendo consequncias claras para o ensino de Cincias. Tambm a capacidade de classificar um pr-requisito para o desenvolvimento de conceitos como plantas, folha, raiz, fruto, etc.
4.
Operatrio-formal crianas acima de 14 anos - Os estudantes
so capazes se analisar desde situaes concretas at hipotticas. Desenvolve o raciocnio proporcional, probabilstico, correlacional e controle de variveis.
Nas atividades, a interao social entre alunos um fator importante para provocar discusses e dvidas que ajudem a resolver problemas e a esclarecer questes.
Sociocultural
Uma importante contribuio s concepo de aprendizagem foi a do pesquisador russo Vygotsky, que enfatizou a importncia sociocultural no processo de aprendizagem da criana. Seus estudos foram fundamentados na observao das crianas no seu dia-a-dia, incluindo a escola e a famlia. Segundo Vygotsky, a relao com os mais velhos uma poderosa fora no desenvolvimento mental da criana. Por meio de dilogos entre pessoas e, mais enfaticamente, graas ao papel do professor, os jovens passam a conhecer o mundo simblico. Assim, no ensino de cincias p importante no s a contato com os objetos, mas tambm com os esquemas conceituais vigentes, que lhe so apresentados pelo representante dessa cincia que com ele interage: o professor.
As ideias bsicas de Vygotsky so: O objetivo maior da escola desenvolver a personalidade dos alunos; A personalidade humana esta vinculada a seu potencial criativo; O processo d ensino aprendizagem a atividade pessoal do aluno; O professor dirige e guia essa atividade individual;
Os mtodos de ensino e de aprendizagem correspondem s suas
particularidades individuais e, portanto, os mtodos no podem ser uniformes.
Construtivismo A partir do trabalho e da metodologia dos cognitivistas, desenvolveram-se outras linhas de pesquisas e teorias para explicar como os alunos adquirem, interpretam e usam informaes construindo o conhecimento. Essa a ideia bsica da vertente de anlise do aprendizado denominada construtivismo, que admite ser o conhecimento edifica do pela prpria pessoa, e, portanto no transmitido nem revelado. Admite-se que todo aluno possui um acervo de conhecimento e que muitas palavras diferentes so usadas para identificar e interpretar esse acervo: ideias, crenas, significados, concepes, estruturas qualificadas como erradas, alternativas, pessoais, intuitivas, ingnuas, espontneas, infantis. Nessas expresses, com frequncia pressupe que os alunos j trazem para a aula ideias prprias, mas inadequadas, que influem
fundamentalmente e criam dificuldades no s no aprendizado do contedo, mas tambm no de habilidades como a de observao, to essencial em Biologia.
Existem seis aspectos importantes apontados por autores numa viso construtivista: Os resultados da aprendizagem dependem no s do ensino ministrado,
mas dos objetivos, da motivao e dos conhecimentos que o aluno traz para a escola. O aprendizado envolve a construo de significados para cada um dos
alunos. Uma vez construdos os significados, cada aluno faz sua avaliao e os
aceita ou rejeita.
A responsabilidade do aprendizado dos estudantes, ou seja, deles
depende a ateno dada tarefa de construir e avaliar determinadas ideias. possvel identificar regularidades nos significados construdos pelos
alunos.
Dados obtidos no Brasil, por meio de pesquisas, confirmam muitas dessas ideias. Admite-se tambm que o conhecimento de histria e filosofia um instrumento para os jovens compreenderem como as descobertas contriburam para o progresso da humanidade e da qualidade de vida. Os efeitos da cincia e da tecnologia na vida moderna oferecem vantagens e causam problemas, e uma anlise histrica permitira aos alunos compreenderem seu significado no momento atual. De outro lado, o progresso cientifico e tecnolgico nos ltimos anos vem levantando problemas ticos sociais que devem ser relacionados a vises de mundo que fundamentaram os processos de descobertas e guiaram a utilizao de seus resultados. Essa vertente justifica a introduo de historia a filosofia na preparao para a cidadania, auxiliando na tomada de decises, principalmente em relao institucionalizao e ao suporte da prpria cincia. O trabalho dos professores, tambm influenciado pela sua concepo de escola, ensino, aprendizado; e pelo conhecimento dos contedos que deve ensinar. Assim, uma postura construtivista implica em criar situaes e envolver os alunos em atividades que promovam o aprendizado de Biologia. Um vez instalada a dvida, o professor deve apresentar novas ideias que sejam mais aceitveis e expliquem um maior nmero de situaes, do que as preexistentes. Um primeiro passo para conseguir a mudana fazer com que os estudantes tenham dvidas sobre a correo e propriedade de seus conceitos, no porque uma autoridade impe, mas porque uma real dvida e insatisfao se instalem neles por meio do professor, pelo colegas ou por algum evento.
A tarefa dos docentes est muito relacionada a investigar como e por que o aluno aprende, deixando de limitar-se a uma apresentao de contedos, temas e atividades listadas em proposta curricular e expostas
discursivamente em livros de texto. Os corolrios pedaggicos da aceitao de diferentes objetivos para o ensino de Biologia ou da adeso a um modelo de cognio devem dar resultados muito diferentes. Quando decises sobre currculos no se baseiam em convices firmes bem fundamentadas, podem resultar em programas desconexos, de resultados duvidosos.
Cap. 3 Planejamento Curricular
Introduo O currculo um caminho a seguir, ou seja, feito por uma instituio que assume tambm a responsabilidade de colocar em prtica uma proposta educacional e avaliar seus resultados. De acordo com essa concepo, o currculo compreende inicialmente um plano, elaborado pelos responsveis por uma escola, uma declarao de intenes, que podemos chamar de currculo terico. Esse plano, ao ser realizado, sofre uma srie de alteraes em funo das contingncias de sua aplicao, de tal forma que a percepo que dele tem os professores e alunos se difere bastante uma da outra. Essas diferenas resultam tanto das experincias de
aprendizagem planejadas, que compe o currculo aparente, como de experincias de aprendizagem no planejadas ou no explicitadas, que compe o currculo latente. Esse currculo latente se revela, em ao , nem sempre com as intenes expressas no currculo terico. Assim, embora em grande parte dos currculos escolares a finalidade explicita do ensino de Biologia seja desenvolver capacidade de pensar lgica e criticamente, a realidade de um ensino diretivo, autoritrio, em que toda a iniciativa e oportunidade de discusso dos alunos coibida, indicar que se deseja na verdade apenas transmitir conhecimentos. Portanto o professor de
Biologia deve estar consciente de que entre suas responsabilidades esto includas tomar decises na fase de planejamento e agir de forma correspondente, para haver harmonia entre o que se diz ou escreve nas propostas curriculares e o que de fato se faz.
Objetivos Alguns problemas bsicos no planejamento curricular so a escolha de objetivos e sua formao, para que sejam plenamente compreendidos por aqueles que tm responsabilidade por sua execuo e avaliao. No entanto mais do que forma, algumas perguntas so essenciais para a escolha dos objetivos, como por exemplo, (Vandervoort, 1988):
Por que ensinar Biologia? Qual a participao da Biologia no currculo escolar? Que tipo de Biologia deve ser ensinada? Qual a importncia social do ensino de Biologia?
Muitas tentativas foram feitas pra classificar as vrias tendncias curriculares e identificar as foras que atuam nas decises tomadas pelos sistemas educacionais, por administradores e por professores. Os governos e as associaes de classe tm um papel importante no nvel de deciso mais geral da construo curricular. No nvel execuo, o tipo de escola, o estilo administrativo, a clientela formada pelos alunos e suas famlias, tambm exercem importante influncia nas escolhas e nas aes do professor que, finalmente, quem pe em prticas o que vai sendo proposto nas vrias instancias decisrias.
Desenvolvimento de progresso cognitivo
Outra tendncia valoriza o desenvolvimento intelectual do aluno, a capacidade de buscar informaes e de us-las em situaes novas. Os modelos pedaggicos so centrados na soluo individual de problemas ou em atividades de grupos interativos. O professor tem como responsabilidade criar situaes que auxiliem a aprendizagem, a qual transcorre de forma autnoma, respeitando-se as caractersticas individuais e os estilos prprios de cada um. Exige do docente um conhecimento amplo dos grandes conceitos da disciplina e tambm a capacidade de criar situaes que demandem uma atitude de investigao. Os adeptos dessa linha vem o ensino de Biologia como uma forma de desenvolver um repertrio de habilidades intelectuais, que ajudam o aluno a resolver problemas por meio da capacidade de buscar dado, analis-los, propor hipteses, organizar investigaes para test-las e avaliar solues.
Sociorreconstrucionista Para alguns ainda, o objetivo da escola e, portanto do ensino de Biologia alcanar mudanas da sociedade, a fim de que esta possa melhor atender aos interesses individuais e coletivos.
Contedo O contedo, na realidade, a preocupao mais presente entre os professores ao fazerem planejamento curricular, tendo que tomar decises de trs tipos: o que ensinar, em que sequncia e como relacionar e integrar os assuntos aos outros tpicos da mesma disciplina e das outras disciplinas.
Abrangncia do contedo A delimitao do escopo do seu currculo , para o professor de Biologia, um problema cada vez mais srio por vrias razes: o conhecimento esta se expandindo enormemente e, em consequncia, as prprias cincias biolgicas vem sofrendo transformaes em sua organizao. De uma cincia que se concentrava na descrio e nos conhecimentos qualitativos, como o desenvolvimento na bioqumica e na biofsica, de processos experimentais e de mensurao, bem como da anlise estatstica, a Biologia passou a se um campo de conhecimento com leis gerais, o que alargou e aprofundou suas dimenses, tornando muito difcil ao professor decidir o que deve ser fundamental, portanto includos em seu curso e o que deve ser acessrio, podendo consequentemente ser deixado de lado. Essas mudanas, que ocorreram no cenrio educacional internacional e brasileiro, puderam ser constatadas consultando-se, periodicamente, professores de Biologia sobre a importncia que atribuem aos diferentes tpicos do currculo. Considera-se, hoje, fundamental a compreenso de conceitos como o de biosfera uma camada de vida entrelaada numa teia que envolve a Terra. Esse conjunto extremamente variado compe a biodiversidade dos seres vivos, que chegou ao estado atual por uma gradual mudana denominada evoluo. O papel do homem, nesse conjunto, exemplifica bem o processo evolutivo morfofisiolgico, agregando porem um componente evoluo da cultura. Essa cultura permitiu ao homem viver e ocupar diferentes habitat, expandir sua populao e afetar o ambiente de forma a exigir cuidados para nos comprometer a sobrevivncia de outras espcies e sua prpria. Os livros didticos, em geral, tambm continuam apresentando as subdivises clssicas e os professores acabam se acomodando a um tipo de programa por ser mais familiar, embora no seja por eles menos considerado o melhor.
Sequncia Uma vez que tenha decidido o que ensinar, o professor enfrenta outro problema: como ordenar os vrios subtpicos? A organizao pode ir dos assuntos mais simples aos mais complexos; dos aspectos prticos aos tericos, dos conhecimentos fundamentais aos perifricos. Em discusses sobre o assunto frequente que a resposta a essa pergunta seja: na ordem lgica. Na verdade, h muitas ordens lgicas. A demonstrao mais conhecida desse fato foi a deciso do grupo de autores do Biological Science Curriculum Study (BSCS) quando, admitindo que um currculo deveria ter flexibilidade para adaptar-se a diferentes situaes, atender diversidade de gosto dos professores e, ao mesmo tempo, cobrir tpicos importantes, resolveu elaborar uma srie de mdulos que cada um organizaria da forma que melhor lhe conviesse. No entanto, consultados os professores e as editoras, o grupo concluiu que essa soluo no era muito prtica. Para chegar a um consenso que resguardasse a necessidade de atender variabilidade das situaes e da semelhana dos tpicos fundamentais, a comisso responsvel pela escolha do contedo doa livros do BSCS decidiu elaborar trs verses diferentes que, embora tivessem caractersticas diferentes. Essas verses ficaram conhecidas como Verso Azul, Verso Verde e Verso Amarela. Cada um desses projetos reflete uma forma de tratamento dos mesmos tpicos, porem diferentemente ordenados. Na verso azul, a sequncia se inicia com as unidades menores de organizao, como tomos e partculas, seguindo para nveis mais complexos, como o nvel de clula, de tecido. De organismo, de comunidade at ecossistema. A verso verde usa um critrio de organizao, que vai das inter-relaes entre os organismos nos ecossistemas,
comunidades e populaes at as unidades bsicas de organizao, como clulas, partculas e molculas. A verso amarela, que tem a organizao mais tradicional, por sua vez, parte de um nvel intermedirio: a clula. Analisa seus constituintes e funes e suas formas de organizao em tecidos, rgos e indivduos. Outra variao do mesmo assunto seria partir de problemas referentes aplicao da Biologia vida diria, usando tpicos de higiene,
microbiologia, gentica e agricultura, e da, seguir para a anlise dos fenmenos no vrios nveis de organizao: mutabilidade, populao, individuo, rgo, tecido, clula etc. Ainda outro caminho, j explorado por vrios autores, seria o estudo de casos grandes descobertas cientficas para analisar os fenmenos biolgicos. Vemos assim, que h possibilidades de um grande nmero de arranjos de um mesmo contedo para a apresentao a alunos de qualquer nvel de ensino. Numa tentativa de sintetizar os vrios princpios que regem a sequncia do contedo, Possner e Strike (1979) elaboraram um esquema de classificao. Simplificadamente, as categorias definidas por eles so:
a. Sequncias relacionadas ao mundo: so aquelas em que a organizao do contedo e as relaes entre os fenmenos so ordenadas como ocorrem realmente. Por exemplo, os elementos so ordenados de acordo com seu arranjo fsico ou espacial. Inclui-se nesse caso a anlise dos componentes celulares na ordem: membrana, citoplasma e ncleo; dos rgos da planta na ordem: raiz, caule e folha. Ou ento quando se apresenta em ordem cronolgica o aparecimento das vrias teorias de evoluo: lamarckismo, darwinismo e neodarwinismo; b. Sequncias relacionadas ao conceito: refletem a organizao do conhecimento, so exemplos os currculos que se baseiam na estrutura da disciplina e tem como tema central seus conceitos fundamentais. c. Sequncias relacionadas investigao: esta categoria inclui dois aspectos sobre desenvolvimento da pesquisa, baseando-se em descries da maneira como os cientistas realmente trabalharam, em que contexto operam, levando em considerao os fatores sociais e psicolgicos, entre outros, que influem no processo. d. Sequncias baseadas no conhecimento sobre a psicologia da aprendizagem: incluem-se, nesta categoria, as propostas de currculo que tomam como base teorias da aprendizagem para organizar o contedo, usando, por exemplo, os estgios do desenvolvimento cognitivo, com base nos estudos da escola piagetiana; muitos currculos de cincias foram
elaborados na sequncia em que os conceitos possivelmente podem ser aprendidos pelos alunos. e. Sequncia baseada na utilizao do conhecimento: a organizao dos contedos feita de acordo com a necessidade futura dos alunos, prevista em termos pessoais e profissionais. Por exemplo, apresentar elementos de estatstica como introduo gentica, apresentar vidraria e ensinar a usar a balana no incio dos cursos de Qumica.
Relao da Biologia com as outras disciplinas
A integrao de contedos depende da natureza da prpria disciplina, das caractersticas dos alunos e das condies em que o processo ensinoaprendizagem deve transcorrer. Pode referir-se s relaes entre vrios elementos de uma mesma disciplina, integrao intradisciplinar, ou entre vrias disciplinas que so presenteadas simultaneamente ou, em sequncia, integrao interdisciplinar. As relaes entre Biologia, fsica e qumica, tanto como parte da disciplina cincias no ensino fundamental como na categoria de disciplinas independentes na escola de ensino mdio, tem sido tradicionalmente objeto de reflexo e discusso na organizao dos currculos das escolas de ensino mdio e fundamental. Mais recentemente, com a relao cada vez mais estreita entre as cincias biolgica e as cincias humanas. A integrao das vrias disciplinas exige a ruptura dessas barreiras pelo trabalho comum de vrios professores e a unio dos laboratrios e bibliotecas para o desenvolvimento de temas gerais. Esses temas podem representar conceitos centrais como o de energia, ou problemas prticos como a poluio e exigem a participao de um grupo de docentes que compartilhem recursos, instalaes e responsabilidades.
Cap. 4 A comunicao entre professor e aluno
Introduo
Professores e estudantes transmitem mensagens por via oral, ou por meio de textos ou figuras, e os educadores esto cada vez mais conscientes das dificuldades prprias desse vrios tipos de comunicao. Muitas so as possveis explicaes para essas dificuldades: as diferenas entre professores e alunos so obstculos para a compreenso de cdigos e valores peculiares aos dois grupos: os meios de comunicao de massa limitam a capacidade de expresso dos jovens.
Comunicao oral Provavelmente, todos os obstculos citados acabam influindo de alguma forma no resultado final: os alunos tm problemas para compreender e comunicar ideias. Nas aulas de Biologia do ensino mdio, foram arrolados alguns problemas especficos relativos comunicao ora: Incompreenso do vocabulrio: os alunos no acompanham as aulas porque so usadas palavras desconhecidas, ou porque eles atribussem aos termos e significados diferentes dos atribudos pelo professor. Excesso de vocabulrio tcnico: o excesso de vocabulrio tcnico que o professor usa em suas aulas leva muitos aluno a pensar que Biologia s um conjunto de plantas, animais, rgos, tecidos e substancias que devem ser memorizadas. O uso inadequado da terminologia cientfica esta estreitamente ligado ao processo de formao de conceitos. Schaefer (1979) admite que um conceito seja formado por trs partes:
1. O ncleo lgico, que a estrutura constante, representativa de uma classe de objetos ou eventos. 2. Associado ao ncleo lgico h um nome que serve tanto como veculo para comunicao entre os indivduos como de referncia para efeito de memorizao pelo prprio individuo. 3. Envolvendo o ncleo lgico e o nome associado h uma rede compacta de relaes que formam a moldura de associaes. A palavra s passa a ter significado quando o aluno tem exemplos e oportunidades suficientes para us-las, construindo sua prpria moldura de associaes. Como, s vezes, os termo apresentados so desnecessrios, j que nuca mais voltaro a ser usados, o professor deve tomar cuidado para na sobrecarregar a memria dos alunos com informaes inteis.
Falta de interao professor-aluno. O ensino informativo, centrado no professor, representado pela aula expositiva, pode ser transformado pela introduo de discusses na sala de aula, chamadas de exposies dialogadas. As perguntas intercaladas na exposio motivam os aluno, servem para controlar e ganhar sua ateno, auxiliam no raciocnio e expem os aluno e muitas ideias em lugar de limitlo a ouvir apenas o professor. No entanto, os professores no usam o recurso do questionamento em classe porque temem que, de alguma forma, sua autoridade seja abalada e haja perda de segurana e do poder assegurados pelas aulas expositivas. Quando resolvem incentivar a discusso, tem que tomar certos cuidados para garantir o pleno xito da atividade, que depende essencialmente de uma genuna disposio para promover um debate franco e aberto nas aulas. Portanto, a primeira preocupao deve ser a de criar um clima de liberdade na aula, em que os professores ouvem e levam em considerao diferentes pensamentos e opinies. Aps os alunos verificarem que a participao bem vinda, certos comportamentos que podem ser
coercitivos no incio do curso, quando o professor no bem conhecido, passam a no ter mais esse efeito porque todos j se conhecem. bom lembrar que a interao do professor com o aluno ocorre tambm por meio de gestos, e que possvel demonstrar desagrado ou ironia por um olhar, um movimento ou uma expresso facial. Interrupes das respostas dos alunos tambm produzem uma diminuio da participao.
Informao visual Uma parcela muito importante das informaes em Biologia obtida por meio da observao direta dos organismos ou fenmeno ou por meio de observao de figuras, modelos, etc. Ouvir falar sobre um organismo , em geral, muito menos interessante do que ver diretamente a realidade, o que justifica aulas prticas, excurses e demonstraes. Ao usar ilustraes, os professores nem sempre consideram as dificuldades para a compreenso de representaes simblicas, o que requer dos alunos treino especial. Nas aulas de Biologia, alguns problemas especficos nesse campo foram identificados: Os alunos tm dificuldades de imaginar, a partir de figuras
representadas no plano, uma estrutura em trs dimenses. Fica difcil para os jovens vendo, por exemplo, esquemas de clulas ou de fases de desenvolvimento embrionrio em corte ou microfotografias eletrnicas. Para avaliar ou desenvolver o aprendizado, interessante que os alunos faam tambm o exerccio inverso: a partir da observao, reconstruir a estrutura em trs dimenses, desenhando ou construindo um modelo. Relacionar a representao simblica esquemtica realidade
requer treinamento. medida que vo se familiarizando, nas aulas com os smbolos adotados, passam a usar conveno apresentada nos livros e nas aulas.
da
Os estudantes tm dificuldades para usar a representao simblica qumica: frequentemente, nas aulas de Biologia, so feitas
representaes de equaes, modelos autnomos e diagramas moleculares antes que os alunos tenham tido a chance de aprender o que significam nas salas de aula. Verificar se os estudantes dominam esses pr-requisitos essencial no planejamento dos cursos. Grficos: impossvel, hoje, que o aluno faa um bom curso de
citologia, fisiologia ou ecologia sem saber construir grficos, interpret-los, interpolar e extrapolar dados. Como frequentemente chegam aos cursos do ensino mdio, sem que tenham aprendido a faz-lo, necessrio investigar qual o seu conhecimento do assunto no inicio, pois caso no tenham aprendido a trabalhar com grficos, o assunto devera ser includo na propagao. Escalas: quando o estudante comea a fazer observaes ao
microscpio, dificilmente relaciona o que v ao tamanho real do objeto e compreende as diferenas aumento nos vrios campos de observao. A essa dificuldade acrescenta-se a de relacionar o objeto real escala de representao e, alm desse treinamento durante o aprendizado, essencial usar escala ou objeto em tamanho real para que o observador tenha a noo do grau de aumento na representao.
Recursos audiovisuais Embora seja amplamente reconhecido o potencial dos recursos
audiovisuais no ensino de Biologia, os dados disponveis indicam que so pouco e mal usados. De acordo com relatos, nem mesmo no tradicional quadro de giz so feitos esquemas, desenhos, demonstraes, de forma que a expresso pejorativa aulas de saliva e giz atualmente est mais reduzida, com a eliminao do giz, ficando a aula restrita apenas fala do professor ou leitura do livro didtico. O quadro de giz, um recurso inestimvel, cada vez menos usado e mais ineptamente usado. Em certas aulas, colocado, antecipadamente, na lousa todo o esquema da aula que vai ser seguido pelo professor. Os
alunos passam ento a copiar o que foi escrito, em lugar de acompanhar o assunto e, segundo os jovens, usam-no como referncia para contar o tempo que falta pra acabar a aula. Esse erro de fcil soluo, desde que os professores utilizem o quadro de giz para construir o roteiro da aula, passo a passo, para que o aluno possa ir acompanhando o raciocnio desenvolvido. Dessa forma, o recurso usado, dinamicamente, permitindo que o estudante tenha alguma participao, mesmo em aulas expositivas. Comunicao escrita A comunicao escrita entre professor e aluno hoje feita, basicamente, por um mediador: o livro didtico. So trs tipos mais comuns de livros didticos de Biologia: o texto do aluno, manual de laboratrio e o guia do professor. O texto do aluno deve apresentar de forma organizada, lgica e didtica, conhecimentos atualizados e relevantes, e o manual de laboratrios deve orientar os procedimentos e atitudes dos alunos nas aulas prticas. O guia do professor visa a auxiliar os docentes na utilizao dos materiais elaborados para os alunos, informando os usurios das intenes de seus autores. Livro didtico Livro didtico tradicionalmente tem tido, no ensino de Biologia, um papel de importncia, tanto na determinao do contedo dos cursos como na metodologia usada em sala de aula, sempre no sentido de valorizar um ensino informativo e terico.
Guia do professor A crescente importncia do manual do professor, que acompanha os textos didticos, est estritamente ligada s condies de trabalho e formao dos docentes, que ficam cada vez mais dependentes do livro como pea central de seus cursos. Muitos dos guias de professor, existentes no mercado, constam apenas do livro do aluno acrescido das respostas aos
exerccios, estrutura obviamente insatisfatria. Consulta feita pelos docentes revela tpicos que devem prioritariamente ser includos num guia do professor. Objetivos do curso Questes adicionais que no constem do livro-texto Lista de material para atividades Bibliografia complementar
Em seguida, na lista de prioridades, vm os seguintes quesitos: complementao terica; respostas complementadas s questes do livro do aluno, modelos de perguntas para a prova. Utilizao dos textos O livro deve ser base para a discusso em classe, e no apenas fonte de informao inerte. Para tanto, professor deve desenvolver a capacidade critica e avaliao a partir da leitura dos textos. Como os alunos dependem de nota em sala de aula, professor poder aproveitar-se desse fato para ensin-lo a compor resumos a compor resumos com os pontos principais de cada assunto. Computadores Cada vez mais os computadores fazem parte do nosso dia a dia. O seu uso de forma adequada nas escolas vem preocupando os educadores que buscam elaborar atividades que desenvolvam o raciocnio e facilitem o aprendizado. Com a intensificao do uso dos computadores torna-se necessrio evidenciar as possibilidades de influncia da cultura impressa na escola, e da cultura digital, que permite maior interao e participao do aluno na produo do conhecimento.
Cap. 5 Modalidades Didticas
Introduo A escolha da modalidade didtica, por sua vez, vai depender do contedo e dos objetivos selecionados, da classe a que se destina, do tempo e dos recursos disponveis, assim como dos valores e convices do professor. As modalidades didticas podem ser classificadas segundo vrios critrios. Segundo Ascher (1966), podem ser agrupadas de acordo com as atividades que os professores desenvolvem, tais como: falar aulas expositivas, discusses, debates: fazer simulaes, aulas prticas, jogos, projetos e mostrar demonstraes, filmes etc. Podemos ainda separar as diversas modalidades didticas de acordo com sua possibilidade de melhor servir aos objetivos do ensino de Biologia: Para transmisso de informaes: aulas expositivas, demonstrao Para realizar investigao: aula prtica, projetos; Para analisar as causas e implicaes do desenvolvimento da
Biologia: simulaes, trabalho dirigido.
Uma grande classificao agrupar diversas modalidades didticas e de acordo com o tamanho do grupo de alunos: Atividades para grandes grupos; aulas expositivas, demonstraes Atividades para pequenos grupos; seminrios, aulas Trabalhos individuais, projetos.
A prpria existncia de vrias classificaes indica que nenhuma delas totalmente satisfatria, principalmente porque difcil uma apreciao fora do contexto em que a aula se coloca. Aulas expositivas uma modalidade didtica mais comum no ensino de Biologia tem como funo informar os alunos. Em geral os professores repetem s livros
didticos, enquanto os alunos ficam passivamente ouvindo. Argumentos de ordem pedaggica podem ser invocados para justificas o uso de aulas expositivas em certos momentos de um curso: elas permitem ao professor transmitir suas ideias, enfatizando os aspectos que considera importantes, impregnando o ensino com o entusiasmo que tem pela matria. A passividade dois alunos representa uma das grandes desvantagens das aulas expositivas, pois gera uma srie de inconvenientes: a reteno de informaes pequena, porque h decrscimo de ateno dos alunos. Essa constatao cria a necessidade de se encontrarem formas de trabalho que permitam a manuteno de um alto nvel de ateno durante todo o perodo da aula. Outro problema vem da falta de interao entre professor e aluno. E os professores, pretendem dar mais contedo do que possvel no tempo disponvel, eles no estabelecem relaes casuais e as aulas podem ser mal preparadas.
Aulas prticas A principal funo das aulas prticas, reconhecidas na literatura sobre o ensino de cincias, : Despertar e manter o interesse dos alunos Envolver os estudantes em investigao cientifica Desenvolver a capacidade der resolver problemas Compreender conceitos bsicos Desenvolver habilidades
As seguintes funes para as aulas de laboratrio tambm as arroladas p outros grupos: Formular, elaborar mtodos para investigar e resolver problemas
individualmente ou em grupo.
Analisar cuidadosamente, com companheiros e professores, os
resultados e significados de pesquisas Distinguir observao de inferncia, comparar crenas pessoais,
compreenso cientifica e teorias.
As aulas de laboratrio tm um lugar insubstituvel nos cursos de Biologia, pois desempenham funes nicas: permitem que os alunos tenham contato direto com os fenmenos, manipulando materiais e observando organismos. O envolvimento do aluno depende da forma de propor o problema e das instrues e informaes fornecidas pelo professor aos estudantes. O mesmo assunto pode ser usado em um exerccio que apenas vise confirmao de uma teoria, ou usado como objeto de pesquisa. Excurses
A maioria dos professores de Biologia considera de extrema valia os trabalhos de campo e as excurses; no entanto, so raros os que as realizam. Os principais obstculos organizao das excurses so: complicao para obter a autorizao dos pais, da direo da escola que no querem ceder seu tempo de aula, o medo de possveis acidentes, a insegurana e o temor de no reconhecer os animais e plantas que forem encontrados, os problemas de transporte. Porm, as excurses tm uma importante dimenso cognitiva e tambm apresenta que Eisner (1979) chamou de resultados expressivos que, embora no possam ser previstos com antecedncia, so experincias estticas e de convivncia muito ricas e que variam de individuo para individuo. A organizao de uma excurso inclui:
Uma etapa de preparao em que feito o reconhecimento do local
escolhido para o trabalho e a identificao dos problemas que sero investigados Elaborao do roteiro de trabalho contendo as instrues para o
procedimento dos alunos e as perguntas que eles devem responder Trabalho de campo propriamente dito Trabalho em classe para a organizao dos dados e exame do material
coletado Discusso dos dados para a elaborao de uma descrio geral do sitio
visitado e uma sntese final
Projetos So atividades executadas por um aluno ou por uma equipe para resolver um problema e que resultam em relatrio, modelo, coleo de organismos, enfim, um produto final concreto. A funo do professor e orientar, auxiliar a resolver as dificuldades que forem surgindo. Portanto quando os alunos tm interesse por algum problema especfico, cabe ao professor verificar o andamento do projeto. De forma geral, so conhecidas as seguintes fases para o desenvolvimento de um projeto: Seleo do problema a ser investigado Elaborao do plano de trabalho Execuo do plano elaborado Muitos professores consideram parte essencial do projeto a obteno
de produto final. Em muitos casos um projeto pode ser bem sucedido em termos educacionais mesmo sem chegar e um resultado concreto como um relatrio, um modelo, uma coleo, etc. Quando o projeto fica pronto, o professor tem ainda uma tarefa difcil, a de julgar os seus resultados e avaliar o que mais importante o produto final ou o processo de execuo. Como tarefa tem por fim, mais do que a
aquisio de informaes, disciplina e aceitao devem ser levados em conta na avaliao.
Tecnologias educacionais Qualquer que seja a modalidade didtica que o professor escolha e use, ela pode se complementada, com vantagem, por recursos tecnolgicos como vdeo, retroprojetor, filmes, programas e ferramentas tecnolgicas, entre muitos outros. O computador, como j vimos, serve pra fornecer dados e permitir a participao do aluno na soluo de problemas. preciso que o professor auxilie o aluno a analisar e a decodificar as mensagens, tanto no material preparado para a escola como programas de radio, televiso e na leitura de jornais e revistas para o publico em geral.
Cap. 6 O ambiente
Introduo Em algumas escolas onde a concepo de ensino inclui atividades que aumentam a interao aluno-professor e aluno-aluno, os arranjos tradicionais cedem lugar a outro tipo de disposio: o local de trabalho do professor no ocupa posio dominante no conjunto formado por mesas e carteiras mveis, que podem ser combinadas de vrias formas, adequadas ao trabalho individual ou em grupo. Os estmulos a projetos individuais de pesquisa revelado pela existncia de locais para o trabalho e a manuteno de experimentos, que podem ser usados pelos estudantes fora do horrio de aula.
Portanto, como em qualquer outra etapa do planejamento curricular, o planejamento das instalaes para as aulas de Biologia deve levar em conta, prioritariamente, as atividades que ali sero realizadas, em funo dos outros determinantes especficos, como densidade dos alunos, as condies de luz, de gua, gs etc. E ainda a localizao do laboratrio no prdio da escola.
Laboratrio de Biologia A rigor, possvel dar um bom curso pratico, mesmo no dispondo de recursos especiais, quando no se tem mo espcimes de animais e plantas. No entanto, poder ser tanto mais eficiente quanto melhores forem as instalaes e o material disponveis, sendo um direito e um dever dos docentes pleitear a luta pela conquista de instrumentos que lhes permitam trabalhar melhor.
Organizao de trabalho no laboratrio Para as atividades prticas darem o melhor resultado possvel, o comportamento de estudantes e professores deve ser estruturado, para que possa ser aproveitado o Maximo de tempo possvel: Ao professor cabe: Preparo material com antecedncia Realizar o experimento antecipadamente, para verificar se seu
funcionamento previsto Dar aos alunos instrues de forma clara e precisa, sempre que
possvel, combinando uma explicao oral com a distribuio do material Cuidados com as substancias qumicas: Solues concentradas de acido e base s devem ser transportadas
por alunos com o professor. As substancias qumicas do laboratrio no devem ser colocadas na boca.
boca.
As solues devem ser pipetadas com bulbo de suco, nunca na
Frascos contendo substncias orgnicas volteis ou cidos no devem
ficar prximos chama ou expostos ao sol. No se deve adicionar gua a nenhuma soluo concentrada de
acido. Para preparar uma soluo, o acido deve ser acrescentado, lentamente, gua. Cuidados no trabalho com seres vivos: Animais e plantas no devem ser mantidos nos laboratrios, se no
puderem se adequadamente alimentados e limpos em fins de semana e nas frias. Verifique a legislao existente em seu Estado sobre as restries
relativas ao trabalho com animais.
Saindo da escola Os objetivos gerais do trabalho fora da escola so coletar dados e informaes, ver exemplos de princpios e fatos mencionados nas aulas, encontrar problemas para a investigao, desenvolver a percepo e aumentar a interao professor-aluno. Mais recentemente, com a preocupao sobre as implicaes sociais do conhecimento biolgico, houve uma expanso das atividades fora da escola: Desenvolver programas comunitrios Intensificar vnculos com a tecnologia Desenvolver responsabilidades de cidadania.
preciso dar aos alunos liberdade de observao e anlise daquilo que veem e considerar o tipo de bioexposio na hora da anlise e discusso da visita.
O guia deve orientar os alunos para resolver problemas. Por exemplo, quando os espcimes esto depostos de acordo com sua classificao, pode se pedir aos alunos que relacionem suas caractersticas e tipo de alimentao. Quando os espcimes so agrupados de acordo com o habitat que ocupa, os alunos podero relacionar aspectos de gentica, evoluo e ecologia, observando as adaptaes comuns.
Cap. 7 Avaliao
Introduo Para fazer um planejamento adequado da avaliao de um curso, importante lembrar que as provas ocupam um lugar central em todo o processo escolar, desempenhando inmeros e variados papeis: Servem para classificar os alunos em bons ou maus; para decidir se vo ou no passar Informam os alunos do que o professor realmente considera importante Informam o professor sobre o seu trabalho Informa a escola sobre os resultados do trabalho de alunos e professores Informam os pais Foram os alunos a estudar Criam problemas entre professores e alunos, pais e diretores (Krasilchik, 1985) Em decorrncia, surge uma srie de subprodutos dos quais alguns so: Tenso emocional Mudana de comportamento Estimulo competio Alterao de julgamento
A partir da dcada de 1970, estudos comparativos buscaram avaliar o aproveitamento dos estudantes em vrios pases. Os resultados desses trabalhos tiveram ampla repercusso no s na mdia como provocaram medidas visando mudar a situao em pases e regies que tiveram resultados considerados desfavorveis. Um desses estudos foi feito em 1970 pela International Association for the Evalluation of Educational Achievement (IEA), analisando o aproveitamento em cincia, inclusive Biologia, dos estudantes de 19 pases. Esse estudo foi repetido em meados de 1980, co a participao de 24 pases ou sistemas educacionais (IEA 1988). Para verificar o perfil dos alunos que saem do ensino mdio, foi institudo no Exame Nacional do Ensino Mdio (ENEM). Pretende-se, atravs do Enem, analisar: o domnio da lngua portuguesa, das reas de matemtica, artstica e cientifica; a aplicao de conceitos para a compreenso de fenmenos naturais, de processos histrico-geogrficos; a produo tecnolgica e as manifestaes artsticas (MEC Poltica e Resultados 1995-2002. Braslia, 2002). Planejamento da avaliao Vrios fatores que devem ser levados em conta no planejamento da avaliao de um curso ou unidade de estudo: Periodicidade das provas: essencial prever e comunicar ao alunos, no inicio dos trabalhos escolares, o nmero de provas a que sero submetidos e o intervalo entre elas. Tempo: quando o planejamento no inclui um perodo suficiente para a execuo dos trabalhos previstos para a avaliao, os resultados sero prejudicados. Instrumentos: sem dvida, esse um fator da maior importncia para que a avaliao fornea os dados que o professor busca seu trabalho e sobre seus estudantes.
Anlise das provas
No processo de avaliao, a fase de interpretao dos dados obtidos tem tanta importncia quanto as fase de escolha e elaborao dos instrumentos de avaliao.
Os requisitos bsicos de uma prova so: Validade do contedo, ou seja, o instrumento serve para obter um amostra que represente adequadamente os objetivos que o professor tem para seu curso. Validade de construo: verificar se as questes que constituem a prova esto bem elaboradas. Fidedignidade: ou seja, o instrumento de avaliao garante que diferentes observadores atribuam notas iguais a um mesmo trabalho, em diferentes correes. A escolha do tipo de questo que deve compor uma prova est estreitamente relacionada necessidade de fidedignidade. Quando os grupos so numerosos e a importncia das notas muito grande, preciso garantir que no haja variaes no critrio de correo das provas, ou pelo uso de provas objetivas, ou de perguntas de resposta livre cujos parmetros para a correo sejam bem estabelecidos.
Critrios para anlise dos resultados das provas A literatura revela hoje trs tendncias para a interpretao dos resultados de avaliao: O desempenho do aluno comparado a um parmetro estabelecido, como padro aceitvel. O desempenho do aluno comparado aos de outros estudantes da mesma classe ou mesma populao. O desempenho do aluno comparado ao resultado que ele mesmo obteve, em avaliaes anteriores.
Cap. 8 O Professor na sala de aula
Introduo Os estgios so uma forma de introduzir o licenciado na escola, com o auxilio de guias experientes que possam orient-lo e auxili-lo na soluo das dificuldades que venham a surgir. O estgio um canal de comunicao ligando as escolas de ensino superior s escolas de ensino mdio e fundamental, da levando informaes de suas necessidades universidade, que deve responder s escolas de nvel primrio e mdio com um influxo de novas ideias. A extrema importncia dos estgios indiscutvel, bem como as dificuldades para excuta-los de forma que atinjam plenamente suas funes nos programas de formao de professores. Uma dessas dificuldades reside nas relaes entre os dois grupos de instituies envolvidas.
Condies para realizar um estgio de qualidade H uma tendncia em considerar os estgio como forma de interferncia no trabalho dos professores, que se sentem constrangidos na presena dos universitrios que observam suas aulas. Na verdade, os estgios s podero servir plenamente a seus fins se os estagirios passarem a sentir-se elemento da escola e no estranhos nela. Os professores monitores, obviamente, tm papel preponderante na orientao dos licenciados, desde que possam trabalho de comum acordo com os professores de prtica de ensino e encarar sua atividade no como fonte de sobrecarga de trabalho, mas sim como uma oportunidade de instruir e ajudar jovens e colegas, com quem ser possvel trocar ideias e opinies sobre os cursos que esto ministrados e sobre problemas do ensino. Tipos de estgios
Estgio de observao: so aqueles em que o estagirio esta presente sem participar diretamente da aula. A condio de futuro professor, ele deve ver a escola de um ngulo muito diverso do que viu na condio de estudante, precisando de instrues e orientaes sobre o que buscar e focalizar. Geralmente acaba verificando que h muitos aspectos que merecem anlise e pesquisa mais detida. Estgios de participao: aquele em que o aluno auxilia o professor, sem contudo assumir a total responsabilidade pela aula. Geralmente nos estgios de participao, os estudantes ajudam o professor monitor nas aulas prticas. Estgios de regncia: aquele em que o estagirios tem a responsabilidade da conduo da aula. So vrias modalidades de estgio de regncia: execuo de atividades de regncia esparsas durante o curso regular, execuo de uma unidade durante o curso regular, minicursos, recuperao, atividades extraclasse e aulas para turmas-piloto.
Relao aluno-professor As relaes democrticas dependem das possibilidades do aluno expressar sua ideias, sentimentos, e da clareza de que o professor tambm lhe dir, com honestidade, o que pensa e ser justo cumprindo suas promessas. Nada mais enganoso do que pensar que o silencio ordem. Disciplina implica a aceitao mutua de normas de conduta e no de sua imposio autoritria. Nunca desistir. bom lembrar que as pessoas reagem diferentemente s mesmas solues, dependendo de sua classe, e do meio cultural a que pertencem. Cap. 9 Perspectivas do ensino de Biologia
A expanso da rede de ensino pblico, provocou a elitizao do ensino mdio pois, se houve um aumento de vagas na escola publica, o ensino nelas ministrados da menores possibilidades a seus estudantes de entrarem na universidade. Consequentemente os alunos com melhor situao financeira passaram a frequentar escolas particulares muitas das quais organizam seus currculos para atender s exigncias do vestibular. O livro didtico um poderoso estabilizador desse estado de coisas, coibindo a funo do professor como planejador e executor do currculo. Alm das dificuldades para dar um ensino experimental, tem dificuldades de acesso a informao e atualizao. Mudanas necessrias Diante da situao presente do ensino de Biologia, salientam-se alm das j propostas anteriormente, vrias mudanas: Enfoque naturalstico biodiversidade: o significado cientifico, econmico e tico do estudo da diversidade biolgica deve ser compreendido pelos alunos, no s como anlise de espcimes mas entendendo que a observao e sistematizao do observado uma atividade cientifica relevante que se consolida nos sistemas de classificao e na taxionomia. Biologia aplicada: a compreenso dos objetivos e procedimentos usados para a soluo de problemas tecnolgicos deve fazer parte da educao dos jovens. Nesse sentido, o primeiro passo sempre identificar uma necessidade: em seguida, propor algumas alternativas para o seu atendimento, das quais se escolhe a melhor, que ser implementada. Na anlise para a escolha da melhor soluo para um problema tecnolgico entram fatores que normalmente no so cogitadas nas anlises feitas pelos professores, quando tratam de problemas cientficos, tais como custo da produo, potencial de uso, eficcia, etc. Multicultural idade: a diversidade cultural no Brasil um fenmeno sempre lembrado nos livros de historia. Diferenas regionais manifestadas por culinrias diferentes, forma de utilizao do ambiente, relaes com os
seres vivos so caractersticas que esto muito presentes na literatura, e nos meios de comunicao, exigindo dos professores um senso de oportunidade para promover discusso, em dimenso global, de muitos dos assuntos tratados nas aulas de Biologia Implicaes sociais: a limitao dos currculos tradicional viso da cincia apenas como atividades para busca do saber ignora o contexto em que ocorre a produo do conhecimento cientifico e a responsabilidade cvica frente s questes referentes s pesquisas cientficas e tecnolgicas. Os professores de Biologia no podem se acomodar diante dessas facilidades e fugir da responsabilidade de ajudar seus alunos a desenvolver as habilidades necessrias. Problemas biolgicos na espcie humana: a sociobiologia que procura oferecer explicaes cientficas para o comportamento social, incluindo o do homem, tem significados explcitos que devem ser tratados em
profundidade e cuidado com os alunos. Educao ambiental: rgos dedicados proteo do meio ambiente, tanto da esfera federal como estadual, envolveram-se em projetos de educao ambiental. No entanto, apesar de ter havido uma multiplicidade de iniciativas, revelando a premncia da soluo dos problemas ambientais. A educao ambiental dever ter um enfoque global e integrado, no podendo ser reduzida a uma disciplina escolar. Os professores de Biologia tm obrigao de se incorporarem ao processo, tanto discutindo os princpios biolgicos necessrios para compreender os problemas ambientais, nas aulas de seus cursos regulares, como participando, sendo membros de uma equipe interdisciplinar na escola ou no grupo de trabalhos comunitrios, sendo para discutir e levar a cabo medidas de proteo ao meio ambiente. Futuro: As projees sobre a situao do ensino de Biologia, nos prximos anos, indicam que a mudanas previstas vo exigir dos professores um profundo
envolvimento nos processo decisrio e um ajustamento para que eles sirvam de elemento de ligao entre a escola e a comunidade.
Atividades 1. Da 5 8 srie do ensino fundamental, os temas comumente ensinados so os seguintes: a. Ser humano b. Sistemas do corpo humano c. Reproduo e estrutura celular d. Plantas e. Fisiologia animal
2Enfatizou a importncia sociocultural no processo de aprendizagem da criana. Seus estudos foram fundamentados na observao das crianas no seu dia-a-dia, incluindo a escola e a famlia. . baseado nessa informao, podemos dizer que, quem fez essa concluso, foi: a. Vygostsky b. Piaget c. Marx d. Gardner e. Bruner
3. Sobre cognitivismo, podemos afirmar, que: a. Este termo engloba teorias que do nfase aos processos mentais dificilmente observveis, enfatizando a possibilidade de o aluno adquirir e organizar informaes. b. Interao do homem como o seu mundo c. Produto dos relacionamentos
d. Prtica do ensino adquirido e. Produto da capacidade de conviver em sociedade
4. Contextualizao sociocultural : a. Conviver bem com o seu ambientes b. Reconhecer a Biologia como um fazer humano e, portanto, histrico, fruto da conjuno de fatores sociais, polticos, econmicos, culturais, religiosos e tecnolgicos. c. Reconhecer a Biologia como um fator fundamental. d. Adaptar-se ao habitat. e. Valorizar somente a cultura.
5. Quais estgios que Piaget identificou no desenvolvimento: a. Sensrio-motor, pr-operatrio, concreto-operacional e operatrio-formal. b. Sensrio-motor e pr-operatrio c. Concreto operacional d. Operatrio formal e sensrio e. Concreto-operacional e sensrio motor
Gabarito
1c / 2a/ 3a / 4b /5a
10. MARGULIS, L.; SCHWARTZ, K. V. Cinco reinos: um guia ilustrado dos filos da vida na Terra. 3. ed. Rio de Janeiro: Guanabara Koogan, 2001. INTRODUO
Este livro versa sobre a superfcie viva do Planeta Terra. Um catlogo da diversidade da vida. Apresenta um sistema de classificao completo, tendo em vista a literatura variante, fragmentada e, frequentemente inconsistente, de onde a nossa informao foi retirada.
Os bilogos estudam organismos individuais, ou parte de populaes, comunidades ou ecossistemas. Estes organismos so classificados com base na forma do corpo, similaridade gentica, metabolismo, padro de desenvolvimento, comportamento e todas as suas caractersticas junto com organismos semelhantes num grupo denominado de espcie. Os cientistas estimam que pelo menos trs milhes, e talvez 30 milhes, de espcies de organismos vivos existam agora. Um nmero ainda maior tem se tornado extinto.
O esforo para discernir ordem, nesta incrvel variedade, deu origem Sistemtica, a classificao do mundo vivo. Os modernos sistematas agrupam espcies proximamente aparentadas em gneros, os gneros em famlias, as famlias em ordens, as ordens em classes, as classes em filos e os filos em reinos. Esta hierarquia conceitual cresceu gradualmente.
SISTEMA MOLECULAR
A sistemtica molecular, ou evoluo molecular como tambm chamado, responsvel por duas inovaes. A primeira a decifrao da sequncia linear e geneticamente determinada dos monmeros componentes nas macromolculas, tais como protenas e cidos necleicos. A segunda a manipulao baseada em computador destas imensas quantidades de dados. Somente computadores de alta velocidade, adequadamente programados, podem organizar e comparar de forma significativa estas enormes quantidades de dados.
OS REINOS DA VIDA
At meados do sculo XX, praticamente todos classificaram os momentos do mundo vivo em dois reinos: plantas ou animais. Contudo, muitos cientistas
notaram que certos organismos, tais como as bactrias e os mofos-de-lodo, diferiam das plantas e dos animais mais do que plantas e animais diferiam entre si. O terceiro e quarto reinos para acomodar estes organismos anmalos foram propostos diversas vezes. Ernst Haeckel, o proponente fez vrias propostas para um terceiro reino de organismos. As fronteiras do novo reino de Haeckel, o reino Protista, variaram durante o curso de sua longa carreira, mas seu persistente objetivo era colocar os organismos mais primitivos e ambguos separados das plantas e dos animais. Contudo, a maioria dos bilogos ignorou as propostas para reinos adicionais alm das plantas e animais ou as considerou curiosidade sem importncia, um pleito especial de excntricos. As opinies em relao aos reinos da vida comearam a mudar na dcada de 1960, principalmente devido ao conhecimento obtido pelas novas tcnicas bioqumicas e da microscopia eletrnica. Entre estas, um sistema de cinco reinos (plantas, animais, fungos, protoctistas e bactrias). A nica ameaa seria para qualquer dos esquemas de cinco reinos o sistema de trs domnios dos microbilogos. Usando critrios moleculares, especialmente sequncias nucleotdicas de RNA ribossmico, esses microbilogos advogam por trs grandes grupos: dois domnios (Archaea e Bactria) consistindo em clulas procariticas, e um domnio (Eukarya) contendo todos os outros organismos. Os fungos, as plantas e os animais so trs dos reinos do domnio Eukarya, da mesma forma como eles esto no nosso esquema de cinco reinos. Contudo, dentro de cada um dos trs domnios h numerosos reinos adicionais muitos correspondentes aos filos no esquema de cinco reinos.
HISTRIA DA DISTINO PROCARIOTA EUCARIOTA
Esta distino fundamental mantida no nvel de super-reino: o Super-reino Prokarya contm os procariotas e somente procariotas, a grande diversidade de organismos com uma organizao do tipo daquela das clulas bacterianas. Os membros dos outros quatro reinos so todos eucariotas. Tanto em estrutura quanto em bioqumica, os eucariotas e os procariotas diferem muito mais do que a presena ou ausncia de um ncleo celular. As clulas procariticas so usualmente mais simples em estrutura (mas no necessariamente em qumica) e menores do que as clulas eucariticas. A distino entre procariota e eucariota imediata e definitiva a partir de micrografias eletrnicas, que mostram estruturas mltiplas dentro das clulas eucariticas. Qualquer estrutura visvel dentro de uma clula uma organela. Os procariotas as tm (carboxomas ou vacolos de gs, por exemplo), mas as
clulas eucariticas contm organelas nicas herdadas, algumas delas separadas por suas prprias membranas do citoplasma. As mitocndrias, organelas ovoides que se especializam em produzir energia pela oxidao enzimtica de compostos orgnicos simples, so encontradas em praticamente todos os eucariotas. Os procariotas no possuem mitocndrias, mas enzimas associadas s suas membranas podem catalisar as oxidaes.
SEXO E REPODUO: PROKARYA E EUKARYA
Reproduo o aumento do nmero de clulas ou organismos, sejam eles unicelulares ou multicelulares. Crescimento um aumento no tamanho. Todas as espcies de organismos crescem e se reproduzem. Sexo, a recombinao de genes de dois ou mais indivduos, de fato ocorre nos procariotas, mas o sexo procaritico no diretamente exigido para reproduo. Clulas bacterianas procariotas nunca se fundem. Em vez disso, os genes do meio fluido, de outra espcie, de um vrus ou de onde quer que seja penetram nas clulas bacterianas por si prprios. As clulas eucariticas se reproduzem por mitose. Elas formam cromossomos pacotes de genes densamente enrolados, mantidos juntos por protenas e anexados s membranas internas do ncleo. Todos os animais se reproduzem por fertilizao de um vulo por um espermatozide menor, para formar o ovo frtil zigoto que se divide por mitose para produzir um embrio rudimentar. O primeiro estgio embrionrio chamado de blstula. Todas as plantas formam esporos que, por si prprios, crescem em uma de duas formas de gametfitos (plantas que formam gametas): ou um macho e formao de espermatozide, ou uma fmea e formao de vulo. Os fungos reproduzem-se por meio de esporos fngicos, propgulos capazes de gerar de um nico genitor, assexuadamente o organismo fngico inteiro novamente. Alguns fungos entram na sua fase sexuada somente quando o ambiente no favorece mais a reproduo assexuada. Os proctistas apresentam uma grande variabilidade nas suas caractersticas de ciclo de vida mas nenhum se encaixa na descrio de animal, planta ou fungo. O reino proctista inclui os eucariotas microbianos (unicelulares ou de poucas clulas) e seus descendentes diretos multicelulares. Protistas se referem aos menores protoctistas, mas algumas pessoas usam o termo para todos eles.
VRUS
Antony van Leeuwenhoek (1632-1723), o descobridor do mundo microbiano. O vrus, compostos de DNA (cido desoxirribonuclico) ou RNA (cido ribonuclico), envolvidos por uma capa de protena, so muito menores do que as clulas. Embora se repliquem, eles s podem fazer isso, entretanto, numa clula e usando os dispositivos de vida desta. Fora da clula, os vrus no podem reproduzir-se, alimentar-se ou crescer. Alguns vrus podem mesmo se cristalizar, como minerais. Neste estado, podem sobreviver por anos sem transformao at que entrem em contato com tecidos vivos especficos dos quais necessitam.
LENDO ESTE LIVRO
Reconhecemos e descrevemos dois super-reinos: Prokarya (todas as bactrias) e Eukarya (todos os organismos nucleados), nestes dois maiores, os txons mais internos so os reinos. O Super-reino Prokarya contm o Reino Bactria (equivalente aos Prokaryota, Procariota, ou Monera em outros esquemas) com seus dois sub-reinos, Archaea e Eubacteria. O Sub-reino Archaea contm dois filos: B-1, o Euryarchaeota, ou os metanogenes e extremos halfilos, e o B-2, o Crenarchaeota, ou extremos termfilos. O Subreino Eubacteria contm 12 filos. No Reino Eukarya (Protoctista, Animalia, Fungi e Plantae) esto 30 filos protoctistas, 37 filos animais, 3 filos fngicos e 12 filos vegetais. Dentro de cada Reino de Eukarya, os filos esto organizados numa ordem adequada, das formas mais simples (presumivelmente, os que devem ter evoludo mais cedo) at as formas mais complexas (presumivelmente, os mais recentes). Como regra, os txons mais altos dentro de um reino, os filos e as classes, representam as divergncias evolutivas mais antigas; os txons mais baixos, os gneros e as espcies, representam as mais recentes. Contudo, isso no uma regra absoluta, porque as relaes evolutivas de muitos grupos so desconhecidas. Os organismos esto agrupados no mesmo txon somente por enquanto, porque eles tm alguns atributos claramente distintos em comum (por exemplo, as hastes radiais dos actinpodes), independentemente de seus ancestrais comuns terem sido documentados.
SUPER-REINO PROKARYA
Sistemas genticos limitados por uma nica membrana: nucleoides, sntese proteicas em pequenos ribossomos, recombinao somente no nvel do DNA. Sem fuso nuclear; ausncia de fuso citoplasmtica e nuclear (isto , fertilizao), implicando ausncia de gentica mendeliana. Apresentam transferncia gnica unidirecional e, em observao microscopia, ausncia de motilidade intracelular visvel. Reproduo por fisso binria, brotamento, brotamento de filamentos, fisso do parental sssil para produzir uma descendncia flagelada, crescimento polar (de ponta a ponta), ou fisso mltipla.
Super-reino Prokarya Reino Bacteria Sub-reino Archaea (Arqueobacteria) Sub-Reino Eubacteria
CAPTULO UM BACTERIA
O Reino Bacteria compreende todos os organismos com estrutura celular procaritica: eles tm pequenos ribossomos no entorno dos seus nucleoides, mas no possuem ncleos envolvidos por membrana. Em termos de atividade e potencial para um crescimento explosivo, as bactrias so imbatveis entre os organismos vivos. As bactrias so os mais resistentes dos seres vivos. Algumas podem sobreviver a temperaturas muito baixas, bem abaixo do congelamento, por anos; outras vicejam em fontes de gua fervente; e outras ainda, crescem at em cidos muito fortes (hot acid), ou vivem retirando hidrognio e dixido de carbono das rochas. Por meio da formao de propgulos, tais como os esporos partculas de vida viajantes que contm pelo menos uma cpia de todos os genes de um organismo muitas toleram gua em ebulio ou uma total dessecao. As bactrias so as primeiras a invadir e colonizar novos habitat: terra que foi queimada, solos vulcnicos ou ilhas recentemente formadas. No entanto, no se tem conhecimento de nenhum organismo nem mesmo a mais resistente das bactrias que seja capaz de completar seu ciclo de vida suspenso no ar ou em qualquer outro gs. Devido a sua limitada morfologia e escassez de registros fsseis, as bactrias tm relaes evolutivas que so excessivamente difceis de apurar.
Embora sejam pequenas, as bactrias que so extremamente numerosas e de crescimento rpido, so cruciais para a sade dos sistemas digestivos, para a manuteno do solo na agricultura e arboricultura, e para a existncia do ar que respiramos. A maioria das bactrias nunca patognica. Os patgenos so simplesmente bactrias (ou, ocasionalmente, protoctistas ou fungos) capazes de causar doenas infecciosas em animais ou plantas. As bactrias podem curar tanto quanto causar doenas. As bactrias so morfologicamente bastante simples: esfricas (cocos), em forma de basto ou em forma de espiral. As mais complexas sofrem mudanas de forma durante o seu desenvolvimento: bactrias simples podem reproduzirse gerando populaes que se metamorfoseiam em estruturas alongadas, gerando filamentos longos ou ramificados, ou formando corpos altos que liberam microcistos na forma de esporos. Algumas produzem colnias altamente mveis. As bactrias so facilmente agrupadas por propriedades da parede celular, distinguidas por um procedimento de colorao. Um teste diagnstico de bactrias, universalmente aplicado, se elas se coram de prpura ou rosa com o teste de Gram, um mtodo de colorao desenvolvido pelo medico dinamarqus Hans Christian Gram (1853-1938). Os organismos gram-positivos (que se coram de prpura profundo) diferem daqueles gramnegativos (que se coram de rosa-claro). A qumica das suas paredes celulares a presena de uma membrana extra nas bactrias gram-negativas a base para classificao. Embora algumas molculas muito complicadas sejam feitas por certas plantas e fungos, os padres biossintticos e degradativos a qumica do uso do alimento e da gerao de energia so marcantemente similares em todas as plantas e clulas fngicas. Os animais e os protoctistas apresentam at mesmo uma variao menor nos seus repertrios qumicos. Em resumo, o metabolismo dos eucariotas bastante uniforme; seus padres de fotossntese, respirao, quebra da glicose e sntese dos cidos nuclicos e protenas so fundamentalmente os mesmos em todos os organismos eucariotas. As bactrias, por outro lado, so no apenas metabolicamente diferentes dos eucariotas, mas tambm diferentes entre si. A superfcie da Terra seu regolito, sedimentos e guas rica no somente em bactrias vivas, pequenos animais, protistas, leveduras e outros fungos, mas tambm em complexos compostos orgnicos que produzem. Os substratos menos tratveis, como cidos tnicos, lignina e celulose, tendem a se acumular, enquanto orgnicos muito mais ativamente metabolizados, tais como acares, amido, compostos organofosforados e protenas, so produzidos e removidos mais rapidamente. Todos estes compostos orgnicos so, direta ou indiretamente, os produtos da quimiossntese ou da fotossntese,
processos que usam energia qumica ou luz do Sol, respectivamente, para converter o dixido de carbono do ar em compostos orgnicos dos quais obtemos leo, gs e carvo. A quimiossntese limitada a certos grupos de bactrias. A fotossntese, que muitas vezes erradamente atribuda somente s algas e plantas, executada por muitos grupos de bactrias. A quimiossntese e a fotossntese so frequentemente, mas no sempre, correlacionadas com os processos que usam qumicos inorgnicos ou luz, respectivamente, para gerar energia para elaborar compostos orgnicos. Ambos os tipos de sntese so formas de nutrio autotrfica estrita, uma sntese de todo o alimento e a obteno da energia exclusivamente de fontes inorgnicas. A fotossntese, o processo, comeou nas bactrias anaerbias h mais de 3 bilhes de anos. As bactrias que obtinham sua energia da luz do Sol, seu carbono do CO, do ar e seus eltrons do H, HS, HO ou outras fontes inorgnicas, so chamadas bactrias fotolitoautotrficas. Elas se alimentamde luz do Sol. A noo de que a teia alimentar comea com as plantas, seguida pelos herbvoros e termina com os animais carnvoros por demais simplista. O zooplncton dos mares se alimenta dos protoctistas: protoctistas nofotossintetizadores se alimentam de bactrias; bactrias (e fungos necrfagos) decompem as carcaas de animais, plantas e algas, liberando de volta para a soluo elementos como o nitrognio e o fsforo, exigidos pelo fitoplncton. Como filo significa planta, e nenhuma planta flutua no oceano aberto, preferimos o termo fotoplncton para nos referirmos s bactrias e algas flutuadoras que fotossintetizam no mar e nos lagos. As bactrias, por serem comidas por outros, facilitam teias alimentares inteiras. Os caminhos pelos quais ns e outras formas de vida dependemos das bactrias, e nos desenvolvemos a partir delas, sero explicados nas descries dos filos. A vida na Terra desapareceria muito mais rapidamente se os organismos do reino Bacteria se tornassem extintos do que com a extino dos organismos de qualquer outro dos reinos. Acreditamos que a vida bacteriana no nosso planeta floresceu muito antes que os grandes organismos, que se desenvolveram por simbiose a partir de comunidades bacterianas, aparecessem.
SUB-REINO ARCHAEA
As caractersticas distintas das aqueobacterias se referem principalmente s sequncias de genes (DNA) que determinam as sequncias de pequenas subunidades ribossomais de RNA (RNAr). As arqueobacterias so mais
aparentadas umas com as outras do que com as eubactrias. Os ribossomos arqueobacterianos so mais semelhantes na ultra-estrutura aos ribossomos eucariticos do que aos ribossomos eubacterianos. Os grandes lipdios de arqueobactrias so teres conectados com cadeias laterais de fitanol. Em outras bactrias e nos eucariotas, os grandes lipdios so ligados por steres. As arqueobactrias no possuem camadas glicopeptdicas caractersticas das paredes celulares das eubactrias. As arqueobactrias incluem as metanognicas, haloflicas e termoacidoflicas. Todas as outras bactrias isto , a maioria so eubactrias. Chamamos a ateno para os ambientes antigos, dos quais membros dos dois filos arqueobacterianos Euryachaeota e Crenarchaeota tendem a ser encontrados: embora se tenha pensado que os habitat de arqueobactrias eram tpicos da superfcie da Terra durante o on Arqueano, h mais de 3 bilhes de anos (isto , ambientes tectonicamente ativos), estudos recentes tem mostrado que as arqueobactrias estavam dispersas na gua do mar, lago, solos e outros ambientes no sujeitos a condies extremas.
B-1 EURYARCHAEOTA
O Filo Euryarchaeota compreende dois grupos muito estranhos e diferentes da arqueobacterias, as metanogenes e as halfilas. Elas esto agrupadas juntas, com base na similaridade na sequncia de RNAr. As metanogenes podem ser gram-positivas ou gram-negativas, com motilidade (atravs de flagelos) ou sem. Todas as trs formas clssicas de bactrias bastonete, espiral e coco esto representadas: as metanogenes variam de bastonetes curtos e cocos irregulares at espirais, grandes cocos em grupos, e filamentos. At o momento, nenhuma metanogene com filamentos ramificados ou com flagelos internos (periplasmticos) foi descoberta. A maioria das espcies pode tolerar temperaturas moderadas ou altas. As metanogenes so encontradas em todo o mundo nos esgotos, nos sedimentos marinhos e de gua doce, e nos tratos intestinais de animais tanto nos ruminantes tanto nos insetos comedores de celulose. Formalmente, os gneros reconhecidos so distinguidos pela morfologia e fisiologia. Todos tm nomes pr-fixados por metano-: Methanosarcina, Methanobacterium, Methanococcus, Methanobacillus e Methanothrix. As halfilas vivem em ambientes externos, salgados ou alcalinos, e no formam esporos. Antes do uso das tcnicas filogenticas moleculares, o parentesco prximo entre as halfilas e as metanogenes era desconhecido.
As bactrias metanognicas no podem usar acares, protenas ou carboidratos como fontes carbono e energia. De fato, a maioria pode usar somente trs compostos como fontes de carbono: formiato, metanol e acetato. As metanognicas so caracterizadas por sua forma extraordinria de obter energia: elas formam metano (CH4) atravs da reduo do dixido de carbono (CO) e da oxidao do hidrognio (H), uma comodidade rara. Como obtm ambos estes gases do ar sua volta e no podem tolerar oxignio, sua distribuio limitada. Alm de usar H, algumas usam formiato, metanol ou acetato como uma fonte de eltrons para reduo do CO. A reao metablica geral caracterstica :
CO + 4H -> CH4 + 2HO
As halfilas so aerbios que respiram oxignio, diferentes das metanogenes, intolerantes a este gs. As bactrias que demandam sal so incapazes de produzir metano, mas, como as metanogenes, so adaptadas a condies ambientais extremas e nenhuma forma esporos. As halfilas sobrevivem em diversos ambientes hipersalinos recentemente descobertos, assimilando como alimentos diversos compostos de carbono. Alguns exemplos incluem as bactrias em forma de bastonete, como Halobacterium salinarium, que vive em peixes salgados; H. sodomense, que requer altas concentraes de magnsio; e a consumidora de acar e tolerante ao sal H. saccharovorum. As clulas em forma de disco achatado ou de clice so membros do gnero Haloferax, enquanto as halfilas triangulares e retangulares so usualmente associadas ao gnero Haloarcula. A maioria das halfilas tem metabolismos aerbicos obrigatrios e motilidade. As halfilas vivem em ambientes de alta fora inica. Notavelmente, a maioria das halfilas estudadas resistem a solues saturadas de cloreto de sdio das salinas em todo o mundo. Elas produzem carotenides rosa-brilhantes e podem mesmo ser visualizadas a partir de avies de satlites orbitais como espuma rosa sobre plancies de sal. Muitas de suas protenas tm modificaes que as permitem funcionar somente em altas concentraes de sal. Suas paredes celulares so bastante diferentes das outras bactrias, no sentido de que elas no tm derivados de cidos diaminopimlicos e murmicos. As membranas comuns de lipoprotenas se rompem ou se desfazem em altas concentraes de sal, mas os lipdios especiais das halobactrias incluem derivados de diter glicerol que estabilizam as membranas sob altas concentraes de sal.
B-2 CRENARCHAEOTA
As arqueobacterias do Crenarchaeota so encontradas em fontes quentes por todo o mundo. Habitam fontes geotrmicas na Islndia, giseres do Parque Nacional de Yellowstone, fluidos de erupes vulcnicas submarinas e outros habitat com condies muito quentes, muito cidas e muito ricas em enxofre e pobres em oxignio, condies que so muito mais familiares para as eubactrias. As termoacidfilas estudadas, tais como Thermoproteus e Sulfolobus, tm paredes celulares fortes e resistentes a cidos, compostas de um material glicoprotico arranjado num padro de subunidades hexagonais. A Thermoplasma, como os micoplasmas do Filo B-9, dois quais aparentada, no possui nenhuma parede. As clulas do gnero Sulfolobus, que crescem em gua a 90C e valores de pH de 1 a 2 (a acidez de cido sulfrico concentrado), foram primeiro isoladas em cultura em 1972. Algumas morrem de frio em temperaturas abaixo de 55oC. A Sulfolobus acidocaldarius tem um nome bem adequado, pois crescem em ambientes que vo de um pH de 0,9 a 5,8, preferindo guas cidas num pH de 2 a 3, tem paredes celulares sem glicopeptdios e so autotrficas facultativas. As termoplasmas so os nicos procariotas conhecidos que contm DNA coberto com protenas bsicas semelhantes s histonas, as protenas cromossmicas da maioria dos eucariotas. Acredita-se que a cobertura proteicas protege seu DNA da desnutrio no cido quente. A Methanothermus, uma metanognica, tambm tem sua cobertura de protena no seu DNA. A Thermoplasma pode ser relacionada com o ancestral do ncleo-citoplasma dos eucariotas.
SUB-REINO EUBACTERIA
Todas as bactrias que no so arqueobactrias so eubactrias. Este ltimo grupo, que compreende a grande maioria das bactrias descritas na literatura, extremamente diverso em morfologia e metabolismo. Todas as bactrias multicelulares com estruturas complexas, ou capacidade para diferenciao celular, pertencem a este grupo, assim como todas as bactrias que retiram energia diretamente da luz do sol e do carbono do ar (isto , bactrias fototrficas). Todas as clulas, incluindo as bactrias, devem manter uma membrana celular intacta todo o tempo, e a parede celular est sempre do lado de fora desta membrana celular intacta (ou plasma) (que tambm o envelope da clula): assim, os procariotas so classificados pelos bacteorologistas de acordo com suas paredes. A classificao baseada em paredes no
necessariamente consistente com aquela baseada no RNAr 16S; a classificao consistente e abrangente deste livro reconciliou as vises e os dados publicados por tantos bilogos quanto possvel, numa forma mais vivel para os estudantes seguirem. As vrias classificaes para paredes celulares de eubactrias so: paredes gram-negativas (Filo B-3 at B-8), sem paredes (Filo B-9) e paredes grampositivas (Filo B1-10 at B-14). A reao de colorao gram-negativa se correlaciona com a presena de uma camada externa de lipoprotena da parede celular, e uma fina camada interna de glicopeptdios (unidades de peptdio anexadas a acares nitrogenados) ligada ao lado de dentro pela membrana do plasma. As paredes gram-positivas esto associadas com certas protenas ligadas parede. As trs divises eubacterianas so Firmicutes, Gracilicutes e Tenericutes. A diviso Firmicutes (do latim firmus, forte durvel) inclui todas as eubactrias gram-positivas, que tendem a ter paredes glicopeptdicas frequentemente espessas que no possuem camadas lipoproteicas externa. A diviso Gracilicutes (do latim gracilis, delgado, fino) inclui todas as eubactrias gram-negativas que, mesmo que tenham uma membrana lipoproteicas externa, tm geralmente paredes celulares mais finas. A diviso Tenericutes (do latim tener, suave, delicado) compreende as eubactrias que no possuem paredes celulares e so geneticamente incapazes de sintetizar precursores de glicopeptdios. Baseados nas suas molculas de RNA e outras caractersticas, a maioria das eubactrias, sem parede, evoluram de membros da Firmicutes, as eubactrias gram-positivas.
DIVISO: GRACILICUTES B-3 Proteobacteria (Bactrias prpuras) Os membros deste grande grupo de eubactrias so classificados por dados de RNAr 16S, pela morfologia e pelo metabolismo.
B-4 Spirochaetae Do latim spira, espiral; e do grego khaite, plo longo As espiroquetas se parecem muito com serpentes helicoidais. Diferente de outras bactrias moveis, elas tem de 02 a mais de 200 flagelos internos (filamentos axiais ou endoflagelos), no espao entre a membrana interna
(plasma) e a membrana celular externa da parede celular gram-negativa, isto , os flagelos esto na parede celular.
B-5 Cyanobacteria (verde, cloroxibactria e bactria azul-esverdeada) Do grego kyanos, azul-escuro Estes dois grupos de bactrias fotossintetizadoras dependentes de oxignio diferem em cor e pigmentao. De longe o maior grupo, com talvez mil gneros, esto as azul-esverdeadas. Somente uns poucos gneros, recentemente descritos, de bactrias verdes fotossintetizadora, as cloroxibacterias, so conhecidas: Prochlorococus, Prochloron e Prochlorothrix.
B-6 Saprospirae (Deslizadores de fermento) Do grego sapros, apodrecido, decomposto; spira, rosca, espiral A sequncia gnica da pequena subunidade da molcula RNAr 16S revelou esta linhagem de eubactrias gram-negativas. Os bacterides e seus parentes, fermentadores anaerbios, formam um subgrupo; o outro, subgrupo de flavobactrias, unifica um conjunto de respiradores de oxignio que se movem por deslizamento: Capnocytophaga, Cytophaga, Flexibacter, Microscilla, Saprospira e Sporocytophaga.
B-7 Chloroflexa (Fottrofas verdes no-sulfurosas) Do latim flexus, curvo; e do grego chloro, verde Novos conhecimentos, especialmente da sequncia de nucleotdeos nos seus genes RNAr 16S, levaram a separao por completo dessas bactrias fototrpicas de outras bactrias fototrpica verdes de enxofre, com as quais tinham sido agrupadas.
B-8 Chlorobia (Bactrias verdes anoxignicas sulfurosas) Do grego chloro, verde; bios, vida Nem todos os organismos fotossintetizadores so fototrficos. Qualquer forma de vida que usa a luz para sntese de material celular fotossintetizadora, mas somente aqueles organismos que no precisam fixas compostos orgnicos no meio so fototrficos.
DIVISO: TENERICUTES B-9 Aphragmabacteria (Micoplasmas) Do grego a, sem; phragma, parede Todas as clulas, incluindo as das eubactrias sem parede, os micoplasmas, so limitadas por uma membrana celular de sua prpria fabricao, que permite a passagem de gua, ons de sais e pequenos compostos orgnicos.
DIVISO: FIRMICUTES B-10 Endospora (Formao de endsporo e bactrias gram-positivas de baixa relao G + C) Do grego endos, dentro; spora, semente As Endospora incluem anaerbios fermentadores obrigatrios e aerbios facultativos ou obrigatrios, a maioria das quais se coram como clulas grampositivas com parede, e todos pertencem ao grupo RNAr 16S de baixo GC de eubactrias.
B-11 Pirellulae (Bactrias de paredes proteicas e seus parentes) Denominao baseada no gnero Pirelulla (do latim pirus, pra) Este novo e diverso grupo foi revelado por informao de sequncia de RNAr 16S. pelo menos dois destes gneros Pirelulla e Planctomyces (e provavelmente Gemmata) tem paredes celulares no-glicopeptdicas e proteicas.
B-12 Actinobacteria (Actinomicetos, actinomicotas; e bactrias grampositivas com alta relao G + C) Do latim aktis, raio; bakterion, bastonete
Este Filo, unificando bactrias semelhantes determinadas pelos critrios da morfologia, fisiologia e RNAr 16S, inclui as bactrias conformes e actinobactrias filamentosas.
B-13 Deinococci (Bactrias gram-positivas resistentes ao calor ou resistentes radiao) Do grego deino, terrvel, estonteante; do latim coccus, gro Deinococci, um novo Filo, baseado principalmente na nica identificao da sequncia de gene RNAr 16S, compreende bactrias gram-positivas, altamente resistentes, heterotrficas, que exigem oxignio para o seu crescimento.
B-14 Thermotogae (Fermentadores termoflicos) Denominao baseada no gnero Thermotoga (do grego Thermo, calor; do latim, toga, roupa de cidado romano) Estas bactrias recentemente descobertas so to impressionantemente diferentes nas suas sequncias de RNAr de todas as outras, que garantiram para si prprias um novo Filo.
SUPER-REINO EUKARYA
Organismos compostos de clulas que se reproduzem por mitose, cada uma contendo mais que dos cromossomos. Na mitose, ncleos circundados por membranas contendo poros se dissolvem total ou parcialmente, e se refazem como dois novos ncleos descendentes. Apresentam fuso nuclear e citoplasmtica vivel e seus processos recprocos (por exemplo, duplicao citoplasmtica e cromossomial com subsequente contedo meitico ou reduo equivalente de contedo citoplasmtico; desta forma, gentica mendeliana). Clulas com citoesqueletos de tubulina-actina capazes de mobilidade intracelular. No possuem transferncia unidirecional de genes (DNA desprotegido).
Super-reino Eukarya Reino Protoctista
Reino Animalia Reino Fungi Reino Plantae
PROTOCTISTA Do Grego protos, primeiro; kristos, estabelecer
Microorganismos nucleados e seus descendentes, exclusivos de fungos, animais e plantas; evoludos a partir da integrao de simbiontes microbianos anteriores. Meiticos ou no-meiticos, com variaes no crculo de fertilizao da meiose. Os registros fsseis se estendem desde a era Proterozoica Mdia Inferior (cerca de 1,2 bilho de anos atrs) at o presente.
O Reino Protoctista compreende os microorganismos eucariticos e seus descendentes imediatos: todas as algas, incluindo as ervas-do-mar (seaweeds), mofos-de-gua undulipodiados (flagelados), os mofos-de-lodo (slime molds) e os slime nets, os tradicionais protozorios e outros organismos aquticos ainda mais obscuros. Seus membros no so animais (que se desenvolvem de uma blstula), nem plantas (que se desenvolvem de um embrio), nem fungos (que no possuem undulipdios e se desenvolvem de esporos). Nem so protoctistas procariotas. Todas as clulas protoctistas tm ncleos e outros atributos caracteristicamente eucariticos. Muitos fotossintetizam (tm plastdios) muitos so aerbios (tm mitocndrias) e muitos tm undulipdios [9(2)+2], com as suas bases de cinetossomo em algum estgio do seu ciclo de vida. Todos os protoctistas se desenvolveram por simbiose dentre pelo menos dois tipos diferentes de bactrias em alguns casos, dentre muito mais do que dois. medida que os simbiontes se integraram, um novo nvel de individualidade surgiu. Os perfis de pigmentao fotossinttica, essenciais funo do cloroplasto, so critrios importantes, tambm empregados pelos taxonomistas para resolver a desconcertante diversidade do Reino Protoctista. Os undulipdios estavam presentes nos ancestrais comuns de todos os filos, mesmo antes das mitocndrias, dado que os arqueoprotistas anaerbicos os carregavam. Seu comportamento durante a mobilidade e a reproduo est relacionado com a diviso celular mittica.
Os protoctistas so aquticos: alguns marinhos, alguns de gua doce, alguns terrestres em solos midos e alguns parasitas ou simbiticos em tecidos midos de outros. Aproximadamente todos os animais, fungos e plantas talvez todos tm protoctistas associados. Os filos, tais como Microspora (Filo Pr-2) e Apicomplexa (Filo Pr-9), incluem milhares de espcies, todas as quais vivem em tecidos de outros. Com uma reverncia diversidade protoctista, um reconhecimento da sua herana eucaritica comum e um senso de humildade tanto em relao sua complexidade quanto nossa ignorncia, apresentamos nossos 30 filos protoctistas.
Pr-1 Archaeprotista Do grego karyon, ncleo, centro; blastos, boto, broto Embora a vasta maioria de organismos nucleados de fato, todos eles, exceto aqueles neste filo e outro sejam aerbios com obrigao de assimilar oxignio atmosfrico, os ancestrais dos eucariotas eram originalmente anaerbios, mortos pelo oxignio.
Pr-2 Microspora (Microsporida) Do grego mikos, pequeno; spora, esporo Os micrsporos, o segundo filo de amitocondriados, so micrbios heterotrficos, que no possuem mitocndria e produzem algum tipo de filamento ou cordo polar.
Pr-3 Rhizopoda (Amebas amastigotas e mofos-de-lodo celulares) Do grego rhiza, raiz; pous, p Como definido aqui, os membros do Filo Rhizopoda amebas amastigotas so amebas que tem mitocndrias e no possuem undulipdios em nenhum dos estgios de suas historias de vida.
Pr-4 Granuloreticulosa Do latim granulum, gro pequeno, semente; reticulum, rede
Granulorreticulosandeos (granukireticulosans) so facilmente definidos: estes organismos possuem reticulpodes, clulas que se fundem para formar redes, nas quais o movimento bidirecional (de mo dupla) pode ser visto.
Pr-5 Xenophyophora Do grego xenus, estranho; pheyin, carregar para adiante, gerar; pherein, portar Habitantes bentnicos (do fundo do mar) grandes e mesmo abissais, estes protoctistas enigmticos so objeto de grande curiosidade
Pr-6 Myxomycota (Myxogastria, mofos-de-lodo primordiais) Do grego myxa, muco; mykes, fungo Os xomicetos gozam de muitos nomes, incluindo mixomicetos, micetozona, mofos-de-lodo plasmodianos, mofos-de-lodo verdadeiros, Myxomycotina e outros. Como os mofos-de-lodo celulares formam um estgio amebide que no possui paredes celulares, e se alimentam de bactrias engolfando-se com pseudpodes por exemplo, fagocitose.
Pr-7 Dinomastigota (Dinoflagellata, Dinophyta) Do grego dinos, retoro, rotao, convoluo; mastigio, chicote Dinomastigotas, Ciliophora e Apicomplexa so classificadas juntas como alveoladas porque todas tem alvolos de superfcie, ou fossas, e sequncias comuns de RNA ribossmicos. Investigaes posteriores podem levar aos seus reposicionamentos como trs classes de um novo filo, Alveolata.
Pr-8 Ciliophora (Ciliados) Do latim cilium, clio, plpebra inferior; do grego pherein, portar A maioria dos ciliados que esto entre os protoctistas mais bem conhecidos, so clulas nicas bactervoras. Os ciliados so caracteristicamente cobertos com clios curtos undulipdios com cinetos somos embutidos num crtex duro externo fibrilar (camada celular proteicas) da clula.
Pr-9 Apicomplexa (Sporozoa, Telosporidea)
Do latim apex, picol complexus, uma envoltria, revestimento A Apicomplexa compe simbitrofos unicelulares modificados para penetrar tecidos e obter alimento de animais. Todos formam esporos. Diferente dos esporos bacterianos, os esporos dos apicomplexos no so resistentes ao calor e dessecao.
Pr-10 Haptomonada (Prymnesiophyta, Haptophyta, cocolitofordeos) Do grego haptein, apertar; phyton, planta As haptomnadas planctnicas, pequeninas tm sido vistas pelos bilogos marinhos e paleontlogos como dois tipos diferentes de organismos: algas douradas e com mobilidade, semelhantes a crisomnadas planctnicas e cocolitofordeos cobertos com cocolita. Os coclitos, estruturas microscpicas em forma de disco, de carbono de clcio, de renome para os paleontlogos, so produzidos por cocolitofordeos, que as carregam como placas de superfcie empacotadoras.
Pr-11 Cryptomonada (Cryptophyta) Do grego kriptos, escondido; do latim monas, unidade; phyton, planta As criptomnadas so as clulas nadadoras, elpticas e achatadas. So heterotrficas e fotossintticas, e encontradas em todo o mundo nos lugares midos.
Pr-12 Discomitochondria (Flagelados, zoomastigotas, zooflagelados) Do grego zoyon, animal; mastix, chicote At que detalhes de suas relaes com o resto do mundo protoctista sejam revelados, este novo grupo convenientemente retm quatro subgrupos bem definidos de mastigotas unicelulares, moveis bactervoros ou osmotrficos, nadadores, e suas colnias derivadas.
Pr-13 Chrysomonada (Crisfita) Do grego chrysus, dourado; phyton, planta
Um novo agrupamento do Filo Pr-13 at Pr-12, de crisomnadas at hipoquitrdeas, foi estabelecido com base na similaridade das sequncias de genes RNA ribossmico, o que sugere que elas tem um ancestral em comum.
Pr-14 Xanthophyta Do grego xanthos, amarelo; phyton, planta As xantofitas, como as eustigmatfitas, so verde-amareladas. Contudo, a organizao nica de suas clulas, a sua tendncia a formar colnias estranhas, sugere que elas so aparentadas com as eustigmas somente por pigmentao.
Pr-15 Eustigmatophyta Do grego eu, verdadeiro, original, primitivo; stigma, marca colocada num escravo (em referncia a um sinal), marca, ponto; phyton, planta As eustigmatfitas foram originalmente agrupadas junto com as xantfitas, com as quais se assemelham, devido a ser verde-amareladas, formarem clulas vegetativas cocoides imveis e se propagam por meio de zosporos assexuadas alongados e mveis.
Pr-16 Diatomceas (Bacilarifita) Do latim bacillus, graveto; do grego phyton, planta Belos protistas aquticos talvez cerca de 10.000 espcies vivas as diatomceas sai unicelulares, ou formam colnias ou filamentos simples. Cerca de 250 gneros destes fantsticos protoctistas so usualmente descritos, e os especialistas reconhecem a enorme quantidade de 100.000 espcies, incluindo aquelas de 70 gneros fsseis.
Pr-17 Phaeophyta (Alga parda) Do grego phaios, escuro, pardo; phyton, planta As fefitas so as algas-do-mar pardas: praticamente todas so marinhas. Elas so os maiores protoctistas. As kelps gigantes, por exmeplo, chegam at 10m. Cerca de 900 espcies em 250 gneros j foram descritas, todas elas fotossintticas.
Pr-18 Labyrinthulata (Slime nets e traustoquitrdeos) Do latim labyrintulum, pequeno labirinto Estudos moleculares de RNA ribossmico tm confirmado a relao entre as duas classes deste filo: os traustoquitrdeos e as slime nets, ou labitintuldeos.
Pr-19 Plasmodiophora Do latim moderno plasmodium, massa multinucleada de protoplasma no dividida em clulas; do grego pherein, portar Os plasmodiforos, 10 gneros e 29 espcies, todos micrbios osmotrficos, so simbitrofos obrigatrios. A maioria das espcies vive dentro de plantas.
Pr-20 Oomycota (Oomicetos) Do grego oion, ovo; mykes, fungo Os oomicetos saem distinguidos de outros protoctistas semelhantes aos fungos pelos cinetdeos e outros detalhes da estrutura do zosporo, e tambm pela natureza de seus ciclos de vida sexual.
Pr-21 Hyphochytriomycota Do grego hyphos, teia: chytra, pequeno pote de cozimento de barro: mykes, fungo Os hifoquitrdeos, com quitrdeos e oomicetos, tm sido tradicionalmente considerados fungos. Estes osmtrofos aquticos de fato se assemelham aos fungos no seu modo de nutrio, que pode ser simbiotrfico ou saprbico.
Pr-22 Haplospora Do grego haplo, nico, simples; do latim spora, esporo A evoluo convergente que levou a estruturas simbiotrficas, pequenas e escuras em tecidos animais unifica este filo, assim como tambm o Filo Pr-23 e Pr-24 trs filos de parasitas em forma de propguos (por exemplo, formadores de cisto) com um filo alveolado, o Apicompleca, e outro protozorio parastico na categoria Sporozoa.
Pr-23 Paramyxa Do grego para, ao lado de; myxa, muco Anteriormente ignorados ou agrupados com os esporozorios, os paramixanos simbiotrficos so imediatamente distinguidos de todos os outros organismos pelo seu comportamento de clulas de aninhamento.
Pr-24 Myxospora (Myxozoa, mixospordeos) Do grego myxa, muco; do latim spora, esporo Este filo consiste nos tradicionais mais bem conhecidos esporozorios, organismos multicelulares que alguns acreditam serem animais. Estes formadores de esporos aerbicos so frequentemente confundidos com microscpios amitocondriados, devido a membro de ambos os grupos terem filamentos polares, embora seus respectivos filamentos difiram grandemente em estrutura e funo.
Pr-25 Rhodophyta (Alga vermelha) Do grego rhodos, vermelho; phyton, planta Os organismos sexuais deste filo e do Gamfita provavelmente desenvolveram seu sistema de acasalamento de conjugao peculiar independentemente de qualquer outro, e independentemente dos fungos, a maioria dos quais apresenta comportamento similar.
Pr-26 Gamophyta (Conjugaphyta, algas verdes de conjugao) Do grego gamos, casamento; phyton, planta As gamfitas so algas verdes que no possuem undulipdis em todos os estgios de sua histria de vida. Sem espermatozides mveis, qualquer outro espermatozide ou outros meios de locomoo, elas normalmente entram em processos sexuais.
Pr-27 Actinopoda Do grego actinos, raio; pous, p
Os protistas marinhos que Ernst Haeckel tradicionalmente chamou de radiolarianos e outros plnctons superficialmente semelhantes protistas grandes com alguma simetria radial, so agrupados como classes no Filo Actinopoda por convenincia e didtica.
Pr-28 Chlorophyta (Algas verdes) Do grego chloros, verde-amarelado; phyton, planta As clorfitas so algas que tem cloroplastos verdes, circundados por duas membranas, e que formam zosporos ou gametas com undulipdios, normalmente pelo menos dois, de igual tamanho.
Pr-29 Chytridiomycota Do grego chytra, pequeno pote de barro de cozinhar; mykes, fungo Dados recentes de sequncia gentica de RNA ribossmico reforaram a hiptese de que os quitrdeos evoluram, pela perda de undulipdios, nos ancestrais da linhagem dos fungos.
Pr-30 Zoomastigota (Zoomastigotas, zooflagelados) Do grego zoion, animal; mastix, chicote Os zoomastigotas so unicelulares fagotrficos ou nadadores aerbicos osmotrficos. Antigos membros amitocondriados foram removidos para o Archaeprotista. Aqueles com mitocndrias achatadas e cristadas esto agora na Discomitochondriates, deixando aqui uma miscelnea de unicelulares.
ANIMALIA Do latim anima, respirao, alma
Organismos diploides que se desenvolvem de embries (blstulas) e que se formam por fuso (fertilizao: citogamia e cariogamia) de vulos e espermatozoides haploides (anisogametas). A meiose de gametas produz anisogametas.
Na classificao de dois reinos (animais e plantas) mais antiga e no usada neste livro os animais compostos de muitas clulas (multicelulares) eram referidos como Metazoa para distingui-los dos Protozoa (animais unicelulares). No nosso sistema, no h animais unicelulares: os protozorios tradicionais esto colocados no Reino Protoctista. Definimos animais como organismos heterotrficos, diploides, multicelulares, que normalmente (exceto as esponjas) se desenvolvem a partir de uma blstula. A blstula, um embrio multicelular que se desenvolve do zigoto diploide, produzido pela fertilizao de um grande vulo haploide por um pequeno espermatozoide haploide, nica aos animais. Os detalhes de desenvolvimento embrionrio posterior diferem largamente de filo para filo. Contudo, padres de desenvolvimento comuns proporcionam pistas para as relaes entre os filos. Em muitos filos, os detalhes do desenvolvimento so conhecidos para muito poucas espcies at o momento; em alguns filos, para nenhuma das espcies. A multicelularidade no nica aos animais; organismos multicelulares so abundantes em todos os reinos. Exemplos incluem a maioria das Cyanobacteria (Filo B-5) e Actinobacteria (Filo B-12) no Reino Bacteria; Phaeophyta (Filo Pr-17), Oomycota (Filo Pr-20) e Rhodophyta (Filo Pr-25) no Reino Protoctista; a maioria dos membros do Reino Fungi; e todos os membros do Reino Plantae. A maioria dos animais ingere nutrientes. Muitos animais levam o alimento para dentro de seus corpos atravs de uma abertura oral e ento os engolfam partculas slidas em clulas digestivas por fagocitose (comer clulas) , ou gotas de lquido por pinocitose (beber clulas), ou absorvem as molculas de alimento atravs de membranas celulares. Os parasitas, tais como os ortonectdeos (Filo A-8) e vermes-de-grdio (Filo A-11), frequentemente no possuem sistemas digestivos. Animais movidos a luz solar, tais como Convoluta paradoxa (um platelminto, Filo A-5) e Elysia (um molusco, Filo A-26), adquirem simbiontes, fotossintetizadores, como os proctotistas que se tornaram plantas (Filo Pr-28). De todos os organismos, somente os animais foram bem-sucedidos em invadir ativamente a atmosfera. Representantes de todos os cinco reinos (por exemplo, esporos de bactrias, fungos e plantas) gastam fraes significantes de seus ciclos de vida suspensos na atmosfera, mas nenhum em qualquer reino gasta toda sua historia de vida no ar. O vo ativo evoluiu somente nos animais. A locomoo dos animais atravs do ar evoluiu de forma independente diversas vezes, mas em somente dois filos: Mandibulata, classe Insecta, e Craniata, Classes Aves (aves), Mammalia (morcegos) e Reptilia (diversos dinossauros voadores extintos).
Por muitos anos, e mesmo agora, alguns bilogos associam animais a um de dois grandes grupos: os invertebrados animais sem espinhas dorsais, e os vertebrados animais com espinha dorsal. Todos os animais, exceto os membros do prprio filo, Craniata, so invertebrados. Hoje em dia, cerca de 98% do todos os animais vivos so invertebrados. Esta dicotomia, invertebrado-vertebrado, considera de forma significativa a nossa perspectiva distorcida. Nossos animais de estimao, bestas de cargas e fontes de alimento, couro e ossos isto , animais terrestres mais prximos do nosso tamanho e mais familiares so membros do nosso prprio filo. De um ponto de vista menos antropocntrico, os atributos que no h ausncia de uma espinha dorsal so melhores indicadores da divergncia evolutiva primitiva. Preferimos descrever estes animais, na maioria marinhos, por seus atributos nicos a diferencia-los coletivamente como invertebrados. Resumos amplos de sistema orgnicos que executam circulao, respirao, digesto, sustentao e reproduo sero discutidas na descrio de cada filo. Certos organismos tm circulao sangunea aberta em pelo menos parte dos seus corpos em vez de veias, artrias e capilares. Em outros, o sangue est confinado em artrias, capilares e veias, as quais so referidas como um sistema circulatrio fechado. Um sistema circulatrio transporta gases dissolvidos oxignio e dixido de carbono enquanto um sistema excretor funciona para livrar um organismo de rejeitos txicos como, por exemplo, algumas espcies so monoicas (uma casa, hermafrodita), tendo ambos os sexos em um nico organismo individual; outras so dioicas (duas casas), com organismos machos e fmeas separados. Os organismos monoicos podem ser hermafroditas simultneos ou hermafroditas sequenciais primeiro macho e depois fmea e depois macho. provvel que os filos animais, com exceo dos porferos, especialmente os eumetazorios, tiveram ancestrais entre os proctotistas.
A-1 Placcozoa (Trichoplax) Grego plakos, plano; zoion, animal Somente uma espcie, Trichoplax adhaerens, conhecida neste filo. Macia e to pequena a ponto de ser dificilmente visvel a olho nu. T. adhaerens est entre os animais mais simples.
A-2 Porifera (Esponjas, porferos) Latim porus, poro; ferre, portar
Os porferos, as esponjas, recebem esse nome por causa dos seus poros. A maioria das esponjas tem milhares de poros pelos quais a gua flui: assim suas formas corporais contribuem em grande parte para o fluxo de gua corrente induzido atravs de seus corpos.
A-3 Cnidaria (Cnidrios, hidras) Grego knide, urtiga As anmonas-do-mar, as guas vivas, as hidras e os corais entre as 9.400 espcies de cnidrios. Estes invertebrados radialmente simtricos so os membros com menos complexos morfologicamente do Sub-reino Eumetazoa, os metazorios verdadeiros.
A-4 Ctenophora (Gelias-de-pente) Grego kteis, pente; pherein, portar Os corpos dos ctenforos so flexveis e mveis, com a consistncia de uma gelia num saco membranoso. Placas de gelia com a forma de um remo (ctenas), nicas aos ctenforos, impulsionam estes invertebrados translcidos, com simetria birradial, atravs do mar.
A-5 Platyhelminthes (Vermes-chatos) Grego platys, chato; helmis, verme Os platelmintos so os vermes chatos em forma de fita e de folha. O corpo macio do verme chato tem simetria bilateral. As estruturas para capturar e consumir as presas esto localizadas na extremidade anterior, excetos nos vermes chatos turbelarianos, nos quais a boca est localizada ventralmente.
A-6 Gnathostomulida (Vermes-de-presa) Grego gnathos, mandbula; stoma, boca Os gnatostomuldeos so vermes-de-presa translcidos, caracterizados por presas nicas, dentadas, prximas s suas bocas ventrais. Estes vermes de vida livre esto em fungos, bactrias e protoctistas entre os gros da areia do mar.
A-7 Rhombozoa (Rombozorios) Grego rhombus, um topo torcido; zoion, animal Dois grupos de invertebrados Rhombozoa e Orthonectida eram designados antigamente como duas classes Rhombozoa (diciemdeos e heterociemdeos) e Orthonectida (ortonecndeos) dentro do Filo Mesozoa.
A-8 Orthonectida (Ortonectdeos) Grego orthos, reto; nektos, natao Os ortonecndeos, como os membros do Filo Rhombozoa, eram anteriormente considerados uma classe do Filo Mesozoa. Uma observao mais detalhada elevou os ortonectdeos ao status de filo, baseado em suas caractersticas nicas.
A-9 Nemertina (Vermes-de-fita, Nemertinos, Nemertes, Rinocelos) Grego Nemertes, uma ninfa do mar Nemertina um filo que consiste na maioria em vermes de vida livre encontrados em habitats marinhos, de gua doce e de solo. Seu nome comum de verme-de-fita se refere a seus corpos chatos e aos padres de cores brilhantes.
A-10 Nematoda (Nematdeos, vermes-de-cordo, vermes-redondos) Grego nema, fio Os nematdeos so vermes pseudocelomados no-segmentados, no percebidos at que eles chamem nossa ateno infestando nossas plantas, nossos animais ou a ns mesmos.
A-11 Nematomorpha nematomorfos) Grego nema, fio; morphe, forma
(Vermes-de-grdio,vermes-de-crina,
Os nematomorfos so comumente chamados de vermes -de-crina. Seus nomes de originam da crena de que estes vermes macios e cilindricos, observados em lavagens de cocheiras tinham origem nas crinas dos cavalos.
A-12 Acanthocephala (Vermes-de-cabea-espinhosa) Grego akantha, espinho; kephale, cabea Os vermes-de-cabea-espinhosa (vermes-de-espinho), ou acantocfalos, so parasitas que vivem no intestino de vertebrados e no seu ciclo de vida inicial dentro de invertebrados. A probscide protrtil com espinhos globular ou cilndrica; o corpo tambm pode conter espinhos pontudos.
A-13 Rotifera (Rotferos) Latim rota, roda; ferre, portar O rotfero um animal aqutico diminuto, cujo nome provm de uma iluso de tica as ondas dos clios batendo na sua cabea parece uma roda em rotao. Os clios tm um duplo propsito: propelem o retfero na natao e dirigem correntes de alimentos para sua boca.
A-14 Kinorhyncha (Quinorrincos) Grego kinein, mover; rynchos, tromba Os quinorrincos so animais marinhos de vida livre, geralmente com 1mm ou menos, um tanto quanto maiores do que os rotferos e os gastrotrqueos.
A-15 Priapulida (Priapuldeos) Latim priapulus, pequeno pnis Os priapuldeos so vermes curtos, gordos e exclusivamente marinhos. A probscide dos priapuldeos termina numa boca e inverte medida que ela se retrai, tal que a probscide (pressoma) chamada de invaginao.
A-16 Gastrotricha (Gastrticos) Grego gaster, estmago; thrix, plo Cristas, espinhos ou escamas ornamentam as costas e as laterais dos gastrotrqueos, mas no suas barrigas. Este filo deriva seu nome dos clios que cobrem suas barrigas o gastrotrqueo composto das palavras gregas para estmago peludo.
A-17 Loricifera (Loricferos) Latim loricus, colete, cinte; fero, carregar, portar Um loricfero um animal microscpico com uma boca cnica um tubo de boca anterior flexvel -, uma cabea contendo espinhos em forma de taco e em forma de garra, um pescoo (trax) e uma cinta de placas chamadas de lorica que cobrem o abdmen.
A-18 Entoprocta (Entoproctos) Grego entos, dentro; proctus, nus Os entoproctos so animais transparentes, pequeninos principalmente marinhos. A maioria sssil, vivendo em colnias permanente e firmemente anexadas por hastes, estolhos horizontais e discos basais a rochas, empilhamentos, conchas, algas ou outros animais.
A-19 Chelicerata (Quelicerados) Grego cheli, garra Os quelicerados, que chegam ao nmero aproximadamente 75.000 espcies, incluem trs classes: os caranguejos-ferradura, as aranhas-do-mar e as aranhas, escorpies, caros, carrapatos, micuins e ceifeiros, tambm chamados de papai-de-pernas-longas.
A-20 Mandibulata (Uniramia) (Mandibulados, artrpodes mandibulados) Latim mandere, mastigar; mandibulum, uma mandbula De mais de 30 milhes de espcies de animais que se estima existirem agora (somente cerca de 10 milhes das quais tem sido formalmente descritas e nomeadas), o Filo Mandibulata reivindica o maior nmero.
A-21 Crustacea (Crustceos) Latim crustaceus, que possui uma concha ou crosta A caracterstica que define o Filo Crsutacea possuir dois pares de antenas na cabea, distinguindo os crustceos de todos os outros artrpodes.
A-22 Annelida (Vermes aneldeos) Latim annellus, pequeno anel Os vermes aneldeos poliquetas, minhocas (oligoquetas) e sanguessugas (hirudneos) so distinguidos por sries lineares de segmentos externos em forma de anel; as fendas entre os segmentos coincidem com os compartimentos internos, frequentemente separados por bainhas transversais de tecido (septos), contendo sries repetidas de nervos, msculos e sistemas excretores.
A-23 Sipuncula (Sipnculos, sipunculdeos, vermes-amendoim) Latim siphunculus, pequeno tubo Os sipnculos so animais marinhos no-segmentados, habitantes de fendas e escavadores. A invaginao contrtil varia em tamanho de menos de metade do comprimento do tronco at vrias vezes o comprimento do tronco em algumas espcies.
A-24 Echiura (Vermes-colher, equiros, equiurdeos) Grego echis, serpente; latim ura, com cauda Cerca de 140 espcies de equiros conhecidos so denominados por suas probscides flexveis, que podem estender-se at cerca de 1,5 m de seus corpos macios e arredondados.
A-25 Pogonophora (Vermes-de-barba, pogonforos, vermes-tudo) Grego pogon, barba; pherein, portar Os pogonforos, os vermes-de-barba, so vermes marinhos bentnicos, que vivem em tubos de quitina eretos, fixos, que eles secretam nos sedimentos, conchas ou madeira em decomposio no fundo do oceano.
A-26 Mollusca (Moluscos) Latim molluscus, macio
Do nutilus da cmera propelido a jato (mas de movimento bastante lento) at a clula de cone venenosa, a diversidade dentro dos moluscos notvel.
A-27 Tardigrada (Ursos-dgua, tardgrados) Latim tardus, lento; gradus, passo Devido a locomoo por patas dos tardgrafos, o naturalista ingls do sculo dezenove Thomas Huxley os denominou ursos-dgua, um nome pomposo.
A-28 Onychophora ( Vermes-veludo, unicforos, perpatos) Grego onyx, garra; pherein, portar Os onicforos so comumente conhecidos como vermes-veludo: suas cutculas so ornamentadas com pequeninos ressaltos que tem a consistncia de veludo.
A-29 Bryozoa (Ectoprocta, ectoproctos, animais-musgo) Grego bryon, musgo; zoion, animal; ektos, exterior; proktos, nus Os briozorios, tambm chamados de ectoproctos ou animais de musgo, so primordialmente filtradores ssseis. Como os branquipodes e os forondeos, os briozorios tm rgos com tentculos chamados de lifforos preenchidos com fluido.
A-30 Brachiopoda (Conchas-de-lmpada(lampshells), braquipodes) Latim brachium, brao; grego pous, p Os branquipodes so chamados de conchas-de-lmpada porque se assemelham s antigas lamparinas a leo. Como os mariscos e outros moluscos bivalves, o branquepode tem duas conchas duras opostas (vlvulas).
A-31 Phoronida (Forondeos) Grego pherein, portar; latim nidus, ninho
Todos os forondeos so vermes marinhos sedentrios, raros na maioria dos locais, mas encontrados em todo o mundo. O forondeo, um lofoforado, carrega uma crina em forma de ferradura ou de mola espiralada, com tentculos ciliados, em torno de sua boca.
A-32 Chaetognatha (Vermes-de-seta) Grego chaite, cabelo; gnathos, mandbula O nome vernacular verme-de-seta frequentemente do aos quetognatos devido aos seus corpos em forma de seta. A parte queto de seus nomes se refere a seus ganchos mveis, com os quais os quetgnatos seguram presas vivas.
A-33 Hemichordata (Vermes-bolota, pterobrnquios, enteropneustos, vermes-delngua) Grego hemi-, metade; latim chorda, cordo Os hemicordados so celomados pequenos, de corpos macios, caracterizados por uma probscide, um colar e um tronco com fendas branquiais.
A-34 Echinodermata (Equinodermos) Grego echinos, urtiga-do-mar; derma, pele Os adultos equinodermos so invertebrados de simetria radial, com trs caractersticas nicas: simetria de cinco partes; um esqueleto interno de carbono de clcio; e um celoma distintamente dividido, incluindo um sistema vascular de gua.
A-35 Urochordata (Tunicados, seringas-do-mar, ascdias, larvceos, salpas) Grego oura, cauda; latim chorda, cordo Os tunicados so considerados cordados acraniados, porque os tunicados e os cordados te as seguintes caractersticas em comum: uma notocorda; um cordo nervoso oco dorsal; fendas branquiais faringianas em algum perodo de suas vidas.
A-36 Cephalochordata (Anfioxo, lanceolados Acrania) Grego cephalo, cabea, latim chorda, corda Os cefalocordados, como os tunicados, so cordados acraniados, isto , os cordados que no possuem um crnio. Os cefalocordados variam de cerca de 5 a 15 cm de comprimento e vivem em fundos de mar arenosos e rasos.
A-37 Craniata Grego kranion, crebro Os membros deste filo, o nosso prprio filo, so os mais familiares de todos os animais. Os craniados incluem cerca de 45.000 espcies, incluindo a maioria dos animais de importncia econmica direta, exceto moluscos e artrpodes.
O REINO FUNGI Latim fungus, provavelmente do grego sp(h)ongos, esponja
Conjugao (fuso de hifas ou celular) de osmtrofos dicariticos ou haplides (monocariticos) que se desenvolvem de esporos fngicos resistentes sem motilidade. As clulas, incluindo os esporos, tm paredes quitinosas. Os esporos so produzidos por mitose ou meios zigtica. Undulipdios (quinetossomos e axonemas) inexistente em todos os estgios. Equivalente aos Reinos Mychota ou Eufungi. Os registros fsseis se estendem desde a era Paleozica inferior (450 milhes de anos atrs) at o presente.
O Reino Fungi, est limitado aos eucariotas que formam propgulos quitinosos resistentes (esporos fngicos) e paredes celulares quitinosas, e que no possuem undulipdios (isto , so amastigotas ou imveis) em todos os estgios do seu ciclo de vida. Das 1.500.000 espcies de fungos que se estima existirem, cerca de 60.000 foram descritas; a maioria terrestre, embora umas poucas espcies verdadeiramente marinhas sejam conhecidas. Devido aos fungos muitas vezes diferirem somente em caractersticas sutis, tais como detalhes da estrutura, dos pigmentos e de complexos compostos orgnicos, provvel que muitos ainda no tenham sido reconhecidos como espcies distintas.
Os fungos foram tradicionalmente colocados com as plantas e, alguns esquemas de classificao anteriores, os consideravam como um sub-reino do Reino Plantae. Contudo, os fungos esto claramente mais prximos dos animais do que das plantas. Em qualquer classificao, os fungos diferem dos ciclos de vida dos animais e das plantas, do modo de nutrio, do padro de desenvolvimento e de muitas outras formas. Portanto, concordamos com muitos mitlogos que os fungos constituem seu prprio reino. Os fungos no possuem estgio embrionrio e se desenvolvem diretamente de esporos. Frequentemente nascidos em esporngios, os esporos podem ser de origem mittica ou meitica. A reproduo sexual dos fungos por conjugao, na qual as hifas dos tipos sexuais complementares se unem e se fundem. Um fungo pai pode ter hifas de dois tipos diferentes de acasalamento. Quase todos os fungos so aerbios, e todos eles so hetertrofos que caracteristicamente absorvem seu alimento. Eles excretam enzimas poderosas que decompem o alimento fora dos fungos em molculas menores; os nutrientes dissolvidos so ento transportados para o fungo atravs da membrana Fonseca. Os fungos so persistentes, resistindo a dessecao severa e outros desafios ambientais. Suas paredes celulares, composta de quitina polissacardea nitrogenada, so duras e resistentes, e suportam perda de gua. Muitos fungos causam doenas, especialmente em animais, incluindo humanos, e em plantas. Contudo, muitos formam associaes ntimas e construtivas com as plantas. Muitos fungos produzem substncias teis aos humanos. Alguns fungos so fontes de cidos ntricos e produtos farmacuticos. Milhares de outros antibiticos tm sido acrescentados penicilina, que um antibitico produzido naturalmente pelo mofo Penicillium chrysogenum. Os fungos adicionam sabor, cor, contedo proteico e preservam qualidades quando usados na produo de bebidas e alimento.
F-1 Zygomycota (Zygomycotes, zygomycetes) Grego zigon, par; mikes, fungo Os membros deste filo no possuem paredes de separao (septos), exceto para separao entre as estruturas reprodutivas e o resto do miclio.
F-2 Basidiomycota (Basidiomycotes, basidiomycetes) Grego basidion, base pequena; mikes, fungo Os basidiomicetos incluem a fuligem, a ferrugem, os fungos gelatinosos, os cogumelos, os bufa-de-lobo e os stinkhorns. O basdio, uma estrutura reprodutiva microscpica em forma de basto, da qual seu nome originrio, distingue os basiomicetos dos fungos nos outros filos.
F-3 Ascomycota (Ascomycotas, ascomecetos) Grego askos, vescula, mykes, fungo Conhecidos como leveduras, os fungos azul-esverdeados, morchelas, trufas, liquens e os antigos deuteromicetos, os ascomicetos so um grupo grande, diverso e economicamente importante de fungos. Cerca de 30.000 espcies so conhecidas.
REINO PLANTAE
PLANTAE Do latim planta, planta
Organismos haploides de sexos complementares crescem de esporos produzidos pela meiose (meiose esporognica) que ocorre no diploide adulto. Estes haploides produzem gametas por mitose. A fertilizao por espermatozoides (citogamia e cariogamia) ou ncleo do plen (cariogamia) conduz a embries diploides retidos pelo organismo haploide feminino durante o desenvolvimento inicial. Os registros fsseis se estendem desde a Era Paleozoica Inferior (450 milhes de anos atrs) at o presente. Os membros do reino das plantas se desenvolvem de embries estruturas multicelulares envolvidas em tecido maternal. Devido a todas as plantasformarem embries, elas so todas multicelulares. Alm disso, devido aos embries serem os produtos da fuso sexual de clulas, todas as plantas potencialmente tm um estgio sexual em seus ciclos de vida (embora isso no seja sempre realidade). No estgio sexual, o gameta masculino (ncleo espermtico, haploide) fertiliza o gameta feminino (vulo, ou ncleo do saco embrionrio, haploide). Muitas plantas crescem e se reproduzem de modos que sobrepassam a fuso sexual binria todas devem ter evoludo de ancestrais
que formavam embries por fuso celular sexuada. Todas as plantas so compostas de clulas eucariotas, muitas com plastdios verdes. Distinguimos as plantas de todos os outros organismos mais por seus ciclos de vida do que por sua capacidade de fotossntese, porque algumas plantas (gotas -de-faia, Epifagus, por exemplo) no executam a fotossntese durante toda a sua existncia. Todas as plantas que fotossintetizam produzem oxignio (em comparao, nas espcies procariotas fotossintticas as enzimas so ligadas como cromotforos s membranas celulares, e no empacotadas separadamente; os padres procariotas da fotossntese anaerbica e aerbica incluem a formao de produtos finais como enxofre, os sulfatos e o oxignio). As plantas so os organismos mais responsveis, na terra e em ambientes marinhos rasos, pela transformao da energia solar, da gua e do dixido de carbono em produtos primrios: alimentos, fibras, carvo, leo, madeira e outras formas de armazenamentos de energia (no oceano aberto, os protoctistas planctnicos so os produtores primrios). Dois grandes grupos as plantas avasculares (informalmente chamadas de brifitas, e tambm chamadas de Bryata) e as plantas vasculares (Traquefitas) constituem o reino das plantas. As espcies de traquefitas, as familiares plantas lenhosas e herbceas, so distinguidas por sistemas vasculares tecidos condutores lignificados, chamados de xilema e floema. As plantas vasculares podem ser agrupadas em plantas vasculares portadoras de sementes (Filos Pl-8 at Pl-12) e no-portadoras de sementes (Filos Pl-4 at Pl-7). A grande maioria das plantas existentes atualmente so traquefitas que pertencem ao Filo Anthophyta, as plantas com flores (Filo Pl-12). O nmero atual de gimnospermas (do grego gymno, nu; sperma, semente) de cerca de 720 espcies em 65 gneros, comparados com aproximadamente 240.000 espcies de plantas com flores. Quatro filos de plantas vasculares no produzem sementes: os licopdios (Filo Pl-4), os fetos-espanador(Filo Pl-5), as cavalinhas (Filo Pl-6) e os fetos (Filo Pl-7). Todas se reproduzem com esporos. A disperso de esporos em hepticas, antocerfitas e cavalinhas auxiliada por elatrios (do grego, diretor) estruturas oblongas que se formam dentro dos esporngios. O esporngio um rgo no qual as clulas sofrem meiose e produzem esporos (haploides). Um esporo uma clula reprodutiva capaz de se desenvolver numa planta madura sem se fundir com outra clula; em comparao, um vulo ou gro de plen se funde com sua clula reprodutiva complementar para produzir uma nova planta. As plantas vasculares no portadoras de semente desenvolveram um grande conjunto de estruturas foliais. Chamamos os trs filos de plantas avasculares (Pl-1 at Pl-3) de Bryata musgos, hepticas e antocerfitas. As opinies variam em relao a Takakia, considerada um musgo por alguns, incluindo ns, e um filo separado (ou
diviso) por outros. Todas as plantas avasculares tm um talo um corpo vegetal sem folhas verdadeiras, caule ou razes. Como elas no possuem razes nem sistema vasculares verdadeiros, obtm umidade e nutrientes do ambiente por difuso direta atravs de seus tecidos. Dentro de seus corpos, a difuso, a ao capilar e a corrente citoplasmtica conduzem os fluidos. Os musgos tambm tem clulas condutoras chamadas de leptoides e hidroides, mas estas so no-lignificadas. Os hidroides so clulas alongadas que no possuem citoplasma vivo na sua maturidade. As suas paredes de extremidades finais so muito permeveis aos solutos e gua. As clulas leptoides coondutoras de nutrientes circundam as hidroides em alguns musgos. Filamentos delicados uni- ou multicelulares (rizoides) ancoram as plantas avasculares ao solo, rochas ou cascas de rvores. As licfitas e as psilfitas tm sido colocadas como representantes vivos da primeira diviso das linhagens primitivas de plantas terrestres vasculares. Estudos de DNA de cloroplastos tendem confirmar a evidncia geolgica de que os licopdios so parentes mais prximos das plantas avasculares (Filos Pl-1 at;e Pl-3), enquanto as psilfitas so parentes mais prximos das plantas vasculares (Filos Pl-6 at Pl-12) do que dos licopdios. Se parece que h muito menos grupos de plantas do que grupos de animais, isto parcialmente devido aos txons animais e vegetais serem definidos por critrios morfolgicos, e a diversidade da anatomia interna e externa mais extensa nos animais do que nas plantas. As diferenas entre muitas plantas so sutis, frequentemente envolvendo distines qumicas. As plantas produzem muitos compostos qumicos que so metablitos secundrios usados na defesa das plantas contra os fungos, animais e outras plantas, e por isso so exigidos somente indiretamente para a sobrevivncia e a reproduo. Os compostos secundrios incluem toxinas, compostos psicoativos com as alcaloides da maconha e venenos respiratrios como o cianeto todos os quais impedem que os predadores comam a planta. Todas as plantas se desenvolvem de embries, organismos jovens multicelulares diploides sustentados por tecidos estreis ou no-reprodutivos; nas conferas, gnetfitas e angiospermas, o cotildone (folha embrionria de semente) proporciona os nutrientes ao jovem embrio. O cotildone das monocotiledneas absorve alimentos, enquanto os cotildones das dicotiledneas armazenam alimentos. Os ciclos de vida de todas as plantas avasculares so dominados pelas notveis gametfitas verdes (haploides), exemplificadas por um manto verde de musgo. Para seguir o ciclo de vida dos musgos, em termos simples, um exemplo do ciclo de vida vegetal em geral, comeamos com a reproduo sexuada. O rgo reprodutivo masculino o anterdio produz anterozoides com um par de undulipdios apontando para frente. O rgo reprodutivo
feminino o arquegnio produz uma oosfera. Os anterozoides dos musgos so dispersados do gametfito masculino maduro pelo choque de gotas de chuva. Numa vista amplificada do arquegnio, os espermatozoides podem ser vistos nadando em direo oosfera. Os espermatozoides das cicadceas, gincos, gnetfitas, conferas e angiospermas so carregados para o vulo num tubo de plen, formando aps a germinao do gro de plen. O ncleo do tudo do gro de plen maduro dirige o crescimento do tudo para o vulo. Nas angiospermas o tubo cresce atravs do estigma e do estilete. O tubo do plen proporciona um ambiente mido para o espermatozoide; assim sua evoluo eliminou a necessidade do ambiente aqutico durante a fertilizao. L dentro do arquegnio e ainda no gametfito feminino a fertilizao do vulo pelo espermatozoide acontece. A fertilizao restaura o diploidismo (a condio 2n) e inicia o desenvolvimento do zigoto no embrio. O embrio de desenvolve no esporfito diploide, que emerge do arquegnio. O jovem esporfito obtm nutrientes do gametfito feminino, no qual ele se aninha permanentemente. As plantas de sementes existentes que produzem gametas no-mveis, em comparao com os tubos de plen, constituem o fim de uma srie que se estende desde os musgos, as hepticas, as antocerfitas, os licopdios, as cavalinhas, as psilfitas e os fetos todos tendo gametas nadadores passando pelos gincos e as cicadceas, que tm tubos de plen assim como gametas mveis, at as conferas, as gnetfitas e as plantas de flores, com tubos de plen como condutores para gametas masculinos no-mveis. As histrias da vida das plantas so to elegantemente diversas quanto suas formas e cores.
Pl-1 Bryophyta (Musgos) Do grego bryon, musgo; phyton, planta As plantas avasculares, informalmente chamadas de brifitas, so as hepticas, as antocerficas, Takakya e os musgos, mas somente musgos e Takakya esto agora includos no Filo Bryophita.
Pl-2 Hepatophyta (Heptica) Do grego hepat, fgado; phyton, planta Os espcimes do Filo Hepatophyta so comumente chamados de hepticas um termo derivado da forma de fgado de seus gametfitos.
Pl-3 Anthocerophyta (Antocerfitas)
Do grego anthos, flor; keros, cera; phyton, planta Os espcimes do Filo Anthocerophyta, chamados de hepticas chifrudas, ou simplesmente antocerfitas, derivam seu nome comum do esporfito alongado em forma de chifre que est encaixado no gametfito por um p.
Pl-4 Lycophyta (Musgos-de-basto, licfitas, licopdios) Grego lykos, lobo; phyton, planta Os espcimes de locfitas licopdios, musgos-de-prego e plantas-pena so relictos (sobreviventes) de um glorioso passado de 400 milhes de anos de idade.
Pl-5 Psilophyta (Psilfitas,feto-espanador) Do grego psilo, nu; phyton, planta As espcimes de psilfitas, Psilotum e Tmesipteris, so nicas entre as plantas vasculares sem semente. Elas constituem o nico filo de plantas vasculares que como as hepticas, antocerfitas e musgos avasculares no possuem nem razes nem folhas.
Pl-6 Sphenophyta (Esfenfitas, equisetfita, cavalinhas) Do grego sphen, cunha; phyton, planta A cavalinha comum pertence s esfenofitas de plantas vasculares sem sementes, facilmente reconhecidas por seus caules ocos e articulados com costelas speras.
Pl-7 Filicinophyta (Pterophyta, Pterodatina, pteridfitas, fetos) Do latim felix, feto; do grego phyton, planta; e pteridium, asa pequena, pena Os fetos so plantas vasculares sem sementes que, como as brifitas, as psilofitas, as licfitas e as esfenfitas, reproduzem-se e dispersam-se atravs de esporos.
Pl-8 Cycadophyta (Cicadcias)
Do grego kikos, uma palmeira; phyton, planta Algumas cicadceas so pequenos arbustos, como Zamia, que tem cerca de 0,3m de altura, enquanto outras, como Cycas e Microcycas, so rvores do tipo de palmeira com mais de 18m de altura.
Pl-9 Ginkgophyta Do japons ginkyo, damasco prateado; do grego phyton, planta A rvore ginco, Ginkgo biloba, [e o nico gnero e espcie do Filo Ginkgophyta, um filo de plantas e sementes vasculares. Em nmero de espcies (uma), este o menor filo vegetal.
Pl-10 Coniferophyta (Conferas) Do latim conus, cone; ferre, portar; do grego phyton, planta A maioria dos gminospermas de cone so rvores, embora alguns sejam arbustos e conferas procumbentes arrepiadas. Como em todas as gminospermas, o esporfito da confera a gerao notvel, enquanto o gametfito menor e nutricionalmente dependente do esporfito.
Pl-11 Gnetophyta (Gnetfitos) Do latim gnetum, do danemu malsio das ilhas molucanas, uma espcie gnetfita encontrada na ilha de Ternate; do grego phyton, planta As gnetfitas so plantas de semente vasculares, distintas de outras gimnospermas; elas tem tubos longos condutores de gua chamados de vasos.
Pl-12 Anthophyta (Angiospermas, magnoliceas, plantas florferas) Do grego anthos, flor; phyton, planta As plantas florescentes, ou de flores, as angiospermas, so as superestrelas da diversidade e da abundncia. Mais de 230.000 espcies de angiospermas esto agrupadas em cerca de 350 familias.
QUESTIONRIO
1. Quais so os fatores que fazer as bactrias serem as mais resistentes dos seres vivos? As bactrias so os mais resistentes dos seres vivos. Algumas podem sobreviver a temperaturas muito baixas, bem abaixo do congelamento, por anos; outras vicejam em fontes de gua fervente; e outras ainda, crescem at em cidos muito fortes (hot acid), ou vivem retirando hidrognio e dixido de carbono das rochas.
2. Como os protoctistas se desenvolvem? Todos os protoctistas se desenvolveram por simbiose dentre pelo menos dois tipos diferentes de bactrias em alguns casos, dentre muito mais do que dois.
3. O que blstula? A blstula, um embrio multicelular que se desenvolve do zigoto diploide, produzido pela fertilizao de um grande vulo haploide por um pequeno espermatozoide haploide, nica aos animais.
4. Descreva o que so os eucariotas. Est limitado aos eucariotas que formam propgulos quitinosos resistentes (esporos fngicos) e paredes celulares quitinosas, e que no possuem undulipdios (isto , so amastigotas ou imveis) em todos os estgios do seu ciclo de vida.
5. Descreva o estgio sexual do reino das plantas.
No estgio sexual, o gameta masculino (ncleo espermtico, haploide) fertiliza o gameta feminino (vulo, ou ncleo do saco embrionrio, haploide). Muitas plantas crescem e se reproduzem de modos que sobrepassam a fuso sexual binria todas devem ter evoludo de ancestrais que formavam embries por fuso celular sexuada.
11. RAVEN, P. H.; EVERT R. F.; EICHHORN, S. E. Biologia Vegetal. 7. ed. Rio de Janeiro: Guanabara Koogan, 2007. sees 4, 5, 6 e 7. SEO 4 Diversidade Sistemtica A sistemtica - estudo cientfico da diversidade biolgica - compreende a taxonomia (a identificao), a nomenclatura, a classificao das espcies e a filogentica (o entendimento das inter-relaes evolutivas entre os organismos). Os nomes cientficos obedecem a uma nomenclatura binomial, ou seja, a primeira palavra no binmio corresponde ao gnero; a segunda palavra, que corresponde ao epteto especfico, combinada ao nome do gnero, completa o nome da espcie. Os gneros so agrupados em famlias; as famlias em ordens; as ordens em classes; as classes em fils e os fils em reinos. Baseando-se em estudos taxonmicos recentes, os reinos so agrupados em domnios. Os organismos so classificados filogeneticamente usando-se caracteres homlogos, em vez de anlogos. Novas tcnicas em sistemtica molecular esto fornecendo um mtodo, relativamente objetivo e explcito, para comparar organismos, o gene. Os estudos obtidos a partir de sequncias da pequena subunidade do RNA ribossmico, agrupam os organismos vivos em trs domnios:, Bactria, Archaea e Eukarya. Bactria e Archaea so duas linhagens distintas de organismos procariticos. Archaea mais estreitamente relacionada com eukarya do que com Bactria. Eukarya consiste inteiramente em eucariotos. Protista, Fungi, Plantae e Animlia so reinos dentro de Eukarya.
Procariotos e vrus
Os procariotos so muito simples estruturalmente. Em termos evolutivos, eles so tambm os mais antigos organismos da Terra e consistem em duas linhagens distintas, os domnios Bactria e Archaea. Os procariotos no possuem ncleo organizado nem organelas celulares envoltas por membranas. A maior parte do seu material gentico est incorporada em uma nica
molcula circular de DNA de fita dupla, que se replica antes da diviso celular. Frequentemente, fragmentos adicionais do DNA circular, conhecidos como plasmdeos, tambm esto presentes. Com exceo dos micoplasmas, todos os procariotos tm parede celular rgidas. Nas Bactria, esta parede celular composta principalmente de peptidioglicanos. As bactrias Gram-negativas, cuja parede celular no fixa o corante violeta cristal, possuem uma camada externa de lipopolissacardios e protenas sobre a camada de peptideoglicano. Muitos procariotos secretam substncias mucilaginosas ou gelatinosas na superfcie de suas paredes celulares, formando uma camada denominada glicoclice ou cpsula. Muitos procariotos se reproduzem por diviso binria. Mutaes, combinadas ao curto tempo de gerao dos procariotos, so responsveis por sua alta adaptabilidade. A recombinao gentica pode ocorrer atravs de conjugao, transformao e transduo. Certas bactrias tm a capacidade de formar endsporos. Alguns procariotos so auttrofos, a maioria hetertrofa. Os procariotos saprfitas, juntamente com os fungos, so os recicladores da biosfera. Os auttrofos podem ser fotossintetizantes ou quimiossintetizantes. De todos os organismos vivos, somente certas bactrias conseguem fixar o nitrognio. Sem as bactrias, a vida na Terra, como ns a conhecemos, no seria possvel. Os procariotos podem ser aerbios, anaerbios estritos, ou ainda, anaerbios facultativos. Podem ser encontrados a O C ou menos (psicrfilos), at os que se desenvolvem em temperaturas acima de 100 C (termfilos extremos). Muitas bactrias so patgenos importantes, tanto de plantas quanto de animais. Bactrias fotossintetizantes podem ser divididas em trs grupos principais: as cianobactrias, as proclorfitas e as bactrias purpreas e verdes. Muitos gneros de cianobactrias podem fixar nitrognio. Isto leva a crer que as Bactria esto envolvidas na origem simbitica dos cloroplastos. No incio da histria dos eucariotos, um fato simbitico similar a este ocorrido na origem dos cloroplastos parece ter dado origem s mitocndrias. As Archaea tambm podem ser divididas em trs grandes grupos: halfilos extremos, metanognicos e termfilos extremos. Um quarto grupo representado por um nico gnero, Thermoplasma, que no possui parede celular. Acreditava-se que as Archaea ocupavam, primariamente, ambientes hostis; hoje, sabe-se que constituem os principais componentes do picoplncton (organismos com menos de 1 micrometro) ocenico. Os vrus so estruturas no-celulares, constitudas de DNA ou RNA, envoltas por uma capa de protena. Os vrus possuem genomas que se replicam dentro de um hospedeiro vivo, direcionando o mecanismo gentico da clula para a sntese de cidos nuclicos e protenas virais. Os vrus so responsveis por causar diversas doenas em humanos e outros animais, e ainda so responsveis pela causa de mais de 2.000 tipos
diferentes de doenas em plantas. A transmisso dos vrus, de plantas doentes para sadias, frequentemente envolve insetos vetores. Uma vez dentro da clula hospedeira, a partcula viral, ou vrion, desprende seu capsdeo, liberando o cido nuclico. A maioria dos vrus de plantas constituda de RNA.
Fungos
Os fungos, juntamente com as bactrias heterotrficas, so os principais decompositores da biosfera, quebrando a matria orgnica e reciclando o carbono, o nitrognio e outros componentes, que so liberados no solo e no ar. A maioria dos fungos saprbia. Muitos fungos atacam organismos vivos e so agentes causadores de doenas em plantas, animais domsticos e no homem. O reino Fungi inclui as leveduras, que so fungos utilizados na fabricao de queijos, os cogumelos comestveis, Penicillium e outros produtores de antibiticos. So organismos que crescem rapidamente e produzem filamentos chamados hifas, classificadas em septadas e asseptadas. Na maioria dos fungos, as hifas formam miclio. Fungos parasitas apresentam haustrios (hifas especializadas), que extraem nutrientes das clulas vivas de outro organismo. Os fungos reproduzem-se por esporos; secretam enzimas sobre a fonte de alimento e ento absorvem as molculas liberadas. O principal componente da parede de um fungo a quitina. Glicognio o principal polissacardeo de reserva. Os fungos so isgamos (gametas similares na forma e no tamanho); a meiose zigtica. O reino Fungi inclui quatro fils: Chytridiomycota, Zygomycota, Ascomycota e Basidiomycota, bem como um grupo artificial conhecido como deuteromicetos. H muitas evidncias de que animais e fungos divergiram a partir de um ancestral comum, representado por um protista colonial. Os Chytridiomycota so principalmente aquticos. Formam o nico grupo de fungos com clulas reprodutivas mveis, as quitrdias, em geral hifas cenocticas, com poucos septos maturidade. Algumas espcies so parasitas, outras so saprbias. Muitas so fitopatgenas. Os Zygomycota ou zigomicetos tem miclio cenoctico. Os esporos so produzidos em esporngios. Os zigomicetos so assim chamados porque, a partir da reproduo sexuada, formam esporos de resistncia denominados zigsporos. Estes desenvolvem-se dentro de estruturas com parede espessa chamadas zigosporngios. No filo /Ascomycofa, comumente denominados ascomicetos, h cerca de 32.300 espcies. Em nenhum outro grupo encontram-se tantas espcies como neste. A caracterstica distintiva so os ascos, encontrados dentro de estruturas chamadas ascomas (antigamente conhecidas como ascocarpos). No
filo Basidiomycota encontram-se muitos fungos macroscpicos e bem familiares. Incluem-se os cogumelos comestveis, chapus-de-sapo, orelhasde-pau e outros; bem como as ferrugens e carves, estes importantes fitopatgenos. A caracterstica distintiva do filo a produo de basdios. Cada um destes produz quatro basidisporos. As leveduras so fungos unicelulares que, tipicamente, se reproduzem por brotamento, um mtodo assexuado de reproduo. A maioria das leveduras do grupo dos ascomicetos, e se reproduzem sexuadamente, por intermdio da produo de ascsporos. Os deuteromicetos ou Fungi Imperfecti, incluem milhares de espcies sem ciclo sexuado conhecido. Os lquens (micobionte mais fotobionte), so associaes simbiontes e mutualsticas entre um fungo e uma populao de algas verdes ou de cianobactrias. 98% dos fungos dessa associao pertencem aos ascomicetos. O restante pertence aos basidiomicetos. As micorrizas so associaes mutualsticas entre fungos e razes.
Protista l - Euglenofceas, mixomicetos, criptofceas, algas vermelhas, dinoflagelados e haptfitas
So considerados protistas os organismos fotossintetizantes (auttrofos), as algas, e os organismos heterotrficos, antigamente tratados por fungos. Nos ltimos incluem-se os Myxomycota e Dictyosteliomycota, bem como os Oomycota. Nas algas incluem-se os protistas fotossintetizantes bem como organismos incolores relacionados, que correspondem a importantes componentes da cadeia alimentar aqutica e fazem parte, junto com as cianobactrias, do fitoplncton. Os mixomicetos, filo Myxomycota, exibem massas de protoplasma deslizantes e multinucleadas chamadas plasmdios, comumente diploide. Esses plasmdios formam esporngios com esporos diploides em seu interior. Cada esporo desses sofre meiose; trs, dos quatro ncleos resultantes, degeneram e apenas um esporo com ncleo haploide permanece. Em condies favorveis, os esporos germinam produzindo amebas que podem tornar-se flageladas. Amebas e clulas flageladas podem funcionar como gametas. Os dictiostelidas, filo Dictiosteliomycota, so organismos ameboides que, em determinada fase do ciclo da vida, agregam-se formando pseudoplasmdios. Clulas flageladas no so conhecidas. A reproduo assexuada por esporos
comum. Ao contrrio dos fungos, mas semelhantes a muitos outros protistas, os mixomicetos e os dictiostelidas alimentam-se por fagocitose. As Algas absorvem nutrientes de modos diferentes. Embora muitas delas sejam capazes de realizar a fotossntese, a utilizao de compostos orgnicos dissolvidos tambm comum. A fagocitose ocorre em alguns membros de euglenofceas, criptofceas, dinoflagelados e haptfitas. Aproximadamente um tero das euglenofceas so fotossintetizantes e possuem cloroplastos, que contem clorofilas a e b e diversos carotenoides. Grnulos de paramido so estocados no citoplasma. As clulas contm um vacolo contrctil e flagelos, A reproduo sexuada desconhecida. As criptofceas so indivduos unicelulares flagelados que parecem ter surgido da fuso de duas diferentes clulas eucariticas, uma heterotrfica e outra fotossintetizante. Alm das clorofilas a e c e carotenoides, alguns cloroplastos de criptofceas contem ficocianina ou ficoeritrina, pigmentos solveis em gua, encontradas somente em cianobactrias e algas vermelhas. Os dinoflagelados so biflagelados e unicelulares. Muitos apresentam placas de celulose formando a parede ou teca. Outros no apresentam placas celulsicas, ou estas so muitos finas. Aproximadamente metade dos dinoflagelados fotossintetizante, possuindo clorofilas a e c. Alguns dinoflagelados marinhos produzem compostos txicos, como os que provocam a mar vermelha (Gonyaulax sp.). Outros produzem alimento na endossimbiose, com clulas de corais formadores de recifes. As algas vermelhas, Filo Rhodophyta, formam um vasto grupo, facilmente encontrado em ambiente marinho (tropical). Geralmente, crescem fixas ao substrato e, s vezes, em grandes profundidades. Bioquimicamente, os cloroplastos so muito semelhantes s cianobactrias, das quais eles provavelmente derivaram. Tambm so fontes de carboidratos teis e valiosos, tais como o gar. As haptfitas, Filo Haptophyta, so encontradas com maior frequncia no fitoplncton marinho. Sua caracterstica mais distintiva a presena de uma estrutura semelhante a uma linha, que parece auxiliar a clula em sua localizao, evitando obstculos ou na captura de partculas de alimento. A maioria das haptfitas fotossintetizante, possuindo clorofila a e c. Tem importncia fundamental no ciclo do carbono e do enxofre.
Protista II - Heterocontas e algas verdes
As algas pardas, filo Phaeophyta, esto intimamente relacionadas a um conjunto diverso de protistas microscpicos, incluindo os oomicetos, filo Oomyhcota, diatomceas, filo Bacillariophyta, e crisfita, filo Chrysophyta. Coletivamente, estes organismos so conhecidos como heterocontfitos,
devido ao fato de possuirem dois flagelos que diferem em comprimento e ornamentao. Os oomicetos compreendem organismos desde unicelulares at formas filamentosas. A parede celular composta principalmente por celulose. A reproduo assexuada faz-se por meio de zosporos. A reproduo sexuada envolve uma oosfera grande e imvel e um gameta masculino pequeno e mvel. As diatomceas so organismos unicelulares ou coloniais que ocorrem tanto em gua doce como marinha. Elas so importantes componentes do fitoplncton e nicas por suas paredes silicosas divididas em duas partes. As Crisfitas podem apresentar pigmento dourado ou ser incolores; unicelulares ou coloniais. Abundantes em gua doce e oceanos de todo mundo. Algumas crisfitas alimentam-se de bactrias ou outras partculas orgnicas. As algas pardas abrangem as maiores e mais complexas algas marinhas. Em muitas formas, o talo vegetativo bastante diferenciado em apressrio, estipe e lmina. Algumas tm tecidos condutores de alimentos, prximos aos tecidos das plantas vasculares. Os Kelps gigantes formam florestas marinhas ecologicamente importantes. Estas algas so fontes de polissacardeos industrialmente importantes. Os ciclos de vida da maioria dos Kelps envolvem alternncia de geraes. filo Chlorophyta divide-se em trs classes: as Chlorophyceae, as Ulvophyceae e as Charophyceae. As algas verdes apresentam uma grande diversidade de formas de gua doce e terrestre, alm das marinhas. As Ulvophyceae so primariamente marinhas e a alternncia de geraes ocorre em algumas espcies. As Chlorophyceae so primariamente de gua doce. As Charophyceae de gua doce so as algas verdes mais intimamente relacionadas com brifitas e plantas vasculares.
Brifitas
Evidncias moleculares sugerem que as plantas terrestres so descendentes de algas verdes, e que as brifitas so as plantas vivas mais antigas, que divergiram a partir da linha principal de evoluo das plantas. As brifitas esto representadas hoje pelas Hepticas, Antceros e Musgos. O gametfito a gerao dominante. Os rgos sexuais masculinos, os anterdios, produzem nmerosos anterozoides, o arquegnio, rgo sexual feminino, fabrica uma nica oosfera. Os anterozoides so biflagelados e nadam at a oosfera. Da fecundao surge o esporfito que transitrio.
As hepticas diferem dos musgos e antceros pela ausncia de estmatos. Os antceros tem um tpico meristema basal e nele falta tecido condutor especializado. Os musgos, pelo menos em alguns grupos, tm tecidos condutores especializados e estmatos que lembram aqueles das plantas vasculares. Os tecidos condutores dos musgos, quando presentes, consistem nos hidroides, clulas condutoras de gua, e leptoides, clulas condutoras de alimento. As brifitas so abundantes e apresentam grande diversidade. Desempenham papel essencial no ciclo global do carbono.
Plantas vasculares sem sementes
As plantas vasculares so caracterizadas plos tecidos vasculares xilema e floema, alm de exibirem uma alternncia de geraes heteromorfas, na qual o esporfito maior e complexo, constituindo a fase nutricionalmente independente. Os corpos de muitas plantas vasculares consistem inteiramente em tecidos primrios. Atualmente, o crescimento secundrio est confinado, em grande parte, s plantas com sementes. Os tecidos vasculares primrios e tecidos fundamentais associados exibem trs arranjos bsicos: a) o protostelo, que consiste em um tecido vascular slido, centralmente localizado; b) o sifonostelo, que contm uma medula circundada pelo tecido vascular; e c) o eustelo, que consiste em um sistema de feixes circundando uma medula, com os feixes separados uns dos outros pelo tecido fundamental. Quando comparamos a origem de razes e folhas, percebemos que as primeiras surgiram de pores subterrneas do corpo primitivo da planta. As folhas no tm uma origem nica. Microfilos, folhas com uma nica nervura, evoluram ou como apndices laterais superficiais do caule ou de esporngios estreis. Megafilos, folhas com venao complexa, evoluram de sistemas de ramos. As plantas vasculares homosporadas produzem somente um tipo de esporo, que podem dar origem a um gametfito bissexuado. As plantas heterosporadas produzem micrsporos e megsporos, que germinam e do origem a gametfitos masculinos e gametfitos femininos, respectivamente. Tem havido uma longa e contnua tendncia evolutiva em direo reduo no tamanho e na complexidade do gametfito, que culminou nas angiospermas. Os ciclos de vida das plantas vasculares sem sementes representam modificaes de uma alternncia de geraes heteromorfas essencialmente similares, na qual o esporfito dominante e de vida livre. Os gametfitos das espcies homosporadas so nutricionalmente independentes do esporfito. Embora potencialmente bissexuados, produzindo tanto anterdios quanto arquegnios, estes gametfitos so funcionalmente unissexuados. Todas as plantas vasculares sem sementes tem anterozoides mveis e a presena de gua faz-se necessria para que o anterozoide nade at a oosfera.
As plantas vasculares sem sementes so atualmente classificadas em quatro fils: as Lycophyta (incluindo Lycopodium, Selaginella e Isoetes), as Psilotophyta (Psilotum e Tmesipteris), as Sphenophyta (Equisetum) e as Pterophyta (samambaias). A maioria homosporada, mas a heterosporia exibida por Selaginella, Isoetes e samambaias aquticas (Salviniales e Marsileales).
Gimnospermas
As plantas com sementes consistem em cinco fils com representantes vivos. Um desses o das bem-sucedidas angiospermas, filo Anthophyta. Este apresenta um conjunto interessante de caractersticas. Os outros quatro, que no apresentam estas caractersticas, so agrupados como gimnospermas. As estruturas semelhantes a sementes mais antigas que se conhecem ocorrem em estratos do perodo Devoniano Superior, h cerca de 365 milhes de anos. Os provveis ancestrais das gimnospermas e angiospermas so as progimnospermas, um grupo extinto de plantas vasculares sem sementes do Paleozoico. As gimnospermas atuais compreendem quatro fils: Cycadophyta, Ginkgophyta, Coniferophyta e Gnetophyta. Seus ciclos de vida so, em essncia, semelhantes, ou seja, uma alternncia de geraes heteromrficas com esporfitos grandes, independentes, e gametfitos muito reduzidos. Os vulos (megaesporngios mais tegumentos) apresentam-se expostos sobre megasporfilos ou estruturas anlogas. Na maturidade, o gametfito feminino da maioria das gimnospermas uma estrutura multicelular com vrios arquegnios. Os gametfitos masculinos desenvolvem-se dentro de gros de plen. Em todas as plantas com sementes, no se observam anterdios. Nas gimnospermas, os gametas masculinos formam-se diretamente a partir da clula gametognica. Em quase todas as plantas com sementes, observamos gametas imveis. Nestas, a gua no necessria para que o gameta masculino chegue oosfera, porque este transportado por uma combinao de polinizao e formao de tubo polnico. A fecundao ocorre quando um gameta masculino do microgametfito (o gro de plen germinado) une-se oosfera, que na maioria das gimnospermas est localizada num arquegnio. O segundo gameta masculino aparentemente no tem funo e se desintegra. Aps a fecundao, cada vulo desenvolve-se e forma uma semente. Existem quatro fils de Gimnospermas com representantes vivos. As conferas, filo Coniferophyta, so o maior e mais disperso filo de gimnospermas atuais, com cerca de 50 gneros e aproximadamente 550 espcies. Elas dominam
muitas comunidades vegetais em todo o mundo, com pinheiros, abetos e outras rvores muito observadas no Hemisfrio Norte. As cicadfitas, filo Chcadophyta, consistem em 11 gneros e cerca de 140 espcies, predominantemente tropicais, mas afastando-se do equador nas regies mais quentes. As cicadfitas so plantas parecidas com palmeiras, com troncos e crescimento secundrio extremamente lento. H apenas uma espcie viva do filo Ginkgophyta (Ginkgo biloba), conhecida apenas sob condies de cultivo. Os trs gneros do filo Gnetophyta correspondem s plantas vivas com maior afinidade em relao s angiospermas.
Introduo s angiospermas
As Angiospermas, ou plantas com flores, so do filo Anthophyta. As duas grandes classes deste filo so as monocotiledneas (65.000) e as eudicotiledneas (165.000). As plantas com flores diferem das outras plantas com sementes em diversas caractersticas , como a presena de endosperma nas sementes, devido aos vulos estarem encerrados nos megasporfilos, os carpelos. Ainda diferem pela presena de suas estruturas reprodutivas distintas, as flores, que se caracterizam por possurem carpelos e microsporfilos tambm especializados, os estames. As flores podem ter at quatro vertidos de apndices. De fora para dentro, os verticilos so as spalas (coletivamente, o clice); as ptalas (coletivamente, a corola); os estames (coletivamente, o androceu); e os carpelos (coletivamente, o gineceu). A polinizao nas angiospermas acontece pela transferncia do plen da antera para o estigma. Os gametas masculinos das angiospermas, ou clulas espermticas, so levados pelo gro de plen, que o microgametfito imaturo. O gametfito feminino das angiospermas denominado saco embrionrio. Ambas as clulas espermticas atuam na fecundao das angiospermas, a chamada dupla fecundao. Uma delas se une oosfera, produzindo o zigoto diploide. A outra se une aos dois ncleos polares, formando o ncleo primrio do endosperma, que comumente triploide (3n). Este ncleo se divide formando um tecido nutritivo chamado endosperma, que pode ser absorvido pelo embrio em desenvolvimento ou pode persistir na semente madura. Os ovrios desenvolvem-se em frutos, que envolvem as sementes. Juntamente com as flores das quais derivam, os frutos so uma caracterstica distintiva das angiospermas.
Nas angiospermas, ocorre a fecundao cruzada, mas tambm pode ocorrer a autopolinizao.
Evoluo das angiospermas
Os fsseis mais antigos atribudos claramente s angiospermas so do Cretceo inferior, cerca de 130 milhes de anos atrs. Eles incluem flores e plen. As plantas com flores se tornaram dominantes no mundo inteiro entre 80 e 90 milhes de anos atrs. A polinizao por insetos bsica nas angiospermas. Os primeiros agentes polinizadores foram, provavelmente, os besouros. As abelhas, entretanto, so os mais especializados e constantes insetos visitantes das flores e, provavelmente, tiveram o maior efeito sobre a evoluo da morfologia floral das angiospermas. Algumas angiospermas tornaram-se polinizadas pelo vento, pela gua e por diversos grupos de animais. Os frutos podem atrair animais que carregam as sementes e as lanam a quilmetros de distncia, facilitando, dessa forma, a disperso. A co-evoluo bioqumica tambm foi importante para muitas angiospermas, assim, algumas fabricam produtos secundrios que favorecem a sua adaptao, como alcaloides, que protegem a planta contra os herbvoros, apesar de alguns desses conseguirem lidar com a toxina e, assim, observamos uma co-evoluo gradual das interaes.
SEO 5 O corpo das angiospermas: estrutura e desenvolvimento Desenvolvimento inicial do corpo da planta
Iniciando-se com o zigoto, os sistemas caulinar e radicular da planta jovem so, a princpio, uma estrutura contnua. Com a diviso assimtrica do zigoto, a polaridade do embrio estabelecida. Atravs de uma progresso ordenada de divises, o embrio se diferencia em um suspensor e em um embrio propriamente dito, no interior do qual os meristemas primrios (os precursores da epiderme, do tecido fundamental e dos tecidos vasculares) e o padro radial so finalmente formados.
As sementes das angiospermas so formadas por um embrio, um envoltrio e alimento armazenado. Quando completamente formado, o embrio consiste basicamente em um eixo, portando um ou dois cotildones, e um meristema apical no pice do sistema caulinar e no pice da raiz. Os cotildones da maioria das eudicotiledneas so carnosos e contm o alimento armazenado na semente. A germinao desta depende de fatores ambientais, incluindo gua, oxignio e temperatura. Muitas sementes devem passar por um perodo de dormncia antes de serem capazes de germinar. A raiz a primeira estrutura a emergir pra a maioria das sementes em germinao, permitindo, assim, que a plntula se fixe ao solo e absorva gua.
Clulas e tecidos vegetais
Aps a embriognese, a maior parte do desenvolvimento da planta ocorre por intermdio da atividade dos meristemas. Os meristemas apicais esto envolvidos primariamente com o crescimento vertical das razes e dos caules, tambm chamado de crescimento primrio. O desenvolvimento compreende trs processos que se sobrepem: crescimento, morfognese e diferenciao. As plantas vasculares so formadas por trs sistemas de tecidos: drmico, vascular e fundamental, que esto presentes na raiz, no caule e nas folhas.
Raiz: estrutura e desenvolvimento
Sustentao, absoro, armazenamento e conduo so as funes desempenhadas pelas razes. Gimnospermas, magnoliideas e eudicotiledneas comumente produzem sistema radicular do tipo pivotante, enquanto monocotiledneas, usualmente, produzem sistemas radiculares fasciculados. Durante o crescimento primrio, o meristema apical d origem aos trs meristemas primrios: protoderme, meristema fundamental e procmbio, os quais se diferenciam, respectivamente, em epiderme, crtex e cilindro vascular. Alm disto, o meristema apical produz a coifa, cuja funo proteger o meristema apical e auxiliar a raiz na penetrao do solo. Muitas clulas da epiderme da raiz desenvolvem plos radiculares, que aumentam sensivelmente a superfcie de absoro da raiz. O xilema primrio usualmente ocupa o centro do cilindro vascular e possui proje-es que se alternam com os cordes de floema primrio. As razes laterais originam-se do periciclo e foram seu caminho para fora, atravs do
crtex e da epiderme. O crescimento secundrio das razes envolve o cmbio vascular e o cmbio da casca. A cenoura, a batata-doce e a beterraba caracterizam-se por serem plantas cujas razes so rgos de reserva.
O sistema caulinar: estrutura primria e desenvolvimento
Os pices do sistema caulinar vegetativo de grande parte das plantas florferas tm um tipo de organizao tnica-corpo que consiste em uma ou mais camadas perifricas de clulas (a tnica) e uma massa interna de clulas (o corpo). A maior parte das angiospermas tem pices compostos por trs camadas de clulas superpostas (duas camadas de tnica e uma camada inicial do corpo). O caule cresce verticalmente, em grande parte, devido ao alongamento internodal. Assim como na raiz, o meristema apical do sistema caulinar origina a protoderme, o procmbio e o meristema fundamental, que desenvolvem os tecidos primrios. Independente do tipo de organizao, o floema comumente localizado externamente ao xilema. O termo "sistema caulinar" designa no apenas um termo coletivo para o caule e suas folhas, mas tambm, uma expresso de sua ntima associao fsica e ontogentica. Os primrdios foliares so originados por grupos de clulas, denominadas clulas fundadoras, na regio perifrica do pice do sistema caulinar, e sua posio refletida no padro do sistema vascular do caule. As variaes na estrutura foliar normalmente esto relacionadas com o habitat. As lminas de algumas folhas so divididas em fololos (folhas compostas), enquanto outras no o so (folhas simples). Os estmatos so geralmente mais nmerosos na superfcie inferior da folha. O tecido fundamental da folha, o mesofilo, tem propriedades de um tecido fotossintetizante e, em mesfitas, est diferenciado em parnquima palidico e parnquima lacunoso. O mesofilo amplamente permeado por espaos aerferos e pelas nervuras, que so compostas por xilema e floema circundados por uma bainha do feixe de natureza parenquimtica. O xilema normalmente ocorre na parte superior da nervura, enquanto o floema ocorre na parte inferior. As folhas apresentam crescimento determinado e os caules apresentam crescimento indeterminado. Na florao, em muitas espcies, o pice do sistema caulinar vegetativo direta-mente transformado em pice reprodutivo. Assim como as razes, os caules podem ser especializados em armazenagem de alimentos. Tubrculos, bulbos e cormos so exemplos de caules carnosos. As plantas armazenadoras de gua so conhecidas como suculentas.
Crescimento secundrio em caules
Para um botnico, o lenho e a casca so indicadores do crescimento secundrio, evidncia de que uma planta cresceu mais em dimetro do que em altura. O crescimento secundrio ocorre em todas as gimnospermas e na maioria das angiospermas (as monocotiledneas), envolvendo a atividade de dois meristemas laterais (o cmbio vascular e o cmbio da casca ou felognio). As plantas herbceas podem apresentar pouco ou nenhum crescimento secundrio, enquanto as plantas lenhosas (rvores e arbustos), podem continuar a aumentar em espessura por muitos anos. Nos caules e razes velhos, a maior parte do floema no-funcional. Os elementos crivados possuem vida curta e, geralmente, somente aqueles formados no ano de crescimento vigente so condutores ou funcionais. As madeiras so classificadas como softwoods e hardwoods. As primeiras correspondem madeira de conferas e as ltimas correspondem madeira de angiospermas (magnliidas e eudicotiledneas lenhosas). O lenho - xilema secundrio - das conferas mais simples do que o lenho das angiospermas. As camadas de crescimento que correspondem ao incremento anual de crescimento so denominadas anis de crescimento anuais. SEO 6 Fisiologia das plantas com sementes Regulando o crescimento e o desenvolvimento: os hormnios vegetais
Os hormnios vegetais so reguladores qumicos que induzem o organismo a produzir respostas fisiolgicas, mesmo quando em quantidades extremamente pequenas. Tradicionalmente, cinco grupos de fitormnios so alvo de maior ateno: auxinas, citocininas, etileno, cido abscsico e giberelinas. A auxina produzida nos meristemas apicais dos caules e nas extremidades dos coleptilos. Seu transporte unidirecional, sempre em direo base da planta, onde controla a extenso do caule, principalmente pela promoo do alongamento celular. A auxina tambm exerce papel na diferenciao de tecidos vasculares e inicia a diviso no cmbio vascular. Frequentemente, inibe o crescimento das gemas laterais, mantendo a dominncia apical. Esse hormnio vegetal promove a formao de razes adventcias em estacas e
retarda a absciso de folhas, flores e frutos. Nos frutos, a auxina produzida nas sementes estimula o crescimento da parede do ovrio. As citocininas esto quimicamente relacionadas a certos componentes dos cidos nuclicos. Elas so mais abundantes em tecidos com alta atividade de diviso, como as sementes, os frutos e as folhas. O etileno o nico hormnio vegetal gasoso. Exerce grande influncia em diversos aspectos do crescimento e do desenvolvimento das plantas, incluindo o crescimento de muitos tecidos, a absciso de frutos e folhas e o amadurecimento de frutos. O cido abscisico (ABA) um hormnio inibidor de crescimento encontrado em gemas e sementes dormentes. O ABA estimula a produo de protenas de sementes e o fechamento de estmatos. As giberelinas controlam o alongamento do caule. Podem substituir a quebra de dormncia pelo frio ou pela luz requerida para a germinao de suas sementes. Os avanos na pesquisa hormonal e na bioqumica do DNA possibilitaram a manipulao das caractersticas genticas das plantas de modo especfico. O potencial de desenvolvimento das clulas at a formao de plantas completas denominado totipotncia.
Fatores externos e crescimento vegetal
O fototropismo uma curvatura, em direo luz, de um sistema caulinar em crescimento. O gravitropismo a resposta de um sistema caulinar ou radicular gravidade. O clcio aparentemente desempenha um importante papel na resposta gravitrpica de sistemas caulinares e radiculares. O hidrotropismo o crescimento em resposta a um gradiente de umidade. resposta ao contato com um objeto slido, tal como o enrolamento de gavinhas, d-se o nome de tigmotropismo. As respostas fotoperidicas controlam o desencadeamento do processo de florao em diversas plantas. Algumas delas, conhecidas como plantas de dias longos, florescem apenas quando os perodos de luz excedem uma durao crtica. Outras plantas, as de dias curtos, florescem apenas quando os perodos de luz so mais curtos do que uma durao crtica. Plantas neutras florescem independentemente de fotoperodo. Experimentos tm demonstrado que o fator crtico o perodo de escuro, e no, o de luz.
Os movimentos das plantas que ocorrem em resposta a um estmulo, cuja direo independe da direo deste, so chamados movimentos nsticos. Entre eles, esto os movimentos de sono, de larga ocorrncia, tambm chamadosnictinsticos. Os movimentos nsticos, que resultam do toque, incluem o fechamento da folha da planta carnvora Dionaea. As plantas podem responder a estmulos mecnicos, alterando os seus padres de crescimento. Fenmeno conhecido como tigmomorfognese. Alm disso, as folhas e flores de variadas plantas acompanham, ou rastreiam, o sol ao longo do dia.
Nutrio vegetal e solos
Um total de 17 nutrientes inorgnicos so necessrios maioria das plantas para o crescimento normal. Desses, carbono, hidrognio e oxignio so derivados do ar e da gua. Os demais so absorvido pelas razes sob a forma de ons. Esses 17 elementos so categorizados como micronutrientes ou macronutrientes, dependendo da quantidade na qual eles so necessrios. Molibdnio, nquel, cobre, zinco, mangans, boro, ferro e cloro so os micronutrientes. Enxofre, fsforo, magnsio, clcio, potssio, nitrognio, hidrognio, carbono e o oxignio so os macronutrientes. Nutrientes inorgnicos regulam a osmose e afetam a permeabilidade celular. Alguns deles funcionam como componentes estruturais das clulas, como componentes de metablitos essenciais e como ativadores e componentes de enzimas. Os ciclos de nutrientes envolvem tanto organismos vivos quanto o ambiente fsico. Em decorrncia disso, eles so tambm denominados ciclos biogeoqumicos. Os ciclos dos nutrientes possuem vazamentos, ou seja, nem todos os nutrientes que retornam ao solo tornam-se disponveis para o uso das plantas. O processo de oxidao da amnia ou ons amnio para formao de nitritos e nitratos denominado nitrificao. Um tipo de bactria responsvel pela oxidao da amnia em nitrito, e outro para oxidao do nitrito em nitrato. O nitrognio absorvido pelas plantas, quase exclusivamente, na forma de nitrato. As bactrias fixadoras de nitrognio mais eficientes so aquelas que
mantem relaes simbiticas com plantas, as quais produzem ndulos radiculares no local da infeco. Na agricultura, o nitrognio e outros elementos no so reciclados quando as plantas so removidas do solo, como acontece na natureza. Por isso h a necessidade de reposio desses nutrientes em forma orgnica ou inorgnica. O ciclo do fsforo difere em parte daquele do nitrognio, pois a crosta terrestre (e no a atmosfera) o reservatrio para reposio do fsforo. Este circula das plantas para os animais e retorna ao solo em formas orgnicas, as quais so posteriormente convertidas em formas inorgnicas por microrganismos. A remoo realizada nas colheitas e o aumento da eroso tm acelerado a perda de fsforo nos solos; os pntanos e os charcos, stios primrios de desnitrificao, tm sido destrudos; os ambientes aquticos esto sujeitos a danos generalizados devido chuva cida. Tanto leguminosas quanto bactrias esto sendo geneticamente selecionadas para combinaes que podem resultar no aumento da fixao de nitrognio em ambientes especficos. A engenharia gentica oferece a possibilidade de transferncia de genes necessrios fixao de nitrognio de um organismo para outro.
O movimento de gua e solutos nas plantas
A maior parte da gua absorvida pelas razes de uma planta perdida para o ambiente na forma de vapor d'gua. Um par de clulas-guarda pode mudar sua prpria forma para controlar a abertura e o fechamento do estmato ou poro. O fechamento dos estmatos evita a perda de vapor d'gua pela folha. Um estmato se abre quando suas clulas-guarda se tornam trgidas e se fecha quando estas se tornam flcidas. Os fatores ambientais tambm afetam o movimento estomtico, incluindo a concentrao de dixido de carbono, luz e temperatura. A teoria da tenso e coeso a mais aceita para explicar o mecanismo do movimento da gua at o pice de rvores altas, atravs do xlema. De acordo com essa teoria, a gua "puxada" ou tensionada atravs do corpo da planta. Esta tenso deve-se transpirao e/ ou ao uso de gua nas folhas, resultando em um gradiente de potencial hdrico das folhas para a soluo do solo em contato com a superfcie das razes. Os plos radiculares fornecem uma enorme superfcie para a absoro de gua. Em algumas plantas, a absoro da gua presente no solo resulta na gerao de uma presso positiva ou presso de raiz, quando a transpirao muito lenta ou ausente. A gutao um processo no qual a gua, na forma lquida, forada a sair atravs de estruturas especiais (hidatdios) nas pontas ou margens das folhas.
Os nutrientes inorgnicos tomam-se disponveis para as plantas na soluo do solo, sob a forma de ons. Quantidades substanciais de ons inorgnicos, que so importadas at as folhas atravs do xilema, so trocadas com o floema das nervuras foliares e exportadas destas juntamente com a corrente de assimilados. De acordo com a hiptese do fluxo de massa, os assimilados se deslocam da fonte para o dreno, segundo um gradiente de presso de turgor desenvolvido osmoticamente.
SEO 7 Ecologia
A dinmica das comunidades e ecossistemas
Os ecossistemas so sistemas auto-sustentados que incluem os organismos vivos e os elementos inertes (fsicos) do ambiente com os quais eles interagem. As comunidades consistem em todos os organismos que vivem numa dada rea. Alguns dos relacionamentos que ocorrem em comunidades podem ser agrupados em trs categorias: mutualismo, competio e interaes plantas-herbvoros (e plantas-patgenos). No mutualismo, duas populaes interagem em benefcios de ambas. Interaes competitivas so encontradas entre a maioria das espcies de plantas que crescem em estreita proximidade e tambm entre muitas plantas isoladas. Um dos tipos mais importantes de competio aquela por luz. As plantas reagem aos efeitos dos herbvoros, os quais limitam o potencial reprodutivo destas, pelo desenvolvimento de espinhos, folhas duras e estruturas semelhantes, e, o que mais importante, defesas qumicas. Um ecossistema consiste em elementos inertes e em dois tipos de elementos vivos, auttrofos (produtores primrios) e hetertrofos (consumidores). Entre os hetertrofos, esto os consumidores primrios ou herbvoros; os consumidores secundrios ou carnvoros e parasitas; e os decompositores. A energia flui atravs dos ecossistemas. 1% ou menos da energia luminosa incidente convertida em energia qumica pelas plantas verdes. Quando essas plantas
so consumidas, menos de 20% da sua energia potencial armazenada no prximo nvel trfico. Dessa forma, cadeias alimentares raramente tm mais de trs ou quatro elos. Na maioria dos ecossistemas, a maior concentrao de energia, biomassa e indivduos ocorre nos nveis trficos inferiores, o que d origem ao fenmeno conhecido como pirmides de energia, de biomassa e de nmeros. Quando uma rea naturalmente aberta ou foi devastada, ocorre a sucesso ecolgica. As espcies pioneiras recorrentes nas clareiras crescem rapidamente sob condies de alta luminosidade, desenvolvendo
caractersticas diferentes daquelas das rvores que dominam a floresta adulta. Finalmente, a sucesso pode resultar na produo de uma comunidade clmax.
Ecologia global
Os biomas so ecossistemas terrestres caracterizados por uma vegetao que lhes peculiar. A distribuio dos biomas o resultado de complexas interaes entre a distribuio do calor do sol, padres de circulao do ar e fatores geolgicos. Alm do clima, as diferenas nas superfcies dos continentes, tais como a composio do solo e a altitude, afetam os tipos de plantas e animais encontrados nos mais variados biomas da Terra. As florestas pluviais tropicais, nas quais a gua e a baixa temperatura no so os fatores limitantes, so, de longe, o bioma mais rico em nmero de espcies. As rvores so sempre verdes e caracterizadas por folhas coriceas de tamanho mdio. Os solos tropicais so frequentemente cidos e muito pobres em nutrientes; tais solos perdem sua fertilidade rapidamente quando a floresta derrubada. A maioria das comunidades tropical e subtropical, caracterizadas pelo fenmeno da seca sazonal, chamada savana. As rvores e arbustos dessas comunidades so total ou parcialmente decduos, perdendo suas folhas durante as pocas secas. Afastando-se do equador, comunidades tropicais e subtropicais vo gradualmente se convertendo em desertos e semidesertos, que so caracterizados pela baixa precipitao. As plantas suculentas e ervas anuais so comuns nesses ambientes. As campinas, que podem gradualmente dar lugar a savanas, desertos e florestas temperadas, so caracterizadas pela ausncia quase total de rvores,
exceto ao longo dos cursos d'gua. Os solos das campinas so os mais produtivos para a agricultura temperada. Nas florestas temperadas decduas, a maioria das rvores perde suas folhas durante a poca mais fria dos invernos. Elas so margeadas por florestas temperadas mistas e florestas de conferas ao norte, nas quais as conferas desempenham um importante papel. A taiga uma vasta floresta de conferas que se estende em faixas contnuas atravs da Eursia e Amrica do Norte e da Costa do Pacfico, descendo at o norte da Califrnia. Nos seus limites meridionais, a taiga dominada por rvores altas com um enorme crescimento de brifitas e liquens; ao norte, ela consiste em vastas faixas montonas de florestas com pouqussimas espcies arbreas. A tundra uma regio sem rvores e tambm se estende ao redor do hemisfrio norte, predominantemente, sobre o Crculo rtico, em uma faixa que interrompida apenas por massas de gua. A vegetao composta por arbustos baixos, gramneas e liquens. Tanto os limites setentrionais da taiga como toda a tundra esto embasados sobre permafrost.
A perspectiva humana
A raa humana originou-se na frica. O gnero Australopithecus existia nessa regio h, pelo menos, 5 milhes de anos. O gnero Homo, aparentemente evoluiu a partir de Australopithecus, h cerca de 2 milhes de anos. A nossa espcie, Homo sapiens, existe h, pelo menos, 500.000 anos. Iniciando-se cerca de 11.000 anos atrs, no Crescente Frtil, o homem comeou a cultivar plantas como cevada, lentilha, trigo e ervilha. A agricultura espalhou-se pela Europa, chegando at a Bretanha h cerca de 6.000 anos. H evidncias de que ela tenha se estendido tambm para o sul atravs da frica. Muitas plantas tornaram-se culturas primeiramente na frica, incluindo o inhame, o quiabo, o caf e o algodo, que foi tambm domesticado independentemente no Novo Mundo e talvez na sia. Neste continente, desenvolveu-se uma agricultura fundada em alimentos bsicos, como o arroz e a soja e, mais para o sul, os ctricos, a manga, a taioba, a banana e outras culturas. Rebanhos de animais de pastoreio foram ecologicamente destrutivos para muitas reas semi-ridas do Velho Mundo, especialmente quando os animais se reproduziam, mas eles tambm foram importantes fontes de alimento. A agricultura foi desenvolvida independentemente no Novo Mundo. Ela teve incio h, pelo menos, 9.000 anos, no Mxico e no Peru. Colombo e outros que o seguiram encontraram uma verdadeira opulncia de novas culturas para levar ao Velho Mundo. Dentre essas o milho, o feijo comum, o feijo-de-lima, o tomate, o tabaco, a pimenta, a batata, a batata-doce, a mandioca, a moranga, a
abbora, o abacate, o cacau e as principais espcies cultivadas de algodo. As especiarias - partes de plantas fortemente aromticas e geralmente ricas em leos essenciais - podem ser derivadas de razes, cascas, sementes, frutos ou gemas, enquanto as ervas aromticas so geralmente folhas de plantas nolenhosas. O suprimento mundial de alimento baseado em um nmero relativamente pequeno de plantas agrcolas. Seis culturas: trigo, arroz, milho, batata, batatadoce e mandioca, so responsveis por mais de 80% das calorias consumidas pela populao humana mundial. A populao humana cresceu de um nmero estimado em 5 milhes de pessoas, na poca em que a agricultura comeou a se desenvolver, para 6 bilhes no final do sculo XX. Como consequncia desse crescimento e da misria generalizada, e tambm porque, relativamente, pouco tem sido feito para o desenvolvimento de prticas agrcolas adequadas s regies tropicais, os trpicos esto sendo ecologicamente devastados. A soluo para problemas atuais requer uma abordagem integrada, ou seja, o fornecimento mundial de alimento pode ser dinamizado plos mtodos tradicionais do melhoramento gentico e seleo, pelo cultivo de novas culturas e plos mtodos da engenharia gentica. A destruio generalizada de habitats dos trpicos est ameaando a sobrevivncia de muitas espcies vegetais potencialmente teis, antes mesmo delas virem a ser identificadas e exploradas.
Sntese elaborada por Paulo Bueno Guerra
12. RIDLEY, M. Evoluo. 3. ed. Porto Alegre: Artmed, 2006. Este livro narra a trajetria da evoluo, comeando por seus aspectos histricos, chegando at os avanos mais recentes na teoria evolutiva. Evoluo significa descendncia com modificaes ou alterao na forma, da fisiologia e do comportamento de organismos ao longo de muitas geraes de tempo. As mudanas evolutivas dos seres vivos ocorrem em um padro arbreo ramificados de linhagens. Os seres vivos possuem adaptaes: isto , eles so bem justados em forma, fisiologia e comportamento para a vida no ambiente natural. Muitos pensadores precursores de Darwin discutiram a
possibilidade de as espcies transformarem-se, ao longo do tempo, em outras espcies. Lamark o mais conhecido deles. Porm em meados do sculo XIX, a maioria dos bilogos acreditava que as espcies tinham formas fixas. A teoria da evoluo por seleo natural de Darwin explica mudanas e adaptaes evolutivas. Os contemporneos de do mesmo em geral aceitaram a ideia de evoluo, mas no a sua explicao com base na seleo natural. Darwin no postulou uma teoria da hereditariedade. Quando as ideias de Mendel foram redescobertas na virada do sculo XX, pensava-se, inicialmente que elas contrariavam a teoria da seleo natural. Fisher, Haldane e Wright demonstraram que a herana mendeliana e a seleo natural so compatveis, a sntese dessas duas ideias chamada de neodarwinismo ou de teoria sinttica da evoluo. Durante a dcada de 1930 e 1940, o neodarwinismo gradualmente se espalhou por todas as reas da Biologia e tornou-se amplamente aceito. Ele unificou a gentica, a sistemtica, a paleontologia, a morfologia comparativa clssica e a embriologia. Em relao a Hereditariedade, determinada por uma molcula chamada de DNA. Essa molcula pode ser dividida em regies chamadas de genes, os quais codificam protenas. O cdigo no DNA lido para produzir uma protena em duas etapas: transcrio e traduo. O cdigo gentico j foi decifrado. O DNA est fisicamente organizado em estruturas chamadas cromossomos. Cada individuo possui um conjunto duplo de cromossomos (um herdado de seu pai e outro herdado de sua me) e portanto, dois conjuntos de todos os seus genes. Uma combinao particular dos genes de um indivduo chamada de gentipo. Novas variaes genticas originam-se por alteraes mutacionais no DNA. As taxas de mutao podem ser estimadas por observao direta. Quando dois indivduos com determinados gentipos se acasalam, as propores dos gentipos da respectiva prole aparecem em propores mendelianas previsveis. As propores exatas dependem dos gentipos no cruzamento. Diferentes genes so preservados ao longo das geraes no sistema de herana mendeliana, e isso permitem que a seleo natural opere. Antes de Darwin, pensava-se (incorretamente) que os materiais hereditrio materno e paterno se misturavam em um indivduo, em vez de serem preservados. Se a hereditariedade fosse por mistura, a seleo natural seria muito menos poderosa do que em uma situao de herana mendeliana. Quanto s evidncias da evoluo, vrias linhas sugerem que as espcies evoluram a partir de um ancestral comum, em vez de terem formas fixas e terem criadas separadamente. Em uma pequena escala, a evoluo pode ser observada enquanto acontece na natureza, como no caso dos padres de colorao de mariposas e nos experimentos de seleo artificial, como aqueles utilizados no cruzamento de variedades agrcolas. A variao
natural pode ultrapassar a fronteira da espcie como, por exemplo, nas espcies em anel de salamandras, e novas espcies podem ser criadas artificialmente, como processo de hibridizao e de poliploidia, pelo qual muitas variedades agrcolas e hortcolas foram criadas. A observao da evoluo em pequena escala, combinada com princpio extrapolativo do uniformitarianismo, sugere que toda a vida poderia ter evoludo a partir de um nico ancestral comum. Na seleo Natural e Variao, agindo sobre quaisquer entidades capazes de se reproduzirem, que apresentarem herana de suas caractersticas de uma gerao para outra, e que variam em aptido (isto , o nmero relativo de descendentes diretos que elas produzirem) de acordo com as caractersticas que possurem. A seleo pode ser direcional, estabilizadora ou disruptiva. Gentica Evolutiva, a transferncia de genes de uma gerao para a prxima no um processo perfeitamente exato, porque a amostragem aleatria pode modificar a frequncia de um gene. Os efeitos da amostragem aleatria so poderosas quando todos os gentipos diferentes possuem o mesmo valor adaptativo e quando os tamanhos populacionais so pequenos. A teoria da deriva aleatria tem sido muito importante para o raciocnio sobre evoluo molecular. A teoria neutra da evoluo molecular sugere que a evoluo molecular principalmente devida a deriva neutra. Sob esse aspecto, as mutaes que surgiram na evoluo foram seletivamente neutras em relao aos genes que elas substituram. Alternativamente, a evoluo molecular pode ser guiada, sobre tudo, por seleo natural. A gentica de populaes para dois ou mais locos est preocupada com trocas nas frequncias de hapltipos, os quais so os equivalentes de mltiplos locos dos alelos. A gentica quantitativa, a qual est relacionada com caractersticas controladas por muitos genes, considera as trocas nas distribuies de frequncias fenotpica e genotipica entre geraes, em vez de seguir o destino de genes individuais. Adaptao e seleo natural, Darwin e pensadores prdarwinianos haviam tentado explicar a sua existncia, sustentando que somente a seleo natural consegue explic-la. Algumas caractersticas, especialmente molecular, evoluram por outros processos que no a seleo natural, mas elas no so adaptaes. Nem toda evoluo ocorre por seleo natural, mas toda evoluo adaptativa sim. S a seleo natural funciona como teoria cientifica. As adaptaes evoluem por meio da seleo natural. Quando a seleo natural age, altera as frequncias das entidades em vrios dos nveis
da hierarquia da organizao biolgica. Ela tambm produz adaptaes que beneficiam entidades em vrios nveis. Adaptaes na Reproduo Sexuada, o sexo tem uma desvantagem adaptativa de 50% em relao reproduo assexuada. As populaes de reproduo sexuada evoluiro mais rapidamente do que um conjunto de clones assexuados, desde que a taxa de mutaes favorveis seja suficientemente alta. Evoluo e Diversidade, na Biologia evolutiva, as espcies podem ser entendidas como conjunto de genes, conjuntos de organismos que intercruzam e eles so unidades importantes porque, na teoria da gentica de populaes, a seleo natural ajusta a frequncia dos genes nos conjuntos gnicos. Como disse Darwin, os milhes de espcies que atualmente habitam este planeta evoluram de um ancestral comum e a multiplicao do nmero de espcies foi gerada do mesmo modo como uma nica espcie que se separou quando duas populaes evoluam independentemente e acumulavam diferenas genticas incompatveis. Especiao significa a evoluo do isolamento reprodutivo entre duas populaes. Filogenias revelam as relaes ancestrais entre espcies. Ela mostra com que outras espcies (ou grupo delas) uma determinada espcie compartilha o ancestral comum mais recente. As relaes filogenticas so inferidas por meio dos caracteres compartilhados pelas espcies. Eles podem ser morfolgicos, de espcies viventes ou fosseis, ou moleculares. Classificao e Evoluo, diz respeito a distino e a descrio das espcies viventes e fosseis e organizao dessas espcies em uma classificao hierrquica com vrios nveis. H dois grandes princpios e trs grandes escolas de classificao biolgica: o princpio fenetico o filogentico, e as escolas fentica, cladstica e evolutiva. Biogeografia Evolutiva, a cincia que procura explicar a distribuio das espcies e de txons mais elevados na superfcie da terra. Macroevoluo tem sido estudada com tcnicas genticas, usando observaes e experimentos com durao de uma existncia humana, por meio da evidncia fssil, da morfologia comparada e da inferncia filogentica. Macroevoluo refere-se s mudanas intrapopulacionais,nas frequncias gnicas. Coevoluo - ocorre quando duas ou mais espcies influenciam as evolues umas das outras. Ela invocada frequentemente para explicar a coadaptao entre espcies.
Extino e Irradiao, fatores que governam a diversidade no documentrio fssil.
13. SCHMIDT-NIELSEN, K. Fisiologia Animal. Adaptao e meio ambiente. 5. ed. So Paulo: Livraria Santos, 2002. O que fisiologia?
A fisiologia trata das funes dos organismos vivos-como se alimentam, respiram e se movimentam, e o que fazem para se manter vivos. Para usar palavras mais tcnicas, a fisiologia aborda o alimento a alimentao, digesto, respirao, transporte de gases no sangue, circulao e funo cardaca, excreo e funo renal msculos e movimentos, etc. O animal morto possui as estruturas que realizam estas funes; no animal vivo, essas estruturas funcionam. A fisiologia tambm estudada como o organismo vivo ajuda-se s adversidades do ambiente- obtm gua suficiente para viver ou evitar excesso de gua, escapa do congelamento que leva a morte ou morrer de calor excessivo movimenta-se para encontrar locais, alimentos e companheiros adequados e como ele obtm
informaes sobre o ambiente por meio de seus sentidos. Finalmente, a fisiologia trata da regulao de todas essas funes-como elas esto correlacionadas harmonioso. Ao se examinar como um animal enfrenta seus ambientes, com frequncia, possvel demonstrar o que pe benfico para ele. Isso pode nos levar, com certo incomodo, a explicaes que sugerem evidncia do propsito, ou teleologia, e muitos bilogos consideram isto cientificamente inadequado. Com tudo tendemos a perguntar; Por qu? Ou, quo benfico para o e integradas em um organismo de funcionamento
animal? De qualquer modo, o animal tem de sobrevier e no h nada de inadequado ou cientifico em descobrir como e por ele bem-sucedido. Se o animal no chegasse a solues para o problema de sobrevivncia, no estaria mais a disposio para ser estudado. E o estudo do organismo Vivo a razo de ser da fisiologia.
1: Oxignio / respirao
Por que o oxignio importante? Pois a maioria dos animais satisfaz sua necessidade de energia por meio da oxidao de alimentos, com a formao de dixido de carbono e gua no processo. O processo de tomada de oxignio e liberao de dixido de carbono denominado respirao. Os animais aquticos captam oxignio a parti da pequena deste gs dissolvido na gua e os animais terrestres, por meio do meio do oxignio abundante no ar. Muitos animais pequenos podem captar oxignio em quantidades suficientes pela superfcie corprea geral, mas a maioria dos animais necessita de rgos respiratrios especiais para a captao deste gs. O dixido de carbono segue o caminho oposto, sendo liberado atravs da superfcie corprea ou dos rgos respiratrios. A gua formada nos processos de oxidao faz meramente parte do reservatrio geral de gua no corpo e no representa qualquer problema especial. O mais importante, s vezes, o nico processo fsico responsvel pelo movimento de oxignio do meio externo para a clulas a difuso, processo no qual uma substancia passa de uma concentrao maior para uma menor. O movimento de dixido de carbono na direo oposta tambm seque os gradientes de concentrao. A difuso pode ser auxiliada por movimento de massa, tal como o movimento do ar entrando e saindo dos pulmes, mas os gradientes de concentrao continua sendo a fora motriz fundamental para movimentar os gases
respiratrios. Para entender a respirao necessrio conhecer os gases respiratrios, suas solubilidades e a fsica dos processos de difuso.
A ATMOSFERA
Composio do ar atmosfrico seco
Os gases mais importantes do ponto de vista fisiolgico so o oxignio, o dixido de carbono e o nitrognio. Alm disso, a atmosfera contm vapor de gua em quantidade de muito varivel. A composio da atmosfera permanece extremamente constante. As correntes de convico causam extensa mistura a uma altitude de pelo menos 100 km e nenhuma mudana aprecivel na composio percentual tem sido demonstrada apesar de a presso do ar ser muito reduzida em grandes altitudes. A afirmao de que os gases mais leves, notadamente o hidrognio e o Helio, so enriquecidos nos limites externos da atmosfera se aplica as camadas mais perifricas que no so de interesse fisiolgico. Para os nossos propsitos, a atmosfera aberta tem uma composio gasosa constante, exceto pelo vapor de gua. A composio do ar mantida como um equilbrio entre o uso de oxignio em processo de oxidao (principalmente a oxidao de compostos orgnicos em dixido de carbono) e a assimilao de dixido de carbono pelas plantas que no processo liberam oxignio.
O efeito estufa
O ligeiro aumento de dixido de carbono causado pela combusto de todo combustvel teria da mesma forma efeitos fisiolgicos desprezveis, mas no
quer dizer que seriam inofensivos. Mesmo uma leve variao no dixido de carbono altera a absoro de radiao solar na atmosfera e poder haver um efeito estufa imprevisvel, que com o passar dos anos pode mudar drasticamente as condies climticas na superfcie terrestre. A atmosfera mais transparente entrada de radiao de ondas curtas que radiao de ondas longas emitidas pela terra. A emisso de radiao de ondas longas absorvida na atmosfera principalmente dixido de carbono e pelo vapor de gua. Estima-se que a duplicao da quantidade de dixido de Atmosfrico aumentaria a temperatura mundial em aproximadamente 1,3 C se a gua e a atmosfera permanecem constantes. Isto causaria extensa funo do gelo polar e elevao do nvel do mar de 20 a 30m ou mais, o suficiente para submergir reas litorneas imensas e muitas das principais cidades do mundo, alm disso, a uma temperatura mais alta, a atmosfera pode reter vapor de gua, O que intensifica o efeito de cobertura e causa maior aumento de temperatura. Contudo, uma elevao do vapor de gua atmosfrico pode aumentar a formao de nuvens, que, por sua vez, refletem, mas a entrada de radiao solar, tendo assim o efeito oposto. A complexidade destas relaes tornam as previses relativas ao efeito estufa por aumento do dixido de carbono altamente insertas.
Difuso de gases
Nos rgos respiratrios dos animais (p.ex. brnquias, pulmes), os gases se difundem entre o ambiente e o organismo; o oxignio e captado e o dixido de carbono liberado pelo animal. Por isso, de interesse saber com que velocidade os gases se difundem, suas velocidades de difuso. Este assunto precisa de ateno especial, e muitos bilogos so levados a acreditar que o dixido de carbono se difunde muito mais rapidamente que o oxignio. Na realidade isso no acontece.
Quando o dixido de carbono funde-se entre o ar e a gua, sua alta solubilidade na gua faz com que parea estar se difundindo para o interior da gua.
Comparao entre a gua e o ar
As diferenas entre a gua e o ar so extremamente importantes para os animais no tocante ao suprimento de oxignio. Por exemplo, compare a qualidade de oxignio disponvel na gua e no ar. A gua em equilbrio com o ar atmosfrico a 15 C contem 7ml de O2 por litro (1.000 ml). Estes 7 ml de O2 pesam 0,01 g esta qualidade encontrada em um peso de gua 100.000 vezes maior. Para a obteno de uma data
quantidade de oxignio, deve-se, por tanto mover 100.000 vezes seu peso * de gua pelos rgos respiratrios. Em contraste, 1 litro de ar contm 209 ml O2, que pesa 280 mg. O restante do ar 791 ml de N2, pesa 910 mg. Para a obteno de oxignio devem-se mover apenas 3,5 vezes na sua massa de gs inerte essa enorme diferena na massa do meio inerte tem uma enorme importante consequncia: nos sistemas respiratrios aquticos, o movimento de guas quase sempre unidirecional. Seu fluxo de gua fosse para dentro e para fora ou para trs e para frente, uma grande massa de gua teria de ser acelerada, em seguida desacelerada, e novamente acelerada em direo oposta. Um alto dispndio de energia seria necessrio para as continuas modificaes na emergncia cintica da gua. Para rgos respiratrios que utilizam o ar atmosfrico, tais como o pulmo, um fluxo para dentro e para fora no muito dispendioso, pois a massa inerte apenas poucas vezes maior a massa de oxignio utilizada. Alm da massa do meio respiratrio, outro fator contribui para a quantidade de trabalho necessrio para mov-la ponto. A gua tem uma viscosidade maior que a do ar e isto aumenta o trabalho necessrio para bombear o
fluxo, pois a fora motriz deve ser aumentada proporcionalmente a maior viscosidades. Como a energia necessria para mover o fluido aumenta em proporo direta a presso, ela tambm aumenta em proporo viscosidade.
Respirao na gua Alguns dos ajustes mais simples para a troca de gases respiratrios so encontrados nos animais aquticos. Muitos pequenos organismos obtm oxignio por difuso, atravs das superfcies do corpo, sem a presena de quaisquer rgos respiratrios especiais e na ausncia de circulao sangunea. Os animais maiores e mais complexos, frequentemente, apresentam superfcies especializadas para a troca de gases e tambm um sistema sanguneo para transportar o oxignio mais rapidamente que o processo de difuso sozinho poderia faz-lo.
Animais sem rgos respiratrios especiais
A forma geomtrica mais simples de um organismo a esfera. Para as consideraes aqui feitas importante que uma esfera tenha a menor superfcie possvel em relao a um dado volume; qualquer desvio da forma esfrica implica num aumento relativo da rea da superfcie. Se for admitido que um organismo esfrico deva ser suprido de oxignio por difuso atravs da superfcie e da para todas as pores do corpo, a mais longa distancia de difuso a da supercie para o centro. A concluso que o organismo, para ser suprido de oxignio, deve ser bem menor ou apesar uma taxa metablica bem mais baixa.
Um organismo que no apresenta a forma esfrica tem uma superfcie relativa maior e uma distncia maior, uma difuso menor que uma esfera. Isso valido para uma variedade de organismo simples. Eles so achata dos ou filamentosos, tem pseudpodes, ou superfcies bastante extensas ou complexas (tais como, corais e esponjas) e podem assim obter oxignio suficiente por difuso.
Animais com rgos respiratrios
Apesar de um organismo pequeno pode seguir oxignio suficiente por difuso atravs da superfcie, geralmente isso no vale para organismos maiores. claro que qualquer forma que no seja esfrica tenha uma superfcie maio e as distancias de difuso so tambm reduzidas. Contudo, na maioria dos casos isso no suficiente e so observados rgos respiratrios especializados com superfcies muito aumentadas.
Frequentemente, esses rgos exibem uma cutcula mais fina do que nas outras partes do corpo, facilitando assim a troca de gases. Se a superfcie respiratria estiver voltada para o lado esterno, formando uma evaginao o rgo restante normalmente chamado de brnquia. Secundariamente uma brnquia pode estar no interior de uma cavidade, tal como um peixe, mais isso no muda o fato de que seja fundamentalmente evaginao. O termo pulmo usado tanto se o meio for respiratrio for a gua como o ar. Os insetos tm uma forma especial de sistema respiratrio. Pequenas aberturas na superfcie corprea do inseto uniram-se a um sistema de tubos (traquias) que se ramifica e se distribui para todas as partes do corpo. Neste caso, o rgo respiratrio combina com sistema de distribuio (os tubos) com o sistema de troca de gases, pois a maior parte do gs passa atravs
das paredes das ramificaes mais finas deste sistema e difundi-se diretamente para as clulas. Em geral, as brnquias servem principalmente para a respirao aqutica e os pulmes para a respirao area. Existem excees: os pepinos do mar tm pulmes equferos nos quais parece ocorrer a maior parte da traa de gases. As brnquias podem tambm ser modificadas para uso o ar, mas em geral so inadequadas para a respirao area.
Ventilao das brnquias
Se uma brnquia remover oxignio de gua completamente estagnada, a camada estacionaria de gua, imediatamente adjacente, logo ter o seu contedo de oxignio esgotado. A renovao desta gua , portanto, importante para o suprimento de oxignio e vrios dispositivos mecnicos servem para aumentar o fluxo de gua sobre a superfcie da brnquia. O fluxo aumentado pode ser conseguido de duas formas: pela movitaao da brnquia pela gua ou da gua sobre a brnquia. A movimentao da brnquia atravs da gua vivel apenas para pequenos organismos. Algumas larvas de insetos aquticos larvas de insetos da famlia Ephaemeridae, por exemplo ventilam suas branqueias desta forma. A dificuldade reside no fato de que, se a branqueia movida atravs da gua, com sua base servindo como um ponto de piv, a fora necessria para superar a resistncia ao movimento muito grande. Essa resistncia aumenta com o quadrado da velocidade linear do rgo e, portanto, a energia necessria para a movimentao da branqueia aumenta na mesma proporo. A fora mecnica desenvolvida pela brnquia tambm precisaria ser aumentada, novamente com o quadrado da velocidade linear, como ocorre com a fora aplicada junto base da mesma para faz-la movimenta atravs da gua. A grande salamandra aqutica norte americana
conhecida como mudpuppy (Netunos) movimenta suas brnquias, mas muito lentamente. A movimentao da gua sobre a superfcie respiratria uma soluo muito mais factvel. O movimento pode ser realizado pela ao de clios, como nas brnquias dos mexilhes e mariscos. As esponjas movimentam a gua atravs de seus hostes pela ao de flagelos. A movimentao da gua por meio de dispositivo semelhante a uma bomba mecnica mais comum. Os peixes e caranguejos, por exemplo, movimentam dessa forma a gua sobre suas brnquias. Na realidade, menos dispendioso movimentar a gua lentamente sobre uma grande superfcie que mov-la rapidamente sobre uma superfcie menor.
Outras funes das brnquias
As brnquias podem ter outras funes alm de trocar gases respiratrios e, s vezes, fica difcil decidir qual a funo principal ou nica, de uma brnquia algumas, chamadas de brnquias anais de larvas de mosquito, agem na regulao osmtica, absorvem os ons da gua e duvidoso que exeram qualquer papel importante na respirao. As brnquias de peixes e caranguejos ajudam na regularizao osmtica, mas nestes casos bastante claro que elas tambm possuem como funo principal a respirao. Para avaliar o papel nas trocas gasosas de uma brnquia, ou outro eventual rgo respiratrio, devem-se obter informaes sobre a quantidade de oxignio captada, de preferncia sobre o dixido de carbono liberado por meio deste rgo. Ao comparar isso com o consumo de oxignio de todo o organismo, possvel verificar se o rgo responsvel por toda a captao de oxignio, uma grande parte, uma pequena parte ou uma quantidade trivial.
Troca gasosa e fluxo de gua
O fato de que a rea de superfcie de uma brnquia deva ser grande o suficiente para prover uma troca de gases adequada bem definido nos peixes. As brnquias dos peixes alternamente ativos apresentam as maiores relativas. A rea de superfcie da brnquia da cavala que nada rpido expressa por unidade de peso, aproximadamente 50 vezes maior que a do lento peixe-ganso que vive no fundo do mar. Para que as trocas gasosas sejam adequadas, so necessrio uma alta velocidade de fluxo de gua e um estreito contato entre gua e brnquia. Isto conseguido por meio da estrutura anatmica do aparelho branquial. As brnquias encontram-se no interior da cavidade branquial, a qual d proteo a estes frgeis rgos e permite, de forma igualmente importante, que a gua passe sobre estas da maneira mais afetiva possvel. Um vantagem especial conseguida por meio de um arranjo que faz com que a corrente de gua sobre a brnquia e o sangue no interior dela flutuam em direo oposta, tal denominado contracorrente.
Fluxo contracorrente
As brnquias dos peixes consistem de diversos arcos branquiais principais de cada lado. De cada arco branquial, estendem-se duas fileiras de filamentos branquiais. As pontas desses filamentos, proveniente de arco adjacente, encontram-se forando a gua e fluir entre os filamentos. Cada filamento possui lamelas achatadas densamente enfileiradas. As trocas gasosas ocorrem nessas lamelas medida que a gua flui entre elas em direo ao sangue, na direo oposta. Esse tipo de fluxo contracorrente tem uma importante consequncia. Quando o sangue est saindo da lamela branquial, e ele encontra gua cujo oxignio ainda no foi removido. Dessa forma, esse sangue obtm oxignio da gua que tem ainda o contedo de oxignio da gua inspirada, permitindo que o contedo de oxignio do sangue atinja p nvel mais alto possvel.
medida que a gua corre entre as lamelas, ela encontra o sangue com o
contedo cada vez mais baixo de
oxignio e, portanto, continua a liberar
mais oxignio. Assim, a lamela, ao longo de toda sua extenso, serve para captar o oxignio da gua, esta, por sua vez, pode deixar a brnquia tendo perdido de 80 a 90% de seu contedo inicial de oxignio. Isso considerado uma extrao bastante alta de oxignio; por outro lado, os mamferos removem apenas aproximadamente um quarto do oxignio presente no ar dos pulmes antes que seja expirado. O fluxo contracorrente tambm encontrado nas brnquias de alguns caranguejos, mas neles a eficincia de remoo de oxignio da gua frequentemente muito menor que nos peixes. Em parte, isso ocorre porque o fluxo de gua menos efetivo, mas provvel que seja devido principalmente maior barreira de difuso brnquia-sangue.
Bombeamento da gua
Para mover a gua sobre as brnquias, os peixes telesteos fazem o uso do bombeamento combinado da boca e dos oprculos, com auxilio de vlvulas adequadas para controlar para controlar o fluxo. Em realidade, o sistema consiste de duas bombas. O volume da primeira, a cavidade bucal, pode ser aumentado pelo abaixamento da mandbula e especialmente do assoalho da boca. O volume da segunda, a cavidade opercular enquanto o refluxo de gua ao longo das bordas impedindo por um prolongamento de pele que atua como uma vlvula passiva. A ao das duas bombas tal qual um fluxo de gua atravs das brnquias mantido aproximadamente por todo o ciclo respiratrio. Esse fluxo continua embora a presso na boca, durante parte do ciclo, possa ser menor que na gua circundante; a razo simplesmente porque a presso nas cavidades operculares mantida mais baixa na boca.
Ventilao forada Alguns peixes so incapazes de respirar dessa forma. Bilogos que estudam peixes, h muito sabem que os grandes atuns no conseguem ser mantidos vivos em cativeiro a menos que possam nadar continuamente; isso pode ser conseguido se forem mantidos em grandes tanques redondos onde
podem nadar ininterruptamente.
O peixe nada com a boca parcialmente
aberta, no a movimento respiratrio visveis e a gua flui continuamente sobre as brnquias; isso chamada de ventilao forada. A ventilao forada no restrita aos peixes pelgicos grandes, que nadam rapidamente. Muitos peixes respiram por bombeamento a velocidade baixa e passam para a ventilao forada a velocidade mais alta. A passagem para a ventilao forada no significa que as brnquias sejam ventiladas sem custo; isso apenas significa que o trabalho da respirao transferido dos msculos das bombas operculares para os msculos natatrios do corpo e da cauda. Aboca aberta faz com que a fora de arrasto aumente o que deve ser compensado por um maior trabalho muscular. Contudo, o fluxo continuo durante a ventilao forada mais econmico do ponto de vista energtico que o bombeamento opercular no ritmo elevado necessrio para a natao rpida.
Respirao area
Os animais que respiram o ar tem trs tipo de principais de rgos respiratrios: brnquias, pulmes e traquias. Brnquias: em geral, as brnquias so muito pouco adequadas respirao area e so usadas apenas por poucos animais principalmente alguns que num perodo relativamente recente invadiram o habitar terrestre, trazendo consigo os vestgios de seus modos prvios de respirao aqutica. Os caranguejos terrestres so bons exemplos. O caranguejo coco (birgus latro) que adotou uma existncia terrestre quase completa (incluindo a escala de coqueiros) possui brnquias que so suficientemente rgidas para continuarem funcionais durante a respirao area. Entre os peixes que conseguem respirar o ar, as brnquias funcionais fora preservadas em alguns, mas no em todos. A enguia comum (Anguilla vulgaris) sobrevive muito bem fora da gua se for mantida em ambiente razoavelmente frio e mido. A maior parte do oxignio obtida atravs da pele e uma menor guantidade atravs das brnquias, pois os filamentos tende a aderir-se, deixando exposta uma pequena superfcie em contato com
o ar contido na cmara. Consequentemente, a enguia no obtm o seu suprimento normal de oxignio e a tomada de oxignio do ar reduzida para apenas cerca da metade do ocorre na gua. Pulmes. Podemos distinguir dois tipos de pulmes: pulmes de difuso e de ventilao. Os pulmes de difuso so caracterizados pelo fato de que a troca de ar com a atmosfera se da apenas por difuso. tais pulmes so encontrados em animais relativamente pequenos, como caracois, escorpies e algumas ispodes. Os pulmes de ventilao so tpicos dos vertebrados. Uma renovao substancial e regular do ar no pulmo necessria para animais de grande tamanho corpreo, combinada com uma elevada taxa metablica. Os sistemas dos vertebrados so ventilados por fluxos de ar que entra e sai, ou fluxo corrente. Entretanto, o sistema respiratrio das aves que mais complexo que dos mamferos, organizado de tal modo que o ar possa fluir numa mesma direo atravs do pulmo durante a inspirao e a expiraao.
Traquias.
Esse tipo de rgo respiratrio e caracterstico dos insetos.
Consiste de um sistema de tubo que fornece oxignio diretamente para os tecidos, evitando assim necessidade de uma circulao sangunea de uma finalidade de transportar gases as trocas gasosas no sistema traqueal podem ocorrer apenas por difuso, mas em muitos insetos, especialmente naqueles com atividade elevada, existe um bombeamento ativo e unidirecional de ar em partes do sistema traqueal. A vantagem do fluxo unidirecional permitir uma troca de gases muito melhor que a obtida pelo bombeamento de ar para dentro e para fora do organismo.
Movimentos respiratrios
Os pulmes dos vertebrados so ventilados pelo bombeamento ativo do ar. A ventilao pulmonar dos vertebrados pode ser obtida de duas formas diferentes. O enchimento pulmonar pode se dar com a utilizao de uma
bomba de presso, como nos anfbios, ou por meio de uma bomba de suco, como na maioria dos repteis, pssaros e mamferos. O mecanismo normal para a insuflao dos pulmes dos repteis emesmo das aves e dos mamferos: os pulmes so inflados por suco. A expirao, em seguida a uma inspirao, pode ser passiva, em decorrncia do recolhimento elstico, ou pode ser ativamente auxiliada pela contrao muscular. Uma bomba de suco que requer uma cavidade torcica fechada na qual a presso durante a inspirao seja menor que a preo atmosfrica. Nos mamferos a inspirao auxiliada pela contrao do diafragma muscular. As aves tm um diafragma membranoso preso parede do corpo com o intemdio do msculo, mas sua funo difere daquela do diafragma dos mamferos. A afirmao de que as aves tm diafragma incorreta.
O papel da pele na respirao
A troca de gases atravs da pele normal e importante para os anfbios, que tem pele mida e bem vascularizada. De fato, algumas pequenas salamandras no tm pulmes e toda a troca de gases ocorre por meio de superfcies da pele, exceto por uma pequena contribuio mucosa. Nas rs, os papeis relativos da pele e pulmes mudam durante o ano. No inverno, quando a tomada de oxignio muito baixa, a pele transfere mais oxignio do que os pulmes. No vero, quando o consumo de oxignio alto, a tomada atravs dos pulmes aumenta diversas vezes e excedem muito a tomada cutnea. O fato de que a tomada de oxignio atravs da pele permanece quase constante quase o ano todo est relacionado com a concentrao constante de oxignio na atmosfera, que permite a existncia de uma diferena de presso de difuso constante se a concentrao de oxignio no sangue permanecer uniformemente baixa durante o ano a difuso atravs da pele no devera ser muito modificada, pois a taxa de difuso ser pouco alterada com a temperatura. Com o grande aumento da demanda no vero, a pele sozinha no conseguiria lidar com tal acrscimo
havendo portando necessidade de tomada adicional pelos pulmes como de fato ocorre. Acredita-se com frequncia que os repteis, em contraste com os anfbios, possuem uma pele quase impermevel. Eles respiram com os pulmes e a maioria deles terrestre. Contudo, as serpentes marinhas verdadeiras so de fato marinhas e algumas suportam at viver no mar quando jovens. Elas so excelentes nadadoras e capazes de mergulhar a uma profundidade de menos 20 m. Quando a serpente marinha Pelamis platurus fica submersa, consegue obter oxignio atravs da pele a uma proporo de at 33% de sua captao padro total de oxignio, e excreta dixido de carbono a uma proporo de mais de 94% do total. Apesar de o pulmo ser o principal rgo de troca gasosa, a pele evidentemente de considervel auxilio na troca de gases quando esta serpente vai ao alcano do peixe pequeno para predlos. Nos mamferos, a troca de gases atravs da pele pouco importante. O morcego tem uma superfcie cutnea relativa muito maior que a dos outros mamferos; as membranas das asas, que so grandes, finas e sem plos, so altamente vascularizadas e podem contribuir para a troca de gases. A captao de oxignio atravs das membranas das asas, contudo, no suficientemente grande para ser significativa; como discutido anteriormente, a difuso entre a gua e o ar de cerca de 25 vezes mais lenta para dixido de carbono que para o dixido de carbono.
Os pulmes dos mamferos
medida que se progride em direo a outras classes de vertebrados, observa-se que os pulmes tornam-se cada vez mais complexos. Nos anfbios, o pulmo o nico saco, subdividido pela trabcula que aumentam a superfcie. O pulmo dos mamferos muito mais amplamente subdividido em pequenos sacos delicados, os alvolos, que aumentam enormemente a rea de superfcie disponvel para as trocas gasosas. Medidas da rea da superfcie do pulmo da r indicam que 1 cm (um centmetro cbico) do
pulmo da r indica 1 cm3 (um centmetro cbico) de tecido pulmonar tem a superfcie total de trocas gasosas de 20 cm sendo que o valor correspondente para um camundongo normal acima de 800 cm para cada m de tecido pulmonar. A grande rea da superfcie essencial para a alta taxa de tomada de oxignio necessria para manter a elevada taxa metablica de animais de sangue quente. A membrana que separa o ar do sangue nos pulmes de ser fina, de modo que o oxignio possa difundi-se rapidamente para o sangue, enquanto o dixido de carbono se move na direo oposta.
Volume pulmonar
O volume pulmonar de um mamfero constitui aproximadamente 5% do volume corpreo, independentemente do peso corpreo.
Inspirao e expirao
A troca gasosa no pulmo ocorre nos alvolos; a traquia, os brnquios e suas ramificaes so apenas tubos repletos de ar usado do pulmo e, quando e, seguida ocorre a inspirao, este ar puxado novamente para o interior dos pulmes antes que o ar fresco externo entre. O volume de ar nas vias respiratrias reduz a quantidade de ar fresco que entra no pulmo chamado de espao morto. O volume de ar inalado em uma nica respirao o volume corrente. Um aspecto importante da respirao que os pulmes nunca so completamente esvaziados do ar. Mesmo se um indivduo expirar o maximo de ar possvel, restaro, aproximadamente, 1.000 cm (mil centmetro cbicos) de ar em seus pulmes.
Tenso superficial
Qualquer um que tenha soprado bolhas de sabo sabe que, quando a conexo com a atmosfera esta aberta, a bolha tende a ser contrair, expelindo at colapsar. O pulmo dos vertebrados algo semelhante; os alvolos com
formato de bolha e grande curvatura indicam que a tenso superficial na face interna midas tende a fazer com que as bolhas se contraiam e desapaream. A tenso superficial deveria causar o colapso do pulmo, mas essa tendncia minimizada pela presena, na superfcie interna dos alvolos, de substncias que reduzem muito a tenso superficial. Essas substncias so fosfolipdios, cujo efeito sobre a tenso superficial lhes conferiu o nome de surfactante. Os surfactantes so encontrados nos pulmes de todos os vertebrados - mamferos aves e anfbios. Mesmo a superfcie do pulmo do peixe pulmonado recoberta com surfactantes. A quantidade de surfactantes presente no pulmo dos vertebrados parece estar sempre acima do mnimo necessrio para recobrir a superfcie pulmonar com uma camada monomolecular.
Trabalho mecnico de respirao
Para movimentar o ar para dentro e para fora do pulmo h necessidade de trabalho de interesse fazer a comparao entre o custo do bombeamento e a qualidade de oxignio provida. O mtodo mais simples comparar a quantidade para acionar a bomba com a qualidade de oxignio que o organismo obtm no mesmo perodo. As determinaes do trabalho respiratrio so difceis e apresentam uma margem substancial de erro, porque a esta qualidade pequena quando comparada com o consumo total de oxignio. A maioria das determinaes indica que o custo da respirao em um indivduo, durante o repouso, de 1,2% do consumo total de oxignio. Com o aumento da ventilao, o custo da respirao tambm se eleva e, nos exerccios mais intensos, ela pode chegar a 3% do total de oxignio consumido.
Sincronizao da respirao com a locomoo
Quando um canguru percorre rapidamente um plancie australiana com seus saltos longos, a sua taxa respiratria sincronizada com os saltos em pequena proporo de 1:1. O mecanismo bastante simples. No comeo de um salto, enquanto o corpo acelerado para frente e para cima, os rgos moles da cavidade abdominal so forados na direo posterior pela inrcia isso, Poe sua vez, fora o diafragma e os pulmes se enchem de ar. Quando no final do salto a perna toca o cho, as vsceras so foradas contra o diafragma e o ar forado a sair do pulmo. como se as vsceras agissem como um pisto, movimentando o ar para dentro e para fora dos pulmes. A sincronizao entre a respirao e a locomoo tem sido observada em muitos mamferos e aves, mas nem sempre fcil analisar este mecanismo. As aves no possuem diafragma muscular e dependem dos movimentos do esterno para ventilao para ventilao. J que os msculos que permitem o vo so unidos ao esterno, parece razovel que os movimentos das asas influenciem a ventilao; em muitas aves, existe, na realidade, um elo entre o bater das as e a respirao. Em algumas espcies, esta proporo de 1; 1, mas em outras ela pode ser 2:1 ou 2:3 e em outras ainda a proporcionalidade pode nem existir. Os motivos de tais diferenas so facilmente explicados. No apenas os cangurus, mais diversos outros mamferos, mostra um sincronizao entre respirao e a locomoo.
Regulao da respirao
Se a demanda de oxignio aumentar, a ventilao dos rgos respiratrios devera ser aumentada de modo compatvel. Da mesma forma, se for
reduzida a concentrao de oxignio do meio, dever haver compensao por meio do aumento da ventilao, ou pelo aumento da qualidade de oxignio removido do ar respirado, ou por ambos. interessante notar que nos vertebrados de sangue quente mamferos e aves a ventilao pulmonar regulada de forma bastante precisa em relao a demanda de oxignio, ma o principal agente responsvel pela regulao a concentrao de dixido de carbono no ar. Isso facilmente
demonstrado pela edio de dixido de carbono ao ar inspirado, o que causa um rpido aumento na ventilao pulmonar. O ar atmosfrico normal quase no conte dixido de carbono (0,03%) e se 2,5% de CO2 forem adicionados ao ar inspirado, o volume ventilatorio aproximadamente dobrado. O efeito semelhante tento nos mamiferos como nas aves.
O centro respiratrio
Os movimentos respiratrios nos mamferos so realizados pelos msculos do diafragma e intercostais. Os movimentos rtmicos desses msculos so controlados pelo centro respiratrio, que est localizado na rea da medula alongada e da ponte no celebro. O centro respiratrio contm grupos separados de neurnios para a inspirao, que funcionam de forma alternada. Quando os neurnios inspiratrios ecitam os msculos da respirao, impulso inibitrio junto aos neurnios envolvidos expirao mantm os msculos inativos. Quando a atividade dos neurnios inspiratrios cessa, os neurnios expiratrios tornam-se ativos e dessa forma a ao alternadas dos dois grupos continua indefinidamente. O centro respiratrio sensvel ao aumento do dixido de carbono ou ao aumento da acidez no liquido cerebroespinhal, sendo que um aumento do PCO2 no sangue faz com que o dixido de carbono difunda-se rapidamente do sangue para o liquido cerebroespinhal.
Animais aquticos
Tem se afirmado que foca e baleias so menos sensveis ao dixido de carbono que outros animais e, portanto, so capazes de ficar debaixo da gua por mais tempo. Contudo, a durao de um mergulho provavelmente limitada pela quantidade de oxignio disponvel e pouco provvel que um
mergulho possa ser prolongado meramente por uma diminuio na sensibilidade ao dixido de carbono. Caso se queira examinar a resposta ao dixido de carbono inspirado em diversos animais, no suficientemente fazer a medida da frequncia respiratria. Em alguns animais, afreguencia respiratria aumenta
consideravelmente em resposta ao dixido de carbono, mais em outros pode haver pouca, ou nenhuma, alterao na frequncia. O volume corrente pode toda via aumentar consideravelmente. Isso tem sido observado, por exemplo, na equitina (tachyglossus). A informao necessria o volume ventilatorio (ou seja, o produto entre a frequncia respiratria e o volume corrente).
Comparao entre as respiraes area e aqutica
Em muito invertebrados aquticos, a regulao da respirao precria ou mesmo ausente. Isso valido, em especial nas espcies marinha que normalmente vivem em gua bem arejadas, com um suprimento de oxignio relativamente constante. Alguns animais inferiores so bastante tolerveis a falta de oxignio; os moluscos bivalves podem manter as conchas fechadas durante longos perodos e, na ausncia de ventilao, utilizam processos metablicos anaerbios. Para a maioria dos animais aquticos, o principal estmulo a respirao a falta de oxignio. Isto comum nos crustceos, polvos, peixes, etc. o efeito de dixido de carbono nos invertebrados aquticos nunca muito pronunciado e pode estar ausente. A tenso de dixido de carbono na gua natural que quase sempre baixa e, devido a alta solubilidade do dixido de carbono na gua, os animais aquticos no tm como desenvolver uma alta tenso deste gs. Se esses animais dependessem de um aumento da tenso de dixido de carbono para o estimulo respiratrio, no seria possvel a segurar um suprimento adequado de oxignio.
Peixes e respirao area
Muitos peixes responderam diminuio do contedo de oxignio na gua nadando para a superfcie para sorver um bolha de ar pela boca, uma resposta que melhora enormemente o suprimento de oxignio. Um peixe dourado mantido na gua em uma tenso de oxignio de apenas 20 mmHg engole ar na superfcie e a uma frequncia de 1800 x por hora, o que quase dobre o contedo de oxignio do sangue arterial do peixe carente de oxignio. Outros peixes so especificamente mais bem adaptados a respirao area que o dourado. Os peixes pulmonados so bem conhecidos, mas no so os nicos que dependem de ar para respirar. Muitos peixes recorreram a respirao area apenas quando o contedo de oxignio na gua for baixo; relativamente, poucos depende de ar a tal ponto que, se forem mantidos submersos, acabaram sendo afogados. Existem duas razoes ecolgicas principais para a respirao area acessria: (1) depleo do oxignio na gua e (2) a ocorrncia de secas peridicas. Um peixe pulmonado, por exemplo, durante os perodos de
secas, enterra-se na lama, encerra-se dentro de um casulo e permanece inativo at a prxima cheia. A maioria dos peixes de respirao area de gua doce tropical ou pertence a espcies estuarinas; poucos so realmente marinhos. A gua doce deficiente em oxignio muito mais comum nos trpicos de que em climas temperados. Isso ocorre porque existem muitas substncias orgnicas em decomposio na gua, a temperatura alta e acelera a ao bacteriana, pequenos corpos de gua so frequentemente muitos sombreados pela floresta (que conduz a fotossntese e a produo de oxignio na gua), e a uma pequena alterao na temperatura entre o dia e a noite e, portanto, uma conveco trmica mnima para levar a gua rica de oxignio da superfcie para as camadas mais profundas. Contudo, nem todos os peixes de respirao area pertencem aos trpicos. O conhecido peixe de gua doce Amia calva encontrado no norte dos Estados Unidos, mesmo onde os lagos ficam congelados durante o inverno. Durante tais perodos, eles vivem bem sem respirar o ar porque a temperatura baixa reduz seu consumo de exigindo.
Foi explicado que as brnquias dos peixes no so muito adequadas para a respirao no ar. Falta-lhes a rigidez necessria e elas tendem a aderir umas as outras, mais mesmo assim uma pequena quantidade de oxignio pode ser obtida por meio das brnquias no ar. Qualquer outra superfcie mida pode suplementar as trocas gasosas, contanto que seja uma superfcie que tenha acesso ao ar e seja suprida de sangue. Algumas trocas gasosas podem ocorrer atravs da pele e da superfcie da cavidade bucal, mas, alm disso, pode haver outros rgos anatomicamente mais especializados que auxiliam nas trocas gasosas. Os rgos comumente utilizados na respirao area so: brnquias, pele, boca, cavidades operculares, estomago, intestino, bexiga natatria e pulmo.
Peixes pulmonados
Os peixes pulmonados africanos e sul americano (protopterus e lepidosiren) foram relacionados como sendo de respirao area obrigatria. Eles vivem em guas paradas e em lagos, onde a falta prolongadas de chuvas podem causar casamento completo de seu habitat. Eles estivam at o prximo perodo de chuvas , quando saem de seus casulos, enterrados na lama, e retornam vida normal. O peixes pulmonados australianos (neoceratodus) vivem em rios e em cursos de guas lentas. eles tambm estiva nos perodos secos mais depende muito menos do pulmo e respira principalmente pelas brnquias. Os peixes pulmonados esto entre os mais estudados peixes de respirao area. As informaes sobre muitos outros so, ainda, insuficiente ou virtualmente nulas. O estudo desses peixes poderia ser bastante
recompensador apesar de ser com frequncia difcil do ponto de vista tcnico, pois muitos so bem pequenos.
A respirao das aves
A estrutura do sistema respiratrio
Os rgo respiratrios da aves so bastantes diferentes da aqueles dos mamferos. os pulmes pequenos e compactos das aves comunicam se com sacos areos volumosos, de paredes finas e espaos de ar que se estende entres os rgos internos, ramificando se no interior dos ossos das extremidades e do crnio. Pode-se dizer que a presena de espao de ar no corpo de uma ave a tornar mais leve, mais apenas no sentido bem limitado. As aves precisam de um sistema digestivo, fgado, rins, etc., e apenas acrescentar grandes sacos areos atividade abdominal no as tornam mais leves. Se esse ar for removido, ou seu tamanho, dobrado, as aves teriam exatamente o mesmo peso para carregar durante o vo. por outro lado, se o tutano do osso for substitudo por volume idntico de ar, o osso pesara menos. os ossos preenchidos de ar, portanto contribuem para tornar as aves mais leves, mais o mesmo no o correm em outro espaos de ar. Se for feita comparao entre os volumes dos sistemas respiratrios de aves e mamferos, verificam se algumas diferenas notveis. O volume do pulmo de uma ave tpica apenas uma pouco maior que ametade do de um pulmo de um mamfero do mesmo tamanho corpreo. Em contraste o volume traqueal muito maior do que de um mamfero. Isso pode ser facilmente entendido, tendo em vista o longo pescoo das aves. Os sacos areos de uma ave so grandes, muitas vezes do tamanho de um pulmo, e os mamferos no possuem sacos areos. Portanto, o volume total do sistema respiratrio de uma ave trs vezes maior o de uma mamfero. A diferena entre aves e mamferos no est restrita a sacos areos; em termos estruturais, pulmes das aves diferentes radicalmente daqueles dos mamferos. Nesse, ramificaes mais finas dos brnquios terminam nos alvolos. Nas aves, nas ramifies mais finas do sistema bronquial (conhecidas como parabronuquios) permitem a passagem continua de ar. Assim o ar pode fluir atravs do pulmo das aves, passando continuamente pela superfcie de troca; nos mamifero o ar deve fluir para dentro e para fora. Essa a diferena mais importante entre o sistema respiratrio das aves e dos mamferos, o que acarreta importantes consequncias fisiolgicas.
importante, contudo, saber que os sacos areos formam, anatmica e funcionalmente, dois grupos: um grupo posterior ou caudal, que inclui os grandes sacos abdominais, e um grupo anterior ou craniano, que consiste em diversos sacos menores. A traqueia divide-se em dois brnquios: cada brnquio alcana um dos pulmes e atravessa-o, terminando no saco abdominal. Os sacos cranianos ligam-se a esse brnquio principal, na parte anterior do pulmo; os sacos caudais ligam-se parte posterior do brnquio principal. O brnquio principal tambm se liga ao pulmo e, alm disso, alguns dos sacos areos ligam-se diretamente ao tecido pulmonar.
Funo do sistema respiratrio
A morfologia dos sacos areos no indica que eles desempenho qualquer papel importante nas trocas gasosas entre o ar e o sangue. Suas paredes so finas, frgeis, pouco vascularizada, no existindo dobras ou salincias para aumentar a rea da superfcie. Uma hiptese mais plausvel para a funo dos sacos areos que eles funcionem como um fole para movimentar o ar para dentro e para fora. No inicio de uma inspirao, a queda simultnea da presso nos sacos craniano e caudal medida que esses se expandem. Isso significa que, durante a inspirao, o ar flui para o interior dos sacos, mas, como ser visto a diante, nem todos os sacos so preenchidos com ar externos. Durante a expirao, a presso nos sacos aumenta e o ar flui para fora novamente. O fluxo de ar no pulmo das aves no de fato um sistema de troca contracorrente ideal, mais muito mais um fluxo do tipo corrente cruzada. O resultado, com respeito as tenses do gs arterial semelhante aquele acima discreto, apensar de o sistema decorrente cruzada no ser efetivo quando um verdadeiro sistema contracorrente em relao a obteno do proveito Maximo nas trocas gasosas,
Altitude: a efetividade do fluxo de ar unidirecional no pulmo das aves particularmente importante em grandes altitudes. O padro do fluxo pulmonar das aves explicao mais plausvel, pois permite que o sangue remova o oxignio de um ar que tem uma concentrao deste gs mais alta que a encontrada no sistema respiratrio dos mamferos. Alm disso, por causa do fluxo unidirecional e continuo do pulmo, uma quantidade maior de oxignio pode ser extrada do ar.
O canto do canrio
Muitos de ns, ouvindo o canto de um canrio macho, j se perguntaram como possvel como um pssaro to pequeno cante continuamente at meio minuto, aparentemente sem fazer nenhuma pausa para respirar. Como esta ave consegue produzir cantos to extensos, tendo em vista a capacidade limitada de seu sistema respiratrio? A resposta que o canto do canrio sempre produzido durante o fluxo do ar expiratrio na traquia. Ele consiste em notas ou em silabas simples, repetida em alta velocidade, cada uma delas dura durante 11 a 280 milissegundo. Entre essas silabas ou notas existem breves intervalos silenciosos, que duram de 20 a 35 MS, durante o tempo em que ocorre a inspirao. Em outras palavra, o canto do canrio no e contnuo, e sim consiste de uma srie extensa de slabas, com pausas to curtas que as no percebemos.
A respirao nos ovos
O ovo recm posto um microcosmo que contm tudo que necessrio para produzir um pintinho vivo, com uma exceo-oxignio. Portanto, a casca deve ser permevel ao oxignio, e consequentemente, tambm a outros gases, pois, o vapor de gua e dixido de carbono so liberados durante a incubao. A casca de um ovo consiste de uma camada externa rgida de carbonato de clcio, que no lado interno possui duas membranas moles, chamadas de
membranas externa e interna da casca. A casca rgida muito menos permevel aos gases que as suas membranas. No inicio, quando o ovo posto, a permeabilidade da casca com suas membranas e baixas, mais aumenta diversa vezes durante os primeiros das aps a postura. Esse aumento esta correlacionado com declnio no contedo de gua das membranas e, a partir da a maior parte resistncia a difuso e consequncia da casca rgida. Na extremidade romba do ovo a uma cmara de ar, que fica entre as membranas internas e externas da casca. Esse espao de ar aumenta de tamanho durante a incubao porque a gua perdida pelo ovo por evaporao. O consumo de oxignio e a produo de dixido de carbono aumentam durante todo perodo de incubao, que dura aproximadamente 21 dias. Como as caractersticas de difuso da casca do ovo, durante a ltima parte de incubao, permanecem constantes, ou quase, deve haver um aumento progressivo do gradiente dos gases em que o ar ambiente e o embrio. Os gradientes mais exagerados so produzidos por uma queda progressiva da presso parcial do oxignio dentro do ovo e um aumento concomitante de dixido de carbono durante a incubao. Durante o ltimo dia de incubao, rapidamente ocorre modificaes. Aproximadamente 28 horas antes do rompimento da casca, o pintinho perfura a membrana da cmara de ar, o incio da respirao pulmonar e ele respira novamente o gs contido no espao de ar (que, devido a evaporao, aumenta seu volume em aproximadamente 10 cm). Cerca de 12 horas depois, o pintinho comea a quebrar a casca, estgio conhecido como pipping, a principal barreira as trocas gasosas neste momento anulada. Durante as horas seguintes, enquanto o pintinho se esfora para sair completamente da casca, a respirao pulmonar assume toda a tarefa das trocas gasosas.
Os ovos dos rpteis Os ovos dos reptis so muitos mais variveis que os das aves. A estrutura do mesmo pode variar desde os que apresentam casca finas, elsticas, parecidas com membranas, ate aqueles com casca rgida
calcrias, mais parecida com os ovos das aves. essa diferencia esto relacionadas com o habitat e especialmente com as condies de umidade sob as quais os ovos so postos. Os ovos dos rpteis podem ser distribudos em trs grandes categorias. Os ovos da maioria dos largartos e de todas as cobras so relativamente fina e altamente elstica. Tais ovos so normalmente depositados em um ambiente relativamente seco e, seu meio for mido tende a reter gua e aumentar de tamanho. Esses ovos so muitos sensveis umidade caracterstica do ambiente e a capacidade de ruptura da casca depende de condies adequadas. Em contraste as tartarugas possuem ovos com as casca um tanto flexveis com uma camada calcaria bem definida. So relativamente independentes das condies de unidade, as quais no afetam de forma excessiva a capacidade de ruptura da casca. Os ovos de todos os crocodilos, de algumas tartarugas, e de alguns poucos largatos, so duros e rgidos. Esses ovos, portanto, no incharam excessivamente em ambientes midos, tais com os ninhos utilizados por jacars e crocodilos.
A respirao dos insetos
A vida terrestre cria um conflito continuo entre a demanda de oxignio e a necessidade de gua. As condies que favorecem a entrada de oxigenio tambm favorecem a perda de gua. Os insetos, os animais terrestres de maior xito, tm uma cutcula rgida que altamente impermevel, aos gases e cuja camada de cera a torna virtualmente impermevel a gua. As trocas gasosas ocorrem por meio de um sistema de tubos internos repletos de ar, as traqueias, que se comunicam com o exterior por aberturas denominadas espiraculos. Os espiraculos, normalmente, tm um mecanismo de fechamento que permite um controle acurado da trocas entre o ar no sistema traqueal e atmosfera.
As traqueias se dividem, ramificam-se e estendem-se para todas as partes do corpo. As ramificaes mais finas, as traqueias, as traquolas, de cerca de 0,5, um ou menos de um dimetro, podem at se estender para o interior de clula individuais, tais como fibras musculares. As traqueias no podem ser menores porque a difuso impedida quando seu tamanho se aproxima do trajeto livre mdio das molculas no ar (a distncia mdia que uma molcula percorre antes de colidir com outra). O sistema traqueal conduz o oxignio diretamente para os tecidos e os dixidos de carbono em direo oposta. Isso torna a respirao dos insetos independente de um sistema circular. O sangue nos insetos no desempenha um papel direto no transporte de oxignio, ao contrrio que acontece nos vertebrados. A comparao com os vertebrados no de forma alguma irrelevante, pois as taxas de consumo de oxignio de uma grande mariposa e um beija-flor durante o vo so semelhantes; no entanto, os mecanismos de suprimento de oxignio difere radicalmente. No se pode concluir que o sistema circulatrio nos insetos no seja importante ele possui muitas outras funes. Um papel bvio. Durante o vo, os msculos desenvolvem uma alta potencia e, enquanto o oxignio provido pelo sistema traqueal o combustvel deve ser suprido pelo sangue.
O sistema traqueal
Os sistemas traqueais so apropriados principalmente para a respirao no ar e devem ter evoludo no meio areo os insetos que secundariamente se tornaram aquticos, em princpio mantiveram o a respirao area mais seus sistemas traqueais apresentam muitas modificaes que os tornam adequados para as trocas gasosas na gua. A maioria dos sistemas traqueais apresentam variaes a partir de um padro bsico no qual existem caracteristicamente 12 pares de espirculos, 3 pares no trax e 9 no abdmen. Frequentemente existem menos espirculos e em alguns casos podem estar ausentes. Num padro tpico, as traqueias maiores conectam-se de tal modo que o sistema consiste de tubos que se interligam. Esse padro comumente
modificado pela adio de poro expandida das traquias, ou sacos areos, que so compressveis. O volume desses sacos pode ser alterado pelo movimento corpreo causando dessa maneira o bombeamento do ar para dentro e para fora do sistema traqueal. Isso importante, pois em insetos grandes e muitos ativos, apenas a difuso no suficiente para estabelecer as trocas gasosas. As traqueias contm anis espiralados internos e so um tanto incompreensveis; os grandes sacos areos so, portanto, necessrios para a ventilao. A abertura e fechamento dos espirculos, em sincronia
com os movimentos respiratrios, porm defasados entre si, permitem que uma corrente unidirecional de ar seja obtida ao longo das trocas traqueais maiores. A maioria dos espirculos no funcional e apenas os dois ltimos abremse pare o meio externo. Eles esto localizados de tal modo que o inseto, ao penetrar na superfcie da gua com a extremidade do abdmen, pode fazer contato com a atmosfera e realizar as trocas gasosas, por difuso ou, mais evidentemente por meio dos movimentos respiratrios. O sistema traqueal pode ser completamente fechado, sem qualquer apertura para a superfcie, embora seja preenchido com ar. Nesse caso, as trocas gasosas devem ocorrer inteiramente por difuso, atravs da cutcula. Muitos insetos aquticos pequenos conseguem realizar suas trocas gasosas dessa forma; a cutcula desses insetos relativamente fina e eles conseguem permanecer submersos sem a necessidade de contato com a atmosfera. No entanto, o sistema traqueal necessrio para o transporte de fases no interior do animal, j que a difuso atravs dos tecidos seria muito lenta.
Difuso e ventilao
A difuso isolada suficiente para as trocas gasosas no sistema traqueal de muitos insetos pequenos e de insetos grande relativamente inativo.
Os espirculos
As aberturas do sistema traqueal, para o exterior, os espirculos so estruturas altamente complexas que podem ser abertas ou fechadas para permitir uma quantidade varivel de trocas gasosas. O controle acurado dos espirculos ajuda a impedir a perda de gua. Os espirculos se abrem, mais frequente e amplamente, a temperaturas altas, e quando h um aumento de atividade, de acordo com a maior demanda de oxignio. Todos os espirculos no se abrem necessariamente ao mesmo tempo; eles esto sob o controle do sistema nervosos central e a abertura e o fechamento defasados permitem o controle do fluxo de ar pelo sistema traqueal. A ventilao do sistema traqueal, e especialmente a funo dos espirculos, influenciada pelo dixido de carbono e pela falta de exigindo. o dixido de carbono parece ser o principal estimulo para a abertura dos espirculos. se uma corrente diminuta de dixido de carbono for direcionada para um espirculo, apenas este se abrir; isso demonstra que um nico espirculo pode responder de forma independente a concentrao de dixido de carbono muito pequena; na barata, por exemplo, 1% de dixido de carbono no ar tem um efeito perceptvel; 2% mantm os espirculo abertos; e 3% fazem com que eles se mantenham amplamente abertos. O efeito da abertura dos espirculos na perda de gua considervel. Para um tenebrionideo, que passa toda vida num ambiente seco da farinha, o controle. Espirculos so muito importantes. Se os espirculos forem forados a permanecer abertos, pela adio de dixido de carbono ao ar, a perda de gua imediatamente aumentar diversas vezes. O dixido de carbono no o nico agente que controla os espirculos; Eles tambm podem ser forados a se abrir na presena de nitrognio puro (ou seja, a deficincia do oxignio tambm um estimulo). Isso poderia ser interpretado como um efeito do dixido de carbono, pois, quando h falta de oxignio, frequentemente existe acmulo de metablitos acido(p.ex.,acido dixido de carbono. O efeito
lctico ) e os cidos aumenta ateno de
atxico, portanto, poderia ser um efeito ser um efeito indireto de dixido de
carbono. Contudo, mesmo uma deficincia moderada de oxignio tenha um efeito direto.
Ventilao traqueal
As traqueias so relativamente rgidas e no se colapsam facilmente, porm, as dobras em espiral da parede permite um encurtamento do tipo acordeo. Isso pode servir para reduzir o volume o suficiente para contribuir para a ventilao ativa do sistema. As traqueias ovais ou achatadas na seco transversal so compressveis. Contudo, a ventilao efetiva depende de dilataes de paredes finas, ou sacos areos, que esto conectados com as traqueias maiores, pois quando esses sacos so comprimidos, um grande volume de ar pode ser expelido. A expirao normalmente a fase ativa e a expirao, a passiva. Em muitos insetos, os principais movimentos respiratrios so realizados pelo abdmen. Durante o vo, variaes sincrnicas de presso e movimentos podem aumentar a ventilao; mesmo a cabea pode ser ventilada ativamente devido a transmiao da presso sangunea. O volume ventilatrio pode ser muito grande. Um tero ou a metade da capacidade total do sistema pode ser expelido numa nica expirao, resultando em uma renovao de cerca da metade do volume do sistema respiratrio. Isso muito maior que num mamifero em repouso no qual a renovao a partir de uma nica respirao de aproximadamente um dcimo do ar contido no sistema. Em exerccio, a renovao mxima num mamfero prxima de dois teros do volume total do sistema respiratrio. Os movimentos so sincronizados com a abertura e o fechamento dos expiraculos e so tambm controlados pela falta de oxignio ou excesso de dixido de carbono. O excesso de oxignio pode causa completa parada dos sistemas respiratrios. Como concentrao alta de oxignio no deve influenciar a produo de dixido de carbono, provvel que o sistema nervoso seja diretamente afetado. Os movimentos respiratrios so controlados por centros no cordo nervoso ventral segmentado e segmentos isolados podem realizar movimentos respiratrios. A coordenao entre os segmentos realizada por centros superiores localizados no protrax. A cabea aparentemente no est envolvida. Os insetos, em geral, tm menos
centros superiores na cabea e a decapitao tem pouco efeito na respirao assim como em outras funes.
Respirao nos insetos aquticos
Os insetos que vivem evoluram a partir de formas terrestres e mantiveram muitas caractersticas de seus ancestrais. Quase todos so de guas doces; muito poucos vivem em gua salobada, e praticamente nenhum inseto verdadeiramente marinho. Alguns insetos mantiveram a respirao area atravs do espiraculos. Esses so em geral, reduzidos em nmero e esto localizados na extremidade posterior do corpo, que esta em contato com a atmosfera. As aberturas dos espirculos so hidrofbicas e frequentemente rodeado de pels hidrofbicos. A superfcie da gua , portanto, facilmente transpostas e o contato com o ar estabelecido.
Reservas externas de oxignio
Muitos insetos transportam ar do lado do corpo e os espirculos se abrem para o interior dessa massa de ar. O ar fica retido por meio de superfcies impermeveis, frequentemente auxiliadas por plos hidrofbicos.
Plastro
Outro tipo de dispositivo respiratrio conhecido como plastro. Um inseto que respira com um plastro tem parte da superfcie corprea densamente coberta com pels hidrofbicos que formam uma superfcie impermevel, o plastro, onde o ar mantido permanente. Esse dispositivo permite que o inseto continue submerso sem limite de tempo. Os insetos que respiram com um plastro apresentam grande vantagem sobre aqueles que carregam uma nica bolha como uma brnquia de difuso, pois a gua no penetra entre os pels hidrofbicos. Como resultado, o plastro serve como uma brnquia no compreensvel para o interior da qual o oxignio da gua se difunde. Pelo fato de o volume
permanecer constante, a difuso lquida de nitrognio nessa brnquia dever ser zero e a presso total dos gases no interior da massa de ar negativa em relao as tenses dos gases na gua circundante. Isso possvel porque os pels hidrofbicos do suporte ao filme superficial da gua, para que esta seja forada para dentro dos espaos do ar entre os pels, necessria de uma presso de 3,5 a 5 atm. Como resultado, os insetos que respira com auxilio de plastro seguem de maneira fcil permanecer indefinidamente em profundidades que exedam muitos metros.
Reserva interna de oxignio
Os insetos hempteros, buenoa e anisops, da famlia do inseto aqutico notonecta, so notrios por permanecerem suspensos no meio da gua, durante vrios minutos, quase sem fazer esforos. Isto bastante incomum entre os insetos, mas permitem que os animais predadores explorem a zona central de guas pobres em oxignio. Em pequenas lagoas temporrias, onde no existem peixes, essa zona relativamente livre de predadores, o fundo perigoso por causas das vorazes larvas de liblula e besouros; outros predadores caam na superfcie, mas na zona central h segurana, assim como pouca competio por alimento. Buenoa e anisops possuem grandes clulas abdominais repletas de hemoglobina. Elas servem para armazenar oxignio, que consumido durante o mergulho, os insetos carregam consigo apenas uma pequena massa de ar, mas esse suprimento externo esgotado lentamente enquanto oxignio proveniente da hemoglobina vai sendo consumido. Quando o
oxignio da hemoglobina esgotado, o oxignio externo rapidamente consumido e os insetos perdem a flutuabilidade ficando mais pesado que a gua. Ento, rapidamente, nadam para a superfcie para enxergar novamente as servas de oxignio. A reserva interna de oxigeno de longa durao confere um benefcio a esses animais. Aps terem reservado suas reservas de oxignio na superfcie, eles nadam rapidamente para permanecer em suspenso na zona
central, onde exibem flutuabilidade quase neutras durante um longo perodo, enquanto as reservas de oxignios vo sendo utilizadas.
Respirao descontinua ou cclica
Muitos insetos apresentam um fenmeno peculiar, caracterizado pela liberao peridica ou cclica de grandes quantidades de dixido de carbono. O oxignio captado a uma taxa mais ou menos constante, mas o dixido de carbono liberado durante breves perodos ou em surtos. Os surtos de dixidos de carbono podem duram poucos minutos e se alterar regularmente com longos perodos de liberao mnima do gs, que podem durar vrias hora, e at mesmo dias. A respirao cclica tem sido observada numa variedade de grupos de insetos, incluindo as baratas, gafanhoto, besouro, larvas pupas, assim como borboletas e mariposas adultas em diapausa.
O movimento dos espirculos
Uma observao direta do espirculos com o microscpio indica que eles fiam amplamente abertos duram o fluxo de dixido de carbono. Logo depois, a vlvula de cada espircula vibra durante um curto perodo e depois se fecha completamente, permanecendo assim por mais de uma hora. Em seguida a esse perodo, a vlvula comea a pulsar ou vibrar lentamente aumentando ligeiramente a abertura espiracular. A vibrao continua, frequentemente, durante vrias horas, at o prximo perodo de ampla abertura e surto de dixido de carbono.
Sangue
Foi visto que o transporte de oxignio e de dixido de carbono por difuso suficiente, exceto para animais muito pequenos. Quase todos os grandes animais, a menos que tenham uma demanda de oxignio bastante pequena,
possuem um sistema de distribuio voltado para o movimento de um lquido, o sangue. A uma tendncia apesar de transporte de gs como a principal funo do sangue, mais ele tem muitas outras funes que podem no ser imediatamente aparente. Os insetos, p. ex., no utilizam o sangue para o transporte gases; utilizam tubos cheios de ar. Esses animais possuem, no entanto, sangue que bombeado por todo o corpo, pois muitas outras substncias precisam ser transferidas mais rapidamente que o transporte proporcionado apenas pela difuso. Dessa forma, o sangue serve para transportar nutrientes que so absorvidos, no trato digestivo, e para transferir produtos de excreo para o rgo excretor.
Um variedade de produtos metablicos intermedirios, incluindo hormnios e outros compostos importantes, precisam de um sistema entre o local de produo e o de uso. Uma importante funo do sangue, frequentemente esquecida, a
transmisso de forma hidrulica, que utilizada em muitos processos tal como na utrafiltrasao renal, locomoo da minhoca, rompimento da carapaa na muda dos crustceos, ereo peniana, etc. Nos grandes animais, com uma elevada taxa metablica, o sangue essencial para o transporte de calor, caso contrario seus rgos internos rapidamente ficariam aquecidos. A capacidade de uma coagulao uma caracterstica inerente do sangue, necessria para reduzir a perda deste importante fluido quando ocorre dano no sistema vascular. A maioria das funes de transporte, assim como a transmisso da fora e de calor , pode ser executada por praticamente por qualquer meio aquoso. As excees so o transporte de gases e a coagulao, que esto associadas a propriedades bioqumicas do sangue altamente complexa.
Transporte de oxignio no sangue
Pigmentos respiratrios
Em muitos invertebrados o oxignio transportado pelo sangue ou hemolinfa em soluo fsica simples. Isso ajuda a transporta o oxignio da superfcie para vrias partes do organismo, pois a difuso sozinha muito lenta para qualquer organismo, exceto os menores. Entretanto, a quantidade de oxignio que pode ser transportada em soluo simples pequena, e muitos animais altamente organizados (vertebrados quase sem exceo) possuem sangue que tem a capacidade de se ligar recersivelmente a grandes quantidades de oxignio, aumentando bastante, dessa maneira a quantidade desse oxignio transportada. As substncias conhecidaa como transportadoras de oxignio no sangue so protenas que contm um metal (comumente ferro ou cobre). Normalmente elas so coloridas e, por isso so frequentemente chamadas de pigmentos de respiratrios. A hemoglobina o pigmento respiratrio mais conhecido e difundido. Elas consistem de uma molcula de protena associada a uma estrutura anelar de quatro membros, chamada porfirina. O ferro contido em cada molcula de hemoglobina ligado a porfirina com uma tomo de ferro de valente (fe2+), ligado a cada unidade de porfirina. A clrocruorina tem uma estrutura bastante similar, um grupo porfirnico contendo ferro associado a uma molcula de protena. A hemeritrina,
entretanto bastante diferem; apesar de conter ferro, ela no tem a estrutura porfinica. a hemocionina um pigmento que contem cobre e ocorre apenas nos moluscos e artrpodes, no entanto ao lado da hemoglobina, o pigmento respiratrio mais amplamente distribudo.
Corpsculos sanguneos
Em alguns animais, o pigmento respiratrio est dissolvido no fluido sanguneo; em outros,tais como os vertebrados, est contido no interior de celular e no contm pigmento respiratrio dissolvido. As grandes molculas de pigmentos respiratrios dissolvido no plasma, so de fato, agregados de subunidades menores, cada uma delas capaz de se unir ao oxignio. Esta estrutura com unidades mltiplas aumenta a quantidade total do pigmento sem aumentar o nmero de molculas de protenas dissolvido no ponto do plasma sanguneo. Um grande aumento no
nmeros de partculas devolvida aumentaria a presso coloidosmatica plasmtica que, por sua vs influiria em muitos outros processos fisiolgicos tais como a passagem de lquido atravs das paredes do capilares e a ultrafitraao (processo inicial de formao de urina nos rins). Existem diversas vantagens em si encerrar a hemoglobina nas clulas, a mais importante provavelmente que o ambiente no interior dos glbulos vermelhos pode ser diferente daquele do plasma sanguneo. A reao entre o oxignio e a hemoglobina muito influenciada por ons inorgnicos e tambm por determinados composto orgnicos, principalmente os fosfatos orgnicos, e a hemoglobina localizada no interior das clulas pode estar em ambiente isolado e bem regulado, diferente do plasma.
Tamanho do formato do glbulo vermelho
Os glbulos dos mamferos so discos esfricos, ligeiramente bicncavos. H uma exceo: todos os membros da famlia dos camelos tm lbulos vermelhos ovais. Os glbulos vermelhos dos mamferos no tm ncleos, embora tenham um curto perodo de vida - cerca de 100 dias - e realizem funes metablicas complexas. Os glbulos vermelhos de todos os outros vertebrados: aves, rpteis, anfbios e peixes possuem ncleos e so quase que,
universalmente, ovais; ele frequentemente so muito maiores que os mamferos; os glbulos vermelhos das salamandras, por exemplo, so 100 ou 200 vezes maiores em volume do que os glbulos vermelhos dos mamferos. A importncia, se houver, de glbulo vermelho nucleado versus um no-nucleado no compreendida. Nem claro se o tamanho dos glbulos vermelho tem importncia funcional. Os nicos vertebrados que no possuem hemoglobina e glbulos vermelhos so as larvas enguias (larvas de leptocephatalus) e alguns peixes antrticos da famlia chaenichthyidae.
O efeito da temperatura
Se a curva da dissociao for determinada para uma amostra de sangue de mamifero a temperatura ambiente e depois a temperatura corprea do animal, os resultados sero completamente diferentes. Isso ocorre porque temperaturas elevadas tornam mais fraca a ligao entre a hemoglobina e o oxignio causando maior dissociao como consequncia a uma maior temperatura a hemoglobina libera o oxignio mais rapidamente e a curva de dissociao deslocada para a direita. Essa ocorrncia de importncia fisiolgica, especialmente em animais de sangue frio, porque uma maior temperatura normalmente acompanhada por um alto aumento da taxa metablica (o demanda de oxignio), por isso vantajoso que hemoglobina libere oxignio mais facilmente a uma temperatura mais alta mesmo nos animais de sangue quente isso pode ter alguma importncia na febre ou durante o exerccio, por exemplo, quando a uma elevao na taxa de consumo de oxignio.
O efeito da altitude
A baixa presso atmosfrica a uma altitude elevada significa que a presso parcial do oxignio mais baixa do que no nvel do mar. Os animais vivem em ambiente de relativa deficincia de oxignio apresentam adaptaes correspondentes das curvas de dissociao de oxignio no sangue.
Animais mergulhadores
Seria esperado que mamferos mergulhadores, como as focas que, normalmente, ficam expostas a uma deficincia relativa de oxignio quando mergulham, pudessem obter vantagens se o sangue tivesse uma alta afinidade pelo oxignio. Todavia, esse no o caso e, com um pouco raciocnio, ficara claro que esses animais no obteriam qualquer vantagem especial dessa maneira. Quando uma foca respira na superfcie ela inspira o ar atmosfrico. A tomada de oxignio pelo sangue no pulmo, portanto,
ocorre sob circunstncias semelhantes quelas de outros mamferos e o suprimento do sangue no pulmo no requer uma afinidade no oxignio especialmente alta. Durante o mergulho, a liberao de oxignio nos tecidos (principalmente no sistema nervoso central e no msculo cardaco) no seria auxiliada por uma alta afinidade do sangue pelo oxignio; ao contrrio, talvez, uma alta afinidade da hemoglobina pelo oxignio tenderia a impedir a liberao desse gs para os principais rgos vitais. Desse modo, as curvas de dissociao de oxignio de animais mergulhadores geralmente diferem um pouco daquelas de animai similares que no mergulham.
Difuso facilitada
fcil observar que a hemoglobina carregada no sangue, melhora muito o transporte de oxignio, no entanto a hemoglobina no ocorre somente no sangue, mas tambm em vrios tecido, e nos animais superiores esta principalmente presente nos msculos. Uma maior capacidade de oxignio dos tecidos pode amenizar pequenas flutuaes quando ocorrem rpidas elevaes na demanda de oxignio. A difuso facilitada esta associada a habilidade da hemoglobina em ligar-se reversivelmente ao oxignio a hemoglobina transformada em meteglobina (obtida pela oxidao do ferro de divalente em trivalente) no pode mais se ligar a oxignio uma soluo de metemoglobina no causa qualquer facilitao da difuso do oxignio. Contudo, a mioglobina e vrias hemoglobinas, assim como de invertebrados apresentam difuso facilitada. um aspecto importante da difuso facilitada que uma maior difuso ocorre somente quando a presso de oxignio na extremidade descendente da trajetria de difuso encontra-se prxima de zero.
Explicaes fisiolgicas
Nos vertebrados a hemoglobina esta presente em alta concentrao, principalmente nos glbulos vermelhos e secundariamente nos msculos vermelhos. discutvel se difuso facilitada tenha muita importncia nos eritrxilos, pois a facilitao requer uma presso de oxignio prxima de zero
na extremidade receptoras do processo da difuso os glbulos vermelhos, que devido o seu tamanho reduzido j tenha uma distancia de difuso muito curta, so raramente, ou nunca, exposto a presso de oxignio prximo de zero, exceto nos capilares sanguneos de msculos com atividades intensas. Para a mioglobina a situao diferente: a certa distncia de um capilar sanguneo, a tenso de oxignio do msculo em atividade pode facilmente se aproximar de zero. Isto estabelece uma situao ideal para a difuso
facilitada na qual a tenso de oxignio no capilar encontra-se entre a tenso arterial no tecido (extremidade receptora do processo de difuso), prxima a zero. Uma difuso facilitada aumenta em muitas vezes, tal como foi
observada em experincias em laboratrio seria realmente importante para melhorar o suprimento de oxignio junto ao msculo. Experimentos de msculos de pombo confirmam que a mioglobina pode transportar uma frao significativa consumido pelo msculo.
Implicaes evolutivas
curioso que uma molcula to complexa como a hemoglobina ocorra to amplamente, ainda que de forma to dispersa e espordica. Alm de estar quase que universalmente presente nos vertebrados, a hemoglobina ocorre aqui e ali num nmero grande de filos animais, incluindo os protozorios, alm de vegetais e bactrias. Frequentemente, encontrada em uma ou poucas espcies dentro de um grande filo; s vezes num parasita, e no em um parente seu de vida livre, e assim por diante. Contudo, uma vez estabelecida a vantagem da hemoglobina no processo de difuso, e facilitada a sua distribuio peculiar, mais fcil de ser entendida. A estrutura bsica do componente heme da molcula de hemoglobina um ncleo de porfirina que est universalmente presente em vegetais e animais como parte das enzimas celulares que pertencem ao sistema citocrmico. Alteraes bioqumicas, relativamente pequenas, podem transformar essa matria prima num pigmento do tipo hemoglobina que se liga reversivelmente ao oxignio. Para que tais mutaes permaneam, adequado que favorea sua manuteno. necessrio algum benefcio
A vantagem mais obvia imediata da hemoglobina a melhoria da liberao de oxignio devido a difuso facilitada. Uma vez estabelecida a sntese de hemoglobina, a posterior melhoria no transporte de oxignio pode ser conseguida por combinao de um transporte convertvel (ou seja, sangue e sistema circulatrio). O estabelecimento de um sistema circulario completo uma alterao complexa que requereria no somente modificaes bioqumicas
relativamente menores na porfirina, mais tambm alteraes fisiolgicas e morfolgicas complexas e em todo o organismo. Tais mudanas drgicas so possveis, claro, mas dificilmente podem ser conseguidas por meio de um nico evento ou por meio de poucos passos intermedirios. Em
contraste, uma ocorrncia inicial de hemoglobina em qualquer tecido fornece imediatamente ao organismo a melhoria da transferncia de oxignio decorrente da difusa facilitada resultando numa vantagem seletiva para sua manuteno. Um passo posterior seria o de permitir que esta hemoglobina entra-se no j existente sistema sanguneo, e aumentado, assim a liberao de oxignio pelo processo de conveco.
Transporte de dixido de carbono no sangue
Quando o sangue passa pelos tecidos, e libera oxignio, ele capta simultaneamente dixido de carbono; quando passa pelos pulmes, acontece o inverso. Examinamos os eventos no pulmo, quando ocorreu a liberao de dixido de carbono. O sangue venoso do mamifero, quando alcanar os pulmes , contm, aproximadamente, 550 cm de Co por litro de sangue e tem uma tenso de dixido de carbono de aproximadamente 46 mmhg (6,1 kpa).
Quando o sangue deixa os pulmes, contm cerca de 500 cm de Co por litro de sangue A uma tenso de aproximadamente 40 mmhg, assim, o sangue libera somente um pequena parcela da quantidade total de dixido de carbono presente, sendo a diferena arteriovenosa aproximadamente 50 ml de Co por litro de sangue. Para entender o transporte de dixido de carbono, preciso conhecer melhor esse gs e seu comportamento em
soluo a quantidade total de dixido de carbono transportada no sangue excede muito a quantidade que estaria dissolvida na gua a mesma tenso deste gs no sangue. Isso acontece porque a maior parte do dixido de carbono encontra-se combinado quimicamente e no sobre forma gasosa, devolvido livremente. Os dois compostos mais importantes so (1) os ons bicabornato e (2) uma combinao de dixido de carbono com hemoglobina.
Parte 3: circulao
O principal objetivo da movimentao de um lquido no corpo o de propiciar rpido transporte de grande volume a distncia onde a difuso inadequada ou muito lenta. Portanto, a circulao importante virtualmente todos os animais com mais de pouco milmetros de tamanho uma necessidade para os grandes animais com altas taxas metablicas. Alm de transportes de gases, a circulao sangunea serve: (1) para
transportar outros solutos, (2) para transportar o calor e (3) para transmitir fora. As funes que dependem da transmisso de fora esto relacionadas, na maioria das vezes, com o movimento do animal todo, o movimento dos rgos e a manuteno da preo pra a ultrafiltrao nos vasos sanguneos renais; estas funes sero discutidas posteriormente. Para entender como o sangue se movimenta no sistema circulatrio necessrio conhecer alguns princpios fsicos bsicos do movimento de fluidos em tubos. Alm disso, o sangue, visto como um fluido tem algumas propriedades em comum que interferem em suas carecteriscas de fluxos. Um sistema adequado para o transporte convectivo de oxignio invariavelmente suficiente para o transporte de dixido de carbono e outros solutos
Princpios gerais; Bombas
Um sistema circulatrio adequado depende de uma ou mais bombas e de canais ou de condutos nos quais o sangue possa fluir. A bomba, ou o corao, baseia-se na capacidade do msculo de contrair-se o encurtar-se. A existncia de um msculo, no entorno de um tubo ou cmara, permite a reduo de volume. Dois tipos diferentes de bombas podem ser projetadas dessa forma: bombas peristlticas ou bombas com cmara e vlvulas. Uma bomba com cmara pode paredes contrateis ou a reduo volumtrica pode ser conseguida atravs da presso externa proveniente de outras partes do corpo. Os coraes peristlticos so encontrados principalmente em invertebrados; o corao dos vertebrados , quase sem exceo, uma bomba com cmara com paredes contrteis. Uma tpica, na qual a fora provida pela presso externa, encontrada nas grandes pernas dos homens. As paredes dessas veias so relativamente finas; elas apresentam vlvulas que impedem a reverso do fluxo sanguneo quando os msculos das pernas se contraem, as veias so comprimidas as vlvulas a seguram que o sangue impelido na direo do corao. Essa ao de bombeamento auxilia muito o movimento sanguneo para longe das veias das pernas contra a fora da gravidade, que tende a fazer com que o sangue seja acumulado nestas. Como o msculo pode prover fora apenas pelo encurtamento, o corao no pode movimentar o sangue, a menos que sejam feitos ajustes especiais. o corao do elasmobrnquio um exemplo. Ele rodeado por uma cpsula firme e quando e cmara se contrai, a outra cmara preenchida por suco, enquanto as vlvulas impedem o refluxo do lado arterial.
Circulao dos vertebrados
Compartimentos de gua e volume sanguneo
Aproximadamente dois teros do corpo dos vertebrados consistem de gua. O valor exato varia muito, particularmente em consequncia de variaes na quantidade de gordura. A maior parte da gua est localizada no interior das clulas (a gua intracelular); uma frao menor est do lado externo das membranas celulares (a gua extracelular). A gua extracelular esta
localizadas nos espaos teciduais (lquido intersticial) e parcialmente no sangue (gua do plasmas). Portanto, pode-se considera a gua corprea distribuda entre trs compartimentos: intracelular, intersticial e plasma.
Padres circulatrios
Cada classe de vertebrados tem um tipo muito uniforme de circulao, mais as diferenas entres as classes so substanciais. medida de que a vida dos vertebrados passa de aqutica para terrestre, a circulao torna-se mais complexa. Existem duas consequncias importantes decorrentes da disposio da circulao dos mamferos. Primeiro, o fluxo sanguneo atravs dos pulmes deve ser igual ao fluxo sanguneo atravs do restante do corpo (exceto por pequenas variaes transitrias que podem ser provocadas por ligeiras alteraes no volume cardaco durante um ou alguns partimentos). Segundo, como as duas metades do corao contraem-se simultaneamente, conforme o sangue ejetado do corao, todo o volume ejetado deve ser acomodado por meio de variaes no dos vasas sanguneos elsticos.
Peixes
Os peixes, tanto telesteos como elasmobrnquios, apresentam um corao que consiste de duas cmaras em srie, um trio um ventrculo. No lado venoso, o corao precedido, na altura da veia, por uma cmara maior ou seio, o seio venoso, que ajuda a assegurar o fluxo sanguneo para o corao. No lado arterial, o corao do telesteo imediatamente seguido por uma parte muscular espessada da horta ventral o bulbo arterial. O corao alasmobrmquio tem uma parte espessada localizada de forma semelhante, o cone arterial, desenvolvido a partir do msculo cardaco. Ele fibroso e equipado com vlvulas que evitam o fluxo sanguneo reverso para o interior do ventrculo. Isso particularmente importante porque o corao do elasmobrnquio, localizado numa cmara rgida, pode produzir presses negativas. Uma presso negativa no corao facilita o enchimento por suco doa trio a partir dos grandes seios venosos.
Rpteis
Em um rptil no-crocodiliano, os trios so completamente separados, mais o ventrculo parcialmente dividido. Mesmo assim, as correntes de sangue oxigenados e no-oxigenados so mantidas, de modo que ocorre muito pouco a mistura de sangue, existindo de fato uma circulao dupla bem desenvolvida. Dessa maneira, a diviso incompleta do ventrculo dos anfbios e rpteis no-crocodilianos no pode ser interpretada simplesmente com base na aparncia anatmica; as correntes sanguneas permanecem muito mais separadas do que fazem supor as consideraes anatmicas.
Circulao dos invertebrados
Muitos
dos
invertebrados
possuem
sistemas
respiratrios
bem
desenvolvidos; alguns exemplos so os aneldeos, equinodermos, artrpodes e moluscos. A maioria deles tm sistema aberto. Os cefalpodes (polvo e lula) tm circulaes fechadas, apesar de as outras classes de moluscos terem sistemas abertos. bom observar que os insetos, que so animais altamente organizados e complexos, e que conseguem manter a atividade metablica excepcionalmente altas, possuem sistemas circulatrios abertos.
Moluscos
Com exceo do polvo e da lula, os moluscos apresentam
sistemas
circulatrios abertos; o sangue de muitos moluscos contm hemocianina, e alguns poucos possuem hemoglobina. A capacidade de transporte de
oxignio do sangue tende a estar correlacionada com o tamanho do animal e, em particular, com suas atividades. Os moluscos, em geral, tm um corao bem desenvolvido e o batimento cardaco ajustado para satisfazer as demandas fisiolgicas de oxignio. Um crescente retorno venoso sanguneo para o corao causa a um aumento tanto na amplitude como na frequncia do batimento cardaco. O corao dos moluscos continua a bater se for isolado do sistema nervoso: ele
tem uma ritmicidade inerente com um marca passo, contudo, o corao tambm est sobre influncia de neurossecreoes que modificam seus batimentos. A acetilcolina inibe o corao e a serotonina agem como um substncia excitatria.
Insetos
Um inseto normalmente tem um vaso sanguneo principal que se estende ao longo de lado dorsal. A parte posterior desse vaso funciona como um corao e provido de uma srie de aberturas com vlvulas atravs das quais o sangue pode entrar. A parte anterior do vaso sanguneo, que poderia ser chamada de aorta. contrtil e pode apresentar ondas peristlticas que impulsionam o sangue na direo anterior. Esse caso sanguneo principal ramifica-se e prossegue em direo a cabea onde termina. As ramificaes da aorta suprem a maior parte do corpo, onde o sangue flui livremente entre os tecidos e lentamente percola de volta ao corao. Em muitos insetos, membranas longitudinais do alguma direo ao sangue que flui entre o tecidos. Nas antes e nos membros, o sangue pode entrar de um lado e sair do outro. Dessa forma, o sangue conduzido para certo trajetos, apesar de no fluir em vasos sanguneos isolados.
Aracndeos
O sistema respiratrio das aranhas e escorpies semelhante ao dos insetos, mas pode ter um papel maior na respirao em relao a estes. A hemociamina encontrada no sangue de alguns escorpies e tanto escorpies como aranhas apresentam rgos respiratrios definidos que so perfundidos por sangue. Isso no deva ser surpresa, pois os aracndeos no possuem sistema traqueal, caracterstico dos insetos. O corao dos aracndeos est localizado dorsalmente do abdmen. ele se enche atravs das aberturas com vlvulas e esvazia-se no interior das artrias. Nas aranhas, artrias distintas levam sangue as pernas, onde a uma presso sangunea relativamente e importante na locomoo. Nas pernas
das aranhas no h msculos extensores e o sangue utilizado como fluido hidrulico para a extenso das pernas.
4 Alimento e combustvel
Os animais necessitam de alimentos (1) para prover a energia necessria para permanecerem vivos e manterem os processos fsicos, para a contrao muscular e muitos outros processos, (2) como matria-prima para formar e manterem o mecanismo celular e metablico e (3) para crescimento e reproduo. As plantas utilizam a energia solar e o dixido de carbono da atmosfera para sintetizarem acares e, indiretamente, todos os composto complicados que consistem numa planta. Todos os animais utilizam compostos qumicos para fornecerem energia e matria estruturais. Eles devem obter diretamente dos vegetais ou de outras substncias. H excees para esta dependncia universal da luz solar. Em alguns locais no fundo do mar, onde no h penetrao de luz, existem ricas comunidade animais que utilizam das caractersticas qumicas peculiares da gua aquecida por processos geomtricos. A trs aspectos principais no temo alimentos: captura de alimentos, digesto e nutrio. Primeiro, ser tratado a captura de alimentos, que se refere aquisio e ingesto do alimento. Quase todos os alimentos de origem vegetal ou animal, consistem de composto altamente complexo que no podem ser usados sem que sejam antes degradados em composto mais simples. Estes processos so referidos como digesto. Uma variedade de compostos orgnicos podem prover energia, mas, alm disso, os animais tm necessidades especficas por compostos que no conseguem sintetizar, tais como aminocidos e vitaminas. A necessidade de alimento para prover energia, e a necessidade de comportares especficos permanecem ao consulto nutrio.
Captura de alimentos
O alimento obtido por meio de uma diversidade de mtodos mecnico, que determinam a natureza do alimente que um dado animal pode obter e utilizar.
Pequenas partculas
Alga e bactrias microscpicas podem ser capturadas diretamente por uma clula (p.ex. nos vcuos digestivos de uma ameba). Uma variedade de organismos microscpicos com o auxilio de clios. Alguns animais notadamente tunicados e alguns gastrpodes, utilizam um filme mucoso que captura diminutas partculas suspensas; este filme ingerido e os organismos, nele contidos, digeridos.
Matria orgnica dissolvida
A gua do mar contm uma grande variedade
de compostos orgnicos
devolvidos, mas em concentrao extremamente baixas. No comeo do sculo XX, o bilogo Putter sugeriu que a matria orgnica dissolvida no mar poderia ser tomada e utilizada diretamente pelos animais aquticos. Putter sugeriu que tal matria orgnica dissolvida era mais importante que o plncton, pois os organismos do plncton ficam to espalhados na gua que, frequentemente, a captura destes no possvel.
Alimento obtido a partir do interior da terra
Em 1977, os bilogos que estudam o fundo do mar descobriram densas populaes de animais peculiares, inteiramente novos cincia, em diversos lugares do fundo do oceano. O fato mais surpreendente que estes animais so completamente independentes das usuais cadeias alimentares baseadas de fotossnteses. Eles existem em locais de atividade vulcnica submarina onde as placas rgidas da crosta terrestre se separam. O solo do mar nessas reas marcado por fendas onde uma fina camada rochosa aquecida a altas temperaturas pelo magma quente subjacente. Quando a gua
enfeltrasse na rocha, ela aquecida e emerge novamente como fontes termais, ou aberturas hidrotmicas.
Essas aberturas so circundadas por ricas comunidades de mexilhes, muitos bivalves, lapas, caranguejos, pogonfores e celenterados. Alguns dos animais atingem tamanhos enormes; os moluscos podem ter 30 cm e os pogonforos mais de 2 m de comprimento. A maioria desses animais rara, anteriormente desconhecidos da cincia.
Digesto
O alimento dos animais consiste de matria orgnica e grande parte dele pertence a trs grupos principais: protenas, lipdios, e carboidratos. Esse trs tipo de compostos orgnicos dominam completamente a composio de quase todos os vegetais e animais e a maioria tem molculas razoavelmente grandes. seja o alimento utilizado para combustvel ou para desenvolvimento e manuteno, as grandes molculas de alimentos so inicialmente degradas em unidades, mais simples: essas so ento absorvidos e podem ser incorporados ou metabolizadas para fornecer energia. A principal funo da digesto decompor as molculas grandes e complexas, presentes nos alimentos, de modo a torn-las absorvidas e disponveis para utilizao no corpo. Essa degradao executada no trato digestivo com o auxlio de enzimas.
Madeira como alimento: digesto simbitica.
O material estrutural mais importante dos vegetais a celulose, um polmero constitudo por glicose, que extremamente insolvel e refratrio ao ataque qumico. As enzimas que digerem as celuloses, as celulases, ausente nas secrees digestiva dos vertebrados e, no entanto, muitos deles dependem da celulose como fonte energtica. A presena das celulases verdadeiras foi verificadas no trato intestinal de muitos invertebrados que se alimentam de madeiras e produtos vegetais similares, mais em muitos casos a digesto de celulose realizada por microorganismos simbinticos que vivem no trato digestvo hospedeiros.
Digesto de celulose nos invertebrados
A questo se a celulase produzida por um animal ou por um microorganismo simbintico tem de ser, frequentemente, confirmada com segurana.
Nutrio
As propriedades mecnicas do alimento devem corresponder ao aparelho de tomada alimentar e a natureza qumica do alimento deve ser adequada aos mecanismos digestivos disponveis. Uma vez satisfeitas estas
necessidades a prxima pergunta : qual a adequao do alimento relao as necessidades do organismos? A nutrio trata desta questo. O problema pode, convenientemente, ser dividido em duas partes: a quantidade e a qualidade do alimento. O organismo precisa de (1) energia para as atividades externa e manuteno interna e (2) suprimento e substancias especficas para a conservao e crescimento. Entre estas substancias especficas esto includas os aminocidos, vitaminas e outro nutrientes essenciais alm de vrios elementos, alguns deles em quantidade to diminutas que tem sido estabelecer se so realmente necessrios.
Regulao de a ingesto alimentar
Em muitos animais, a ingesto alimentar extremamente bem ajustada em relao ao gasto energtico. Se as necessidades energticas forem aumentadas por caudas das atividades fsicas, a ingesto alimentar ser regula de acordo. Os mecanismo regulatrios para a ingesto alimentar dos mamferos esto localizadas na rea hipotalmica do celebro, mas influncia perifricas especficas tem um papel principal no nvel momentneo de fome e ingesto alimentar. Uma das influncias importante o nvel do sanguneo do acar. Foi demonstrado que pontadas de fome conhecido com as
contraes do estmago, e que elas ocorrem especialmente quando o nvel sanguneo de glicose diminui. Alm disso, o grau de enchimento do
estmago de evidente importncia na induo de uma sensao de saciedade e trmino da ingesto alimentar.
Defesa qumica
Um meio de se evitar ser comido torna-se no comestvel. Este princpio simples amplamente utilizado por plantas e animais. Os vegetais so os produtores primrios na cadeia alimentar e, como no conseguem fugir, esto sempre prontos para serem devorados. Tornar-se no comestvel com o auxilio de compostor txicos uma defesa bastante utilizada, e um nmero surpreendente de plantas so venenosas para os animais. Seus predadores, por sua vez, desenvolveram mecanismos para evitar ou neutralizar essas defesas, de modo que dificilmente qualquer planta esteja livre de inimigos.
Venenos de plantas
Vrios compostos vegetais afetam quase todos os sistemas fisiolgicos. Pode agir na pele, sistema digestivo, sangue, vrias partes do sistema nervoso, metabolismo, valor nutricional do alimento, sistemas hormonais e reprodutivos dos animais. A variedade de compostos e efeitos tornam difcil uma classificao. Contudo, podem ser divididos em grupos principais, em parte por causa das semelhanas qumicas e da similaridades quanto aos efeitos.
Uso animal de venenos prprios
Os animais tambm usam uma ampla variedade de venenos e toxinas que eles prprios produzam. Exemplos bem bom conhecidos so cobras, escorpies, aranhas, abelhas e vespas. Entre os tipos mais incomuns de defesa incluem-se o cianeto de hidrognio produzem pelos milpedes quando so perturbados e, mais dramtico ainda, o jato qumico ejetado a na temperatura de 100 C pelo besouro bombardeiro.
5 Metabolismo energtico
Os animais precisam de energia qumica para realizar suas vrias funes e o uso dessa so com frequncia denomino metabolismo enrgico. Os animais obtm energia principal por meio de oxidao dos alimentos. A quantidade de oxignio que eles consomem pode, portanto, ser usada como uma medida do metabolismo energtico. Contudo, nem sempre isso acontece. Alguns animais podem viver na ausncia do oxignio livre. Eles tambm utilizam energia qumica para suas necessidades energticas, embora as vias metablicas sejam diferentes. Tais processo metablicos so conhecidos como metabolismo aneorbico. Essa situao normal para animais que vivem nos ambientes pobres em oxignio ou toleram a falta prolongado ou permanente de oxignio. Eles obtm energia atravs de processos que no utilizam oxignio molecular. Os processos que requerem energia e as reaes no organismo vivo utilizam uma fonte comum de energia, o trifosfato de adenosina (ATP). Esse composto ubquo, por meio da hidrlise de uma ligao de fosfato rica em energia, a fonte de energia imediata para processos como a contrao muscular, o movimento ciliar, a luminescncia dos vaga-lumes, a descarga de peixe eltrico, os processo de transporte celular, todos os tipo de reaes de sntese e assim por diante. A molcula de ATP formada nas vrias etapas geradora de energia na oxidao dos alimentos e tambm em processos anaerbios gerados de energia. O ATP o intermedirio universal no fluxo de energia para processos que requerem energia nos organismos aerbios e anaerbios.
Armazenamento de energia: gordura e glicognio.
Para a maioria dos animais adultos, o consumo de alimentos e gastos de energia permanecem aproximadamente iguais. Se o gasto de energia exceder o consumo de alimentos, o excesso de energia ser coberto pelo
consumo de substancia corporais, primeiramente gordura. Se o consumo de alimentos utilizados o excede, ser armazenado como gordura, independente
da composio do alimento. Por exemplos, os porcos so engordados com alimentos que consistem principalmente de carboidratos gros.
Efeito da concentrao de oxignio
Em geral, supe-se que o consumo de oxignio (taxa metablica), dentro dos limites amplos, seja indepetemente da concentrao de oxignio. Por exemplo, se substitumos o ar comum por oxignio puro, um mamifero continuar consumir o oxignio da mesma taxa embora oxignio esta taxa presente numa concentrao cinco vezes maior. O inverso tambm verdadeiro. Se reduzirmos a concentrao de oxignio pela metade, o que pode ser feito reduzindo a presso atmosfrica total metade (o que equivale,, aproximadamente, a 6.000 m de altitude), a taxa de consumo de oxignio continuara, da mesma forma, quase inalterada. Essa independncia na concentrao de oxignio no universal. A situao pode ser bastante diferente. Em temperaturas altas, o consumo de oxignio mais alto que em temperatura baixas, mas em qualquer temperatura a tomada de oxignio relacionada de maneira linear com o oxignio disponvel.
Aclimao baixa de oxignio
Muitos peixes ativos que nadam rapidamente so muitos sensveis ao baixo teor de oxignio na gua. Isso verdadeiro para a maioria de salmondeospor exemplo, a truta pintada. Para esse peixe, o nvel letal de oxignio na gua depende da exposio anterior a nveis baixos, mas tolerveis de oxignio. Se o peixe for mantido em gua relativamente pobre em oxignio, ele ficar mais tolerante a baixos teores de oxignio. A melhor tolerncia provavelmente atribuvel a uma ampliao de a capacidade extrair oxignio da gua. Se houver um aumento real na capacidade do tecidos de tolerarem condies atxicas, um peixe aclimatada a baixos nveis de oxignio poder viver mais que um peixe mais aclimado, quando colocado em condies anaerbias. No h evidncias de tal diferena.
Metabolismo anaerbio
Os animais que toleram exposies prolongadas a oxignio zero, devem necessariamente obter energia metablica a reaes no oxidativas. Isso verdadeiro para muitos parasitas intestinais, para animais que vivem no lodo livre de oxignio de lagos e represas, para os bivalveis que ficam fechados durante longos perodos e para muitos outros. Os animais que podem sobreviver, indeterminadamente, sobre perodo sob condies anxidas so chamados de organismo anaerbios.
A camada mnima de oxignio
A maioria dos oceanos do mundo contm, em profundidades intermedirias, zonas onde o contedo de oxignio muito baixo. H uma grande rea desse tipo no nordeste do Pacfico tropical, que conhecida como a camada mnima de oxignio a partir da costa central do sul do Mxico; a camada se estende, aproximadamente, entre 100 e 900 m de profundidade e o contedo de oxignio devolvido geralmente menor que 0,1 cm (centmetro cbico) por litro de gua no mar, com relativamente pouca variao durante o ano. No entanto, uma populao surpreendente de animais vive na gua pobre de oxignio. Alguns dos animais particularmente os peixes fazem migraes dirias para a superfcie, acendendo no crepsculo e retornando as profundezas antes do amanhecer, em movimentos verticais que perfazem 300m ou mais. Paradoxalmente, os peixes presentes na regio de baixo teor de oxignio geralmente tm uma bexiga natatria cheia de gs que contem uma porcentagem alta de oxignio.
Embolia gasosa
Essa sndrome perigosa conhecida por vrias denominaes: doenas do mergulhador, mal dos caixes e aerbolea. Ela ocorre guando um mergulhado humano retorna para a superfcie, aps um perodo prolongado
em profundidade considervel, abaixo de 20 m mais ou menos. A sndrome agravasse quando maior a profundidade e o tempo de mergulho. causada por bolhas de ar nos tecido e nas correntes sanguneas, do mesmo modo que as bolhas formam em uma garrafa de refrigerante quando a tampa removida. Ambos os casos, as bolhas aparecem guando a presso reduzida em lquido que est saturada de gs em alta presso. A aerbolia tambm pode ocorrer durante a sesso rpida de aeronaves com cabines no pressurizadas: nesses casos, o perodo de aerbolia ocorre quando a presso reduzida para cerca de 0,5 ATM (cerca de 6 mil metros).
Toxidade do oxignio
A importncia da toxidade do oxignio no mergulho obvia. Se o mergulhador descer a 40m e o suprimento de ar comprimido, ele respirar a uma presso total de 5 atm. Como 1/5 do ar e oxignio, a presso principal de oxignio 1 atm (i.e., prximo limite txico). Os mergulhos com ar
comprimido em profundidades ainda maiores so, claro, cada vez mais prejudiciais, pois a presso parcial do oxignio precede a 1 atm. O nico modo prtico de o mergulhador trabalhar 40m ou mais evitar a toxidade do oxignio e diminuir a o contedo de oxignio do ar que respira. Um mergulhador que trabalha a 40m (5 atm) poderia ter um suprimento com gs que contenha O2 a 10% e N2 a 90%. Ento ele respiraria o oxignio a uma presso de 0,5 atm, que completamente seguro em uma profundidade maior, o contedo do oxignio da mistura gasosa deve ser ainda mais reduzido para diminuir o risco aeroenbolia durante a descompresso, o nitrognio deve ser parcial ou completamente substitudo por gs hlio.
Suprimento de oxignio durante o mergulho
Para uma pessoa que mergulha sem equipamento, a necessidade mais imediata e obter o oxignio. Em geral, ela enche os pulmes para aumentar a quantidade de ar que carrega com sigo adiando o momento para voltar para a superfcie para respirar novamente. Existem, contudo, muitas
possibilidades tericas para aumentar o tempo de mergulho. Algumas delas
so, do pondo de vista fisiolgico, no factveis ou no se sabe que sejam utilizadas pelos animais. A maiorias dos animais que mergulha, adotou uma combinao de vrios mtodos possveis, em geral resultando em desempenhou de mergulho que so surpreendentes quando comparados aos padres humanos. Respirao cutnea e retal
J foi visto quo importante a respirao cutnea em rs como a salamandra que, sem pulmo, obtm todo oxignio atravs da pele. No h razo para acreditar que um pssaro ou um mamfero mergulhador utilize uma via similar para cobrir qualquer parte substancial da sua demanda por oxignio. Por um lado, esses animais so grandes e as suas reas
superficiais relativas, e so, portanto, pequenas. Por outro lado, eles so animais de sangue quente e suas demandas de oxignios so altas. Finalmente. A natureza de suas peles muito diferente de do tegumento fino, mido altamente vascularizado dos anfbios. Os rpteis que mergulham constituem um problema diferente. As tartarugas aquticas possuem uma carapaa espessa e, provavelmente
impermevel, mas podem usar a mucosa oral como auxiliar nas trocas gasosas quando permanecem submersas por longos perodos. Algumas tartarugas captam gua pelo reto, mas incerta a importncia disto para as trocas gasosas; mais provvel que elas utilizem gua para ajustar suas flutuabilidades. Para reduzir sua flutuabilidade, o animal expele ar dos pulmes, mas como a carapaa rgida, a reduo do volume pode ser compensada pela captao de gua (ou recolhendo os membros para dentro da carapaa).
Taxa metablica e tamanho corpreo
Mamferos
Retornemos sobre a discusso sobre os mamferos mergulhadores. As grandes baleias podem ficar submersas, mas o mergulhador muito pequeno como o masaranho aqutico, faz mergulhos que realmente
excedem meio minuto. De como geral, quanto maior for o mamfero mergulhador, mais longos so os mergulhes que eles podem realizar. A resposta que a taxa de consumo de oxignio relativa ao tamanho corpreo muito mais alta no mamifero pequeno do que no mamifero maior.
Custo energtico da locomoo
Nadar, correr e voar requer mais energia do que ficar sentado quieto, mais como pode se comparar os diferentes tipos de locomoo? Andar e correr so mais familiares para nos do que nada e voar e tem interesses mais imediato e, portanto, sabemos mais sobre esses modos de locomoo. Alm disso, o homem o nosso melhor experimental, porque ele extremamente cooperativo e, s vezes, at altamente motivado. Isso muito til nos estudos sobre desempenho mximo, porque quando um animal se recusa a correr mais rpido ou mais longe porque ele no quer - ou no pode? Os ces foram extensivamente estudados porque geralmente so cooperativos e fceis de trabalhar - s vezes, eles at parecem estar altamente motivados. O ar uma que est diferente, j que o homem tem experincia mnima em voar com a sua prpria fora, um nadador desajeitado ineficaz comparado com as focas, baleias e peixes. Quando comparamos os vrios tipos de locomoo temos que coonprender que as maiores diferenas entre mover-se na gua, na terra e no ar so atribuveis a diferentes qualidades fsicas desses meios. As duas diferenas mais importante resistem (1) no suporte do corpo animal, proporcionado pelo ar e pela gua (2) na resistncia diferente a realizao de movimentos que os dois meios apresentam. A maioria dos animais nadadores apresenta flutuabilida de quase neutra; o peso totalmente suportado pela gua circundante e no a esforo para suportar o corpo. Os animais que correm e voam esto em situao muito diferente eles precisam suportar todo o peso de seus corpos. Animal que corre tem apoio slido sob os ps, mas o animal que voa deve apoiar seu peso continuamente contra um fluido de baixa densidade e baixa viscosidade.
A resistncia do meio muito importante para o animal que nada, porque a gua tem alta viscosidade e densidade, em contraste os animais que correm e voam tem a vantagem de mover-se em meio de baixa viscosidade e baixa densidade. As caracteriza fsicas tem efeitos profundos sobre as adaptaes esturrais que os animais apresenta em seus modos de locomoo. Os animais aquticos tem corpos hidrodinamicos e geram autopropulso com nadadeiras e cauda ou modificaes desses estruturas. Os pssaros tm corpos aerodinmicos e asas que funcionam basicamente sobre os mesmos princpios da dinmica dos fluidos como a caudas dos peixes. Quanto menor o animal, contudo menos efetivo a hidro ou aerodinmica para reduzir a repetncia e aos pequenos obviamente, no so aerodinmicos. Os animais que correm usam as extremidades como a lavanca para remover-se sobre um substrato solido. A resistncia do ar pouca importncia aos animais que correm no so rticulamente aerodinmicos. A aerodinmica muito mais importante para os pssaros porque eles se movem muito mais rpido que os mamifero que correm e a resistncia do ar aumente aproximadamente com o quadro da velocidade. A maioria dos mamferos so quadrpedes mais algumas raridades como o homem e o canguru, so principalmente bpedes.
Tempo fisiolgico
Os animais pequenos vivem num ritmo muito mais acelerado que os animais grandes. Eles movem as pernas muito mais rapidamente e o corao bate mais depressa. Tudo acelerado e parece o tempo no relgio no pode o mesmo significado para os animais grandes. O corao de um musuranho bate 1000 vezes por minuto e do elefante talvez 30 vezes. necessrio 0,5 horas para que um corao de um elefante bata mil vezes, um nmero que o corao do musurano atinge em um minuto, a unidade de tempo no relgio deve ser um significado diferente nos dois animais.
6 Efeito da temperatura
A vida ativa dos animais limita-se a uma faixa restrita de temperatura, que varia alguns graus abaixo do ponto de congelamento da gua pura (0c)
at aproximadamente +50 graus. O enfoque agora ser dado a tenpera do organismo, e no do seu meio ambiente. Por exemplo, o ser humano mantm a sua temperatura corprea a 37C, independentemente se fica exposto ao um frio intenso ou se vai a uma sauna, onde a temperatura aproximadamente a do ponto de ebulio da gua. Os limites de temperatura para a vida animal so muito restritos se comparados a temperaturas csmicas.Apesar disso, h a ocorrncia de temperaturas adequadas no oceanos, e na maior parte da superfcie terrestre, pelo menos em partes do ano.
Terminologia confusa
Os termos cientficos correspondentes, poquilotrmicos, para animais de sangue frio e homeotrmicos para animais com sangue quente, so tambm imprecisos. A palavra poiquilotrmico refere-se ao fato de temperatura de um animal de sangue frio variar de acordo com o meio ambiente em que vive. um peixe tem a temperatura da gua em que vive e uma minhoca, a do solo onde encontrado. Entretanto, um peixe de guas profundas, que vive toda sua vida em guas que apresentam oscilaes trmicas dificilmente mensurveis, realmente um animal com a temperatura corprea praticamente constante. O lgico seria denoomin-lo um peixe
homeotrmico, porm esse termo usado especificamente para aves e mamferos, animais que na realidade apresentam temperatura corprea que normalmente auxiliam em vrios graus e que durante a hibernao podem at mesmo chegar a cerca de 0c animais que ocasionalmente apresentam temperaturas corpreas elevadas e bem reguladas e, outras vezes,
corportam-se de maneira similar aos animais de sangue frio, so frequentemente denominados de heterotrmicos. Animais homeotrmicos geralmente mantm uma temperatura corprea elevada e permanecem ativos em ambiente quentes ou frios, ao passo que a maioria dos animais poquilotrmicos ficam mais inativos devido ao fato de que a temperatura diminui. H algumas excees dignas de notas, como, por
exemplo, um largato que, sob o sol, consegue manter a temperatura bem acima aquela do ar ambiente. Para diferenciar o largato de aves e mamferos os quais mantm a temperatura corprea por meio de produo metablica de calor, dispe-se dos termos ectotrmico e endotrmico.
Adaptao a temperatura
Os limites da tolerncia temperatura para um lado animal no so fixos; a exposio a um temperatura prxima da letal frequentemente acarreta um certo nvel de adaptao, de modo que uma temperatura anteriormente seja tolerada. Com frequncia, o intervalo de tolerncia trmica diferente para a mesma espcie no vero e no inverno. Um animal universal geralmente tolera e permanece at mesmo ativo temperatura to baixa que seria letal a um animal estival; inversamente, o animal universal menos tolerante que um animal estival temperaturas elevadas. Essas modificaes na tolerncia a temperatura com as mudanas climticas so chamadas de aclimao. Efeitos semelhantes podem ser simulados em experimentos de laboratrios, mantendo-se animais para algum tempo a determinadas temperaturas. Para distinguir a adaptao ou ajustes, que ocorrem em experimentos de laboratrios da aclimatizao natural, a resposta as condies experimentais geralmente desligada pelo termo aclimatao.
7 A regulao de temperatura
A temperatura da maioria dos animais segue, de modo passivo, a temperatura do meio em que os circunda. Seria vantajoso se os animais pudessem no depender da imprevisibilidade dessa varivel ambiental e, neste captulo, trataremos do animais que mant a sua temperatura corprea mais ou menos independe da temperatura ambiental. Aves e mamferos vivem a maior parte de suas vidas em temperaturas corpreas que no oscilam mais que alguns graus. Esses animais sero os primeiros a serem considerados e, em seguida, ser discutido o que outros
animais conseguiram para manter suas temperaturas, independente da do ambiente. Em primeiro lugar, deve ser esclarecido o que se entende por temperatura corprea no sendo este, de modo algum, um conceito simples. Segundo, para a manuteno de uma temperatura constante, o ganho e perda de calor do organismo devem ser iguais. Para a compreenso desses dois processos, devemos estar familiarizados com a fsica de transferncia de calor. Em um ambiente frio, uma alta temperatura pode ser mantida pela reduo da perda de calor e o aumento do ganho de calor. A maioria das aves e mamferos fazem isso de modo bastante satisfatrio. Entretanto, alguns mamferos, e umas poucas aves, parecem desistir de resistir ao frio e permitem que suas temperatura caia vagarosamente, entrando em um estado de torpor ou hibernao. No entanto, a regulao no abandonada; ao contrrio, a hibernao um estado fisiolgico bem regulado. Em um ambiente quente, os problemas para a manuteno da temperatura corprea so inversos: o animal deve evitar que sua temperatura corprea aumente e, geralmente, forado a resistir pela evaporao da gua.
Regulao da temperatura no frio
Conservao de Calor Para a manuteno de temperatura, um animal deve se satisfazer
condio do estado de equilbrio no qual a taxa de produo de calor metablico equivale taxa de perda de calor. Por enquanto, ser considerado que a perda de calor (no frio) ocorre somente por conduo e radiao, e a perda de calor por evaporao pode ser desprezada.
Distribuio de material isolante
Devemos recordar que a pelagem dos animais rticos no consegue isolar, de modo uniforme, todas as partes do corpo; esses animais tambm necessitam de superfcies a partir da quais o calor possa ser dissipado, principalmente durante o esforo como a principal parte do corpo bem isolada para a reteno mxima de calor, eles precisam de reas recobertas
com pele fina nos ps, face e outras localizaes perifricas, a partir das quais possa ocorrer perda de calor quando aumenta a demanda para dissip-lo.
gua e regulao osmtica
O volume do organismo e a concentrao dos solutos devem ser mantidos dentro de limites bastantes estreitos. A razo que o funcionamento tico de um animal requer uma composio relativamente e bem definida de seus fluidos corpreos, e desvios substanciais so geralmente incompatveis com a vida. O problema que as concentraes adequadas dos fluidos corpreos dos animais, invariavelmente, diferem daquelas do meio ambiente. Os animais precisam manter concentraes apropriadas, porm as diferenas de concentrao tendem a diminuir, alterando o estado de condies internas. Os animais podem minimizar as dificuldades pela reduo da permeabilidade e dos gradientes de concentrao entre os fluidos corpreos e o meio, sendo que ambas as estratgias podem ser utilizadas.
Vertebrados aquticos
Esta sesso abordar, essencialmente, peixes e anfbios. No se contesta que as baleias e as tartarugas marinhas sejam vertebrados aquticos, porm, sero tratadas em um contexto diferente porque so descendentes de ancestrais terrestre e respiram ao ar, e mais conveniente discuti-las como animais terrestre que vivem em meio onde no h gua doce disponvel. 9 Excreo
As funes dos rgos excretores esto associadas a um princpio bsico: para a manuteno de um meio interno constante: qualquer material captado pelo organismo deve ser equilibrado pela remoo de igual quantidade de material que, por sua vez requer, das funes excretoras, uma capacidade
varivel, que pode ser ajustada para eliminar quantidades criteriosamente controladas de cada uma das diferentes substncias existentes. As principais funes dos sistemas excretores so:
1. Manuteno de concentrao apropriada de solutos 2. Manuteno de volume corpreo adequado. 3. Eliminao de produtos metablicos 4. Eliminao de substncias estranhas ou de seus produtos metablicos.
10 Movimento msculo biomecnicas
O nmero de mecanismos utilizados para a realizao de movimentos limitadas, embora os seus usos variem muitos. Sero discutidos trs mecanismos bsicos, os movimentos ameboide, ciliar e muscular. O movimento ameboide tem o seu nome derivado da ameba, um organismo unicelular descrito em qualquer livro de Biologia. A locomoo ameboide implica em grandes modificaes da forma das clulas, fluxos de citoplasma e atividade de psedpodes. A locomoo ciliar um modo caracterstico de movimentao protozorios ciliados, como o paramcio. Entretanto, os clios so observados em todos os filos animais e servem para uma variedade de funes como, por exemplo, os clios criam correspondente que movimentam guas atravs das brnquias dos bivalves ou movimenta fluido no sistema hidrulicos dos equinodermis. As vias respiratrias dos vertebrados que respiram o ar atmosfrico so revestida por clulas ciliadas que, lentamente, removem partculas estranhas que se alongam em suas superfcies. Os
espermatozoides da maioria dos animais movimentam-se com o auxlio de uma cauda, que em princpio, agem como um clio. O movimento muscular o mecanismo fundamental ultilizado em uma imensa maioria de diferentes movimentos. Ele depende dos usos de msculos, os quais em todo o reino animal, tm uma caracterstica universal - a capacidade de exercer encurtamento de fora dos mesmos.
Controle e integrao
Integrao significa a unio de partes. Em Fisiologia, essa palavra abrange o controle de todos os componentes funcionais, incorporados em um organismo que opera de forma integrada, na qual nenhum processo isolado pode ocorrer a um ritmo indenpente. As funes fisiolgicas podem ser controladas por hormnios ou pelo sistema nervos, mas h duas diferenas fundamentais: uma, est relacionada velocidade de ao e a outra, ao tamanho do alvo.
O controle e teoria do controle
O mecanismo de controle responsvel pela manuteno do estado de equilbrio nos organismos vivos constitui um dos principais captulos da Fisiologia e conveniente examinar seu funcionamento na teoria do controle simples. Por exemplo, aves e mamferos mantm a temperatura central aproximadamente constante, apesar de amplas variaes na temperatura externa e na produo interna de calor. Essa regulao de temperatura corprea realizada com o auxilio de um sistema de controle complexo.
12 Controle hormonal
Ento, o que se entende pelo termo hormnio? Um hormnio mais bem definido como uma substncia que liberada a partir de um rgo, de estrutura bem delimitada, e que exerce um efeito bem especfico sobre algumas outras estrutura bem distintas. Para que um hormnio afete um alvo ou um rgo especfico, o alvo deve ser capaz de receber o sinal e as clulas devem possuir receptores que recebem ao sinal. De igual importncia, outros rgos que entram em contato com a mesma concentrao de hormnio precisam se refratrios: eles devem ser desprovidos dos receptores que so essenciais para a ativao da maquinaria da clula que resulta na resposta das clulas.
13 Informaes e sentidos
A maior parte das informaes acerca do meio obtida por meio de rgos sensoriais especializados. Tradicionalmente, os rgos dos sentidos so devidos em exteroceptores que respondem a receptores do meio externo, como luz e som e os propioceptores que esto voltados as informaes internas, como a posio dos membros essa separao no tem muito significado, e, na melhor das hipteses, apenas uma questo de convenincia. Outra classifiao tradicional dos sentidos baseia-se nos cincos sentidos mais evidentes dos seres humanos: viso, audio, paladar, olfato e tato. Na realidade, o equipamento sensorial dos seres humanos no to limitado.
Qualidades sensoriais
A relao dos estmulos externos, aos quais, pelo menos alguns animais respondem, bastante extensa. As categorias relacionadas no so completamente distintas e a separao um tanto arbitrria. Entretanto, por convenincia, essa sequncia ser seguida na presente discusso acerca das possibilidades e limitaes que se aplicam ao uso de vrios tipos de informaes disponveis. As informaes, naturalmente, podem ser
subdivididas em trs categorias principais: energia eletromagntica e trmica, energia mecnica e agentes qumicos. A ampla variedade de estmulos e as enormes diferenas estruturais nos rgos sensoriais parecem causar confuso at verificarmos que a sequncia geral de eventos a mesma para todos. O estmulo externo emerge sobre um estrutura assessorial, que pode ser altamente complexa como o olho ou a orelha ou muito mais simples, com os receptores tteis da pele.
Atividades
1. O sistema respiratrio semelhante ao dos insetos, mas pode ter um papel em relao a estes, mas apresentam rgos respiratrios definidos que so perfundidos por sangue. De que espcies de animais estamos falando:
a. Insetos b. Moluscos c. Peixes d. Aracndeos e. Mamferos
2. Alguns dos ajustes mais simples para a troca de gases respiratrios so encontrados nos____________. Complete a lacuna.
a. Animais aquticos b. Aves c. Resteis d. Anfbios e. Aracndeos
3. O que Fisiologia?
a. A fisiologia trata da respirao dos animais b. A fisiologia trata de apenas do funcionamento dos rgos c. A fisiologia trata das funes dos organismos vivos - como se alimentam, respiram e se movimentam, e o que fazem para se manter vivos. d. A fisiologia trata das funes vitais apenas aos mamferos e. A fisiologia trata das funes de excreo
4. O podemos dizer que a respirao nos ovos?
a. Que a casca deve ser permevel ao oxignio, e consequentemente, tambm a outros gases, portanto, o vapor de gua e dixido de carbono liberado durante a incubao. b. Que no existe respirao nos ovos. c. Que a casca deve ser impermevel ao oxignio, e consequentemente, tambm a outros gases.
d. Que apenas dixido de carbono liberado durante a incubao. e. Que o ovo recm posto um microcosmo
5. Se a curva da dissociao for determinada para uma amostra de sangue de mamfero a __________ ambiente e depois a
_________corprea do animal, os resultados sero completamente diferentes. Assinale a alternativa que completa as lacunas.
a. Circulao / temperatura b. Temperatura / temperatura c. Temperatura / respirao d. Temperatura / excreo e. Atmosfera / excreo
Gabarito: 1 d /2 a /3 c /4 a /5 b 14. SENE, F. M. Cada caso, um caso... puro acaso Os Processos de evoluo biolgica dos seres vivos. Ribeiro Preto: SBG, 2009. Conhea a Obra: Livro-texto do professor de Histria Natural, Fbio de Melo Sene, do Departamento de Gentica da Faculdade de Medicina (FMRP), Cada caso, um caso... Puro acaso, aborda, de forma descontrada, e com exemplos de acontecimentos de seu cotidiano, os processos de evoluo biolgica dos seres vivos.
Ao terminar a leitura, acessvel tambm para aqueles que s tiveram acesso ao tema nas aulas de cincias, no ensino fundamental e no mdio, todos podem ter a sensao de que conheceram mais sobre evoluo de uma maneira diferente, de trs para frente. Utilizando sua experincia em sala de aula, o autor, logo no primeiro captulo, fala das dvidas da humanidade sobre a origem e evoluo do homem e as
hipteses cientficas para o fato. Descreve a teoria criacionista e evolucionista sem tomar partido, mas tambm sem deixar de lado a tendenciosidade e o ceticismo que acompanha a maioria dos cientistas. Neste captulo fala, ainda, sobre o medo da finitude que acompanha o homem e a criao do divino e misterioso para explicar o depois da morte. Cita o raciocnio lgico do matemtico francs Pascal:
Embora a existncia de Deus seja altamente improvvel, h uma grande assimetria na punio por errar no palpite. Portanto, melhor acreditar em Deus porque, se o palpite estiver certo, voc poder ganhar o cu e, se estiver errado, no far nenhuma diferena. Porm, se voc disser que no acredita em Deus, e estiver errada, a chance de ir para o inferno enorme e, se estiver certo, no far diferena.
Uma rpida explanao sobre a evoluo dos seres vivos por meio da seleo natural proposta por Wallace e Darwin e o impacto causado por ela, assim como as premissas e concluses dessa teoria esto no captulo dois. Neste captulo o professor explica de forma didtica a seleo natural, a origem das variaes e a teoria hereditria de Galton. Finaliza falando do descrdito teoria de Wallace e Darwin. Wallace e Darwin Tentar explicar por que s Wallace e Darwin tiveram, independentemente e quase simultaneamente, a mesma ideia para explicar a origem de tanta diversidade um exerccio puramente especulativo. Dizer que eles eram mais inteligentes, mais geniais ou mais espertos do que os outros tambm uma afirmao leviana. Porm, um fato muito importante aconteceu aos dois: depois de terem visto a grande diversidade biolgica em reas continentais sul-americanas, Darwin contornando o continente, e Wallace viajando pela Amaznia, ambos entraram em contato com a fauna e flora de arquiplagos. Darwin, em Galpagos, Wallace, na Malsia. E, o que diferencia os arquiplagos das reas continentais? A descontinuidade das populaes, isoladas uma das outras pelos limites das ilhas. A diferenciao entre populaes isoladas muito maior do que se elas tivessem uma distribuio geograficamente contnua e, embora o processo evolutivo atue igualmente nas ilhas ou nos continentes, o resultado da diferenciao fica muito mais evidente nas ilhas.
Charles R. Darwin e Alfred R. Wallace
O processo de evoluo dos seres vivos atravs da seleo natural foi imaginado por Wallace e Darwin quase simultaneamente. Diz-se que quase simultaneamente porque pode haver uma diferena temporal entre a ocorrncia da formulao por um e outro. Darwin estava na Inglaterra; Wallace, na Malsia, e o contato entre eles era precrio. A hiptese evolutiva tornou-se pblica atravs da apresentao em 1 de julho dec 1858, na Lynnean Society, em Londres, com o ttulo Sobre a Tendncia das Espcies de Formarem Variedades; e sobre a Perpetuao das Variedades e Espcies por Meios Naturais de Seleo.
A teoria causa impacto ao falar da evoluo do Homem
Em 1871, doze anos aps a publicao da Origem das Espcies, Darwin publicou o livro que realmente levantou a polmica sobre a teoria, uma vez que ele estendeu, explicitamente, para a espcie humana, todos os conceitos que havia postulado para os demais seres vivos, em 1858 e 1859. A publicao do livro A Descendncia do homem e Seleo em Relao ao Sexto (The Descent of Man, and Selection in Relation to Sex, 1871), por ter inserido o homem no restante do mundo animal, considerada uma das ideias que produziram os trs maiores traumas pelos quais passou a humanidade. Os outros dois traumas teriam sido infringidos por Coprnico, ao afirmar que a Terra no era o centro do Universo, e por Freud, ao postular que o ego estava sujeito a determinismos internos cuja verdadeira natureza permanecia inconsciente. A reao publicao de 1871, nos meios cientficos e na sociedade, foi imediata. Afirmar que a espcie humana teria surgido a partir de outra espcie e, portanto, no seria obra direta de uma criao, abalou a sociedade. Historicamente, consta que Darwin, ao ser pressionado, teria procurado Wallace como um aliado, porm Wallace no concordou com a extenso da teoria da seleo natural espcie humana. Segundo Wallace, a teoria s se aplicaria aos demais seres vivos e a origem do Homem seriam mais transcendentais. Assim, Wallace afastou-se da polmica e diminuiu sua responsabilidade pelas consequncias da teoria e, praticamente, abriu mo, em vida, de assumir a polmica sobre a teoria que propunha a evoluo dos seres vivos atravs da seleo natural.
O captulo trs, mesmo sendo mais apropriado para aqueles j iniciados no estudo da gentica, d uma viso geral sobre cromossomos, genes, divises celulares, fentipo e gentipo, mutaes gnicas, reproduo, tambm para
aqueles que engatinham na busca por entender esses processos. Conclui o captulo com o que nem o ser humano gosta de ouvir, a regra a extino e ao observarmos a natureza, estamos olhando para os sobreviventes e isso nos d a falsa impresso de que todos se adaptam ou especiam. Segundo ele, no documento fssil o registro mostra que a maioria das espcies extinguiu-se. Conceitos Bsicos para o Entendimento da Evoluo Biolgica.
A compreenso da complexa interao dos diversos fenmenos biolgicos responsveis pela enorme diversidade da vida fundamental para que se possa entender o processo evolutivo. Assim, apresento alguns fatos e conceitos bsicos para o entendimento do processo evolutivo e para possveis inferncias a respeito do mesmo.
S variaes hereditrias tm importncia evolutiva
As nicas alteraes e caractersticas que tm significado evolutivo so as que podem ser transmitidas hereditariamente. Todas as outras modificaes que os indivduos sofrem, ao longo da vida, e que afetam suas clulas somticas e no afetam o material gentico das suas clulas germinativas, no so transmitidas s geraes seguintes.
Os cromossomos
O nmero e a forma dos cromossomos de uma clula recebe o nome de caritipo (cario = ncleo). Todos os indivduos de uma mesma espcie e todas as clulas de um mesmo indivduo tm o mesmo caritipo. Os cromossomos formam pares e os cromossomos de um mesmo par recebem o nome de homlogos. Por exemplo, a espcie humana tem 23 tipos de cromossomos homlogos, ou seja, 46 cromossomos (que formam o conjunto diploide, 2n, gentipo). Nas drosfilas, o nmero 6 e o 2n 12. Os organismos pluricelulares de reproduo sexuada so formados a partir de uma clula-ovo resultado da funso dos gametas masculinos e femininos dos pais.
Os gametas so haploides, n e, ao se juntarem no ovo, forma, clulas diploides, 2n. Assim, a partir do desenvolvimento da clula-ovo, todo o organismo ser 2n.
Os genes esto nos cromossomos Os genes so as unidades do material gentico, esto localizados, ao longo dos cromossomos e, consequentemente, no se encontram soltos, como partculas, dentro do ncleo das clulas. No se sabe exatamente o nmero de genes de cada espcie, mas, com certeza, da ordem dos milhares. Como o sequenciamento dos genomas a estimativa atual para moscas do gnero Drosophila em torno de 13.600 genes, para os camundongos 20.200 e, para a espcie humana, 19.000. Como as espcies do gnero Drosophila tm 6 pares de cromossomos e o homem tm 23, evidente que cada filamento cromossmico possui milhares de genes.
Divises celulares: mitose e maiose
Aps a fecundao, a clula-ovo passa por um processo de seguidas divises celulares, chamadas mitose, dando origem s milhares ou milhes de clulas do organismo, as clulas somticas. Os principais eventos que ocorrem no ncleo das clulas durante a mitose so:
- duplicao cada filamento cromossmico se duplica, resultando um filamento original e um filamento cpia, que permanecem ligados por uma regio chamada centmero. - separao a seguir o filamento cpia de cada cromossomos se separa do filamento original e cada um deles segue para o ncleo das duas clulas resultantes da diviso. Como isso ocorre com todos os cromossomos, os dois ncleos resultantes recebem o mesmo nmero e tipo de cromossomos e cada um contm 2n, iguais clula inicial da qual so originrios. O processo iniciado com uma clula 2n,e termina com duas clulas 2n.
Os gametas, espermatozoides e vulos, so formados a partir das clulas germinativas, localizadas, respectivamente, nos testculos dos machos e nos ovrios das fmeas. As clulas germinativas so 2n, ou seja, possuem os filamentos cromossmicos aos pares e os gametas, a que do origem, so n, ou seja, tm apenas um filamento cromossmico de cada par. O processo de diviso celular que resulta na formao dos gametas a meiose. Durante a meiose, os principais eventos que ocorreram no ncleo das clulas germinativas so: - duplicao filamento cromossmico se duplica, resultando num filamento original e num filamento cpia, que permanecem ligados pelo centrmero. - separao dos cromossomos neste momento ocorre a primeira diferena em relao mitose porque, ao se dividir em duas clulas, os cromossomos homlogos se separam e cada um vai para uma celular nova. Assim, cada clula nova tem apenas um cromossomo de cada par, duplicado (original e cpia); Separao entre os originais e as cpias a segunda diferena em relao mitos que, em seguida, cada clula nova entra novamente em processo de diviso celular sem que ocorra, previamente, duplicao cromossmica, e os filamento originais de cada cromossomo se separam das cpias e cada um vai para uma celular nova. Assim, o processo inicia-se com uma clula 2n, na qual ocorre duplicao, sofre uma diviso, que separa os pares de cromossomos homlogos e sofre mais uma diviso que separa filamentos originais de cada cromossomo de suas cpias, dando origem a quatro gametas n. cada um com um filamento cromossmico de cada par.
Fentipo e gentipo
O termo fentipo refere-se aparncia estrutural ou constituio metablica de um organismo, relutante da interao entre a ao do gentipo e do ambiente em que o organismo viva ou tenha se desenvolvido. Entende-se como ao do ambiente tudo que no seja decorrente da ao do gentipo. Gentipo, o conjunto diploide, ou 2n, de material genrico de um indivduo, formado por metade de cromossomos herdado do pai (n) e a outra metade, da me (n), os quais formam os pares de cromossomos homlogos.
Locos, genes e alelos
A religio do cromossomo onde se localiza um gene recebe o nome de loco. Numa clula, quando o gene de um loco sofre uma mutao, surge um gene novo que, ao ser copiado na duplicao, ocupa o mesmo loco do gene original no cromossomos. Assim, passam a existir cromossomos com o gene original e cromossomos com a cpias mutante. Genes diferentes (em decorrncia de mutaes) e que ocupam o mesmo loco de cromossomos homlogos so chamados de genes alelos.
O material gentico
O material gentico est contido nos cromossomos. Sob o ponto de vista qumico, os cromossomos so formados basicamente por protenas e cidos nuclicos. O cido nuclico chamado DNA (cido desoxirribonuclico) a substncia qumica que contm todas as informaes genticas da maioria dos seres vivos (em alguns o RNA cido ribonuclico que executa essa funo). Essas molculas codificam, na sequncia e organizao das suas bases nitrogenadas, todas as informaes hereditrias de cada indivduo. So essas informaes que organizam o arranjo dos aminocidos para a sntese de todas as protenas em cada organismo. Cada segmento do DNA, responsvel pela sntese de uma protena, chamado gene ou cistron. Dependendo da funo que a protena sintetizada venha a exercer no organismo, o gene chamado de estrutural, quando afeta a estrutura ou a forma de um carter, ou regulador, quando afeta a manifestao de outros genes. Antes de cada diviso celular, ocorre a duplicao das fitas de DNA (com o mesmo resultado de uma cpia por contato), e uma das duas novas clulas recebe o DNA original e a outra clula recebe a cpia. Assim, sucessivamente, a capacidade de autoduplicao permite que cpias do DNA do ovo no momento da fecundao, metade recebido da me e metade recebido do pai, estejam presentes em todas as clulas do indivduo adulto.
Mutaes gnicas
As mutaes gnicas so alteraes no DNA decorrentes de alteraes nas bases nucleotdicas que contm toda a informao do material gentico e fazem parte do cdigo gentico. Mudana de uma base ou da ordem das bases altera o produto final do gene. Na maioria dos casos, as mutaes ocorrem naturalmente no momento da duplicao do DNA. A mutao gnica s pode ser passada aos descendentes se ela ocorrer em clulas germinativas.
Mutaes ocorrem sobre sequncias preexistem de DNA
Um dos fatores que torna o processo evolutivo conservador o fato de as mutaes gnicas ocorrerem em uma sequncia de DNA preexistente e, por isso, fica afastada a possibilidade de aparecimento de uma estrutura totalmente nova devido a um evento mutacional nico. Uma mutao num segmento codificante de DNA (um gene), que seja, por exemplo, responsvel pela produo de melanina, que determina a cor da pele do indivduo, afeta, exclusivamente, a cor da pela de seu portador, ou seja, no possvel que mudanas nesse gene dem origem a rgos novos ou estruturas novas e isso vlido para qualquer outro segmento de DNA ou carter que se considere.
Reproduo sexuada
Em algum momento do processo evolutivo surgiu a reproduo sexuada. Por este tipo de reproduo, cada indivduo precisa interagir com outro indivduo para se reproduzir. A reproduo sexuada depende do processo de diviso celular chamado meiose. Pelo fato de o processo da meiose, que produz os gametas, ser basicamente o mesmo no reino animal, no reino vegetal e nos fungos, pode ser afirmado que a reproduo sexuada surgiu antes da separao evolutiva desses grupos de organismos, ou seja, no incio da diversificao da vida.
Reproduo assexuada
O outro tipo de reproduo, a assexuada, ocorre por duplicao do DNA ou Rna, como nos vrus, ou por simples diviso celular (como na maioria dos micro-organismos).
Todos os seres vivos atuais tm uma origem nica
Todos os indivduos de todas as populaes e colnias de seres vivos existentes atualmente tm uma relao ancestral descendente com a origem da vida. Por hiptese, mesmo que se admita que no incio possam ter surgido vrias linhas de vida independentes, certamente apenas uma delas deu origem a todos os seres vivos atuais. A afirmao possvel porque entre a origem de uma primeira estrutura, capaz de se autoduplicar, e a origem de uma clula, decorreram 2 a 3 bilhes de anos. A primeira estrutura, provavelmente uma molcula de RNA, no tinha metabolismo prprio e dependia de elementos do meio ambiente para sobreviver.
Evoluo no significa progresso
Evoluo biolgica no sinnimo de progresso. Uma das causas do mau entendimento desse assunto uma questo semntica de o processo ser chamado de evoluo. A palavra evoluo, alm da ideia de mudana, sugere uma mudana para melhor. Talvez, no sculo 19, quando esse termo foi usado para denominador mudana dos seres vivos, ele at tenha todo tal conotao. Porem, hoje, quando se fala em evoluo biolgica no se pensa em progresso e sim na capacidade de sobrevivncia da populao, da espcie ou do grupo de espcies. Nessa avaliao, todos os organismos vivos atuais so igualmente evoludos: os micro-organismos, todas as plantas, insetos, vermes, peixes, rpteis, aves, mamferos, embora no signifique que eles sejam todos iguais, biologicamente. evidente que cada um tem suas peculiaridades e complexidades, mas todos tm em comum o fato de estabelecerem uma relao ancestral/descendente com a origem da vida.
A regra a extino
Como consequncia do processo evolutivo uma populao pode, ao longo das geraes: permanecer adaptada ao ambiente, especiar ou extinguir. Por adaptao entendem-se boas condies de sobrevivncia no ambiente e, por especiao, quando as mudanas produzem uma diferenciao a ponto de a populao vir a ser considerada uma nova espcie. Num ambiente estvel, as populaes tendem a permanecer adpatadas e inalterada ao longo do tempo. Quando o ambiente muda na esmagadora maioria das vezes a populao extinta. A extino de uma populao no significa necessariamente a extino da espcie, pois para que uma espcie se extinga, preciso que todas as suas populaes se extingam. A adaptao vento raro e a especiao, extremamente raro. Assim, regra a extino. Ao observarmos a natureza, estamos olhando para os sobreviventes e isso nos d a falsa impresso de que todos se adaptam ou especiam. No o que observamos no documentrio fssil, cujo registro mostra que maioria das espcies extinguiu-se.
As leis de Mendel e o surgimento da gentica abrem o captulo quatro, que tem tambm teoria sinttica ou neodarwinismo, que rene todo o conhecimento dos mutacionistas e dos selecionistas, que aceitam plenamente a ideia de seleo natural de Darwin, completando-se com a informao que faltava: a origem da variao.
As leis de Mendel
Muitos tentam especular a respeito do trabalho de Gregor Johann Mendel (1822/1884) e sobre qual teria sido a consequncia caso Darwin tivesse tomado conhecimento a respeito das leis de Mendel que explicavam os mecanismos de transmisso dos caracteres do pas para filhos e, consequentemente, a hereditariedade das variaes. Na realidade, publicado em 1866, e redescoberto em 1900, o trabalho de Mendel s foi totalmente aceito em 1910 e s foi ligado teoria de Darwin entre as dcadas de 1930 e 1940, quando seus princpios foram ampliados s populaes e associados ideia de seleo natural, com centenas de pesquisadores no mundo todos se dedicando ao assunto. Imaginar que Darwin, sozinho pudesse ter entendido as leis de Mendel e feito a correlao delas com sua teoria, exigir demais de sua genialidade.
O surgimento da gentica
A descoberta do trabalho de Mendel em 1900 e sua ampla aceitao em 1910, marca o incio de uma cincia, a Gentica. Embora j decorridos mais de 100 anos, a Gentica considerada uma cincia recente se comparada a outras reas a Biologia, a Botnica, a Zoologia, a Fisiologia. Nos 34 anos que se seguiram a 1876, at o trabalho ser redescoberto vrios conhecimentos biolgicos novos convergiram para a hiptese matemtica de Mendel. Entre as mais diretamente correlacionadas, temos: - a descrio da diferena entre clulas somticas e germinativas, ficando resolvida a questo da origem dos gametas; - a descrio das estruturas da clula e as do ncleo; - o entendimento da fecundao a descrio da unio dos gametas (vulos e espermatozoides); - a descrio dos cromossomos; - a descoberta de que os gametas contm metade dos cromossomos das clulas somticas; - a sugesto da correlao entre cromossomos, ncleo e hereditariedade (transmisso das caractersticas hereditrias de pais para filhos); - a descrio da mitose e meiose.
Mesmo assim, s em 1902, dois anos aps a redescoberta dos trabalhos de Mendel quer foi publicado o trabalho de Walter Sutton e Theodor Boveri, estabelecendo a correlao entre meiose e a hiptese de Mendel.
Fentipo = gentipo + meio ambiente
Foi Wihelm Johannsen (1857-1927) que, nos primeiros anos do sculo 20, props o uso do termo gene, derivando o nome do tempo pangeneses usado por Darwin, para os fatores Mendelianos. Dele tambm a criao dos termos gentipo e fentipo.
Mutacionistas versus selecionistas
O termo mutao j havia sido usado por DeVries ao descrever as alteraes que ele encontrou no cultivo de Oenothera lamarckiana, uma planta de jardim conhecida por prmula. Como as variaes que surgiam nas prmulas em seu jardim, a partir da forma abrupta. DeVries props uma nova teoria evolutiva. Segundo a teoria a evoluo se daria ao saltos, de uma gerao para outra, em decorrncia das mutaes que seriam as nicas responsveis por provocar alteraes nos organismos vivos. Nas trs primeiras dcadas do sculo 20, com a denominao geral de mutacionista, foi a teoria evolutiva predominante apoiada pelos principais geneticista da poca, incluindo Morgan e seu grupo. A hiptese de saltos, proposta DeVries, e conceito fundamental para os mutacionistas no consideram a seleo natural proposta por Darwin como fator evolutivo, uma vez que, por seleo natural, o processo de mudanas seria lento e gradual. Nessas primeiras trs dcadas, os mutacionistas s foram contestados pelos ditos selecionistas defendendo a ideia da seleo natural como responsvel pelo processo evolutivo e, medida que foram sendo acumulados exemplos da seleo natural, ficava cada vez mais difcil ignor-la. Mais uma vez os dados mostravam o caminho para explicao dos fenmenos. Entre os exemplos da poca sugerindo a ao da seleo, destacam-se: - o melanismo industrial das mariposas inglesas; - o mimetismo em borboletas, especialmente envolvendo as espcies monarca e vice-rei; - os pssaros de Galpagos; - anemia falciforme; - diversas doenas da espcie humana e de outros animais.
Teoria sinttica ou neodarwinismo
Aps 3 dcadas, foi possvel conciliar as discusses dos selecionistas e dos mutacionistas e o resultado desta juno conveniou-se chamar de moderna sntese ou teoria sinttica, porque se preocupava acomodar uma sntese todo o conhecimento dos mutacionistas e dos selecionistas. tambm
chamada neodarwinismo, uma vez que aceita plenamente a ideia de seleo natural de Darwin, completando-a com a informao que faltava: a origem da variao.
Premissas da teoria sinttica
A teoria sinttica, ou neodarwinismo, foi estabelecida baseada em premissas e as principais so: 1- o processo evolutivo resultado das foras geradas pela taxa de mutao e pela presso de seleo; 2- as mutaes gnicas ocorreram ao acaso e so responsveis por toda a variao gentica encontrada nas populaes; 3- a seleo natural molda as populaes s condies ambientais atravs de um processo de eliminao das variveis (mutaes) no adaptadas e favorecimento das variveis que aumentam a adaptao dos indivduos ao ambiente. Toda variabilidade gentica seletiva, ou seja, se para um dado carter existirem duas formas na mesma populao, uma melhor do que outra e acaba por elimin-la ao longo das geraes; 4- a variabilidade gentica nas populaes pequena em decorrncia da ao da seleo natural; 5- todas as populaes so infinitamente grandes. Estatisticamente, para uma populao ser considerada como infinitamente grande ela deve ter, no mnimo 1000 indivduos; 6- a migrao e o sistema de acasalamento so fatores complementares. Entende-se por migrao a ocorrncia de fluxo gnico entre populaes. O sistema de acasalamento pode ser ao acaso ou preferencial. Ao acaso, quando a frequncia de cruzamentos entre indivduos com caractersticas fenotpicas diferentes depende exclusivamente das frequncias destas caractersticas. Quando em uma populao os cruzamentos ocorrem ao acaso, ele considerado panmtica. O acasalamento considerado preferencial, quando diferentes caractersticas genticas dos indivduos de uma populao interferem na formao de casais e tipos semelhantes preferem cruzar entre si e os cruzamentos deixam de ser ao acaso. Nesse caso a populao no panmtica. O fato de a populao ser a unidade do processo evolutivo significa que o processo no ocorre em outro nvel taxonmico a no ser na populao. Portanto, mesmo diferenas grandes como as encontradas entre populaes
pertencentes a famlias diferentes, como os Feldeo e os Candeo, por exemplo, so decorrentes de alteraes sofridas pela populao ancestral dos dois grupos e acumuladas, aps a diferenciao, ao longo das geraes. Todas as populaes atuais destes animais tm uma relao ancestral/descendente com a populao de carnvoros, ancestral das duas famlias.
Tomando como base as premissas, foi considerado que:
1- como as populaes eram infinitamente grandes, a evoluo teria como fator casual apenas a mutao, uma vez que a ocorrncia de erro amostral (deriva gentica) seria desprezvel; 2- as populaes seriam moldadas, ajustadas ao ambiente pela ao da seleo natural; 3- como as populaes eram infinitamente grandes, o processo de substituio gnica ocorreria de forma lenta e gradual, sem deriva gentica e, consequentemente, o processo de diferenciao de populaes seria lento e gradual. Em decorrncia, algumas das questes eram: o quanto a populao precisaria mudar geneticamente para que surgisse uma espcie nova, e quanto tempo demoraria? Polimorfismo genticos
Uma populao Polimrfica quando, para um mesmo carter, pode ser encontrada mais de uma forma na populao. O Polimorfismo chamado gentico quando as diferentes formas so decorrentes de mutaes genticas. Quando ocorre uma mutao e ela passada s geraes seguintes, a populao inicialmente polimrfica em relao quela caracterstica, ou seja, contm mais de uma forma da mesma caracterstica porque convivem indivduos que possuem alelos mutados e outros que so portadores do alelo original. Nem sempre possvel saber, numa populao polimrtica, qual o alelo original e qual o alelo mutante. Para a teoria sinttica, todo Polimorfismo transitrio, ou seja, toda vez que num mesmo loco existam dois ou mais alelos diferentes, um deles dar maior capacidade adaptativa a seu portador e ser favorecido pela seleo natural que, nesse caso, atua de forma purificadora. Assim, o Polimorfismo transitrio e perdurar pelo nmero de geraes necessrio para que os alelos menos adaptados sejam eliminados.
A teoria sinttica tambm assunto para o captulo cinco. onde ele descreve a deriva gentica, quando a composio gentica de uma gerao diferente da gerao anterior por erro amostral na passagem de gametas de uma gerao para outra - descrita em 1932, e s reconhecida como importante 48 anos depois, e o caso particular dessa deriva, chamado de efeito fundador. O captulo seis foi reservado para a discusso sobre os fatores evolutivos. Entre as afirmaes a de que o resultado da seleo natural no casual e sim um fator determinstico resultando ou na extino ou na maior adaptao das populaes.
Os Fatores Evolutivos
Os processos biolgicos capazes de provocar alteraes na composio gentica de uma populao, de uma gerao para outra, so denominados fatores evolutivos. Numa frase reducionista, vrios geneticistas afirmam que a evoluo simplesmente mudana nas frequncias gnicas de uma populao, ao longo das geraes.
Alterao da composio gentica das populaes
Se a unidade evolutiva a populao, para que se possa compreender o processo evolutivo fundamental que se entenda a dinmica das mudanas do material gentico nas populaes e como atuam os fatores evolutivos responsveis pelas adaptaes, especiao e extines.
O teorema de Hardy-Weinberg
Enfim, se no ocorre mutao, se no ocorrer migrao, se no ocorrer seleo, os cruzamentos forem ao acaso e a populao permanecer grande, as frequncias gnicas e genotpicas permanecero constantes ao longo das geraes. Este o enunciado do Teorema proposto por Hardy e por Weinberg em 1908, considerado a base terica de Gentica de populaes. Para os autores do teorema, uma populao que obedea a todas as condies citadas uma populao em equilbrio. Resumindo, podemos dizer: - se nada acontecer, nada acontecer.
Assim, mutao gnica, migrao, deriva gentica, seleo natural e padres de cruzamentos so os nicos fatores evolutivos. Ao longo das geraes, todas as populaes esto sujeitas a sofrer alterao de cada um desse fatores. Da atuao dos fatores evolutivos resulta o processo evolutivo.
Os fatores evolutivos
Normalmente ocorre interao entre os fatores evolutivos e pode tambm haver variao de intensidade e de importncia para cada um. Essa interao e intensidade tambm podem vrias de uma gerao para outra da mesma populao, ou seja, alm da variao da composio gentica dos indivduos de cada populao, h variao tambm na atuao dos fatores evolutivos em decorrncia das condies ambientais onde vive a populao.
Fontes da variabilidade populacional
A variabilidade gentica de uma populao pode decorrer de mutao gnica, de recombinao cromossmica (tambm conhecida como recombinao gentica) e de migrao.
Mutao gnica
A mutao gnica, desde a origem da visa, a nica fonte primria de surgimento de material genrico novo. resultado de alterao na estrutura do DNA e ser passada aos descendentes se ela ocorrer em clulas germinativas. A mutao gnica um evento considerado ao acaso, quando ao momento de ocorrncia. O fato de ser evento ao acaso, quanto ao momento, significa que a ocorrncia de uma mutao gnica independe de qualquer previso ou de qualquer objetivo adaptativo em resposta a uma presso ambiental. O efeito, ou consequncia fenotipca, causado pelo novo alelo, varia dentro de um conjunto fechado de possibilidades, ou seja, quando um segmento de DNA, responsvel por uma caracterstica muts, o alelo mutante
alterar a manifestao fenotpica do alelo original e, consequentemente, afetar a manifestao do carter que ele determina.
Migrao
Migrao de indivduos para dentro ou para fora da populao tambm pode afetar a composio gentica populacional.
Por uma questo de qualidade, a migrao pode ter um impacto muito maior do que a mutao em termos de introduo de material gentico novo numa populao, se os migrantes forem geneticamente diferentes da populao. A importncia da migrao realada pelo fato de a maioria das espcies serem formadas por vrias populaes e estas por subpopulaes ocasionais de indivduos. Os migrantes geram fluxo gnico entre as subpopulaes diluindo eventuais diferenas genticas que tenham surgido durante o isolamento entre ela.
Seleo natural
A seleo natural proposta por Darwin w Wallace como sendo o principal fator evolutivo, embora seja o mais estudado e discutido e o que acumula maior nmero de trabalho e pesquisa, o mais polemico dos fatores . Quando tal acontece, porque o fenmeno ou processo estudado complexo cientificamente, pois, quando o problema simples, quando mais se estuda, quanto mais se acumula conhecimento, menos, dvidas permanecem. Porm, no caso da seleo natural, estudada h 150 anos, a polmica continua a principal questo atual que ela foi concebida, desde Darwin at a dcada de 70, como um agente selecionado as variveis indesejveis, o que, ao longo das geraes deveria purificar as populaes. Porm, com o avano das tcnicas moleculares, demonstrou-se que a variao das populaes era muitas vezes maior do que a prevista por Darwin, ou o que seria esperado se a seleo natural fosse purificadora.
Seleo natural igual luta pela sobrevivncia?
Embora a seleo natural tenha sido definida e entendida como luta pela sobrevivncia, dificilmente envolve combates fsico entre os competidores. Os animais mais velozes casaram melhor, alimentaram melhor seus filhotes, o que far com que, dentro de uma populao de guepardo, os mais velozes se reproduzem mais do que os menos velozes. A seleo favorvel dos mais velozes contra os mais lentos se processa sem que aja luta direta entre eles. Por questes de limitaes biolgicas no esperado que a seleo produza sempre animais mais rpidos a cada gerao que a espcie possuem.
O resultado da seleo natural no casual
Quando se diz que a evoluo um processo ao acaso, no sentido de que ela no tem direo nem objetivo. No entanto, a seleo natural no considerada um fator casual (o estocstico). Na realidade, um fator determinstico resultando ou na extino, ou na maior adaptao da populao ao ambiente, pois sobrevivem os mais adaptados a cada gerao. A alterao da composio do conjunto gnico das populaes, em cada gerao, favorecendo a sobrevivncia dos mais adaptados a um determinado ambiente, e o resultado da seleo. A seleo natural no cria estruturas novas ou materiais genticos novos, ela no interfere na frequncia dos existentes. De maneira que, ao permanecerem os melhores, no necessariamente estaro sobrevivendo tipos ideias. O processo como todo casual em decorrncia da incerteza sobre a qualidade e a quantidade da variabilidade que estaro sobre seleo, e essa seleo ainda poder variar de acordo com mudanas ambientais.
Padres de seleo
A seleo natural tem trs modos bsicos de expresso: como seleo direcional, estabilizadora ou disruptiva. Ela direcional quando os favorecidos so os fentipos extremos de uma distribuio normal da varivel, resultando em aumento de frequncia deles ou longo das geraes.
A seleo estabilizadora sobre um carter quando, na populao, os tipos intermedirios tem maior valor adaptivel. Isso acontece na maioria dos casos, quando o meio ambiente permanece constante por um longo perodo de tempo. A seleo disruptiva quando favorece tipos extremos para um dado carter, em detrimento do tipo mdio. E o contrario da seleo estabilizadora e o mesmo que seleo direcional, agindo simultaneamente em sentido opostos, favorecendo os dois extremos de uma distribuio norma. Ela pode ocorrer em duas situaes:
- primeira, se a rea de distribuio da populao for muito ampla ou diversificada e os indivduos estiverem expostos a diferentes presses de seleo em locais diferentes; - segunda, se houver polimorfisno para um carter na populao e os cruzamentos forem preferenciais entre indivduos semelhantes.
Depois de anos de estudos e palestras, o pesquisador ainda deixa transparecer sua admirao pelo processo de adaptao das populaes ao ambiente e relata ser este um dos fenmenos mais interessantes que ocorrem na natureza e que serviram de base para a teoria evolutiva de Wallace e Darwin. Esse tema amplamente discutido no captulo sete que traz, inclusive, exemplo de adaptao como a resistncia das moscas a inseticidas, das bactrias a antibiticos, assim como o caso dos coelhos na Austrlia e dos pardais na Frana. A seleo artificial criada pelo homem tambm ganha destaque neste captulo. Para discorrer sobre as interaes entre os efeitos evolutivos, o pesquisador dedica todo o captulo oito. Para isso faz uma reviso dos conceitos sobre os fatores evolutivos, como por exemplo, a deriva gentica, mais a endogamia, mais seleo que igual a processo evolutivo mais rpido. Nos captulos nove e 10, o autor faz uma descrio sobre a diferenciao entre as populaes e origem das espcies. Evoluo dos grandes grupos
A origem dos grandes grupos taxonmicos decorre da somatria de eventos que ocorrem nas populaes das espcies. O tempo decorrido e o
desaparecimento dos tipos intermedirios so os responsveis pelo aumento das diferenas entre os grandes grupos. muito difcil imaginar que existia qualquer tipo de direo no processo evolutivo uma vez que todo ele baseado nos processos de especiao e adaptao que ocorre sempre nas populaes das espcies.
Os grandes grupos.
A espcie, formando pelo conjunto de suas populaes, a nica categoria sistemtica zoologia que tem existncia concreta. O agrupamento de indivduos numa espcie no artificial, pois entre os indivduos de espcie existente uma ligao natural que a reproduo. As demais categorias sistemticas, inventadas pelo homem (Gnero, Famlia, Ordem, Classe e Filo) so agrupamentos de espcies. Quanto mais natural for a classificao, ou seja, quanto mais prxima estiver da historia evolutiva das espcies, mais estes agrupamentos refletem a real historia do grupo. A assim sendo, esperado que todos as espcies que formam um gnero seja descendentes de uma nica espcie ancestral comum. Este raciocnio a valido para todos as categorias sistemticas superiores como Famlia, Ordem, Classe, Filo e Reino. O conjunto de espcies de um gnero escolhido pela semelhana entre elas e, na grande maiorias das vezes, no possvel estabelecer, na pratica, a relao filogentica (ou relao de parentesco) que decorre da relao ancestral/descedente entre essas espcies. Por critrios semelhantes, como um conjunto de espcies formam um gnero, um conjunto d gnero forma um famlia e assim por diante. Durante muito tempo alguns cientistas defenderam a ideia de que as micromutaes seriam responsveis pelo aparecimento de pequenas diferenas acumuladas, acabariam por separar uma populao, enquanto as macromutaes seriam mutaes com grandes efeitos, que tornariam seus portadores diferentes a ponto de serem classificados como outra Espcies, outro Gnero, outra Famlia e Ordem. Na realidade, a conceituao esta errada, pois, normalmente, as mutaes que causam grandes alteraes so deletrias, reduzindo a viabilidade dos seus portadores. No perodo longo de tempo o acumulo de mutaes que causam pequenas alteraes, as responsveis as grandes diferenas entres as categorias superiores. Portando, o mesmo processo que d origem s espcies, atuando um grupo de espcies por um longo
perodo de tempo, acaba por diferenci-lo dos demais em suas categorias bsicas.
Irradiao Adaptativa
A irradiao adaptativa o processo pelo qual dentro de um grupo a taxa de diversificao muitas vezes maior que a de extino em certo perodo de tempo. A irradiao pode ocorrer quando o grupo de organismo adquire uma caracterstica nova que lhe permite explorar de forma diferente o ambiente em que vive ou quando o grupo invade um territrio novo desocupado. Com a expanso do grupo, sero formadas novas populaes as quais podero dar origem as novas espcies, das quais surgiram outras novas populaes, de tal forma que, num intervalo de tempo relativamente curto, nmerosas espcies passam a ter diferentes caractersticas adaptativas.
Constncia Evolutiva
Algumas espcies ou grupo de espcies atravessam longos perodos d tempo sem se alterem de forma substancial, configurando o que chamado de constncia evolutiva. Alguns organismos chegam at a receber a denominao de fssil vivo.
Convergncia Evolutiva
Denomina-se convergncia evolutiva a situao em que organismos com diferentes origens filogenticas e diferentes estruturas, quando submetidos a um mesmo ambiente com o mesmo problema com o mesmo problema adaptativo, evoluem para formas ou solues semelhantes. Ocorre uma convergncia quando forma do organismo. So exemplos so exemplos clssicos de convergncia evolutiva os mamferos aquticos, como as baleias e os golfinhos, que adquiram a forma externa semelhante de peixes, embora originrios de ancestrais terrestres. A distancia filogentica/evolutiva entre mamferos e peixes muito grande. A volta de um mamfero terrestre para a vida aqutica, adquirindo u aspecto externo de peixe, a chamada convergncia evolutiva, no significa
que necessariamente todo vertebrado de origem terrestre, que passe av viver na gua, tenha de adquirir a forma de peixe; basta observar por exemplo, o caso das tartarugas marinhas.
Analogia e Homologia
Quando organismos diferentes apresentam estruturas com forma e funo iguais, porm com origens embrionrias diferentes, essas estruturas so chamadas anlogas. Assim, as asas dos insetos e as asas dos pssaros so rgos anlogos. Quando organismos diferentes apresentam estruturas com forma e funo diferentes, porm com a mesma origem embrionria, essas estruturas so chamadas homologas. Os braos do homem, as patas dianteiras dos mamferos quadrpedes, as nadadeiras das baleias e a asa das aves, so rgos homlogos.
Documento fssil como evidncia direta da histria evolutiva est descrito no captulo 11, que traz, ainda, os processos de fossilizao, a representatividade do documentrio fssil, dificuldades para localizao de fsseis, o tempo geolgico e a escala do tempo evolutivo, mudanas na diversidade ao longo do tempo. O Documentrio Fssil
A rea da cincia que estuda os fosseis a Paleontologia. Fsseis (do termo latino fossile que significa desenterrado) do quaisquer restos ou vestgios preservados de plantas, animais, ou de qualquer outro ser vivo. As caractersticas dos restos podem ser: um organismo inteiro, como no caso dos mamutes congelados da sibria; esqueletos ou parte deles: troncos de plantas petrificados, ou ainda impresses deixadas nas rochas por pegadas ou por folhas. Qualquer vestgio de vida passada, como por exemplo, os coprlitos, que so fezes de animais preservadas naturalmente pela dessecao ou por mineralizao, considerado fssil. O registro fssil uma evidncia direta, concreta, da historia evolutiva dos seres vivos cuja validade e aceitao dependem muito de evidncias desse registro. Estudos de parentescos entre os grupos, atravs de anlises filogenticas, com o uso de marcadores morfolgicos ou genticos ou genticos, so evidncias indiretas da historia evolutiva .
A maioria dos fosseis encontrada em rochas sedimentares, embora existam excees como, por exemplo, fsseis no gelo ou mbar . O mbar uma resina vegetal e tem cor amarela semitransparente. Dentro dessa resina tm sido encontrados fosseis de polens, cianobactrias e outros organismos unicelurares, embora os mais conhecidos sejam os fosseis de invertebrados, como besouros, mosquitos, vespas, aranhas, e pequenos vertebrados como anfbios. A importncia do mbar que nele esto seres que normalmente se desintegram depois da morte e, alm disso, os fosseis preservados no mbar so os melhores porque normalmente o organismo preservado por inteiro.
Os Processos de Fossilizao
Embora a preservao de matria orgnica como fssil no seja um evento raro na natureza, a preservao de restos delicados mais rara porque pode ocorrer a destruio do fssil por decomposio. Em decorrncia, fsseis das partes moles dos organismos so raros. No entanto, a preservao de partes esquelticas biomineralizadas, mais duras e resistentes eroso e decomposio, como os dentes, as conchas, as carapaas e os ossos so muito mais comuns e, consequentemente, a maior parte do registro fssil constituda por fsseis desse tipo. Biomineralizao o processo chamado histometabase pelo qual a matria orgnica se transforma se transforma em matria mineral. As matrias orgnicas so substitudas por mineral, mais frequentemente calcita, molcula por molcula, resultado numa reproduo perfeita do organismo, nos mnimos pormenores celulares. De qualquer forma, para que os restos de qualquer organismo sejam fossilizados, imprescindvel que sejam recobertos por sedimentos logo aps a morte e permaneam enterrados durantes todo o processo de fossilizao. Assim, tiveram muito mais chances de se tornarem fosseis os animais e plantas que viveram em ambientes aquticos, de plancies alagadas e mesmo animais terrestres de plancies no alagadas, formadoras e formadas por rochas sedimentares, que, alm de ambiente favorvel fossilizao, ainda so compostos por populaes maiores em decorrncia do ambientes em que viveram ter sido mais homogneo. O Tempo Geolgico
A dificuldade normal das pessoas imaginarem a escala do tempo evolutivo um dos entraves para que aceitem que o processo que o
processo evolutivo possa ter ocorrido sem direo. A nossa escala de tempo est ligada ao nosso perodo de vida de algumas dezenas de anos e, portanto, j difcil imaginar que tempo o de 500 anos desde que os europeus chegaram s Amricas. Por isso, o tamanho da dificuldade para o entendimento do que so 500 mil anos, 1 milho de anos, 200 milhes de anos, 1 bilho de anos.
O cenrio da evoluo est no captulo 12. A gua, a atmosfera, composio de gases, temperatura, gravidade, presso atmosfrica, composio qumica dos seres vivos, alimentao e alimentos, a digesto, os sentidos e at a percepo de rudos, entre outros fatores do ambiente descritos pelo autor, mostram, segundo ele, que a vida est adaptada ao planeta Terra. O Cenrio da Evoluo
A vida se desenvolveu na terra em condies ambientais e climticas muito especiais, entre limites muito estreitos se comparados com o rosto do Universo. Quando se especula sobre a existncia de vida em outro planeta, devemos ter em mente que a vida, como a conhecemos, adaptada ao ambiente da ter. De uma maneira bem simplificada, pode-se afirmar que a evoluo e diversificao dos seres vivos sempre despendem, a cada momento, das modificaes e das condies geerais do ambiente terrestre. As peculiaridades ambientais de cada regio e as alteraes desses ambientes ao longo do tempo, desde a origem do planeta, so os principais responsveis pela enorme diversidade biolgica.
gua em estado lquido
Tanto quanto se saiba, a terra continua sendo o nico local do Universo onde existe gua em estado lquido. A gua fica em estado lquido entre as temperatura de 0C e 100C. Abaixo de 0C ela se solidifica e, acima de 100C, evapora (esses limites podem variar uma pouco, dependendo da presso atmosfrica). Como se sabe, a vida na terra originou-se na gua e nenhuma forma de vida sobrevive sem ela.
Atmosfera
A atmosfera, que a camada gasosa que envolveu a Terra, dissipa-se gradualmente medida que nos afastamos de superfcie da terra, tornando quase impossvel estabelecer sua espessura e seu limite exato. A estimativa de seu peso em torno de 5 bilhes de toneladas, aproximadamente um milionsimo do peso da terra. A densidade da atmosfera diminuem proporcionalmente em funo da altitude: a 6.000m de altura a metade da existente ao nvel do mar; a 95.000m um milionsimo da existente ao nvel do mar.
Atmosfera
Essa camada gasosa que envolve a Terra, dissipa-se gradualmente medida que nos afastamos da superfcie do planeta, sendo quase impossvel estabelecer sua espessura e seu limite exato. A estimativa de seu peso em torno de 5 bilhes de toneladas, um milionsimo do peso da Terra. Sua densidade diminui proporcionalmente em funo da altitude: a 6 mil metros de altura a metade da existente no nvel do mar, a 95 mil metros um milionsimo as existente no nvel do mar
Gravidade
Ela varia dependendo da altitude, pois o resultado da acelerao de um corpo em queda e diminui de maneira inversamente proporcional ao quadrado da distncia ao centro da Terra. Ao nvel do mar, prximo a linha do Equador o valor da acelerao da gravidade de, aproximadamente, 9,816 m/s2. Como a fora da gravidade em funo do tamanho do planeta, na superfcie lunar a gravidade 1/6 da gravidade da Terra.
Presso Atmosfrica
a fora gerada pela fora da gravidade que atua sobre a atmosfera, em decorrncia do seu peso e da sua densidade. O corpo de todo organismo vivo imerso na atmosfera sofre a presso de fora para dentro e s no esmagado porque h uma presso de dentro para fora que se iguala a ela. Assim, numa
presso atmosfrica muito menor para a qual est adaptado qualquer organismo explode em decorrncia de sua presso interna.
Composio qumica dos seres vivos
Os seres vivos so formados pelos elementos qumicos que estavam presentes na Terra ou que chegaram aqui com meteoritos. Todos os seres vivos tem a molcula da gua como substncia mais abundante em seus organismos correspondendo a 70% do corpo humano, 75% das bactrias, 95% da folha de alface.
Alimentao:
Os primeiros seres vivos eram heterotrficos e s bem mais tarde evoluram os autotrficos. - heterotrficos: so organismos incapazes de transformar energia do Sol em energia qumica. Alimentao de matria orgnica onde essa transformao j foi feita - autotrficos : so os organismos capazes de transformar a energia do Sol em energia qumica; maioria dos vegetais e de algumas bactrias
Alimentos
Existem 3 tipos: 1. Os plsticos ou estruturais so as protenas, tanto de origem vegetal ou animal 2. os energticos 3. os reguladores A digesto
Quando uma clula, ao invs de obter aminocidos do ambiente, ingere outro organismo, recebe protenas do outro organismo. Essas protenas precisam ser
decompostas para liberarem os aminocidos para que a clula possa produzir as prprias protenas. A quebra das protenas nos aminocidos que as constituem, o processo chamado digesto. Ele est presente em todos os organismos; a Maneira e os rgos responsveis por essa funo variam muito de um grupo para outro. Nos vertebrados, o aparelho digestivo (boca, estomago, intestino e rgos acessrios) realiza tal funo. Quando um indivduo ingere protenas, a funo da digesto quebr-las nos aminocidos que a compem para que os aminocidos possam ser reorganizados nas suas protenas de acordo com seu cdigo gentico. Quando um animal ingere protenas vegetais, aps a digesto ele ser capaz de sintetizar protenas animais. o que acontece com os animais herbvoros ou vegetarianos. No caso do Homem, quando come carne de frango, por exemplo, as protenas so quebradas e ele sintetiza protenas humanas, o que vale para qualquer protena ingerida de origem animal ou vegetal.
Percepo do Ambiente
Viso, tato, olfato, paladar e audio so os principais sentidos dos animais que se diferenciaram e se adaptaram em funo das caractersticas fsico/qumicas dos ambientes existentes na Terra
Os sentidos
atravs dos sentidos que o crebro informado sobre o mundo exterior ao corpo. Desde o desenvolvimento embrionrio o crebro recebe estmulos que do ao indivduo a noo do mundo exterior. A estimulao se acentua aps o nascimento, quando principalmente a viso, as variaes de temperatura, o olfato e o paladar tambm passam a atuar. O indivduo nasce sem informao externa e o crebro tende a aprender, atravs dos sentidos, sobre o ambiente que o cerca.
Olfato
tambm chamado de sentido qumico porque seus receptores nervosos so excitados por estimulantes qumicos volteis existentes no ar.
O paladar
Ligado a ingesto dos alimentos, o paladar tambm um sentido qumico porque os receptores gustativos localizados na lngua so excitados por substncias qumicos existentes nos alimentos e informa ao crebro o sabor do que est sendo ingerido. Os trs sabores bsicos so: doce, azedo, salgado, mas a combinao deles permite uma enorme gama de percepo de diferentes sabores. No crebro, o sabor do alimento se completa com a combinao dos estmulos do paladar e do olfato.
Percepo tctil
O tato informa ao crebro sobre o que est em contato com o corpo com a temperatura ambiente, as superfcies que esto sendo tocadas, a presso da gravidade, as dores. Ao contrrio dos sentidos anteriores, no existe um rgo especfico para o tato e seus terminais nervosos esto espalhados por todo o corpo, principalmente prximos epiderme. Pelas funes bsicas exercidas, est claro que importante para a sobrevivncia dos indivduos e tambm est ajustado s condies ambientais da Terra. Porm, sua importncia e sua funo podem variar de organismo para organismo.
A vida est adaptada ao planeta Terra
Pode-se deduzir que desde a origem da vida, todos os organismos e todas as populaes de organismos foram e esto sendo selecionados para sobreviverem nas condies ambientais da sua rea de ocorrncia. Toda a vida na Terra est adaptada s condies ambientais, atuais dessa fina camada da superfcie da Terra onde, na existncia de gua, tanto a composio qumica da atmosfera quanto a temperatura, a gravidade e a
presso atmosfrica variam dentro de uma escala muito pequena, se comparados com as variaes encontradas no universo. Concluso: a vida na forma que conhecemos no Planeta Terra est restrita a esse ambiente.
Aps essa concluso, nada mais justo do que mostrar no captulo 13 a distribuio dos seres vivos na Terra, assim como o deslocamento histrico dos continentes atuais e suas consequncias at a histria dos mamferos neste contexto.
A distribuio dos seres vivos na Terra
Biogeografia o estudo do padro de distribuio dos seres vivos na Terra. Para a maioria das pessoas, a primeira lembrana a distribuio geogrfica dos organismos terrestres, e dentre eles, a dos animais. Alm da vida em reas terrestres, existem as de rea marinha e, diante da enorme diversidade de vida dos oceanos, a presena de animais e plantas que evoluram em ambientes terrestres como os insetos, os anfbios, os rpteis e os mamferos so insignificantes. Todas as reas ocenicas do planeta se intercomunicam, de maneira que, ao contrrio dos animais terrestres, os organismos marinhos deveriam ter o poder de deslocamento para todas as regies da Terra. So poucas as espcies que se deslocam por todos os oceanos, pois, embora a distribuio das guas seja contnua, os continentes constituem barreiras fsicas que, para serem contornadas, exigem que os organismos enfrentem situaes ecolgicas muito adversas, especialmente de temperatura.
A vida em ambientes terrestres
O ambiente da Terra sofreu mudanas muito grandes desde sua origem, em decorrncia de influncias astronmicas e da dinmica da prpria Terra. Antes da origem da vida e, provavelmente por um tempo posterior considervel, a Terra apresentava uma atmosfera reducional, sem oxignio livre e essa condio s mudou aps a evoluo dos organismos fotossintetizadores, a cerca de 3,2 bilhes de anos, cujos resduos metablicos transformaram a atmosfera de reducional a oxidante. Nos continentes, como o produto de
comunidades de vegetao terrestre, os solos orgnicos surgiram no Siluriano, a cerca de 438 milhes de anos. Tambm a distribuio dos mares e das massas continentais mudou drasticamente durante o tempo geolgico por efeito da tectnica de placas.
Relao ancestral/descendente dos seres vivos
Quando se pensa nos seres vivos atuais, deve se lembrar que a atual distribuio geogrfica deles depende de razes histricas, como a posio, a forma e o isolamento entre os continentes, que foram alterados ao longo do tempo geolgico, alm do clima que j sofreu vrias e grandes mudanas desde a origem da Terra. Como a vida surgiu na gua, todas as diferentes formas de vida terrestre descendem e evoluram a partir de ancestrais aquticos e todas essas formas de vida tambm so monofilticas, portanto, todos os vertebrados terrestres atuais so descendentes da populao dos primeiros vertebrados que conseguiram sobreviver fora da gua como anfbios primitivos. Da mesma forma, todos os insetos atuais das primeiras populaes de artrpodos que sobreviveram fora da gua; assim, seguem-se para os demais organismos terrestres, animais ou vegetais. A afirmao de que os grupos so monofilticos significa que, pelo fato de os ambientes terrestres serem descontnuos, uma espcie que evolua num continente s estar presente noutro continente, se emigrar ou for levada de alguma forma para l. O fato de uma espcie, ou um grupo taxonmico, estar em dois continentes diferentes no decorrente do fato de terem surgido duas vezes, de forma independente, em cada um dos continentes.
Padres de Distribuio Geogrfica de Fauna e flora
Aos padres biolgicos de uma rea geogrfica, d-se o nome de Fauna, ou de flora, que significa um conjunto de linhagens evolutivas vivendo em uma determinada regio. A determinao de rea pode receber o nome de ecossistema, que uma paisagem determinada por caractersticas geoclimticas conhecidas. Existe tambm o termo bioma, que se refere a um ecossistema que tenha nome prprio. Conjuntos paisagsticos relativamente homogneos so considerados biomas. So fisionomias ocupando reas relativamente contnuas, regionais e so
unidades teis como foco de polticas conservacionistas. Podemos ter ecossistema de floresta pluvial tropical, como, por exemplo, os biomas: - da Floresta Atlntica, da Floresta Amaznica, da Floresta do Congo. Os domnios morfoclimticos so representaes de biomas. So unidades definidas pela superposio de variveis geogrficas, como vegetao, geomorfologia, hidrologia, paisagem. Para o Brasil foram definidos os domnios do Cerrado, da Caatinga, da Amaznia, da Mata Atlntica, entre outros. Em geral, o que determina a distribuio e os limites de um ecossistema, ou de um bioma, o clima e considerando-se todas as reas continentais, a ocorrncia dos biomas previsvel porque eles localizam-se por zonas de latitudes diferentes que se sucedem medida que nos afastamos da linha do Equador em direo aos plos. A principal causa dessas zonas, alm da temperatura, o padro de circulao dos ventos, os quais determinam, a grosso modo, de todo o planeta, onde chove, quando chove e o quanto chove. A ocorrncia de biomas em locais no previstos pela localizao geogrfica planetria pode ser definida pode ser devida a solos diferenciados por permeabilidade de gua; por deficincia de nutrientes vegetais; por padres anormais e locais de circulao dos ventos; por diferenas de altitude; por proximidade com os oceanos. No existem rplicas dos biomas, no existem replicas de fauna e flora. Porm, dada a proximidade geogrfica e aos isolamentos, a fauna terrestre apresenta alguns grandes padres de estrutura que foram observados pelos pesquisadores desde o inicio das grandes navegaes.
Endemismo e disperso
Quando uma espcie ocorre apenas em uma regio da terra diz-se que endmica daquela rea. Toda espcie inicialmente endmica no seu local de origem, mas, se seus indivduos conseguirem dispersar, podem passar a habitar outras reas, ampliando o territrio de disperso. Se essa disperso levar a espcie a ocupar mais de uma zona biogeogrfica ou mais de um continente, poder ser considerada como semicosmopolita e, ao passar a habitar a maioria dos continentes, atinge o estgio Maximo de disperso tornando-se cosmopolita. So pouca as espcies que se tornaram cosmopolitas por disperso ativa. Uma delas a espcie humana
A invaso das espcies exticas
A maioria das espcies cosmopolitas atuais sofreu disperso passiva, ou seja, foram levadas pelo homem para outros lugares. As consequncias dessas atividades para o equilbrio biolgico da natureza variam desde incuos at muito grandes, embora muitas vezes, os prejuzos tanto financeiros como ambientais sejam difceis de serem avaliados por se tratarem de eventos nicos. De acordo com a Unio Internacional para a Conservao da Natureza (IUCN), as espcies invasoras representam a segunda maior ameaa a biodiversidade em todo o planeta, s perdendo para os desmatamentos.
Deriva continental
Deriva continental (movimentao dos continentes, ou ainda, que as placas se movem) a teoria que diz que a Terra no esttica. Wegener observou que a costa da frica possua contorno que se encaixava na America do Sul. A partir da foram sendo acumuladas evidncias que os dois continentes eram ligados no passado. Uma delas foi descoberta de fosseis de animais da mesma espcie nos dois continentes, cuja nica explicao a unio dos mesmos, pois seria impossvel esses animais terem atravessados o Oceano Atlntico. Estudos sobre a formao geolgica mostraram tambm, uma incrvel coincidncia dos mesmos tipos de rochas exatamente nas mesmas regies da frica e da Amrica do Sul. Para estudar a deriva continental e a expanso dos fundos ocenicos surgiu por volta de 1960 uma linha de pesquisa chamada Tectnica de Placas.
Consequncia dos deslocamentos das Placas
A consequncia dos atritos entre as placas depende do tipo e da intensidade do contato. As mais comuns so os terremotos, a erupo de vulces e o dobramento da costa terrestre dando origem as montanhas. Ilhas e arquiplagos atuais, na maioria dos casos, foram ligados em alguma poca ao uma rea continental. Em termos biolgicos, os isolamentos mais famosos so arquiplagos de Galpagos e da Malsia, cujas faunas peculiares impressionaram Darwin e Wallace, respectivamente, a ponto de os dois terem simultaneamente a ideia de propor a evoluo dos seres vivos pela seleo natural. Outros isolamentos famosos so da Austrlia, Tasmnia e Nova
Zelndia, em cujos territrios no existiam mamferos placentrios atem a chegada da espcie humana a quarenta mil anos.
O arquiplago do Hawaii
nico arquiplago do Pacfico Norte tem uma historia geolgica diferente. Surgiu a partir de acmulos de lavas das erupes vulcnicas da placa do Pacfico e nunca foi conectado a qualquer rea continental.
Um captulo especial foi dedicado espcie humana, o captulo 14, e todo o seu processo de evoluo. Destaque para o detalhe de que diferentemente da maioria dos animais, que ao invadir um novo sistema se adapta biologicamente, a espcie humana tende a ajustar as diferenas do novo ambiente s suas necessidades e no precisa, na maioria dos casos, se adaptar a ele para sobreviver.
A origem da espcie Homo Sappiens
Sobre o viver: (...) viver muito perigoso (...) Joo Guimares Rosa (...) morrer fcil, viver que o difcil (...) Moacyr Felix
Existem atualmente duas hipteses predominantes: - a primeira (modelo para fora da frica) postula que a espcie surgiu primeiro da frica e, saindo dela, entre 100 mil e 200 mil anos atrs, substituiu todas as outras espcies de Homo existentes. Essa hiptese diz que todos os povos atuais so exclusivamente afrodescendentes. - a segunda, conhecida como modelo multirregional diz que a espcie evoluiu simultaneamente em diferentes partes do mundo, a partir de linhagens originais de Homo Erectus, assim, o povo atual da China descenderia das populaes
de Homo Erectus que viviam l; os Aborgenes da Austrlia da prpria frica, Europa, descenderiam de populaes de Homo Erectus daquela regio. Embora as duas hipteses ainda polarizem as discusses sobre a origem do homem, o acumulo de dados indica que, provavelmente, a histria uma fuso dessas duas hipteses.
O processo evolutivo
Como o processo evolutivo biocentrico e no antropocntrico, a espcie humana, surgiu por uma sequncia altamente improvvel de eventos e os fatores que atuaram para que ela acontecesse so os mesmos para todos os seres vivos, ou seja, mutao, seleo natural, deriva gentica, migrao, padres de cruzamento. O processo evolutivo prescinde tanto de contedo moral quanto imoral e no fornece nenhuma base filosfica para a esttica ou para a tica. Porm, como qualquer outro conhecimento, a Biologia Evolutiva pode servir causada liberdade e da dignidade humana, por nos ajudar a aliviar a fome e a doena e por nos ajudar a entender e apreciar tanto a unidade quanto a diversidade humana.
Na concluso, que o autor chama de ltimas Palavras, ele retoma alguns temas e discute outros que permeiam as pesquisas atuais. O avano da cincia, em particular da gentica, est neste captulo. Assim como as migraes recentes e suas consequncias, as presses culturais que afetam o desenvolvimento cultural, e por fim as misrias da humanidade, o tamanho populacional que o autor chama de ameaa sobrevivncia da humanidade, alm da possibilidade no remota da extino provocada por uma destruio nuclear, e as misrias da humanidade.
uma formiguinha atravessa em diagonal a pgina ainda em branco. Mas ele, aquela noite, no escreveu nada. Para qu? Se por ali j havia passado o frmito e o mistrio da vida ... Mario Quintana
15. TORTORA, G. J. Corpo humano: fundamentos de anatomia e fisiologia. 6. ed. Porto Alegre: Artmed, 2006. Organizao do corpo humano O corpo humano consiste em vrios nveis de organizao: qumico, celular, tecidual, orgnico, sistmico e do organismo. As clulas so as unidades estruturais e funcionais bsicas de um organismo. Os tecidos consistem em grupos de clulas de especialidade similar. Os rgos so estruturas compostas por dois ou mais tecidos diferentes, tem funes especficas e normalmente uma forma reconhecvel. Os sistemas consistem em associaes de rgos que tenham uma funo comum. O organismo humano uma coleo de sistemas integrados estrutural e funcionalmente. Os sistemas do corpo humano so o tegumentar, o esqueltico, o muscular, o nervoso, o endcrino, o cardiovascular (circulatrio), o linftico e o imunolgico, o respiratrio, o digestrio, o urinrio e o genital. Entre os processos vitais esto o metabolismo, a responsividade, o movimento, o crescimento, a diferenciao e a reproduo.
Introduo qumica Matria qualquer coisa que ocupe espao e tenha massa. constituda por unidades fundamentais denominadas elementos qumicos, carbono, hidrognio e nitrognio perfazem 96% do peso corporal. Os tomos consistem num ncleo que contm prtons e nutrons, e em eltrons que se movimentam em torno do ncleo em camadas eletrnicas. Uma molcula consiste em dois ou mais tomos quimicamente combinados. As substncias inorgnicas geralmente no possuem carbono e as orgnicas sempre o contm. cidos, bases e sais inorgnicos dissociam-se em ons quando em soluo aquosa. Numa escala de pH, o 7 neutro; abaixo, solues cidas; valores acima, solues alcalinas. Carboidratos so acares ou amidos (compostos orgnicos) e a fonte mais
comum de energia necessria para a manuteno da vida. Lipdios, como os triglicerdeos (gorduras), protegem, isolam e fornecem energia. As protenas so construdas a partir de aminocidos. O DNA uma dupla-hlice e a substncia qumica principal dos genes. O RNA principalmente responsvel pelas reaes de sntese proteicas. O ATP a principal molcula armazenadora de energia. As clulas A clula a unidade bsica, viva, funcional e estrutural do corpo. A membrana plasmtica circunda a clula e a isola. Substncias movem-se atravs de membranas plasmticas entre os fluidos intra e extracelular. Fagocitose a ingesto de partculas slidas por pseudpodos e pinocitose a ingesto de um lquido pela membrana plasmtica. O citosol a poro espessa e semifluida de citoplasma, entre a membrana plasmtica e o ncleo. O ncleo controla as atividades celulares e contm a informao gentica. O retculo endoplasmtico armazena molculas recm-sintetizadas, sintetiza e empacota molculas, desintoxica substncias qumicas e libera ons clcio envolvidos na contrao muscular. Os ribossomos so stios de sntese de protenas. O complexo de Golgi processa, separa e distribui as protenas membrana plasmtica, aos lisossomos e s vesculas secretoras. Os lisossomos so estruturas esfricas que contm enzimas digestivas. As mitocndrias so chamadas de "casa de fora" da clula, uma vez que so responsveis pela produo de ATP. Microfilamentos, microtbulos e filamentos intermedirios formam o citoesqueleto. Clios e flagelos so projees celulares utilizadas no movimento. O centrossomo importante na reproduo celular. Os centrolos tem papel na formao e na regenerao de flagelos e clios. A sntese proteicas utiliza a informao gentica codificada no DNA, que transcrita a um filamento de RNA mensageiro; neste momento, cada base nitrogenada (citosina (C), guanina (G), tmina (T), adenina (A) e uracila (U)) transcrita para sua correspondente: A para U, T para A, G para C e C para G. Alm do RNA mensageiro, o DNA tambm sintetiza o RNA ribossmico e o RNA transportador. O RNA mensageiro contm, ento, a sequncia de aminocidos de uma protena que ser sintetizada no ribossomo (composto por RNA ribossmico e protena). Enquanto isso, o RNA transportador carreia os aminocidos especficos at o RNA mensageiro em forma de antcdon (uma trinca de bases nitrogenadas): por exemplo, se a sequncia no RNA mensageiro DAG, o anticdon trazido pelo RNA transportador ser AUC. O ribossomo, ao se deslocar ao longo da fita de RNA mensageiro, agrega as bases trazidas pelo RNA transportador e, medida que os aminocidos so adicionados, vai se formando a molcula de protena. Esta ltima etapa conhecida por traduo.
Os tecidos
um conjunto de clulas semelhantes, e sua substncia intercelular especializada para determinada funo (epitelial, conjuntivo, muscular e nervoso). O tecido epitelial pode ser de revestimento (escamoso simples, cuboidal simples, colunar simples, estratificado escamoso, estratificado cuboidal, de transio ou colunar pseudoestratificado) ou glandular (uma glndula pode ser endcrina ou excrina). O tecido conjuntivo o tecido mais abundante no corpo; suas clulas so fibroblastos, macrfagos, plasmcitos e mastcitos. A substncia fundamental e as fibras constituem a matriz do tecido conjuntivo, que pode ser embrionrio (mesnquima que forma todos os outros tecidos conjuntivos) ou adulto maduro (frouxo, areolar, adiposo, reticular, denso, elstico, cartilaginoso, sseo e sanguneo). A tnica epitelial (mucosa, serosa ou cutnea) uma camada de epitlio sobre uma camada de tecido conjuntivo. A tnica sinovial reveste cavidades de articulaes e no contm epitlio. O tecido muscular especializado para contrao, o que permite movimento. O tecido nervoso composto de neurnios que geram e conduzem impulsos nervosos.
O sistema tegumentar As principais partes da pele so a epiderme externa e a derme interna. Os rgos acessrios da pele so plos, glndulas da pele (sebceas, sudorparas e ceruminosas) e unhas. O sistema esqueltico Consiste em todos os ossos unidos nas articulaes e em cartilagem entre as articulaes. Tem como funes sustentao, proteo, locomoo, armazenamento mineral, alojamento de tecido hemocitopotico e armazenamento de energia. Os ossos podem ser longos, curtos, planos, irregulares, suturais ou sesamoideos. As clulas so amplamente separadas, circundadas por grandes quantidades de matriz e podem ser osteoprogenitoras, osteoblastos, ostecitos e osteoclastos. O tecido sseo pode ser compacto (protege, suporta e resiste ao estresse) ou esponjoso (trabculas circundando muitos espaos cheios de medula ssea vermelha). Devido atividade do disco epifisrio, a difise de um osso aumenta em comprimento. O osso velho constantemente destrudo pelos osteoblastos, enquanto um osso novo construdo pelos osteoblastos. O crescimento normal depende de sais minerais (clcio, fsforo, magnsio), vitaminas (D, C, A) e
hormnios (de crescimento humano, sexuais, tireoideos, calcitonina e paratireoideo). Fazem parte do esqueleto axial o crnio, o osso hioide, os ossculos da audio, a coluna vertebral, o esterno e as costelas. As partes do esqueleto apendicular so cngulo peitoral (ombro), ossos dos membros superiores, cngulo plvico (cngulo do membro inferior) e ossos dos membros inferiores. Os ossos cranianos incluem frontal (I), parietal (2), temporal (2), occipital (I), esfenoide (I) e etmoide (l). Os faciais so nasal (2), maxilas (2), zigomtico (2), mandbula (I), lacrimal (2), palatino (2), concha nasal inferior (2) e vmer (I). Os da coluna vertebral so as vrtebras cervicais (7), torcicas (12), lombares (5), o sacro (5, fundidos) e o cccix (4, fundidos). O esqueleto torcico consiste no esterno, costelas (12 pares), cartilagens costais e vrtebras torcicas. Cada cngulo peitoral tem uma clavcula e uma escpula. Em cada membro superior tem mero, ulna, rdio, carpo, metacarpo e falanges. O cngulo plvico tem dois ossos do quadril. Cada membro inferior tem fmur, patela, tbia, fbula, ossos tarsais, ossos metatarsais e falanges. As articulaes So as regies de contato entre dois ou mais ossos. Podem ser fibrosas, cartilaginosas ou sinoviais. As funes podem ser de sinartrose (articulao imvel), anfiartrose (articulao levemente mvel) ou diartrose (articulao livremente mvel). Elas permitem movimentos de deslizamento, flexo, extenso, hiperextenso, rotao, abduo, aduo e circunduo. O sistema muscular O tecido muscular esqueltico est inserido nos ossos e, na maior parte das vezes, estriado e voluntrio. O cardaco forma a maior parte da parede do corao, estriado e involuntrio. O liso est localizado nas vsceras, estriado e involuntrio. O tecido muscular tem como funes o movimento do corpo, o movimento de substncias dentro do corpo, a estabilizao das posies do corpo e a produo de calor. Suas caractersticas so excitabilidade, contratilidade, extensibilidade e elasticidade. No tecido muscular esqueltico, a fscia, o epimsio, o perimsio e o endomsio so os componentes do tecido conjuntivo. Tendes e aponeuroses so suas extenses, alm das fibras musculares. Os nervos enviam impulsos e o sangue, nutrientes e oxignio. Cada fibra muscular contm miofibrilas - miofilamentos delgados e espessos compartimentalizados em sarcmeros; os delgados so compostos de actina, tropomiosina e troponina e os espessos, de miosina. A contrao ocorre quando os miofilamentos delgados de um sarcmero deslizam uns contra os outros, quando as cabeas de miosina tracionam os miofilamentos de actina. Um neurnio motor transmite um impulso nervoso a um msculo esqueltico, o que
estimula a contrao (unidade motora). As fibras musculares de uma unidade motora contraem-se em sua extenso total ou no se contraem (tudo-ou-nada). A fadiga muscular resulta da disponibilidade diminuda de oxignio, dos efeitos txicos do dixido de carbono e do acmulo de cido ltico durante o exerccio. Os msculos esquelticos produzem movimento tracionando os ossos. O tecido muscular cardaco contrai e relaxa rpida, contnua e ritmicamente; pode contrair-se sem uma estimulao extrnseca e pode permanecer contrado por mais tempo que o tecido muscular esqueltico. O tecido muscular liso possui fibras musculares que contem filamentos intermedirios. No msculo liso visceral, as fibras esto dispostas numa rede; a contrao ocorre numa onda. O msculo liso multiunitrio encontrado nos vasos sanguneos e nos olhos. O tecido nervoso Auxilia a controlar e integrar todos as atividades do corpo. Est dividido em central (encfalo e medula) e perifrico (nervos cranianos e espinhais), com componentes sensitivos e motores, e est dividido em somtico e autnomo. O somtico conduz impulsos dos receptores cutneos para a musculatura estriada, enquanto o autnomo, para a lisa, a cardaca e as glndulas. A neuroglia sustenta e protege os neurnios, que so constitudos de corpo celular, dendritos e axnio, e conduzem o impulso nervoso. Seu agrupamento forma nervos, tratos, gnglios, ncleos cornos e funculos. As fibras com bainha de mielina e as mais espessas conduzem os impulsos mais rapidamente. A juno entre dois neurnios ou entre ele e uma clula muscular ou glandular chama-se sinapse. O impulso s ocorre do axnio de um neurnio para outro neurnio. O sistema nervoso central e somtico A medula espinhal e o encfalo so protegidos pelo lquido crebro espinhal e por trs meninges: dura-mter, aracnoide e pia-mter. O encfalo ainda tem a proteo dos ossos cranianos, e a medula espinhal, do canal e ligamentos vertebrais. Ela se estende do occipital 2a vrtebra lombar. A substncia cinzenta na medula dividida em cornos e a branca, em funculos. Conduz as informaes sensitivas e motoras atravs dos tratos sensitivos (ascendentes) e motores (descendentes), respectivamente. Um arco reflexo a via mais curta que pode ser tomada por um impulso, de um receptor a um efetor. Seus componentes bsicos so um receptor, um neurnio sensitivo, um centro integrador, um neurnio motor e um efetor. Os nervos espinhais so 8 pares cervicais, 12 torcicos, 5 lombares, 5 sacrais e 1 coccgeo. Os principais plexos (redes de nervos) so o cervical, braquial, lombar e sacral. Os nervos T2 e T11 (intercostais ou torcicos) no formam plexos. O encfalo composto por
tronco (bulbo, ponte e mesencfalo), crebro (diencfalo (tlamo e hipotlamo) e telencfalo) e cerebelo. A interrupo do suprimento de oxignio pode enfraquecer, causar leses permanentes ou matar as clulas do encfalo. bulbo regula a frequncia cardaca, a respiratria, o dimetro dos vasos sanguneos, a deglutio, a tosse, o vmito, o espirro e o soluo. A ponte une as partes do encfalo e controla a respirao. O mesencfalo conecta a ponte ao diencfalo. O tlamo uma estao de retransmisso para os impulsos sensitivos ao crtex cerebral (dor, temperatura, tato, presso e vibrao). O hipotlamo controla a raiva, a temperatura, a ingesto de alimentos e os padres de conscincia e sono. O telencfalo dividido em lbulos (frontal, parietal, temporal e occipital). O hemisfrio esquerdo controla o lado direito do corpo e tem o centro da linguagem falada e escrita, habilidades numricas e cientficas e raciocnio; o direito controla o lado esquerdo, a sensibilidade artstica, a percepo espacial e a gerao de imagens mentais. O cerebelo coordena os msculos esquelticos, o tnus muscular e o equilbrio corporal. Sistema nervoso autnomo Regula as atividades vicerais, as musculaturas lisa e cardaca e as glndulas. controlado pelo crtex cerebral e pelo hipotlamo. Os neurnios motores do SNA liberam noradrenalina, alm de acetilcolina. Divide-se em simptico (gasto de energia) e parassimptico (restaurao e conservao de energia). Sensibilidade e sensao Aps a recepo de um estmulo, acontece a converso em impulso nervoso, a conduo at o crebro e a interpretao do mesmo. Os receptores podem ser mecanoceptores, termoceptores, nociceptores, fotoceptores e quimioceptores. As sensaes tteis, trmicas e de dor so 1 a sensibilidade cutnea. Os receptores do tato so os corpsculos de Merkel e de Meissner, os da presso so os corpsculos de Ruffini e de Valter-Pacini. Os da temperatura so as terminaes nervosas livres. Os receptores olfatrios s percebem substncias gasosas, hidrossolveis e lipossolveis. Os receptores gustatrios s reconhecem substncias em soluo na saliva. A formao da imagem na retina, onde esto os fotoceptores (cones e bastonetes), envolve a refrao da luz, a acomodao da lente (cristalino), a constrio da pupila, a convergncia e a formao da imagem invertida. As ondas sonoras chegam membrana do tmpano, vibram o martelo, a bigorna e o estribo, atingem a janela do vestbulo e, com a alterao da presso interna da cclea, gera-se o impulso nervoso. As mculas do utrculo e do sculo so os rgos dos sentidos do equilbrio esttico e as cristas ampulares, do equilbrio dinmico.
O sistema endcrino Glndulas endcrinas e tecidos endcrinos (presentes em alguns rgos) secretam hormnios na corrente sangunea. Os hormnios regulam o meio interno, o metabolismo, o equilbrio energtico, a contrao muscular, a secreo glandular, certas resposta imunolgicas, o crescimento, o desenvolvimento e a reproduo. A combinao entre o hormnio e o receptor ativa uma cadeia de eventos na clula-alvo, na qual os efeitos fisiolgicos do hormnio so expressos. Os receptores dos hormnios lipossolveis so no interior da clula, enquanto que os dos hidrossolveis, na membrana plasmtica. A adeno-hipfise produz os hormnios do crescimento (hGH), prolactina (PRL), estimulante de tireoide (TSH), folculo estimulante (FSH), luteinizante (LH), adrenocorticotrpico (ACTH) e melancito-estimulante (MSH), e controlada pelo hipotlamo, que tambm produz a ocitocina (OT) e o antidiurtico (ADH). A tireoide est localizada junto laringe e secreta a tiroxina (T4), a triiodotironina (T3) e a calcitonina (CT), tambm controlada pelo hipotlamo. As paratireoides, em contato com as tireoides, so autoreguladas e secretam o hormnio paratireoideo (PTH). As suprarenais esto acima dos rins e secretam mineralocorticoides (aldosterona), glicocorticoides (cortisol) e andrgenos, alm de adrenalina e noradrenalina. Os pncreas possuem as ilhotas de Langerhans, que secretam glucagon, insulina, somatostatina e polipeptdio pancretico. Os ovrios produzem estrgeno e progesterona, e os testculos, a testosterona, todos hormnios sexuais. A pineal secreta melatonina, e o timo, a timosina (FHT), o fatortmico (FT) e a timopoietina.
O sistema cardiovascular: sangue Transporta oxignio, dixido de carbono, nutrientes, resduos e hormnios; ajuda a regular o pH, a temperatura e o contedo hdrico das clulas. composto por 55% de plasma e 45% de elementos figurados, que so: eritrcitos (glbulos vermelhos), clulas anucleadas que possuem hemoglobina que transportam 02 e C02; leuccitos (glbulos brancos), que podem ser granulares (neutrfilos, eosinfilos e basfilos) ou agranulares (linfcitos e moncitos), combatem a inflamao e a infeco, alm de serem importantes nos processos alrgicos; e plaquetas, que trabalham no processo de coagulao. A hemstase envolve vasoconstrio, formao do tampo plaquetrio e coagulao do sangue (rede de fibrina), que requer vitamina K. No sistema ABO, a presena ou ausncia dos isoantgenos A e B determina o grupo sanguneo, e, no sistema Rh, indivduos que possuem nos seus eritrcitos o antgeno Rh so Rh+ e os que no o possuem, Rh".
O Sistema cardiovascular: corao Localiza-se no mediastino, recoberto pelo pericrdio; sua parede possui trs camadas (epicrdio, miocrdio e endocrdio) e dividido em 4 cmaras: duas superiores (trios) e duas inferiores (ventrculos). A circulao do sangue pelo corpo humano tem a seguinte sequncia: o trio direito recebe o sangue venoso (sem oxigenao) atravs das veias cavas superior (vem das partes do corpo acima do corao) e inferior (partes do corpo abaixo do corao) e seio coronrio (drena o sangue dos vasos da parede do corao). O sangue venoso passa para o ventrculo direito atravs da valva tricspide que tem a funo de evitar o refluxo sanguneo. O sangue venoso sai do corao pelo tronco pulmonar (artria pulmonar esquerda e direita), indo para os pulmes onde sofre a oxigenao, retorna ao trio esquerdo pelas quatro veias pulmonares (agora como sangue arterial), passa pela valva mitral (bicspide), que tem a mesma funo do tricspide, e vai para o ventrculo esquerdo. O sangue sai do corao pela aorta ascendente; desta, o sangue arterial passa para as artrias coronrias, ao arco da aorta e s partes torcica e abdominal da aorta descendente. Estes vasos sanguneos e suas ramificaes transportam o sangue oxigenado para todas as partes do corpo. Esse movimento do sangue controlado pela abertura e fechamento das valvas e contrao e relaxamento do miocrdio. O batimento cardaco consiste em uma sstole (contrao) e uma distole (relaxamento) das cmaras do corao. A frequncia cardaca mdia de 75 batimentos/minutos que controlada pelo bulbo cerebral e pode ser alterada por fatores internos (adrenalina, sdio, potssio e idade) e externas (temperatura, emoes e sexo).
O sistema cardiovascular: vasos sanguneos As artrias so elsticas e contrteis, transportam o sangue que sai do corao (veias pulmonares so exceo), passando-o para as arterolas, que possuem papel importante na regulao da presso arterial, e que, por sua vez, ramificam-se em capilares (vasos microscpicos onde acontecem as trocas entre o sangue e as clulas). Feitas as trocas, o sangue comea a voltar ao corao pelas vnulas, que se juntam formando as veias que possuem valvas para evitar o refluxo. O bulbo recebe os impulsos enviados plos barorreceptores e controla a presso sangunea. Epinefrina e norepinefrina aumentam a presso. A pressa arterial mdia de 120/80 mmHg (sistlica/diastlica). A circulao sistmica e a pulmonar so as duas maiores e mais importantes rotas. A sistmica inicia na aorta, passa por todo o corpo e retorna ao corao pelas veias cavas e seio coronrio. A pulmonar sai do ventrculo direito, vai ao pulmo e, atravs das veias pulmonares, traz sangue
arterial para o trio esquerdo. A circulao portal heptica passa pelo fgado para desintoxicao do sangue.
O sistema linftico, a resistncia inespecfica e a imunidade Os linfonodos, o bao e o timo so os rgos desse sistema que inclui ainda a linfa e os vasos linfticos. Ele drena o excesso de fluido dos espaos tissulares, devolve as protenas que escaparam do sistema circulatrio, transporta lipdeos e vitaminas e confere imunidade. O fluido intersticial que banha as clulas chamado de linfa quando est nos vasos e possui menos protena que o plasma. Os vasos possuem mais valvas que as veias. Os linfonodos esto ao longo dos vasos linfticos e filtram a linfa. O bao a maior massa de tecido linftico e produz linfcito B, e o timo, linfcitos T. Resistncia inespecfica uma ampla variedade de respostas corporais (vmito, secrees, substncias antimicrobianas, etc.) contra uma variedade de patgenos. A imunidade adquirida pode ser ativa ou passiva, natural ou artificial, a qual produz linfcitos ou anticorpo especfico (IgG, IgA, IgM, IgD e IgE). Imunidade celular a destruio de antgenos por linfcitos T e a humoral, por anticorpos que so produzidos por plasmcitos originados por linfcitos B. Os tipos de linfcitos T so auxiliares, citotxicos, de hipersensibilidade, supressores e de memria.
O sistema respiratrio O nariz, a faringe, a laringe, a traquia, os brnquios e os pulmes so rgos desse sistema. O nariz tem como funo o aquecimento, umedecimento e filtragem do ar, a olfao e a ressonncia para sons. A poro interna do nariz comunica-se com os seios paranasais e a parte nasal da faringe, atravs dos canos. A faringe um tubo muscular revestido por tnica mucosa que atua na digesto e respirao. A laringe a via area que conecta a faringe traquia, contm a cartilagem tireoide (pomo de Ado), a epiglote (impede a passagem de alimento), as pregas vocais (produzem sons) e as pregas vestibulares (prendem o ar na cavidade torcica). A traquia liga a laringe aos brnquios, estes ramificam a via area chegando aos bronquolos. Os pulmes (dois) so recobertos pela pleura; o direito tem trs lbulos e o esquerdo, dois (adaptao para acomodar o corao). A troca gasosa ocorre na membrana alveolocapilar. A ventilao pulmonar consiste em inspirao (contrao dos msculos intercostais e diafragma) e expirao (relaxamento dos mesmos). A troca gasosa, na respirao pulmonar, ocorre entre os alvolos e os capilares sanguneos, e a tecidual, entre os capilares e as clulas dos tecidos. Quase a
totalidade do 02 transportada pelos tomos de ferro da hemoglobina; a maioria do C02 transportada pelo plasma. O sistema digestrio O processo consiste em ingesto, mistura e movimento, digesto mecnica e qumica, absoro e defecao, para preparo do alimento para uso das clulas. composto por trato gastrintestinal, dentes, lngua, glndulas salivares, fgado, vescula biliar e pncreas. So trs os pares de glndulas salivares (partida, submandibular e sublingual). A saliva lubrifica o alimento e inicia a digesto qumica dos carboidratos (polissacardeos em dissacardeos). A lngua e os dentes iniciam a digesto mecnica. O esfago transporta o bolo alimentar at o estmago por peristalse. O estmago realiza digesto mecnica (ondas misturadoras) e qumica (pepsina - converte protenas em peptdeos). A secreo e a motilidade gstrica so reguladas por mecanismos nervosos e hormonais. Os pncreas derramam o suco pancretico no duodeno, que contm enzimas que digerem o amido (amilase pancretica), a protena (tripsina, quimiotripsina e carboxipeptidase), os triglicerdeos (lipase pancretica) e os cidos nucleicos (nucleases). O fgado produz a bile que armazenada na vescula biliar; a bile emulsifica os triglicerdeos. O intestino delgado dividido em duodeno, jejuno e leo; adaptado digesto e absoro (gua, eletrlitos e vitaminas). O suco entrico composto por enzimas intestinais (dissacardeos em monossacardeos): peptidase (completa a digesto das protenas), lipase pancretica (triglicerdeos em cidos graxos) e nuclease (cidos nucleicos em pentose e bases nitrogenadas). O intestino grosso divide-se em ceco, colos, reto e canal anal. H digesto mecnica e qumica (ao bacteriana). Continua a absoro e ocorre a formao das fezes. Nutrio e metabolismo Existem seis classes de nutrientes: carboidratos, lipdeos, protenas, minerais, vitaminas e gua. O centro da fome e o da saciedade esto localizados no hipotlamo. Uma dieta deve possuir, em calorias, 55% de carboidratos, 30% de gordura e 15% de protenas, aproximadamente. O metabolismo (todas as reaes qumicas do corpo) est dividido em catabolismo (reaes de decomposio) e anabolismo (reaes de sntese). O metabolismo dos carboidratos inicia-se pelo metabolismo da glicose que envolve a gliclise (respirao anaerbica), o ciclo de Krebs (ocorre na mitocndria) e a cadeia de transporte de eltrons (reaes de oxirreduo). O anabolismo da glicose (glicognese: formando glicognio) estimulada pela insulina. A glicogenlise converte glicognio em glicose, e a gliconeognese converte gordura e protenas em glicose. Os lipdeos podem produzir ATP armazenado no tecido adiposo, ou utilizados como molculas estruturais ou na sntese de molculas essenciais. O catabolismo dos lipdeos chamado de liplise, e o anabolismo, de lipognese. As protenas so catabolisadas e convertidas em aminocidos
que, quando entram nas clulas, so imediatamente utilizados na sntese proteica (anabolismo) nos ribossomos. O minerais podem ser macro ou microminerais. As vitaminas podem ser lipossolveis (A, D, E e K) ou hidrossolveis (B e C). Um Kcal igual energia necessria para aumentar em 1C, 1000g de gua. O calor resultante da oxidao dos alimentos conhecido como taxa metablica, que pode ser afetada por diversos fatores.
O sistema urinrio Regula a concentrao e o volume do sangue e excreta as impurezas, atravs dos rins, ureteres, bexiga e uretra. O nfron (corpsculo renal e tbulo renal tbulo contorcido proximal, ramo descendente da ala de Henle, ramo ascendente da ala de Henle e tbulo contorcido distai) a unidade funcional dos rins. O sangue entra nos rins atravs da artria renal e sai pela veia renal. A maioria das substncias do plasma filtrada pela cpsula gromerular; a reabsoro tubular retm substncias necessrias para o corpo, e a secreo tubular elimina na urina as substncias qumicas desnecessrias. A filtragem do sangue atravs de um dispositivo artificial chamada de hemodilise. Os ureteres transportam urina dos rins bexiga urinria. A bexiga armazena urina para mico. A uretra vai do soalho da bexiga ao exterior. A urina contm 95% de gua e 5% de solutos (uria, creatinina, cido rico, urobilinognio, sais e ons).
Equilbrio de fluidos, eletroltico e cido-bsico Aproximadamente dois teros do fluido corporal est localizado no interior das clulas (fluido intracelular); o restante encontrado no fluido intersticial, plasma, linfa, cerebroespinhal, lquido sinovial, fluidos dos olhos e das orelhas, filtrado gromerular e lquidos pleurais, do pericrdio e peritonial. A gua constitui aproximadamente 60% do peso corporal. A desidratao causa a sensao de sede e estimula a ingesto de gua. Os eletrlitos so substncias qumicas que se dissociam em ons nos fluidos corporais. O Na* o on extracelular mais abundante, e o K+, o intracelular mais abundante. O clcio o mineral mais abundante no corpo. O movimento dos fluidos entre os compartimentos intersticial e intracelular ocorre por meio da osmose. O pH normal dos fluidos extracelulares varia de 7,35 a 7,45. Os sistemas tampes evitam mudanas drsticas do pH.
Os sistemas genitais A reproduo produz novos indivduos de uma espcie e passa material gentico de uma gerao para outra. Fazem parte dos rgos genitais as gnadas, os duetos, as glndulas sexuais assessrias e as estruturas de sustentao. Os rgos genitais masculinos incluem testculos, dueto do epiddimo, dueto deferente, dueto ejaculatrio, uretra, vesculas seminais, prstata, glndula de Cowper e pnis. Os testculos alojam-se no escroto e produzem espermatozoides (gametas masculinos) nos tbulos seminferos. Os espermatozoides so haploides (23 cromossomos) e so constitudos de cabea, pea intermediria e cauda. Na puberdade, as clulas intersticiais so estimuladas a produzir testosterona e inicia-se a produo de espermatozoides (espermatognese). Eles so transportados para o epiddimo, que os amadurece e armazena. Atravs dos duetos ejaculatrios, chegam uretra, que atravessa a prstata, o diafragma urogenital e o pnis. As vesculas seminais secretam o smen (neutraliza a acidez da uretra masculina e vagina) que viabiliza os espermatozoides. A prstata secreta um fluido que fornece ATP aos espermatozoides e tambm anticoagulante do smen. A ereo do pnis (rgo masculino de copulao) se d pela expanso de seus seios sanguneos. Os rgos genitais femininos incluem ovrios (gnadas), tubas uterinas, tero, vagina e vulva. Os ovrios se localizam lateralmente ao tero, na cavidade plvica. Produzem e liberam um nico vulo haploide por vez, que transportado pela tuba uterina at o tero (com forma de pra invertida), onde se implanta o vulo ou se desenvolve o feto. Sua camada interna chama-se endomtrio. A vagina, canal de passagem do fluxo menstrual e receptculo do pnis no ato sexual, e a vulva so os rgos genitais externos femininos. O ciclo menstrual prepara o endomtrio para receber o vulo fertilizado. Durante a fase menstrual (1 ao 5 dia), o endomtrio descamado; na fase provulatria um grupo de folculos amadurece, mas apenas um se desenvolve, e o endomtrio reparado (ao dos estrgenos). A ovulao a liberao do ovcito do folculo desenvolvido na cavidade peritonial (ao do hormnio LH). Na ps-ovulao, o endomtrio espessado (ao dos estrgenos e progesterona), se no houver fertilizao, a baixa desses hormnios descama o endomtrio (menstruao).
Desenvolvimento e herana A gestao envolve fertilizao, implantao, desenvolvimento embrionrio e fetal e nascimento. A fertilizao a unio de material gentico de um ocito secundrio com o de um espermatozoide, formando o zigoto. Atravs de
clivagens forma-se a mrula, que se desenvolve para blastocisto. No desenvolvimento embrionrio, as camadas germinativas primordiais (ectoderma, mesoderma e endoderma) e as membranas embrionrias (saco vitelino, mnio, crion e alantoide) so formadas. No desenvolvimento fetal, os rgos originados das camadas germinativas primordiais crescem rapidamente. Os estrgenos, a progesterona e a hCG so os hormnios que mantm a gravidez. A gestao humana dura aproximadamente 38 semanas. No nascimento, h a dilatao do colo do tero, a expulso do feto atravs das contraes uterinas e a eliminao da placenta. O gentipo a composio gentica de um indivduo, e o fentipo o resultado das interaes do gentipo e do seu meio ambiente. Sntese elaborada por Alexandre Miranda
RESPONSVEIS PELA PUBLICAO SECRETARIA DE FORMAO Nilca F. Victorino Magda Souza de Jesus CEPES CENTRO DE ESTUDOS E PESQUISAS EDUCACIONAIS E SINDICAIS Maria Izabel A. Noronha Coordenadora SECRETARIA DE COMUNICAES Paulo Jos das Neves Roberto Guido ASSESSORIA DE COMUNICAES Rosana Incio DIAGRAMAO Carlos Roberto F. dos Santos Rosely Soares ASSESSORIA DE FORMAO Edson Roberto Nunes SECRETRIA Solange Cavalheiro Produo da Secretaria de Formao REVISO Profa. Sandra Andria Ferreira
Polticas Sociais: Marcos de Oliveira Soares; Secretria para Assuntos de Aposentados: Silvia Pereira; Secretrio Adjunto para Assuntos de Aposentados: Gilberto de Lima Silva; Secretria Geral de Organizao: Margarida Maria de Oliveira; Secretrio de Organizao para a Capital: Jos Wilson de Souza Maciel; Secretrio de Organizao para a Grande So Paulo: Douglas Martins Izzo; Secretrio de Organizao para o Interior: Ezio Expedito Ferreira Lima; Secretrio de Organizao para o Interior: Ederaldo Batista.
DIRETORIA ESTADUAL:
Ademar de Assis Camelo; Aladir Cristina Genovez Cano; Alberto Bruschi; Alex Buzeli Bonomo; Ana Lcia Santos Cugler; Ana Paula Pascarelli dos Santos; Anita Aparecida Rodrigues Marson; Antonio Carlos Amado Ferreira; Antonio Jovem de Jesus Filho; Ariovaldo de Camargo; Ary Neves da Silva; Benedito Jesus dos Santos Chagas; Carlos Alberto Rezende Lopes; Carlos Barbosa da Silva; Carlos Eduardo Vicente; Carmen Luiza Urquiza de Souza; Cilene Maria Obici; Deusdete Bispo da Silva; Dorival Aparecido da Silva; Edgard Fernandes Neto; Edith Sandes Salgado; Edna Penha Arajo; Eliane Gonalves da Costa; Elizeu Pedro Ribeiro; Emma Veiga Cepedano; Fernando Borges Correia Filho; Fludio Azevedo Limas; Floripes Ingracia Borioli Godinho; Geny Pires Gonalves Tiritilli; Gerson Jos Jrio Rodrigues; Gisele Cristina da Silva Lima; Idalina Lelis de Freitas Souza; Ins Paz; Janaina Rodrigues; Josafa Rehem Nascimento Vieira; Jose Luiz Moreno Prado Leite; Jos Reinaldo de Matos Leite; Josefa Gomes da Silva; Jovina Maria da Silva; Jucina Benedita dos Santos; Juvenal de Aguiar Penteado Neto; Leandro Alves Oliveira; Leovani Simes Cantazini; Lindomar Conceio da Costa Federighi; Luci Ferreira da Silva; Luiz Carlos de Sales Pinto; Luiz Carlos de Freitas; Luiz Cludio de Lima; Luzelena Feitosa Vieira; Maisa Bonifcio Lima; Mara Cristina de Almeida; Marcio de Oliveira; Marcos Luiz da Silva; Maria Jos Carvalho Cunha; Maria Lcia Ambrosio Orlandi; Maria Liduina Facundo Severo; Maria Sufaneide Rodrigues; Maria Teresinha de Sordi; Maria Valdinete Leite Nascimento; Mariana Coelho Rosa; Mauro da Silva Incio; Miguel Leme Ferreira; Miguel Noel Meirelles; Moacyr Amrico da Silva; Orivaldo Felcio; Ozani Martiniano de Souza; Paulo Alves Pereira; Paulo Roberto Chacon de Oliveira; Ricardo Augusto Botaro; Ricardo Marcolino Pinto; Rita de Cssia Cardoso; Rita Leite Diniz; Roberta Iara Maria Lima; Roberta Maria Teixeira Castro; Roberto Mendes; Roberto Polle; Ronaldi Torelli; Sandro Luiz Casarini; Sebastio Srgio Toledo Rodovalho; Sergio Martins da Cunha; Solange Aparecida Benedeti Penha; Sonia Aparecida Alves de Arruda; Stenio Matheus de Morais Lima; Suzi da Silva; Tatiana Silvrio Kapor; Telma Aparecida Andrade Victor; Teresinha de Jesus Sousa Martins; Tereza Cristina Moreira da Silva; Uilder Ccio de Freitas; Ulisses Gomes Oliveira Francisco; Vera Lcia Loureno; Vera Lcia Zirnberger; Wilson Augusto Fiza Frazo.
DIRETORIA DA APEOESP TRINIO 2008/2011 DIRETORIA EXECUTIVA:
Presidenta: Maria Izabel Azevedo Noronha; VicePresidente: Jos Geraldo Corra Jnior; Secretrio Geral: Fbio Santos de Moraes; Secretrio Geral Adjunto: Odimar Silva; Secretria de Finanas: Luiz Gonzaga Jos; Secretria Adjunta de Finanas: Suely Ftima de Oliveira; Secretrio de Administrao e Patrimnio: Silvio de Souza; Secretrio Adjunto de Administrao e Patrimnio: Fbio Santos Silva; Secretrio de Assuntos Educacionais e Culturais: Pedro Paulo Vieira de Carvalho; Secretrio Adjunto de Assuntos Educacionais e Culturais: Carlos Ramiro de Castro; Secretrio de Comunicaes: Paulo Jos das Neves; Secretrio Adjunto de Comunicaes: Roberto Guido; Secretria de Formao: Nilca Fleury Victorino; Secretria Adjunta de Formao: Magda Souza de Jesus; Secretrio de Legislao e Defesa dos Associados: Francisco de Assis Ferreira; Secretria Adjunta de Legislao e Defesa dos Associados: Zenaide Honrio; Secretrio de Poltica Sindical: Joo Luis Dias Zafalo; Secretria Adjunta de Poltica Sindical: Eliana Nunes dos Santos; Secretria de Polticas Sociais: Francisca Pereira da Rocha; Secretrio Adjunto de
You might also like
- MAPA - Material de Avaliação Prática Da Aprendizagem: DisciplinaDocument6 pagesMAPA - Material de Avaliação Prática Da Aprendizagem: Disciplinajose carlos felizNo ratings yet
- Anotações Biologia e Bioquímica Humana UnicesumarDocument9 pagesAnotações Biologia e Bioquímica Humana UnicesumarSthefany Do NascimentoNo ratings yet
- Bases Biológicas Do Comportamento Aula 1Document27 pagesBases Biológicas Do Comportamento Aula 1Tharick Matheus - Administrativo DFNo ratings yet
- Curso de Biologia Pré-Vestibular: Sumário dos Módulos de Citologia, Histologia, Seres Vivos, Fisiologia, Genética, Evolução e EcologiaDocument162 pagesCurso de Biologia Pré-Vestibular: Sumário dos Módulos de Citologia, Histologia, Seres Vivos, Fisiologia, Genética, Evolução e EcologiaJoaquim JosephNo ratings yet
- Células: estrutura, função e processosDocument8 pagesCélulas: estrutura, função e processosDenise CorreiaNo ratings yet
- Extração DNA Células VegetaisDocument10 pagesExtração DNA Células VegetaisPaulinhaa MesquitaNo ratings yet
- TópicosDocument45 pagesTópicosCamilaNo ratings yet
- 620 Questões de BiologiaDocument961 pages620 Questões de BiologiaPeter Parker75% (8)
- Trabalho de Introducão À Pesquisa Científica: Tema A Ciência Moderna, Seu Impacto Na Vida Das Pessoas em AngolaDocument12 pagesTrabalho de Introducão À Pesquisa Científica: Tema A Ciência Moderna, Seu Impacto Na Vida Das Pessoas em AngolacandelejeronimoNo ratings yet
- Material de Apoio de Biologia Do I Trimestre Da 11 Classe.Document20 pagesMaterial de Apoio de Biologia Do I Trimestre Da 11 Classe.Mateus Eduardo Dos SantosNo ratings yet
- Funções organelas célula vegetal animalDocument3 pagesFunções organelas célula vegetal animalThaís LuzNo ratings yet
- Ribossomos RER REL tradução proteínasDocument8 pagesRibossomos RER REL tradução proteínasAndre Luiz do NascimentoNo ratings yet
- Ensino técnico de nível médio integrado em biologia celularDocument13 pagesEnsino técnico de nível médio integrado em biologia celularJhorginho DanielNo ratings yet
- Respiração CelularDocument40 pagesRespiração CelularBruna Vitória Oliveira dos SantosNo ratings yet
- Trilha Biologia 1 Ano Semana 14Document7 pagesTrilha Biologia 1 Ano Semana 14Angelica Nunes TiepoNo ratings yet
- Carboidratos: da ingestão à produção de energiaDocument4 pagesCarboidratos: da ingestão à produção de energiaRonaldo ValmorbidaNo ratings yet
- A Base Celular Da VidaDocument14 pagesA Base Celular Da VidaCarolina RodriguesNo ratings yet
- Resumo da estrutura e função da célulaDocument13 pagesResumo da estrutura e função da célulaLuciana Taliano50% (2)
- Resumos de Química UniversidadeDocument31 pagesResumos de Química UniversidadeLeonor CabralNo ratings yet
- Características da célula procariótica e eucarióticaDocument24 pagesCaracterísticas da célula procariótica e eucarióticaIsabela Campos TavaresNo ratings yet
- AD1 BioGeral - Vinícius José de Oliveira-BICT-BCA 2023 1Document3 pagesAD1 BioGeral - Vinícius José de Oliveira-BICT-BCA 2023 1Vinícius JoséNo ratings yet
- Curso de pós-graduação em bioquímica aplicadaDocument59 pagesCurso de pós-graduação em bioquímica aplicadaAngela ReboloNo ratings yet
- Observação de células vegetais em divisãoDocument20 pagesObservação de células vegetais em divisãoMatilde MendesNo ratings yet
- Biologia Nejaii-Volume2Document21 pagesBiologia Nejaii-Volume2CLAUDIA CRISTINA TEIXEIRA DA SILVANo ratings yet
- Documento Sem NomeDocument17 pagesDocumento Sem NomeMatilde MendesNo ratings yet
- Trabalho BCMDocument5 pagesTrabalho BCMpedro.lealNo ratings yet
- Sistema Endomembranoso - Wikipédia, A Enciclopédia LivreDocument25 pagesSistema Endomembranoso - Wikipédia, A Enciclopédia LivrejobhunhajosiasmatucaNo ratings yet
- Resumo de Micro e Imuno, Prova 1Document15 pagesResumo de Micro e Imuno, Prova 1Ricardo GandurNo ratings yet
- Lipídeos: biomoléculas essenciais nas membranas celularesDocument14 pagesLipídeos: biomoléculas essenciais nas membranas celularesSGI Gestão IntegradoNo ratings yet
- AVALIacao 20220519021641Document4 pagesAVALIacao 20220519021641jogomortalmkNo ratings yet
- Ficha 8-BIOQDocument8 pagesFicha 8-BIOQcassimo abilio santosNo ratings yet
- Planos de Aulas Enem BiologiaDocument12 pagesPlanos de Aulas Enem BiologiahellberNo ratings yet
- Fundamentos da Bioquímica emDocument4 pagesFundamentos da Bioquímica emSamuel LemosNo ratings yet
- Biologia Molecular: Genoma, Cromossomos, Genes e Replicação do DNADocument8 pagesBiologia Molecular: Genoma, Cromossomos, Genes e Replicação do DNAALLYCE PEREIRA COELHONo ratings yet
- Ob Ten Cao de EnergiaDocument30 pagesOb Ten Cao de EnergiaAna Margarida Lopes Alves N3 8B a3487No ratings yet
- Atividade Contextualizada Celula Eucarionte e FunçõesDocument5 pagesAtividade Contextualizada Celula Eucarionte e FunçõesjosewillyNo ratings yet
- 1 Série Biologia TRILHA Semana 16Document2 pages1 Série Biologia TRILHA Semana 16Adriana BarbosaNo ratings yet
- Célula AnimalDocument6 pagesCélula AnimalFelipe MeloNo ratings yet
- DNA RNA proteínas ciclo celular replicaçãoDocument13 pagesDNA RNA proteínas ciclo celular replicaçãoVinícius Alves Moura67% (3)
- Capa ModeloDocument6 pagesCapa ModeloMaria Fernanda Lui LopesNo ratings yet
- Resumo - GenéticaDocument16 pagesResumo - Genéticaisabeljulia968No ratings yet
- Organelas celularesDocument10 pagesOrganelas celularesJoão LucasNo ratings yet
- Biologia 10o Ano - Organização CelularDocument47 pagesBiologia 10o Ano - Organização Celularcarinafish100% (2)
- PM3 - UFPR - Gabarito ComentadoDocument61 pagesPM3 - UFPR - Gabarito ComentadoKim TaehyungNo ratings yet
- Macromoléculas e constituição celularDocument8 pagesMacromoléculas e constituição celularIsac Francisco RaulNo ratings yet
- Apostila 06 - Estrutura e Função Das Principais BiomoléculasDocument7 pagesApostila 06 - Estrutura e Função Das Principais BiomoléculasNoberto Fontenele FrotaNo ratings yet
- As células: estrutura, funções e sistema de controle do ciclo celularDocument5 pagesAs células: estrutura, funções e sistema de controle do ciclo celularJennifer Isabella Rodrigues TeixeiraNo ratings yet
- Teoria Celular e EstruturasDocument11 pagesTeoria Celular e EstruturasAdriane CardozoNo ratings yet
- Bases químicas da constituição celularDocument55 pagesBases químicas da constituição celularDiego NascimentoNo ratings yet
- Revisão para EnemDocument16 pagesRevisão para EnemJedaNo ratings yet
- 9º Ano v3 - Ciencias PF - v29072021Document34 pages9º Ano v3 - Ciencias PF - v29072021LuisNo ratings yet
- QUESTÕES CITOLOGIADocument8 pagesQUESTÕES CITOLOGIAIvi Liliam Fernandes TabiNo ratings yet
- Práticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoFrom EverandPráticas de Genética, Biologia Molecular, Biotecnologia e EvoluçãoRating: 4 out of 5 stars4/5 (1)
- Trabalho ProntoDocument11 pagesTrabalho ProntoRamalde SantanaNo ratings yet
- Colégio Ciências NaturezaDocument5 pagesColégio Ciências NaturezaRamalde SantanaNo ratings yet
- Avaliação de Biologia com 16 questões sobre hereditariedadeDocument4 pagesAvaliação de Biologia com 16 questões sobre hereditariedadeRamalde SantanaNo ratings yet
- Simulado 1 Unidade - Educação Física - DilaDocument10 pagesSimulado 1 Unidade - Educação Física - DilaRamalde SantanaNo ratings yet
- O segredo reveladoDocument496 pagesO segredo reveladoeltonrluciano100% (1)
- Avaliao 1 Ano A Matutino II UnidadeDocument10 pagesAvaliao 1 Ano A Matutino II UnidadeRamalde SantanaNo ratings yet
- Dila - Cma 2015 - Educação Fisica 1º Ano B - VespertinoDocument7 pagesDila - Cma 2015 - Educação Fisica 1º Ano B - VespertinoRamalde SantanaNo ratings yet
- Fichamento de fontes acadêmicasDocument5 pagesFichamento de fontes acadêmicasDelatus Christian RockNo ratings yet
- 3 - Sociologia Da Educação - RODRIGUES, Alberto TosiDocument42 pages3 - Sociologia Da Educação - RODRIGUES, Alberto Tosiadanievski100% (13)
- Dila - Cma 2015 - Educação Fisica 2º Ano B - VespertinoDocument6 pagesDila - Cma 2015 - Educação Fisica 2º Ano B - VespertinoRamalde SantanaNo ratings yet
- Modelo de FichamentoDocument3 pagesModelo de FichamentoLeone SilvaNo ratings yet
- Literatura InfantilDocument13 pagesLiteratura InfantilRamalde SantanaNo ratings yet
- Dila - Cma 2015 - Educação Fisica 1º Ano A - MatutinoDocument6 pagesDila - Cma 2015 - Educação Fisica 1º Ano A - MatutinoRamalde SantanaNo ratings yet
- A Ilustração No Livro Infantil: Uma Análise Através Dos TemposDocument25 pagesA Ilustração No Livro Infantil: Uma Análise Através Dos TemposOlhares da FaE - UFMG100% (6)
- GraçaDocument7 pagesGraçaSimone TeodoroNo ratings yet
- A Ilustração No Livro Infantil: Uma Análise Através Dos TemposDocument25 pagesA Ilustração No Livro Infantil: Uma Análise Através Dos TemposOlhares da FaE - UFMG100% (6)
- Contrato de Prestação de Serviços - Cursos 24 HorasDocument5 pagesContrato de Prestação de Serviços - Cursos 24 HorasRamalde SantanaNo ratings yet
- Educação Fisica - 2º Ano A - Matutino 2014Document5 pagesEducação Fisica - 2º Ano A - Matutino 2014Ramalde SantanaNo ratings yet
- Montagem de Um Terrario para Caranguejeira 2Document1 pageMontagem de Um Terrario para Caranguejeira 2Aline KrupniskiNo ratings yet
- Educação Fisica - 3 º Ano A - Matutino 2014Document6 pagesEducação Fisica - 3 º Ano A - Matutino 2014Ramalde SantanaNo ratings yet
- Educação Fisica - 3 º Ano B - Vespertino 2014Document6 pagesEducação Fisica - 3 º Ano B - Vespertino 2014Ramalde SantanaNo ratings yet
- Educação Fisica - 2º Ano A - Matutino 2014Document5 pagesEducação Fisica - 2º Ano A - Matutino 2014Ramalde SantanaNo ratings yet
- Cma 2014 - Educação Fisica 4º Ano BDocument5 pagesCma 2014 - Educação Fisica 4º Ano BRamalde SantanaNo ratings yet
- Emsc 7 AnoDocument6 pagesEmsc 7 AnoRamalde SantanaNo ratings yet
- Educação Fisica - 1º Ano C - Vespertino-2014Document4 pagesEducação Fisica - 1º Ano C - Vespertino-2014Ramalde SantanaNo ratings yet
- Educação Fisica - 1º Ano C - Vespertino-2014Document4 pagesEducação Fisica - 1º Ano C - Vespertino-2014Ramalde SantanaNo ratings yet
- Educação Fisica - 3 º Ano B - Vespertino 2014Document6 pagesEducação Fisica - 3 º Ano B - Vespertino 2014Ramalde SantanaNo ratings yet
- Educação Fisica - 3 º Ano A - Matutino 2014Document6 pagesEducação Fisica - 3 º Ano A - Matutino 2014Ramalde SantanaNo ratings yet
- Portalpedagogico Biologia II Unidade 2014Document6 pagesPortalpedagogico Biologia II Unidade 2014Ramalde SantanaNo ratings yet
- Processamento AlternativoDocument2 pagesProcessamento AlternativoAna Rita BragaNo ratings yet
- A Biologia Da CrençaDocument135 pagesA Biologia Da CrençaMarcos Vinicius Lopes100% (1)
- Ciências Do Comportamento Desviante IDocument46 pagesCiências Do Comportamento Desviante Icarina nunesNo ratings yet
- Geovanna Abreu - Estudo Dirigido 1 - Estrutura DNA Replicação Trancrica Traducao - Rogerio UEFSDocument2 pagesGeovanna Abreu - Estudo Dirigido 1 - Estrutura DNA Replicação Trancrica Traducao - Rogerio UEFSJoaoNo ratings yet
- Biologia Molecular e suas aplicações no diagnósticoDocument21 pagesBiologia Molecular e suas aplicações no diagnósticoLuciana dos SantosNo ratings yet
- Ic - Dragon's Blood LPDocument11 pagesIc - Dragon's Blood LPEdersson NaitzNo ratings yet
- Guia Básico Nature, IVs e EVs em PokémonDocument11 pagesGuia Básico Nature, IVs e EVs em PokémonGilney Lias de MacêdoNo ratings yet
- Trindade - Caderno de Questões - Prova IDocument12 pagesTrindade - Caderno de Questões - Prova ILeandro BresegheloNo ratings yet
- Estrutura dos ácidos nucleicos DNA e RNADocument35 pagesEstrutura dos ácidos nucleicos DNA e RNAaudebaranNo ratings yet
- Cap 2 ORGANIZACAO MOLECULAR DOS GENOMAS - C Sol - AlunosDocument9 pagesCap 2 ORGANIZACAO MOLECULAR DOS GENOMAS - C Sol - AlunosEmília Morgado SantosNo ratings yet
- Lista de exercícios sobre estrutura e função dos ácidos nucleicosDocument6 pagesLista de exercícios sobre estrutura e função dos ácidos nucleicosAlessandro G. Mattos100% (1)
- Clique Aqui para Realizar A Atividade de Estudo 01 - Prazo Final - 18-06-2023 - Revisão Da TentativaDocument8 pagesClique Aqui para Realizar A Atividade de Estudo 01 - Prazo Final - 18-06-2023 - Revisão Da TentativaJesse Santana Costa100% (1)
- Wa0041.Document15 pagesWa0041.Shadow FunnyNo ratings yet
- Introdução A Biotecnologia AULA 1Document47 pagesIntrodução A Biotecnologia AULA 1Bila BernardesNo ratings yet
- Jogos Didáticos - BiologiaDocument37 pagesJogos Didáticos - BiologiaWilliam SousaNo ratings yet
- Transporte de seiva bruta no floema de plantasDocument13 pagesTransporte de seiva bruta no floema de plantasEfraim Nobre SoaresNo ratings yet
- Ácido Desoxirribonucleico - DNADocument20 pagesÁcido Desoxirribonucleico - DNACia. Castro (Orgânicos em Sua Vida)No ratings yet
- Livro Ciencias Biologicas GeneticaDocument134 pagesLivro Ciencias Biologicas GeneticajganascimentoNo ratings yet
- Genética 3º Ano - IntroduçãoDocument12 pagesGenética 3º Ano - IntroduçãoDenis Claudio Silva BarrosNo ratings yet
- SanarFlix - Introdução À GenéticaDocument31 pagesSanarFlix - Introdução À GenéticaGabriel Elias100% (1)
- Biotecnologia LIVRO WEBDocument306 pagesBiotecnologia LIVRO WEBAlexandreScheifeleNo ratings yet
- Soma Dos N Primeiros Termos de Uma PADocument31 pagesSoma Dos N Primeiros Termos de Uma PAkilt4No ratings yet
- Estrutura do DNA e processo de replicaçãoDocument24 pagesEstrutura do DNA e processo de replicaçãoRAUL SANTOS SILVANo ratings yet
- Aula 2 GenéticaDocument52 pagesAula 2 GenéticaRafael AlflenNo ratings yet
- Simulado - Biologia - GeneticaDocument2 pagesSimulado - Biologia - GeneticaOzimar FroesNo ratings yet
- Teste 2 PDocument11 pagesTeste 2 PRenata RosárioNo ratings yet
- 2o Simulado Mensal - Prova ComentadaDocument41 pages2o Simulado Mensal - Prova ComentadafisioNo ratings yet
- DNA e RNA: estrutura, função e replicaçãoDocument88 pagesDNA e RNA: estrutura, função e replicaçãoFrancisco Marques100% (5)
- Ficha 03-03-2023Document4 pagesFicha 03-03-2023Marcelo PereiraNo ratings yet
- Banca de Exercícios - Introdução Ao Estuda Da CélulaDocument7 pagesBanca de Exercícios - Introdução Ao Estuda Da CélulaEmanoel Tiago Barbosa CiriloNo ratings yet