You might also like
- Iluzie sau realitate? (Reflecții și fantezii despre misterul vieții și comunicarea temporală)From EverandIluzie sau realitate? (Reflecții și fantezii despre misterul vieții și comunicarea temporală)Rating: 3.5 out of 5 stars3.5/5 (2)
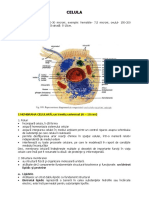
- CELULADocument29 pagesCELULAMadalina AndreeaNo ratings yet
- Membrana CitoplasmaticaDocument12 pagesMembrana CitoplasmaticaMafy20No ratings yet
- Modelul Mozaicului Fluid Al Membranei CelulareDocument12 pagesModelul Mozaicului Fluid Al Membranei CelulareBuliga Claudia50% (2)
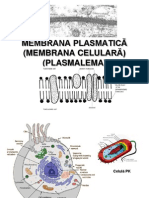
- Membrana CelularaDocument9 pagesMembrana CelularaCorina Bianca MironNo ratings yet
- Referat Membrana CelularaDocument15 pagesReferat Membrana CelularaRazvan_GabrielNo ratings yet
- Celula Eucariota Structura Si UltrastructuraDocument19 pagesCelula Eucariota Structura Si UltrastructuraFlorentina Claudia Calusaru100% (2)
- Structura Membranei CelulareDocument11 pagesStructura Membranei CelulareOana Palade100% (2)
- Medicamente HomeopateDocument10 pagesMedicamente HomeopateMarianSiGeorgianaMihaiNo ratings yet
- HANS Ghidul Doctorului AutismDocument38 pagesHANS Ghidul Doctorului AutismLarisa Ardelean100% (1)
- Mb. Celulara - ReferatDocument17 pagesMb. Celulara - ReferatClaudiu Bud100% (2)
- Membrane Biologice - BunDocument21 pagesMembrane Biologice - BunMarius Alexa67% (3)
- Celula Si SCH Prin MBDocument10 pagesCelula Si SCH Prin MBTeodora VasileNo ratings yet
- Membrana CelularaDocument25 pagesMembrana CelularaFlavius DobreNo ratings yet
- Elemente de Transport Prin Membrana CelularaDocument29 pagesElemente de Transport Prin Membrana CelularaIoana CojocaruNo ratings yet
- Membrana Celulara PDFDocument40 pagesMembrana Celulara PDFOzana CopyShop100% (1)
- Biologie Celulara Curs 2Document4 pagesBiologie Celulara Curs 2Voineta Gheorghiu100% (1)
- 4 Transport Membranar MGDocument19 pages4 Transport Membranar MGmarkopavloski100% (1)
- Biocel - Subiecte RezolvateDocument94 pagesBiocel - Subiecte RezolvateVictorVasile95% (20)
- Subiecte PT Exam Oral BiofarmacieDocument66 pagesSubiecte PT Exam Oral Biofarmaciebiancaaa24No ratings yet
- Subiect eDocument162 pagesSubiect eMarina MaziluNo ratings yet
- Membrana PlasmaticăDocument56 pagesMembrana PlasmaticăAndrada Raicu100% (1)
- Colica RenalăDocument7 pagesColica RenalăBfkt ProNo ratings yet
- Definiţia Şi Funcţiile Membranelor BiologiceDocument23 pagesDefiniţia Şi Funcţiile Membranelor BiologiceDanila ElenaNo ratings yet
- Fenomene de Transport MembranarDocument19 pagesFenomene de Transport MembranarChiriac SimonaNo ratings yet
- WWW - Referate.ro-Membrana Celulara Structura Moleculara Si Functiile Membranelor Celulare 81e12Document25 pagesWWW - Referate.ro-Membrana Celulara Structura Moleculara Si Functiile Membranelor Celulare 81e12Otilia Iftimie100% (1)
- Biologie Celulară Şi Moleculară Curs 4Document56 pagesBiologie Celulară Şi Moleculară Curs 4Andrei GlonțNo ratings yet
- Diversitatea Si Rolul Membranelor BiologiceDocument12 pagesDiversitatea Si Rolul Membranelor BiologiceRaluca RoxanaNo ratings yet
- Membrana Celulara Structura Moleculara Si Functiile Membranelor CelulareDocument25 pagesMembrana Celulara Structura Moleculara Si Functiile Membranelor CelulareAna CirsteaNo ratings yet
- Colesterolul MembranarDocument5 pagesColesterolul MembranarProf. Cristina GheraNo ratings yet
- Permeabilitate Şi TransportDocument14 pagesPermeabilitate Şi TransportLisa Briggs100% (1)
- Curs 9Document17 pagesCurs 9aleahim_aciroivNo ratings yet
- TAM4Document37 pagesTAM4Adriana Loredana SecrieriuNo ratings yet
- Biofizica 7Document34 pagesBiofizica 7Lile George100% (1)
- 1Document41 pages1Viorica IugaNo ratings yet
- Proteinele Si Lipidele MembranareDocument13 pagesProteinele Si Lipidele MembranareFlorentina Claudia CalusaruNo ratings yet
- Histo Partialu 1Document10 pagesHisto Partialu 1Evelina UngureanuNo ratings yet
- BiocelDocument26 pagesBiocelAdelin FrumusheluNo ratings yet
- Membrane Biologice 1Document10 pagesMembrane Biologice 1Anca NegreanNo ratings yet
- Capitole Speciale de Biochimie Medicala-Compozitia Si Arhitectura MembranelorDocument11 pagesCapitole Speciale de Biochimie Medicala-Compozitia Si Arhitectura MembranelorEmma esperanzaNo ratings yet
- Sinteza Histo IDocument38 pagesSinteza Histo IAdriana ElenaNo ratings yet
- CelulaDocument10 pagesCelulaValentina Gina MuaNo ratings yet
- Structura Şi Ultrastructura CeluleiDocument34 pagesStructura Şi Ultrastructura CeluleiNadeaNo ratings yet
- Membrana PlasmaticäDocument53 pagesMembrana PlasmaticäFantasyoflove VallyNo ratings yet
- Celula EucariotaDocument5 pagesCelula EucariotaStefan GhergheNo ratings yet
- Curs MembranaDocument17 pagesCurs MembranaMorgan RosewoodNo ratings yet
- Biofarm ADocument22 pagesBiofarm ASimona StefanescuNo ratings yet
- Catb 5Document3 pagesCatb 5Diana ValentinaNo ratings yet
- Curs 4 BIOFIZDocument72 pagesCurs 4 BIOFIZfloricapopovici168No ratings yet
- Invelisul CelularDocument7 pagesInvelisul CelularDumitrita100% (1)
- Cap 4Document32 pagesCap 4cnajesrnasiferfNo ratings yet
- Membrane 2019Document38 pagesMembrane 2019Urecheanu DorinNo ratings yet
- Ceasul Chinezesc Nu GresesteDocument3 pagesCeasul Chinezesc Nu GresesteBfkt ProNo ratings yet
- ZEFLEMEAUA de Camil PetrescuDocument2 pagesZEFLEMEAUA de Camil PetrescuBfkt ProNo ratings yet
- Diabetul InsipidDocument7 pagesDiabetul InsipidBfkt ProNo ratings yet
- Ghid Ong 2019Document11 pagesGhid Ong 2019IuliNo ratings yet
- AdhdDocument19 pagesAdhdRodica CristeaNo ratings yet
- Idei PoliticeDocument9 pagesIdei PoliticeBfkt ProNo ratings yet
- Uleiul CBD in AutismDocument4 pagesUleiul CBD in AutismBfkt ProNo ratings yet
- VitamineleDocument3 pagesVitamineleBfkt ProNo ratings yet
- 7 Simptome Care Apar Mai Întâi Pe MâiniDocument8 pages7 Simptome Care Apar Mai Întâi Pe MâiniBfkt ProNo ratings yet
- Ghid Utilizator Notebook HP ELITEBOOK Folio G1Document74 pagesGhid Utilizator Notebook HP ELITEBOOK Folio G1Bfkt ProNo ratings yet
- Diabetul InsipidDocument7 pagesDiabetul InsipidBfkt ProNo ratings yet
- Analize HematologiceDocument5 pagesAnalize HematologiceBfkt ProNo ratings yet
- Screening Ministerul SanatatiiTSADocument1 pageScreening Ministerul SanatatiiTSAzb789No ratings yet
- 12 Idei de Briose Pe Care Le Poti Servi La Cina in 30 de MinuteDocument13 pages12 Idei de Briose Pe Care Le Poti Servi La Cina in 30 de MinuteBfkt ProNo ratings yet
- Alimentele Bogate În Colesterol NU Cresc Riscul de AteroameDocument3 pagesAlimentele Bogate În Colesterol NU Cresc Riscul de AteroameBfkt ProNo ratings yet
- 10 Retete Deosebite Cu Broccoli Pentru Copiii MofturosiDocument11 pages10 Retete Deosebite Cu Broccoli Pentru Copiii MofturosiBfkt ProNo ratings yet
- 10 Deserturi Raw-VeganDocument8 pages10 Deserturi Raw-VeganBfkt ProNo ratings yet
- 15 Trucuri in Bucatarie Pe Care Nu Le StiaiDocument7 pages15 Trucuri in Bucatarie Pe Care Nu Le StiaiBfkt ProNo ratings yet
- 10 Sosuri Reci Pentru Salate, Paste, Orez FripturaDocument5 pages10 Sosuri Reci Pentru Salate, Paste, Orez FripturaBfkt ProNo ratings yet
- 10 Idei de Jocuri Pentru Copii Folosind o Cutie de CartonDocument13 pages10 Idei de Jocuri Pentru Copii Folosind o Cutie de CartonBfkt ProNo ratings yet
- 10 Idei de Retete Delicioase Cu DovleacDocument12 pages10 Idei de Retete Delicioase Cu DovleacBfkt ProNo ratings yet
- 10 Jocuri Cu Nasturi Pentru CopiiDocument11 pages10 Jocuri Cu Nasturi Pentru CopiiBfkt ProNo ratings yet
- Proprietați Mecanice, Termodinamice Si Electromagnetice Ale Unor Materiale Biodegradabile.Document25 pagesProprietați Mecanice, Termodinamice Si Electromagnetice Ale Unor Materiale Biodegradabile.Bfkt ProNo ratings yet
- 10 Tratamente Naturiste Pentru 10 Probleme de Sanatate Ale CopiluluiDocument5 pages10 Tratamente Naturiste Pentru 10 Probleme de Sanatate Ale CopiluluiBfkt ProNo ratings yet
- 10 Deserturi Raw-VeganDocument8 pages10 Deserturi Raw-VeganBfkt ProNo ratings yet
- Proprietați Mecanice, Termodinamice Si Electromagnetice Ale Unor Materiale Biodegradabile.Document25 pagesProprietați Mecanice, Termodinamice Si Electromagnetice Ale Unor Materiale Biodegradabile.Bfkt ProNo ratings yet
- Proprietați Mecanice, Termodinamice Si Electromagnetice Ale Unor Materiale Biodegradabile.Document25 pagesProprietați Mecanice, Termodinamice Si Electromagnetice Ale Unor Materiale Biodegradabile.Bfkt ProNo ratings yet