Professional Documents
Culture Documents
Biology
Uploaded by
İsmail AtamOriginal Description:
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Biology
Uploaded by
İsmail AtamCopyright:
Available Formats
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.
gr 2010/2011
Year:
Notes for biology
Human biology
1. Subject, place and significance of biology. Basic methods in biology.
Biology is defined as the study of life. The term life refers to all organisms(plants, animals, bacteria, fungi, and protists) inhabiting Earth and itsatmosphere. Biology is a natural science concerned with the study of life and living organisms, including their structure, function, growth, origin, evolution, distribution, and taxonomy. Five unifying principles that can be said to be the fundamental axioms of modern biology: 1) Cells are the basic unit of life 2) New species and inherited traits are the product of evolution 3) Genes are the basic unit of heredity 4) An organism regulates its internal environment to maintain a stable and constant condition 5) Living organisms consume and transform energy. Basic methods in biology. 1) Biochemistry examines the rudimentary chemistry of life. 2) Molecular biology studies the complex interactions of systems of biological molecules. 3) Cellular biology examines the basic building block of all life, the cell. 4) Physiology examines the physical and chemical functions of the tissues, organs, and organ systems of an organism. 5) Ecology examines how various organisms interact and associate with their environment. Cell theory: The cell theory, in its modern form, includes the following three principles: 1) All organisms are composed of one or more cells, and the life processes of metabolism and heredity occur within these cells. 2) Cells are the smallest living things, the basic units of organization of all organisms. 3) Cells arise only by division of a previously existing cell . Although life likely evolved spontaneously in the environment of the early earth, biologists have concluded that no additional cells are originating spontaneously at present. Rather, life on earth represents a continuous line of descent from those early cells.
Notes made by kozi1989 email:kozi1989@yahoo.gr
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Evolution A central organizing concept in biology is that life changes and develops through evolution and that all life forms known have a common origin . Charles Darwin established evolution theory by articulating its driving force, natural selection. Darwin theorized that species and breeds developed through the processes of natural selection as well as by artificial selection or selective breeding. Genetic drift was embraced as an additional mechanism of evolutionary development in the modern synthesis of the theory. Gene theory Biological form and function are created from and passed on to the next generation by genes, which are the primary units of inheritance. Physiological adaptation to an organism's environment cannot be coded into its genes and cannot be inherited by its offspring. Remarkably, widely different organisms, including bacteria, plants, animals, and fungi, all share the same basic machinery that copies and transcribes DNA into proteins. For example, bacteria with inserted human DNA will correctly yield the corresponding human protein. The total complement of genes in an organism or cell is known as its genome, which is stored on one or more chromosomes. A chromosome is a single, long DNA strand on which thousands of genes, depending on the organism, are encoded. When a gene is active, the DNA code is transcribed into an RNA copy of the gene's information. A ribosome then translates the RNA into a structural protein or catalytic protein. Homeostasis Homeostasis is the ability of an open system to regulate its internal environment to maintain a stable condition by means of multiple dynamic equilibrium adjustments controlled by interrelated regulation mechanisms. All living organisms, whether unicellular or multicellular, exhibit homeostasis. Homeostasis exists at the cellular level, for example cells maintain a stable internal acidity (pH); and at the level of the organism, for example warm-blooded animals maintain a constant internal body temperature. Homeostasis is a term that is also used in association with ecosystems, for example, the atmospheric concentration of carbon dioxide on Earth has been regulated by the concentration of plant life on Earth because plants remove more carbon dioxide from the atmosphere during the daylight hours than they emit to the atmosphere at night. Tissues and organs can also maintain homeostasis.
2. Nucleic acids
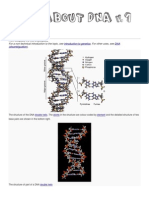
DNA- helix types and functions, RNA- types and functions Deoxyribonucleic acid , or DNA, is a nucleic acid that contains the genetic instructions used in the development and functioning of all known living organisms (with the exception of RNA viruses). The main role of DNA molecules is the long-term storage of information. DNA is often compared to a set of blueprints, like a recipe or a code, since it contains the instructions needed to construct other components of cells, such as proteins and RNA molecules. The DNA segments that carry this genetic information are called genes ,
Notes made by kozi1989 email:kozi1989@yahoo.gr
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
but other DNA sequences have structural purposes, or are involved in regulating the use of this genetic information. DNA consists of two long polymers of simple units called nucleotides, with backbones made of sugars and phosphate groups joined by ester bonds. These two strands run in opposite directions to each other and are therefore anti-parallel. Attached to each sugar is one of four types of molecules called bases. It is the sequence of these four bases along the backbone that encodes information. This information is read using the genetic code, which specifies the sequence of the amino acids within proteins. The code is read by copying stretches of DNA into the related nucleic acid RNA, in a process called transcription. Within cells, DNA is organized into long structures called chromosomes. These chromosomes are duplicated before cells divide, in a process called DNA replication . Eukaryotic organisms (animals, plants, fungi, and protists) store most of their DNA inside the cell nucleus and some of their DNA in organelles, such as mitochondria or chloroplasts. In contrast, prokaryotes (bacteria and archaea) store their DNA only in the cytoplasm . Within the chromosomes, chromatin proteins such as histones compact and organize DNA. These compact structures guide the interactions between DNA and other proteins, helping control which parts of the DNA are transcribed. The backbone of the DNA strand is made from alternating phosphate and sugar residues. The sugar in DNA is 2-deoxyribose, which is a pentose (five-carbon) sugar. The sugars are joined together by phosphate groups that form phosphodiester bonds between the third and fifth carbon atoms of adjacent sugar rings. These asymmetric bonds mean a strand of DNA has a direction. In a double helix the direction of the nucleotides in one strand is opposite to their direction in the other strand: the strands are antiparallel. The asymmetric ends of DNA strands are called the 5 (five prime) and 3 (three prime) ends, with the 5' end having a terminal phosphate group and the 3' end a terminal hydroxyl group. One major difference between DNA and RNA is the sugar, with the 2-deoxyribose in DNA being replaced by the alternative pentose sugar ribose in RNA. The DNA double helix is stabilized primarily by two forces : hydrogen bonds between nucleotides and base-stacking interactions among the aromatic bases. In the aqueous environment of the cell, the conjugated bonds of nucleotide bases align perpendicular to the axis of the DNA molecule, minimizing their interaction with the solvation shell and therefore, the Gibbs free energy. The four bases found in DNA are adenine (abbreviated A), cytosine (C), guanine (G) and thymine (T) . These four bases are attached to the sugar/phosphate to form the complete nucleotide, as shown for adenosine monophosphate. These bases are classified into two types; adenine and guanine are fused five- and six-membered heterocyclic compounds called purines, while cytosine and thymine are sixmembered rings called pyrimidines.A fifth pyrimidine base, called uracil (U), usually takes the place of thymine in RNA and differs from thymine by lacking a methyl group on its ring. Uracil is not usually found in DNA, occurring only as a breakdown product of cytosine.
Notes made by kozi1989 email:kozi1989@yahoo.gr
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
B-DNA B-DNA is the form commonly observed in chromosomes. B-DNA is a right-handed helix with 10 base pairs per turn. B-DNA is replicated and used in the transcription and translation of RNA, which is the molecule used for protein synthesis. B-DNA can be denatured, which means the hydrogen bonds are removed. This is essentially the first step in replicating DNA in the cell. A-DNA A-DNA is also a right-handed helix. However, there are more base pairs per turn. ADNA has 11 base pairs per turn. Other than the more compact structure, A-DNA is similar to B-DNA. It is biologically active in the cell, and it forms crystallized structures in lab experiments. Z-DNA Z-DNA is the type of DNA that is a left-handed helix. It is also known to be biologically active in zigzag formations of repeating base pair sequences. Z-DNA has 12 base pairs per turn, so it carries the most genes between each turn. Z-DNA plays a role in RNA transcription, which is the protein synthesis process of creating mRNA from a strand of DNA. mRNA (message RNA) is the molecule that carries transcribed genes to ribosomes where proteins are synthesized. cDNA cDNA (complementary or clonal DNA) is a type of DNA used to describe libraries of genetic information. cDNA is used in testing for pharmaceuticals and research of diseases. cDNA is a complementary strand that is transcribed in laboratories to create genes. Genetic engineering also uses these DNA libraries to create modified versions of genomic information. Ribonucleic acid (RNA) is one of the three major macromolecules (along with DNA and proteins) that are essential for all known forms of life. Like DNA, RNA is made up of a long chain of components called nucleotides . Each nucleotide consists of a nucleobase (sometimes called a nitrogenous base), a ribose sugar, and a phosphate group. The sequence of nucleotides allows RNA to encode genetic information. Like proteins, some RNA molecules play an active role in cells by catalyzing biological reactions, controlling gene expression, or sensing and communicating responses to cellular signals. One of these active processes is protein synthesis, a universal function whereby mRNA molecules direct the assembly of proteins on ribosomes. This process uses transfer RNA (tRNA) molecules to deliver amino acids to the ribosome, where ribosomal RNA (rRNA) links amino acids together to form proteins. The chemical structure of RNA is very similar to that of DNA, with two differences: 1) RNA contains the sugar ribose while DNA contains the slightly different sugar deoxyribose (a type of ribose that lacks one oxygen atom) 2) RNA has the nucleobase uracil while DNA contains thymine (uracil and thymine have similar base-pairing properties).
Notes made by kozi1989 email:kozi1989@yahoo.gr
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Unlike DNA, most RNA molecules are single-stranded. Single-stranded RNA molecules adopt very complex three-dimensional structures, since they are not restricted to the repetitive double-helical form of double-stranded DNA. RNA is made within living cells by RNA polymerases, enzymes that act to copy a DNA or RNA template into a new RNA strand through processes known as transcription or RNA replication, respectively. Types of RNA Messenger RNA (mRNA) carries information about a protein sequence to the ribosomes, the protein synthesis factories in the cell. It is coded so that every three nucleotides (a codon) correspond to one amino acid. In eukaryotic cells, once precursor mRNA (pre-mRNA) has been transcribed from DNA, it is processed to mature mRNA. This removes its intronsnon-coding sections of the pre-mRNA. The mRNA is then exported from the nucleus to the cytoplasm, where it is bound to ribosomes and translated into its corresponding protein form with the help of tRNA. In prokaryotic cells, which do not have nucleus and cytoplasm compartments, mRNA can bind to ribosomes while it is being transcribed from DNA. After a certain amount of time the message degrades into its component nucleotides with the assistance of ribonucleases. Transfer RNA (tRNA) is a small RNA chain of about 80 nucleotides that transfers a specific amino acid to a growing polypeptide chain at the ribosomal site of protein synthesis during translation. It has sites for amino acid attachment and an anticodon region for codon recognition that binds to a specific sequence on the messenger RNA chain through hydrogen bonding. Ribosomal RNA (rRNA) is the catalytic component of the ribosomes. Eukaryotic ribosomes contain four different rRNA molecules: 18S, 5.8S, 28S and 5S rRNA. Three of the rRNA molecules are synthesized in the nucleolus, and one is synthesized elsewhere. In the cytoplasm, ribosomal RNA and protein combine to form a nucleoprotein called a ribosome. The ribosome binds mRNA and carries out protein synthesis. Several ribosomes may be attached to a single mRNA at any time.rRNA is extremely abundant and makes up 80% of the 10 mg/ml RNA found in a typical eukaryotic cytoplasm.*Regulatory rnas
3. Self-assembly of the macromolecular subunits
A macromolecule is a very large molecule commonly created by some form of polymerization. In biochemistry, the term is applied to the four conventional biopolymers (nucleic acids, proteins, carbohydrates, and lipids), as well as non-polymeric molecules with large molecular mass such as macrocycles. The constituent molecules from which macromolecules are assembled are called monomers (mono=single, meros=part). Self-assembly is a term used to describe processes in which a disordered system of pre-existing components forms an organized structure or pattern as a consequence of specific, local interactions among the components themselves, without external direction. When the constitutive components are molecules, the process is termed molecular selfassembly. Self-assembly can be classified as either static or dynamic. In static self-assembly,
Notes made by kozi1989 email:kozi1989@yahoo.gr
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
the ordered state forms as a system approaches equilibrium, reducing its free energy. However in dynamic self-assembly, patterns of pre-existing components organized by specific local interactions are not commonly described as "self-assembled" by scientists in the associated disciplines. These structures are better described as "self-organized". Nucleic acids Nucleic acids are biological molecules essential for life, and include DNA (deoxyribonucleic acid) and RNA (ribonucleic acid). Together with proteins, nucleic acids make up the most important macromolecules; each is found in abundance in all living things, where they function in encoding, transmitting and expressing genetic information. The term nucleic acid is the over all name for DNA and RNA, members of a family of biopolymers, and is synonymous with polynucleotide. Nucleic acids were named for their initial discovery within the cell nucleus, and for the presence of phosphate groups (related to phosphoric acid). All living cells and organelles contain both DNA and RNA, while viruses contain either DNA or RNA, but not usually both. The basic component of biological nucleic acids is the nucleotide , each of which contains a pentose sugar (ribose or deoxyribose), a phosphate group, and a nucleobase. Nucleic acids can vary in size, but are generally very large molecules. Indeed, DNA molecules are probably the largest individual molecules known. In most cases, naturally occurring DNA molecules are double-stranded and RNA molecules are single-stranded. There are numerous exceptions, howeversome viruses have genomes made of double-stranded RNA and other viruses have single-stranded DNA genomes, and, in some circumstances, nucleic acid structures with three or four strands can form. Nucleic acids are linear polymers (chains) of nucleotides. Each nucleotide consists of three components: a purine or pyrimidine nucleobase (sometimes termed nitrogenous base or simply base), a pentose sugar and a phosphate group. The substructure consisting of a nucleobase plus sugar is termed a nucleoside. Nucleic acid types differ in the structure of the sugar in their nucleotides - DNA contains 2'-deoxyribose while RNA contains ribose (where the only difference is the presence of a hydroxyl group). Also, the nucleobases found in the two nucleic acid types are different: adenine, cytosine, and guanine are found in both RNA and DNA, while thymine occurs in DNA and uracil occurs in RNA. The sugars and phosphates in nucleic acids are connected to each other in an alternating chain (sugar-phosphate backbone) through phosphodiester linkages. In conventional nomenclature, the carbons to which the phosphate groups attach are the 3'end and the 5'-end carbons of the sugar. This gives nucleic acids directionality, and the ends of nucleic acid molecules are referred to as 5'-end and 3'-end. The nucleobases are joined to the sugars via an N-glycosidic linkage involving a nucleobase ring nitrogen (N-1 for pyrimidines and N-9 for purines) and the 1' carbon of the pentose sugar ring. Carbohydrates A carbohydrate is an organic compound with the empirical formula Cm(H2O)n (where m could be different from n); that is, consists only of carbon, hydrogen, and
Notes made by kozi1989 email:kozi1989@yahoo.gr
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
oxygen, with a hydrogen:oxygen atom ratio of 2:1 (as in water). Carbohydrates can be viewed as hydrates of carbon, hence their name.The carbohydrates (saccharides) are divided into four chemical groupings: monosaccharides, disaccharides, oligosaccharides, and polysaccharides. In general, the monosaccharides and disaccharides, which are smaller (lower molecular weight) carbohydrates, are commonly referred to as sugars. The word saccharide comes from the Greek word (skkharon), meaning "sugar". Carbohydrates perform numerous roles in living things. Polysaccharides serve for the storage of energy (e.g., starch and glycogen), and as structural components (e.g., cellulose in plants and chitin in arthropods). The 5-carbon monosaccharide ribose is an important component of coenzymes (e.g., ATP, FAD, and NAD) and the backbone of the genetic molecule known as RNA. The related deoxyribose is a component of DNA. Saccharides and their derivatives include many other important biomolecules that play key roles in the immune system, fertilization, preventing pathogenesis, blood clotting, and development. Lipids Lipids fall into three main categories: fats, steroids, and phospholipids. Lipids are a broad group of naturally occurring molecules which includes fats, waxes, sterols, fat-soluble vitamins (such as vitamins A, D, E and K), monoglycerides, diglycerides, phospholipids, and others. The main biological functions of lipids include energy storage, as structural components of cell membranes, and as important signaling molecules. Lipids may be broadly defined as hydrophobic or amphiphilic small molecules; the amphiphilic nature of some lipids allows them to form structures such as vesicles, liposomes, or membranes in an aqueous environment. Biological lipids originate entirely or in part from two distinct types of biochemical subunits or "building blocks": ketoacyl and isoprene groups. Using this approach, lipids may be divided into eight categories: fatty acyls, glycerolipids, glycerophospholipids, sphingolipids, saccharolipids and polyketides (derived from condensation of ketoacyl subunits); and sterol lipids and prenol lipids (derived from condensation of isoprene subunits). Although the term lipid is sometimes used as a synonym for fats, fats are a subgroup of lipids called triglycerides. Lipids also encompass molecules such as fatty acids and their derivatives (including tri-, di-, and monoglycerides and phospholipids), as well as other sterol-containing metabolites such as cholesterol. Although humans and other mammals use various biosynthetic pathways to both break down and synthesize lipids, some essential lipids cannot be made this way and must be obtained from the diet.
4. Comperative structural and functional characterization of procaryotic and
eucaryotic cells Procaryotic and eucaryotic cells. Origin and evolution of cell. The First Cell arose in the previously pre-biotic world with the coming together of several entities that gave a single vesicle the unique chance to carry out three essential and quite different life processes. These were:
Notes made by kozi1989 email:kozi1989@yahoo.gr
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
1) To copy informational macromolecules, 2) To carry out specific catalytic functions, and 3) To couple energy from the environment into usable chemical forms. These would foster subsequent cellular evolution and metabolism. Each of these three essential processes probably originated and was lost many times prior to The First Cell, but only when these three occurred together was life jump-started and Darwinian evolution of organisms began. The prokaryotes are a group of organisms that lack a cell nucleus (= karyon), or any other membrane-bound organelles. The organisms that have a cell nucleus are called eukaryotes. Most prokaryotes are unicellular, but a few such as myxobacteria have multicellular stages in their life cycles. The word prokaryote comes from the Greek (pro-) "before" + (karyon) "nut or kernel". Prokaryotes do not have a nucleus, mitochondria, or any other membrane-bound organelles. In other words, neither their DNA nor any of their other sites of metabolic activity are collected together in a discrete membrane-enclosed area. Instead, everything is openly accessible within the cell, some of which is free-floating. It is generally agreed that prokaryotes as a group do not have taxonomic value. They comprehend two domains: the bacteria and the archaea. A eukaryote is an organism whose cells contain complex structures enclosed within membranes. The defining membrane-bound structure that sets eukaryotic cells apart from prokaryotic cells is the nucleus, or nuclear envelope, within which the genetic material is carried. The presence of a nucleus gives eukaryotes their name, which comes from the Greek (eu, "good") and (karyon, "nut" or "kernel"). Most eukaryotic cells also contain other membrane-bound organelles such as mitochondria, chloroplasts and the Golgi apparatus. All species of large complex organisms are eukaryotes, including animals, plants and fungi, although most species of eukaryotic protists are microorganisms. Cell division in eukaryotes is different from that in organisms without a nucleus (prokaryotes). It involves separating the duplicated chromosomes, through movements directed by microtubules. There are two types of division processes. In mitosis, one cell divides to produce two genetically identical cells. In meiosis, which is required in sexual reproduction, one diploid cell (having two instances of each chromosome, one from each parent) undergoes recombination of each pair of parental chromosomes, and then two stages of cell division, resulting in four haploid cells (gametes). Each gamete has just one complement of chromosomes, each a unique mix of the corresponding pair of parental chromosomes.Eukaryotes appear to be monophyletic, and so make up one of the three domains of life. Main structures of eukaryotic cell: Plasma membrane: have lipid bilayers with embedded proteins, and fuctions in regulating what passes into and out of cell. Cytoskeleton: gives structural support and helps in movement; made up mostly of networks of protein filaments.
Notes made by kozi1989 email:kozi1989@yahoo.gr
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Nucleus: control center of cell, directs protein synthesis and cell reproduction is sphericall shaped; surrounded by a double membrane that contains chromosomes. Golgi apparatus: packs proteins for export from cell; forms secretory vesicles; made up of stacks of flat vesicles. Lysosomes: digests worn out organells and cell debris; plays role in cell death. The vescicles derived from golgi that contain hydrolytic digestive enzyme. Mitochondria: power house of cell; site of oxidative metabolism; made up of double membrane Chloroplast: site of photosynthesis in plant cells; membrane contains chlorophyll and photosynthetic pigment Chromosomes: contains hereditary information; made up of long threads of DNA that form a complex with protein. Nucleolus: assembles ribosomes; site of gene rRNA synthesis. Ribosome: site of protein synthesis; often bound to ER. Differences: A distinction between prokaryotes and eukaryotes (meaning true kernel, also spelled "eucaryotes") is that Eukaryotes do have "true" nuclei containing their DNA. Unlike prokaryotes, eukaryotic organisms may be unicellular, as in amoebae, or multicellular, as in plants and animals. The difference between the structure of prokaryotes and eukaryotes is so great that it is sometimes considered to be the most important distinction among groups of organisms. The cell structure of prokaryotes differs greatly from that of eukaryotes. The defining characteristic is the Absence of a nucleus. Also the size of Ribosomes in prokaryotes is smaller than that in eukaryotes , but two organelles found in eukaryotic cells contain mitochondria similar in size and makeup to those found in prokaryotes. The genomes of prokaryotes are held within an irregular DNA/protein complex in the cytosol called the nucleoid, which lacks a nuclear envelope. In general,prokaryotes lack the following membrane-bound cell compartments: mitochondria and chloroplasts. Instead, processes such as oxidative phosphorylation and photosynthesis take place across the prokaryotic plasma membrane. However, prokaryotes do possess some internal structures, such as cytoskeletons,and the bacterial order Planctomycetes have a membrane around their nucleoid and contain other membrane-bound cellular structures. Both eukaryotes and prokaryotes contain large RNA/protein structures called ribosomes, which produce protein. Prokaryotes are usually much smaller than eukaryotic cells.
Notes made by kozi1989 email:kozi1989@yahoo.gr
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Prokaryotes also differ from eukaryotes in that they contain only a single loop of stable chromosomal DNA stored in an area named the nucleoid, whereas eukaryote DNA is found on tightly bound and organized chromosomes. Although some eukaryotes have satellite DNA structures called plasmids, in general these are regarded as a prokaryote feature, and many important genes in prokaryotes are stored on plasmids. Prokaryotes have a larger surface-area-to-volume ratio giving them a higher metabolic rate, a higher growth rate, and, as a consequence, a shorter generation time compared to Eukaryotes.
5. The cell as an open biological system. Cell signaling. Cell junctions. Cell
receptors. Cells communicate through any of four basic mechanisms, depending primarily on the distance between the signaling and responding cells. In addition to using these four basic mechanisms, some cells actually send signals to themselves, secreting signals that bind to specific receptors on their own plasma membranes. This process, called autocrine signaling, is thought to play an important role in reinforcing developmental changes. Direct Contact: the surface of a eukaryotic cell is a thicket of proteins, carbohydrates, and lipids attached to and extending outward from the plasma membrane. When cells are very close to one another, some of the molecules on the cells plasma membranes may bind together in specific ways. Many of the important interactions between cells in early development occur by means of direct contact between cell surfaces. Paracrine Signaling Signal molecules released by cells can diffuse through the extracellular fluid to other cells. If those molecules are taken up by neighboring cells, destroyed by extracellular enzymes, or quickly removed from the extracellular fluid in some other way, their influence is restricted to cells in the immediate vicinity of the releasing cell. Signals with such short-lived, local effects are called paracrine signals. Like direct contact, paracrine signaling plays an important role in early development, coordinating the activities of clusters of neighboring cells. Endocrine Signaling If a released signal molecule remains in the extracellular fluid, it may enter the organisms circulatory system and travel widely throughout the body. These longer lived signal molecules, which may affect cells very distant from the releasing cell, are called hormones, and this type of intercellular com-munication is known as endocrine signaling. Synaptic signaling In animals, the cells of the nervous system provide rapid communication with distant cells. Their signal molecules, neurotransmitters, do not travel to the distant cells through the circulatory system like hormones do. Rather, the long, fiberlike extensions of nerve cells release neuro-transmitters from their tips very close to the target cells. The narrow gap between the two cells is called a chemical synapse . While paracrine signals move through the fluid between cells, neurotransmitters cross the synapse and persist only briefly. Adjacent cells can signal others by direct contact, while nearby cells that are not touching can communicate through paracrine signals.
Notes made by kozi1989 email:kozi1989@yahoo.gr
10
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
A cell junction (or intercellular bridge) is a type of structure that exists within the tissue of a some multicellular organism (- for example true for animals, but not plants, which possess plasmodesmata instead). Cell junctions consist of protein complexes and provide contact between neighbouring cells or between a cell and the extracellular matrix. They also build up the paracellular barrier of epithelia and control the paracellular transport. Cell junctions are especially abundant in epithelial tissues. A gap junction or nexus is a specialized intercellular connection between a multitude of animal cell-types. It directly connects the cytoplasm of two cells, which allows various molecules and ions to pass freely between cells. One gap junction channel is composed of two connexons (or hemichannels) which connect across the intercellular space. Gap junctions are analogous to the plasmodesmata that join plant cells.A notable use of gap junctions is in the electrical synapse found in some neurons. Tight junctions, or zonula occludens, are the closely associated areas of two cells whose membranes join together forming a virtually impermeable barrier to fluid . It is a type of junctional complex present only in vertebrates. The corresponding junctions that occur in invertebrates are septate junctions. Adherens junctions (or zonula adherens, intermediate junction, or "belt desmosome") are protein complexes that occur at cellcell junctions in epithelial tissues, usually more basal than tight junctions. An adherens junction is defined as a cell junction whose cytoplasmic face is linked to the actin cytoskeleton. They can appear as bands encircling the cell (zonula adherens) or as spots of attachment to the extracellular matrix (adhesion plaques). A similar cell junction in non-epithelial cells is the fascia adherens. It is structurally the same, but appears in ribbonlike patterns that do not completely encircle the cells. One example is in cardiomyocytes. A receptor is a molecule found on the surface of a cell, which receives specific chemical signals from neighbouring cells or the wider environment within an organism. These signals tell a cell to do somethingfor example to divide or die, or to allow certain molecules to enter or exit the cell. In biochemistry, a receptor is a protein molecule, embedded in either the plasma membrane or the cytoplasm of a cell, to which one or more specific kinds of signaling molecules may attach. A molecule which binds (attaches) to a receptor is called a ligand , and may be a peptide (short protein) or other small molecule, such as a neurotransmitter, a hormone, a pharmaceutical drug, or a toxin. Each kind of receptor can bind only certain ligand shapes. Each cell typically has many receptors, of many different kinds. Simply put, a receptor functions as a keyhole that opens a neural path when the proper ligand is inserted. Ligand binding stabilizes a certain receptor conformation (the three-dimensional shape of the receptor protein, with no change in sequence). This is often associated with gain of or loss of protein activity, ordinarily leading to some sort of cellular response. However, some ligands (e.g. antagonists) merely block receptors without inducing any
Notes made by kozi1989 email:kozi1989@yahoo.gr
11
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
response. Ligand-induced changes in receptors result in cellular changes which constitute the biological activity of the ligands. Many functions of the human body are regulated by these receptors responding uniquely to specific molecules like this. Depending on their functions and ligands, several types of receptors may be identified: 1) Some receptor proteins are peripheral membrane proteins. 2) Many hormone and neurotransmitter receptors are transmembrane proteins: transmembrane receptors are embedded in the phospholipid bilayer of cell membranes, that allow the activation of signal transduction pathways in response to the activation by the binding molecule, or ligand. 3) Metabotropic receptors are coupled to G proteins and affect the cell indirectly through enzymes which control ion channels. 4) Ionotropic receptors (also known as ligand-gated ion channels) contain a central pore which opens in response to the binding of ligand. 5) Another major class of receptors are intracellular proteins such as those for steroid and intracrine peptide hormone receptors. These receptors often can enter the cell nucleus and modulate gene expression in response to the activation by the ligand. Membrane receptors are isolated from cell membranes by complex extraction procedures using solvents, detergents, and/or affinity purification.
6. Individual development of cells. Cell senescence.
Elements of cell senescence. Apoptosis and necrosis. Cellular senescence is the phenomenon by which normal diploid cells lose the ability to divide, normally after about 50 cell divisions in vitro. Some cells become senescent after fewer replications cycles as a result of DNA double strand breaks, toxins, etc In response to DNA damage (including shortened telomeres), cells either age or selfdestruct (apoptosis, programmed cell death) if the damage cannot be easily repaired. In this 'cellular suicide', the death of one cell, or more, may benefit the organism as a whole. The ones that do not self-destruct remain until destroyed by outside forces. Apoptosisis the process of programmed cell death (PCD) that may occur in multicellular organisms. Biochemical events lead to characteristic cell changes (morphology) and death. These changes include: 1) Blebbing 2) Cell shrinkage 3) Nuclear fragmentation 4) Chromatin condensation 5) And chromosomal dna fragmentation. (See also Apoptosis DNA fragmentation.)
Notes made by kozi1989 email:kozi1989@yahoo.gr
12
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Unlike necrosis, apoptosis produces cell fragments called apoptotic bodies that phagocytic cells are able to engulf and quickly remove before the contents of the cell can spill out onto surrounding cells and cause damage. In contrast to necrosis, which is a form of traumatic cell death that results from acute cellular injury, apoptosis, in general, confers advantages during an organism's life cycle. For example, the differentiation of fingers and toes in a developing human embryo occurs because cells between the fingers apoptose; the result is that the digits are separate. Between 50 and 70 billion cells die each day due to apoptosis in the average human adult. The process of apoptosis is controlled by a diverse range of cell signals, which may originate either extracellularly (extrinsic inducers) or intracellularly (intrinsic inducers).Extracellular signals may include toxins, hormones, growth factors, nitric oxide or cytokines, that must either cross the plasma membrane or transduce to effect a response. These signals may positively (i.e., trigger) or negatively (i.e., repress, inhibit, or dampen) affect apoptosis. (Binding and subsequent trigger of apoptosis by a molecule is termed positive induction, whereas the active repression or inhibition of apoptosis by a molecule is termed negative induction.) A cell initiates intracellular apoptotic signalling in response to a stress, which may bring about cell suicide. The binding of nuclear receptors by glucocorticoids, heat, radiation, nutrient deprivation, viral infection, hypoxia and increased intracellular calcium concentration, for example, by damage to the membrane, can all trigger the release of intracellular apoptotic signals by a damaged cell. A number of cellular components, such as poly ADP ribose polymerase, may also help regulate apoptosis. Before the actual process of cell death is precipitated by enzymes, apoptotic signals must cause regulatory proteins to initiate the apoptosis pathway. This step allows apoptotic signals to cause cell death, or the process to be stopped, should the cell no longer need to die. Several proteins are involved, but two main methods of regulation have been identified: targeting mitochondria functionality, or directly transducing the signal via adaptor proteins to the apoptotic mechanisms. Another extrinsic pathway for initiation identified in several toxin studies is an increase in calcium concentration within a cell caused by drug activity, which also can cause apoptosis via a calcium binding protease calpain. Necrosisis the premature death of cells and living tissue. Necrosis is caused by factors external to the cell or tissue, such as infection, toxins, or trauma. This is in contrast to apoptosis, which is a naturally occurring cause of cellular death. While apoptosis often provides beneficial effects to the organism, necrosis is almost always detrimental and can be fatal. Cells that die due to necrosis do not usually send the same chemical signals to the immune system that cells undergoing apoptosis do. This prevents nearby phagocytes from locating and engulfing the dead cells, leading to a build-up of dead tissue and cell debris at or near the site of the cell death. For this reason, it is often necessary to remove necrotic tissue surgically, a process known as debridement. There are seven distinctive morphologic patterns of necrosis:
Notes made by kozi1989 email:kozi1989@yahoo.gr
13
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
1) Coagulative necrosis is typically seen in hypoxic (low-oxygen) environments, such as an infarction. Cell outlines remain after cell death and can be observed by light microscopy. 2) Liquefactive necrosis (or colliquative necrosis) is usually associated with cellular destruction and pus formation (e.g., pneumonia). This is typical of bacterial or, sometimes, fungal infections because of their ability to stimulate an inflammatory reaction. It is interesting to note that ischemia (restriction of blood supply) in the brain produces liquefactive, rather than coagulative necrosis due to the lack of any substantial supportive stroma. 3) Gummatous necrosis is restricted to necrosis involving spirochaetal infections (e.g., syphilis). 4) Haemorrhagic necrosis is due to blockage of the venous drainage of an organ or tissue (e.g., in testicular torsion). 5) Caseous necrosis is a specific form of coagulation necrosis typically caused by mycobacteria (e.g., tuberculosis), fungi, and some foreign substances. It can be considered a combination of coagulative and liquefactive necrosis. 6) Fatty necrosis results from the action of lipases on fatty tissues (e.g., acute pancreatitis, breast tissue necrosis). 7) Fibrinoid necrosis is caused by immune-mediated vascular damage. It is marked by deposition of fibrin-like proteinaceous material in arterial walls, which appears smudgy and eosinophilic on light microscopy.
7. Cell reproduction
Cell cycle. The cell cycle, or cell-division cycle, is the series of events that takes place in a cell leading to its division and duplication (replication). In cells without a nucleus (prokaryotic), the cell cycle occurs via a process termed binary fission. In cells with a nucleus (eukaryotes), the cell cycle can be divided in two brief periods: interphaseduring which the cell grows, accumulating nutrients needed for mitosis and duplicating its DNA and the mitosis (M) phase, during which the cell splits itself into two distinct cells, often called " daughter cells". The cell cycle consists of four distinct phases: G1 phase, S phase (synthesis), G2 phase (collectively known as interphase) and M phase (mitosis). M phase is itself composed of two tightly coupled processes: mitosis, in which the cell's chromosomes are divided between the two daughter cells, and cytokinesis, in which the cell's cytoplasm divides in half forming distinct cells. Activation of each phase is dependent on the proper progression and completion of the previous one. Cells that have temporarily or reversibly stopped dividing are said to have entered a state of quiescence called G0 phase. State Phase Abbreviation Description
Notes made by kozi1989 email:kozi1989@yahoo.gr
14
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
quiescent/ senescent Interphase
Gap 0 Gap 1
G0 G1
A resting phase where the cell has left the cycle and has stopped dividing. Cells increase in size in Gap 1. The G1 checkpoint control mechanism ensures that everything is ready for DNA synthesis. DNA replication occurs during this phase.
Synthesis Gap 2
S G2
During the gap between DNA synthesis and mitosis, the cell will continue to grow. The G2 checkpoint control mechanism ensures that everything is ready to enter the M (mitosis) phase and divide. Cell Mitosis M Cell growth stops at this stage and cellular energy division is focused on the orderly division into two daughter cells. A checkpoint in the middle of mitosis (Metaphase Checkpoint) ensures that the cell is ready to complete cell division. After cell division, each of the daughter cells begin the interphase of a new cycle. Although the various stages of interphase are not usually morphologically distinguishable, each phase of the cell cycle has a distinct set of specialized biochemical processes that prepare the cell for initiation of cell division. Resting (G0 phase) The term "post-mitotic" is sometimes used to refer to both quiescent and senescent cells. Nonproliferative cells in multicellular eukaryotes generally enter the quiescent G0 state from G1 and may remain quiescent for long periods of time, possibly indefinitely (as is often the case for neurons). This is very common for cells that are fully differentiated. Cellular senescence is a state that occurs in response to DNA damage or degradation that would make a cell's progeny nonviable; it is often a biochemical alternative to the selfdestruction of such a damaged cell by apoptosis. Interphase Before a cell can enter cell division, it needs to take in nutrients. All of the preparations are done during the interphase. Interphase proceeds in three stages, G1, S, and G2. Cell division operates in a cycle. Therefore, interphase is preceded by the previous cycle of mitosis and cytokinesis. G1 phase The first phase within interphase, from the end of the previous M phase until the beginning of DNA synthesis is called G1 (G indicating gap). It is also called the growth phase. During this phase the biosynthetic activities of the cell, which had been considerably slowed down during M phase, resume at a high rate. This phase is marked by synthesis of
Notes made by kozi1989 email:kozi1989@yahoo.gr
15
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
various enzymes that are required in S phase, mainly those needed for DNA replication. Duration of G1 is highly variable, even among different cells of the same species. S phase The ensuing S phase starts when DNA synthesis commences, when it is complete, all of the chromosomes have been replicated, i.e., each chromosome has two (sister) chromatids. Thus, during this phase, the amount of DNA in the cell has effectively doubled , though the ploidy of the cell remains the same. Rates of RNA transcription and protein synthesis are very low during this phase. An exception to this is histone production, most of which occurs during the S phase. G2 phase The cell then enters the G2 phase, which lasts until the cell enters mitosis. Again, significant biosynthesis occurs during this phase, mainly involving the production of microtubules, which are required during the process of mitosis. Inhibition of protein synthesis during G2 phase prevents the cell from undergoing mitosis. M phase Mitosisis the process by which a eukaryotic cell separates the chromosomes in its cell nucleus into two identical sets in two nuclei. It is generally followed immediately by cytokinesis, which divides the nuclei, cytoplasm, organelles and cell membrane into two cells containing roughly equal shares of these cellular components. Mitosis and cytokinesis together define the mitotic (M) phase of the cell cyclethe division of the mother cell into two daughter cells, genetically identical to each other and to their parent cell. This accounts for approximately 10% of the cell cycle.
8. Cell reproduction
Notes made by kozi1989 email:kozi1989@yahoo.gr
16
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Consecutive events in mitosis (M-phase) and cytokinesis. Cell cycle control, MPF description. Mitosis is the process by which a eukaryotic cell separates the chromosomes in its cell nucleus into two identical sets in two nuclei. It is generally followed immediately by cytokinesis, which divides the nuclei, cytoplasm, organelles and cell membrane into two cells containing roughly equal shares of these cellular components. Mitosis and cytokinesis together define the mitotic (M) phase of the cell cyclethe division of the mother cell into two daughter cells, genetically identical to each other and to their parent cell. This accounts for approximately 10% of the cell cycle. Mitosis occurs only in eukaryotic cells and the process varies in different species. For example, animals undergo an "open" mitosis, where the nuclear envelope breaks down before the chromosomes separate, while fungi such as Aspergillus nidulans and Saccharomyces cerevisiae (yeast) undergo a "closed" mitosis, where chromosomes divide within an intact cell nucleus.Prokaryotic cells, which lack a nucleus, divide by a process called binary fission. The process of mitosis is fast and highly complex. The sequence of events is divided into stages corresponding to the completion of one set of activities and the start of the next. These stages are interphase, prophase, prometaphase, metaphase, anaphase and telophase. During mitosis the pairs of chromatids condense and attach to fibers that pull the sister chromatids to opposite sides of the cell. The cell then divides in cytokinesis, to produce two identical daughter cells. Because cytokinesis usually occurs in conjunction with mitosis, "mitosis" is often used interchangeably with "mitotic phase". However, there are many cells where mitosis and cytokinesis occur separately, forming single cells with multiple nuclei. This occurs most notably among the fungi and slime moulds, but is found in various different groups. Even in animals, cytokinesis and mitosis may occur independently, for instance during certain stages of fruit fly embryonic development. Errors in mitosis can either kill a cell through apoptosis or cause mutations that may lead to cancer. The primary result of mitosis is the transferring of the parent cell's genome into two daughter cells. These two cells are identical and do not differ in any way from the original parent cell. The genome is composed of a number of chromosomescomplexes of tightly-coiled DNA that contain genetic information vital for proper cell function. Because each resultant daughter cell should be genetically identical to the parent cell, the parent cell must make a copy of each chromosome before mitosis. This occurs during the S phase of interphase, the period that precedes the mitotic phase in the cell cycle where preparation for mitosis occurs. Each chromosome now has an identical copy of itself, and together the two are called homologus pairs. The homologus pairs are held together by a specialized region of the chromosome known as the centromere. Each homologus pair is not considered a chromosome in itself, and a chromosome always contains 2 chromatids. In most eukaryotes, the nuclear envelope which segregates the DNA from the
Notes made by kozi1989 email:kozi1989@yahoo.gr
17
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
cytoplasm disassembles. The chromosomes align themselves in a line spanning the cell. Microtubules, essentially miniature strings, splay out from opposite ends of the cell and shorten, pulling apart the sister chromatids of each chromosome. As a matter of convention, each sister chromatid is now considered a chromosome, so they are renamed to sister chromosomes. As the cell elongates, corresponding sister chromosomes are pulled toward opposite ends. A new nuclear envelope forms around the separated sister chromosomes. As mitosis completes,the cell starts cytokinesis. In animal cells, the cell pinches inward where the imaginary line used to be (the area of the cell membrane that pinches to form the two daughter cells is called the cleavage furrow), separating the two developing nuclei. In plant cells, the daughter cells will construct a new dividing cell wall between each other. Eventually, the parent cell will be split in half, giving rise to two daughter cells, each with a replica of the original genome. Prokaryotic cells undergo a process similar to mitosis called binary fission. However, the process of binary fission is very much different from the process of mitosis, because of the non-involvement of nuclear dynamics and lack of linear chromosomes. Phases of cell cycle and mitosis Interphase The mitotic phase is a relatively short period of the cell cycle. It alternates with the much longer interphase, where the cell prepares itself for cell division. Interphase is therefore not part of mitosis. Interphase is divided into three phases, G1 (first gap), S (synthesis), and G2 (second gap).During all three phases, the cell grows by producing proteins and cytoplasmic organelles. However, chromosomes are replicated only during the S phase. Thus, a cell grows (G1), continues to grow as it duplicates its chromosomes (S), grows more and prepares for mitosis (G2), and finally it divides (M) before restarting the cycle. All these phases in the interphase are highly regulated, mainly via proteins. The phases follow one another in strict order and there are "checkpoints" that give the cell the cues to proceed from one phase to another. Preprophase In plant cells only, prophase is preceded by a pre-prophase stage. In highly vacuolated plant cells, the nucleus has to migrate into the center of the cell before mitosis can begin. This is achieved through the formation of a phragmosome, a transverse sheet of cytoplasm that bisects the cell along the future plane of cell division. In addition to phragmosome formation, preprophase is characterized by the formation of a ring of microtubules and actin filaments (called preprophase band) underneath the plasma membrane around the equatorial plane of the future mitotic spindle. This band marks the position where the cell will eventually divide. The cells of higher plants (such as the flowering plants) lack centrioles; instead, microtubules form a spindle on the surface of the nucleus and are then being organized into a spindle by the chromosomes themselves, after the
Notes made by kozi1989 email:kozi1989@yahoo.gr
18
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
nuclear membrane breaks down. The preprophase band disappears during nuclear envelope disassembly and spindle formation in prometaphase. Prophase Normally, the genetic material in the nucleus is in a loosely bundled coil called chromatin. At the onset of prophase, chromatin condenses together into a highly ordered structure called a chromosome. Since the genetic material has already been duplicated earlier in S phase, the replicated chromosomes have two sister chromatids, bound together at the centromere by the cohesin protein complex. Chromosomes are typically visible at high magnification through a light microscope. Close to the nucleus are structures called centrosomes, which are made of a pair of centrioles. The centrosome is the coordinating center for the cell's microtubules. A cell inherits a single centrosome at cell division, which is replicated by the cell with the help of the nucleus before a new mitosis begins, giving a pair of centrosomes. The two centrosomes nucleate microtubules (which may be thought of as cellular ropes or poles) to form the spindle by polymerizing soluble tubulin. Molecular motor proteins then push the centrosomes along these microtubules to opposite sides of the cell. Although centrioles help organize microtubule assembly, they are not essential for the formation of the spindle, since they are absent from plants, and centrosomes are not always used in meiosis. Prometaphase The nuclear envelope disassembles and microtubules invade the nuclear space . This is called open mitosis, and it occurs in most multicellular organisms. Fungi and some protists, such as algae or trichomonads, undergo a variation called closed mitosis where the spindle forms inside the nucleus, or its microtubules are able to penetrate an intact nuclear envelope. Each chromosome forms two kinetochores at the centromere, one attached at each chromatid. A kinetochore is a complex protein structure that is analogous to a ring for the microtubule hook; it is the point where microtubules attach themselves to the chromosome ( about 1-40 in number, on an average 20 ). Although the kinetochore structure and function are not fully understood, it is known that it contains some form of molecular motor. When a microtubule connects with the kinetochore, the motor activates, using energy from ATP to "crawl" up the tube toward the originating centrosome. This motor activity, coupled with polymerisation and depolymerisation of microtubules, provides the pulling force necessary to later separate the chromosome's two chromatids. When the spindle grows to sufficient length, kinetochore microtubules begin searching for kinetochores to attach to. A number of nonkinetochore microtubules find and interact with corresponding nonkinetochore microtubules from the opposite centrosome to form the mitotic spindle. Prometaphase is sometimes considered part of prophase. In the fishing pole analogy, the kinetochore would be the "hook" that catches a sister chromatid or "fish". The centrosome acts as the "reel" that draws in the spindle fibers or
Notes made by kozi1989 email:kozi1989@yahoo.gr
19
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
"fishing line". It is also one of the main phases of mitosis because without it cytokinesis would not be able to occur. Metaphase Metaphase comes from the Greek meaning "after."Microtubules find and attach to kinetochores in prometaphase. Then the two centosomes start pulling the chromosomes through their attached centromeres towards the two ends of the cell. As a result of this the chromosomes come under longitudinal tension from the two ends of the cell. The centromeres of the chromosomes, in some sense, convene along the metaphase plate or equatorial plane, an imaginary line that is equidistant from the two centrosome poles. This even alignment is due to the counterbalance of the pulling powers generated by the opposing kinetochores, analogous to a tug-of-war between people of equal strength. In certain types of cells, chromosomes do not line up at the metaphase plate and instead move back and forth between the poles randomly, only roughly lining up along the midline. Because proper chromosome separation requires that every kinetochore be attached to a bundle of microtubules (spindle fibres), it is thought that unattached kinetochores generate a signal to prevent premature progression to anaphase without all chromosomes being aligned. The signal creates the mitotic spindle checkpoint. Anaphase When every kinetochore is attached to a cluster of microtubules and the chromosomes have lined up along the metaphase plate, the cell proceeds to anaphase (from the Greek meaning up, against, back, or re-). Two events then occur: first, the proteins that bind sister chromatids together are cleaved, allowing them to separate. These sister chromatids, which have now become distinct sister chromosomes, are pulled apart by shortening kinetochore microtubules and move toward the respective centrosomes to which they are attached. Next, the nonkinetochore microtubules elongate, pulling the centrosomes (and the set of chromosomes to which they are attached) apart to opposite ends of the cell. These two stages are sometimes called early and late anaphase. Early anaphase is usually defined as the separation of the sister chromatids, while late anaphase is the elongation of the microtubules and the chromosomes being pulled farther apart. At the end of anaphase, the cell has succeeded in separating identical copies of the genetic material into two distinct populations. Telophase Telophase (from the Greek meaning "end") is a reversal of prophase and prometaphase events. It "cleans up" the after effects of mitosis. At telophase, the nonkinetochore microtubules continue to lengthen, elongating the cell even more. Corresponding sister chromosomes attach at opposite ends of the cell. A new nuclear envelope, using fragments of the parent cell's nuclear membrane, forms around each set of
Notes made by kozi1989 email:kozi1989@yahoo.gr
20
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
separated sister chromosomes. Both sets of chromosomes, now surrounded by new nuclei, unfold back into chromatin. Mitosis is complete, but cell division is not yet complete. Cytokinesis Cytokinesis, from the greek cyto- (cell) and kinesis (motion, movement), is the process in which the cytoplasm of a single eukaryotic cell is divided to form two daughter cells. It is the division of the cytoplasm. It usually initiates during the late stages of mitosis, and sometimes meiosis, splitting a binucleate cell in two, to ensure that chromosome number is maintained from one generation to the next. In animal cells, one notable exception to the normal process of cytokinesis is oogenesis (the creation of an ovum in the ovarian follicle of the ovary), where the ovum takes almost all the cytoplasm and organelles, leaving very little for the resulting polar bodies, which then die. In plant cells, a dividing structure known as the cell plate forms across the centre of the cytoplasm and a new cell wall forms between the two daughter cells. Cell cycle checkpoints are control mechanisms that ensure the fidelity of cell division in eukaryotic cells. These checkpoints verify whether the processes at each phase of the cell cycle have been accurately completed before progression into the next phase. Multiple checkpoints have been identified, with some of them are less understood than others. An important function of many checkpoints is to assess DNA damage, which is detected by sensor mechanisms. When damage is found, the checkpoint uses a signal mechanism either to stall the cell cycle until repairs are made or, if repairs cannot be made, to target the cell for destruction via apoptosis (effector mechanism). All the checkpoints that assess DNA damage appear to utilize the same sensor-signal-effector mechanism. The cell cycle checkpoints are, therefore, made up of composites of protein kinases and adaptor proteins that all play salient roles in the maintenance of the integrity of the division. Checkpoints are now accepted to exist at every single point in the cell cycle. The DNA damage checkpoint is always active. Nonetheless, most human cells, for example, are terminally differentiated and must exit the cell cycle. There is a phase late in G1 phase called the restriction point (RP, or the restriction checkpoint); cells that should cease division exit the cell cycle and enter G0. Cells that continually divide in the adult human include hematopoietic stem cells and gut epithelial cells. Therefore, the re-entrant into the cell cycle is possible only by overcoming the RP. This is achieved by growth factor-induced expression of cyclin D proteins. These then overcome the G0 barrier and are able to enter the cell cycle.The main checkpoints that control the cell division cycle in eukaryotes include: G1 (Restriction) Checkpoint
Notes made by kozi1989 email:kozi1989@yahoo.gr
21
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The first checkpoint is located at the end of the cell cycle's G1 phase, just before entry into S phase, making the key decision of whether the cell should divide, delay division, or enter a resting stage. Many cells stop at this stage and enter a resting state called G0. Liver cells, for instance, enter mitosis only around once or twice a year. The G1 checkpoint is where eukaryotes typically arrest the cell cycle if environmental conditions make cell division impossible or if the cell passes into G0 for an extended period. In animal cells, the G1 phase checkpoint is called the restriction point, and in yeast cells it is called the Start point. The restriction point is controlled mainly by action of the CKI- p16 (CDK inhibitor p16). This protein inhibits the CDK4/6 and ensures that it can no longer interact with cyclin D1 to cause the cell cycle progression. In growth-induced or oncogenic-induced cyclin D expression, this checkpoint is overcome because the increased expression of cyclin D allows its interaction with CDK4/6 by competing for binding. Once active CDK4/6-CYCLIN D complexes form, they phosphorylate the tumour suppressor retinoblastoma (Rb), which relieves the inhibition of the transcription factor E2F. E2F is then able to cause expression of cyclin E, which then interacts with CDK2 to allow for G1-S phase transition. That brings us to the end of the first checkpoint, which allows the G0-G1-S-phase transition. In simpler terms, the CDK inhibitor p16 inhibits another CDK from binding to its cyclin (D). When growth is induced, the expression of this cyclin is so high that they do bind. The new CDK/cyclin complex now phosphorylates retinoblastoma (a tumour suppressor). Unphosphorylated retinoblastoma inhibits a transcription factor. This factor then brings about the G1-S phase transition. G2 Checkpoint The second checkpoint is located at the end of G2 phase, triggering the start of the M phase (mitosis). In order for this checkpoint to be passed, the cell has to check a number of factors to ensure the cell is ready for mitosis. If this checkpoint is passed, the cell initiates the many molecular processes that signal the beginning of mitosis. The CDKs associated with this checkpoint are activated by phosphorylation of the CDK by the action of a "Maturation promoting factor" (or Mitosis Promoting Factor, MPF). The molecular nature of this checkpoint involves an activating phosphatase , known as Cdc25, which under favourable conditions removes the inhibitory phosphates present within the MPF complex. However, DNA is frequently damaged prior to mitosis, and, to prevent transmission of this damage to daughter cells, the cell cycle is arrested via inactivation of the Cdc25 phosphatase (via phosphorylation with other protein kinases). Metaphase Checkpoint The mitotic spindle checkpoint occurs at the point in metaphase where all the chromosomes have/should have aligned at the mitotic plate and be under bipolar tension. The tension created by this bipolar attachment is what is sensed, which initiates the anaphase entry. This sensing mechanism allows the degradation of cyclin B, which harbours a D-box (destruction box). Degradation of cyclin B ensures that it no longer inhibits the
Notes made by kozi1989 email:kozi1989@yahoo.gr
22
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
anaphase-promoting complex, which in turn is now free to break down securin. The latter is a protein whose function is to inhibit separase, the protein composite responsible for the separation of sister chromatids. Once this inhibitory protein is degraded via ubiquitination and subsequent proteolysis, separase then causes sister chromatid separation . After the cell has split into its two daughter cells, the cell enters G1. Maturation-promoting factor (abbreviated MPF, also called mitosis-promoting factor or M-Phase-promoting factor) is a heterodimeric protein composed of cyclin B or cyclin A and cyclin-dependent kinase (CDK1, also known as Cdc2 or p34 kinase) that stimulates the mitotic and meiotic cell cycles. MPF promotes the entrance into mitosis from the G2 phase by phosphorylating multiple proteins needed during mitosis. MPF is activated at the end of G2 by a phosphatase, which removes an inhibitory phosphate group added earlier. MPF is made of 2 subunits: 1) A subunit that transfers phosphate groups from ATP to specific serine and threonine residues of specific proteins (kinase activity) 2) Cyclin, a regulatory subunit Targets of MPF include: 1) 2) 3) 4) 5) Condensins, which enable chromatin condensation (see prophase) Various microtubule-associated proteins involved in mitotic spindle formation Lamins, interaction contributing to degradation of the nuclear envelope Histones, h1 and h3 Golgi matrix, to cause fragmentation
MPF phosphorylates inhibitory sites on myosin early in mitosis. This prevents cytokinesis. When MPF activity falls at anaphase, the inhibitory sites are dephosphorylated and cytokinesis proceeds. MPF must be activated in order for the cell to transition from G2 to M phase. There are 3 amino acid residues responsible for this G2 to M phase transition. The threonine 161 (Thr-161) on cdk1 must be phosphorylated by a Cyclin Activating Kinase (CAK). CAK only phosphorylates Thr-161 when cyclinB is attached to cdk1. In addition, two other residues on the cdk1 subunit must be activated. cdc25 removes a phosphate on residues Threonine-14 (Thr-14) and Tyrosine-15 (Tyr-15) and adds a hydroxy group. CyclinB/cdk1 activates cdc25 resulting in a positive feedback loop. During G1 and S phase, the MPF subunit cdk1 is inactive due to an inhibitory enzyme, Wee1. Wee1 phosphorylizes the Thr-14 and Tyr-15 residues. During the transition of G2 to M phase, MPF inhibits Wee1, again resulting in a positive feedback loop. MPF is disassembled when anaphase-promoting complex (APC) polyubiquitinates cyclin B, marking it for degradation in a negative feedback loop. As the concentration of CyclinB/cdk1 increases, the heterodimer promotes APC to polyubiquitinate CyclinB/cdk1. MPF is also present in somatic cells, where it induces entry into M phase of the
Notes made by kozi1989 email:kozi1989@yahoo.gr
23
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
mitotic cycle The MPF is also called the M phase kinase because of its ability to phosphorylate target proteins at a specific point in the cell cycle and thus controlling their ability to function. The MPF has two subunits: The Cyclins and the Cyclin dependent Kinases(CDK) The cyclins are the regulatory subunits that are necessary for the kinase to function with appropriate substrate. The mitotic cyclins can be grouped as cyclins A & B. The CDK is the catalaytic subunit that phosphorylates serine and threonine residues in target proteins. The MPF is a heterodimeric protein that act as the B type cyclins . These cyclins have a nine residue sequence in the N- terminal region called as the destruction box, which can be recognized by the ubiquitin ligase enzyme and thus destroys the cyclins. In intact cells, cyclin degradation begins shortly after the onset of anaphase (late anaphase), the period of mitosis when sister chromatids are separated and pulled toward opposite spindle poles. Covalent attachment of chains of ubiquitin, a process called polyubiquitination, marks proteins for rapid degradation in eukaryotic cells by proteasomes, multiprotein cylindrical structures containing numerous proteases. Triggers the formation of mitotic spindle. Promotes mitosis i.e. chromatin condensation. Causes nuclear envelope breakdown by phosphorylating the lamins that form an intermediate filament-type network (nuclear lamina) underlying the inner nuclear membrane. The three lamins present in the nuclear lamina, lamin A,B & C, are phosphorylated by MPF at serine amino residues. This leads to depolymerisation of nuclear lamina & breakdown of nuclear envelope into small vesicles.
9. Molecular mechanisms of the basic genetic processes
DNA replication. Basic modes of replication. DNA repair DNA replicationis a biological process that occurs in all living organisms and copies their DNA, it is the basis for biological inheritance. The process starts with one doublestranded DNA molecule and produces two identical copies of the molecule. Each strand of the original double-stranded DNA molecule serves as template for the production of the complementary strand. Cellular proofreading and error toe-checking mechanisms ensure near perfect fidelity for DNA replication. In a cell, DNA replication begins at specific locations in the genome, called "origins". Unwinding of DNA at the origin, and synthesis of new strands, forms a replication fork. In addition to DNA polymerase, the enzyme that synthesizes the new DNA by adding nucleotides matched to the template strand, a number of other proteins are associated with the fork and assist in the initiation and continuation of DNA synthesis. DNA replication can also be performed in vitro (artificially, outside a cell). DNA polymerases, isolated from cells, and artificial DNA primers are used to initiate DNA synthesis at known sequences in a template molecule. The polymerase chain reaction (PCR), a common laboratory technique, employs such artificial synthesis in a cyclic manner to amplify a specific target DNA fragment from a pool of DNA. DNA usually exists as a double-stranded structure, with both strands coiled together
Notes made by kozi1989 email:kozi1989@yahoo.gr
24
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
to form the characteristic double-helix. Each single strand of DNA is a chain of four types of nucleotides having the bases: adenine, cytosine, guanine, and thymine. A nucleotide is a mono-, di-, or triphosphate deoxyribonucleoside; that is, a deoxyribose sugar is attached to one, two, or three phosphates. Chemical interaction of these nucleotides forms phosphodiester linkages, creating the phosphate-deoxyribose backbone of the DNA double helix with the bases pointing inward. Nucleotides (bases) are matched between strands through hydrogen bonds to form base pairs. Adenine pairs with thymine, and cytosine pairs with guanine. DNA strands have a directionality, and the different ends of a single strand are called the "3' (three-prime) end" and the "5' (five-prime) end". These terms refer to the carbon atom in deoxyribose to which the next phosphate in the chain attaches. In addition to being complementary, the two strands of DNA are antiparallel: They are orientated in opposite directions. This directionality has consequences in DNA synthesis, because DNA polymerase can synthesize DNA in only one direction by adding nucleotides to the 3' end of a DNA strand. The pairing of bases in DNA through hydrogen bonding means that the information contained within each strand is redundant. The nucleotides on a single strand can be used to reconstruct nucleotides on a newly synthesized partner strand. DNA polymerase DNA polymerases adds nucleotides to the 5' end of a strand of DNA. If a mismatch is accidentally incorporated, the polymerase is inhibited from further extension. Proofreading removes the mismatched nucleotide and extension continues. DNA polymerases are a family of enzymes that carry out all forms of DNA replication. However, a DNA polymerase can only extend an existing DNA strand paired with a template strand, it cannot begin the synthesis of a new strand. To begin synthesis, a short fragment of DNA or RNA, called a primer, must be created and paired with the template DNA strand. DNA polymerase then synthesizes a new strand of DNA by extending the 3' end of an existing nucleotide chain, adding new nucleotides matched to the template strand one at a time via the creation of phosphodiester bonds. The energy for this process of DNA polymerization comes from two of the three total phosphates attached to each unincorporated base. (Free bases with their attached phosphate groups are called nucleoside triphosphates.) When a nucleotide is being added to a growing DNA strand, two of the phosphates are removed and the energy produced creates a phosphodiester bond that attaches the remaining phosphate to the growing chain. The energetics of this process also help explain the directionality of synthesis - if DNA were synthesized in the 3' to 5' direction, the energy for the process would come from the 5' end of the growing strand rather than from free nucleotides. In general, DNA polymerases are extremely accurate, making less than one mistake for every 107 nucleotides added. Even so, some DNA polymerases also have proofreading ability, they can remove nucleotides from the end of a strand in order to correct mismatched
Notes made by kozi1989 email:kozi1989@yahoo.gr
25
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
bases. If the 5' nucleotide needs to be removed during proofreading, the triphosphate end is lost. Hence, the energy source that usually provides energy to add a new nucleotide is also lost. Replication process For a cell to divide, it must first replicate its DNA. This process is initiated at particular points in the DNA, known as "origins", which are targeted by proteins that separate the two strands and initiate DNA synthesis. Origins contain DNA sequences recognized by replication initiator proteins (e.g., dnaA in E. coli' and the Origin Recognition Complex in yeast). These initiator proteins recruit other proteins to separate the two strands and initiate replication forks. Initiator proteins recruit other proteins to separate the DNA strands at the origin, forming a bubble. Origins tend to be "AT-rich" (rich in adenine and thymine bases) to assist this process, because A-T base pairs have two hydrogen bonds (rather than the three formed in a C-G pair)in general, strands rich in these nucleotides are easier to separate because a greater number of hydrogen bonds requires more energy to break them. Once strands are separated, RNA primers are created on the template strands. To be more specific, the leading strand receives one RNA primer per active origin of replication while the lagging strand receives several, these several fragments of RNA primers found on the lagging strand of DNA are called Okazaki fragments, named after their discoverer. DNA Polymerase extends the leading strand in one continuous motion and the lagging strand in a discontinuous motion (due to the Okazaki fragments). RNase removes the RNA fragments used to initiate replication by DNA Polymerase, and another DNA Polymerase enters to fill the gaps. When this is complete, a single nick on the leading strand and several nicks on the lagging strand can be found. Ligase works to fill these nicks in, thus completing the newly replicated DNA molecule. As DNA synthesis continues, the original DNA strands continue to unwind on each side of the bubble, forming a replication fork with two prongs . In bacteria, which have a single origin of replication on their circular chromosome, this process eventually creates a "theta structure" (resembling the Greek letter theta: ). In contrast, eukaryotes have longer linear chromosomes and initiate replication at multiple origins within these.[citation needed] Replication fork The replication fork is a structure that forms within the nucleus during DNA replication. It is created by helicases, which break the hydrogen bonds holding the two DNA strands together. The resulting structure has two branching "prongs", each one made up of a single strand of DNA. These two strands serve as the template for the leading and lagging strands, which will be created as DNA polymerase matches complementary nucleotides to the templates; The templates may be properly referred to as the leading strand template and the lagging strand template.
Notes made by kozi1989 email:kozi1989@yahoo.gr
26
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Leading strand The leading strand is the template strand of the DNA double helix so that the replication fork moves along it in the 3' to 5' direction. This allows the new strand synthesized complementary to it to be synthesized 5' to 3' in the same direction as the movement of the replication fork. On the leading strand, a polymerase "reads" the DNA and adds nucleotides to it continuously. This polymerase is DNA polymerase III (DNA Pol III) in prokaryotes and presumably Pol in eukaryotes. Lagging strand The lagging strand is the strand of the template DNA double helix that is oriented so that the replication fork moves along it in a 5' to 3' manner. Because of its orientation, opposite to the working orientation of DNA polymerase III, which moves on a template in a 3' to 5' manner, replication of the lagging strand is more complicated than that of the leading strand. On the lagging strand, primase "reads" the DNA and adds RNA to it in short, separated segments. In eukaryotes, primase is intrinsic to Pol . DNA polymerase III or Pol lengthens the primed segments, forming Okazaki fragments. Primer removal in eukaryotes is also performed by Pol . In prokaryotes, DNA polymerase I "reads" the fragments, removes the RNA using its flap endonuclease domain (RNA primers are removed by 5'-3' exonuclease activity of polymerase I, and replaces the RNA nucleotides with DNA nucleotides (this is necessary because RNA and DNA use slightly different kinds of nucleotides). DNA ligase joins the fragments together. Three types of replication are been proposed by different scientist namely Conservative mode, dispersive mode, and semi conservative mode. Out of these three types conservative mode and dispersive mode are discarded are they are not proved experimentally. Semi conservative mode of replication is proved experimentally and accepted type of replication. Conservative Mode Conservative mode is a postulated type of DNA replication in which double stranded helix remains intact and acts as a template for a new double stranded DNA helix. This type of DNA replication is disproved model, in this type of replication one-half of the daughters DNA molecules will have both strands composed of newly polymerized nucleotides and the old parent double stranded helix will remain unchanged. Dispersive Mode In dispersive type of DNA replication the parent DNA molecule is said undergo fragmentation to produce many fragments of DNA. Each fragment will replicate and then
Notes made by kozi1989 email:kozi1989@yahoo.gr
27
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
the fragments intermix with the newly synthesized fragments to form new double helical DNA molecule. The newly formed DAN would have patchwork of old DNA and new fragments. But this model of DNA replication cannot be accepted and discarded since it is not proved experimentally. Semi Conservative Mode In semi conservative replication process the each molecule of parent DNA double helix results in two hybrid DNAs. Here each of the newly synthesized daughter DNA double helices retains a parent DNA strand and a newly synthesized strand. This mode of DNA replication is proposed by Watson and Crick, for which they were awarded with Nobel Prize. Semi conservative mode of replication is proved by the experiments of Mathew Messelson and Franklin W. Stall. This method is proved using isotopically labeled DNA and a type of isopycnic density gradient centrifugation. DNA repair refers to a collection of processes by which a cell identifies and corrects damage to the DNA molecules that encode its genome. In human cells, both normal metabolic activities and environmental factors such as UV light and radiation can cause DNA damage, resulting in as many as 1 million individual molecular lesions per cell per day. Many of these lesions cause structural damage to the DNA molecule and can alter or eliminate the cell's ability to transcribe the gene that the affected DNA encodes. Other lesions induce potentially harmful mutations in the cell's genome, which affect the survival of its daughter cells after it undergoes mitosis. As a consequence, the DNA repair process is constantly active as it responds to damage in the DNA structure. When normal repair processes fail, and when cellular apoptosis does not occur, irreparable DNA damage may occur, including double-strand breaks and DNA crosslinkages. The rate of DNA repair is dependent on many factors, including the cell type, the age of the cell, and the extracellular environment. A cell that has accumulated a large amount of DNA damage, or one that no longer effectively repairs damage incurred to its DNA, can enter one of three possible states: 1) An irreversible state of dormancy, known as senescence 2) Cell suicide, also known as apoptosis or programmed cell death 3) Unregulated cell division, which can lead to the formation of a tumor that is cancerous The DNA repair ability of a cell is vital to the integrity of its genome and thus to its normal functioning and that of the organism. Many genes that were initially shown to influence life span have turned out to be involved in DNA damage repair and protection. Failure to correct molecular lesions in cells that form gametes can introduce mutations into the genomes of the offspring and thus influence the rate of evolution. Cells cannot function if DNA damage corrupts the integrity and accessibility of essential information in the genome (but cells remain superficially functional when so-
Notes made by kozi1989 email:kozi1989@yahoo.gr
28
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
called "non-essential" genes are missing or damaged). Depending on the type of damage inflicted on the DNA's double helical structure, a variety of repair strategies have evolved to restore lost information. If possible, cells use the unmodified complementary strand of the DNA or the sister chromatid as a template to recover the original information. Without access to a template, cells use an error-prone recovery mechanism known as translation synthesis as a last resort. Damage to DNA alters the spatial configuration of the helix, and such alterations can be detected by the cell. Once damage is localized, specific DNA repair molecules bind at or near the site of damage, inducing other molecules to bind and form a complex that enables the actual repair to take place. Direct reversal Cells are known to eliminate three types of damage to their DNA by chemically reversing it. These mechanisms do not require a template, since the types of damage they counteract can occur in only one of the four bases. Such direct reversal mechanisms are specific to the type of damage incurred and do not involve breakage of the phosphodiester backbone. The formation of pyrimidine dimers upon irradiation with UV light results in an abnormal covalent bond between adjacent pyrimidine bases. The photoreactivation process directly reverses this damage by the action of the enzyme photolyase, whose activation is obligately dependent on energy absorbed from blue/UV light (300 500 nm wavelength) to promote catalysis. Another type of damage, methylation of guanine bases, is directly reversed by the protein methyl guanine methyl transferase (MGMT), the bacterial equivalent of which is called ogt. This is an expensive process because each MGMT molecule can be used only once; that is, the reaction is stoichiometric rather than catalytic. A generalized response to methylating agents in bacteria is known as the adaptive response and confers a level of resistance to alkylating agents upon sustained exposure by upregulation of alkylation repair enzymes. The third type of DNA damage reversed by cells is certain methylation of the bases cytosine and adenine. Single-strand damage When only one of the two strands of a double helix has a defect, the other strand can be used as a template to guide the correction of the damaged strand. In order to repair damage to one of the two paired molecules of DNA, there exist a number of excision repair mechanisms that remove the damaged nucleotide and replace it with an undamaged nucleotide complementary to that found in the undamaged DNA strand. 1) Base excision repair (BER), which repairs damage to a single base caused by oxidation, alkylation, hydrolysis, or deamination. The damaged base is removed by a DNA glycosylase. The "missing tooth" is then recognised by an enzyme called AP endonuclease, which cuts the Phosphodiester bond. The missing part is then
Notes made by kozi1989 email:kozi1989@yahoo.gr
29
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
resynthesized by a DNA polymerase, and a DNA ligase performs the final nick-sealing step. 2) Nucleotide excision repair (NER), which recognizes bulky, helix-distorting lesions such as pyrimidine dimers and 6,4 photoproducts. A specialized form of NER known as transcription-coupled repair deploys NER enzymes to genes that are being actively transcribed. 3) Mismatch repair (MMR), which corrects errors of DNA replication and recombination that result in mispaired (but undamaged) nucleotides. Double-strand breaks Double-strand breaks, in which both strands in the double helix are severed, are particularly hazardous to the cell because they can lead to genome rearrangements. Three mechanisms exist to repair double-strand breaks (DSBs): non-homologous end joining (NHEJ), microhomology-mediated end joining (MMEJ), and homologous recombination. PVN Acharya noted that double-strand breaks and a "cross-linkage joining both strands at the same point is irreparable because neither strand can then serve as a template for repair. The cell will die in the next mitosis or in some rare instances, mutate." DNA ligase is an enzyme that joins broken nucleotides together by catalyzing the formation of an internucleotide ester bond between the phosphate backbone and the deoxyribose nucleotides. In NHEJ, DNA Ligase IV, a specialized DNA ligase that forms a complex with the cofactor XRCC4, directly joins the two ends. To guide accurate repair, NHEJ relies on short homologous sequences called microhomologies present on the single-stranded tails of the DNA ends to be joined. If these overhangs are compatible, repair is usually accurate. NHEJ can also introduce mutations during repair. Loss of damaged nucleotides at the break site can lead to deletions, and joining of nonmatching termini forms translocations. NHEJ is especially important before the cell has replicated its DNA, since there is no template available for repair by homologous recombination. There are "backup" NHEJ pathways in higher eukaryotes. Besides its role as a genome caretaker, NHEJ is required for joining hairpin-capped double-strand breaks induced during V(D)J recombination, the process that generates diversity in B-cell and T-cell receptors in the vertebrate immune system. Homologous recombination requires the presence of an identical or nearly identical sequence to be used as a template for repair of the break. The enzymatic machinery responsible for this repair process is nearly identical to the machinery responsible for chromosomal crossover during meiosis. This pathway allows a damaged chromosome to be repaired using a sister chromatid (available in G2 after DNA replication) or a homologous chromosome as a template. DSBs caused by the replication machinery attempting to synthesize across a single-strand break or unrepaired lesion cause collapse of the replication fork and are typically repaired by recombination. Topoisomerases introduce both single- and double-strand breaks in the course of changing the DNA's state of supercoiling, which is especially common in regions near an
Notes made by kozi1989 email:kozi1989@yahoo.gr
30
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
open replication fork. Such breaks are not considered DNA damage because they are a natural intermediate in the topoisomerase biochemical mechanism and are immediately repaired by the enzymes that created them. Translesion synthesis Translesion synthesis (TLS) is a DNA damage tolerance process that allows the DNA replication machinery to replicate past DNA lesions such as thymine dimers or AP sites. It involves switching out regular DNA polymerases for specialized translesion polymerases (i.e. DNA polymerase IV or V, from the Y Polymerase family), often with larger active sites that can facilitate the insertion of bases opposite damaged nucleotides. The polymerase switching is thought to be mediated by, among other factors, the posttranslational modification of the replication processivity factor PCNA. Translesion synthesis polymerases often have low fidelity (high propensity to insert wrong bases) on undamaged templates relative to regular polymerases. However, many are extremely efficient at inserting correct bases opposite specific types of damage. For example, Pol mediates error free bypass of lesions induced by UV irradiation, whereas Pol introduces mutations at these sites. Pol is known to add the first adenine across the T^T photodimer using Watson -Crick base pairing and the second adenine will be added in it's syn conformation using Hoogsteen base pairing. From a cellular perspective, risking the introduction of point mutations during translesion synthesis may be preferable to resorting to more drastic mechanisms of DNA repair, which may cause gross chromosomal aberrations or cell death. In short, the process involves specialized polymerases either bypassing or repairing lesions at locations of stalled DNA replication. A bypass platform is provided to these polymerases by Proliferating cell nuclear antigen (PCNA). Under normal circumstances, PCNA bound to polymerases replicates the DNA. At a site of lesion, PCNA is ubiquitinated, or modified, by the RAD6/RAD18 proteins to provide a platform for the specialized polymerases to bypass the lesion and resume DNA replication. After translesion synthesis, extension is required. This extension can be carried out by a replicative polymerase if the TLS is error-free, as in the case of Pol , yet if TLS results in a mismatch, a specialized polymerase is needed to extend it; Pol . Pol is unique in that it can extend terminal mismatches, whereas more processive polymerases cannot. So when a lesion is encountered, the replication fork will stall, PCNA will switch from a processive polymerase to a TLS polymerase such as Pol to fix the lesion, then PCNA may switch to Pol to extend the mismatch, and last PCNA will switch to the processive polymerase to continue replication.
10. Molecular mechanisms of the basic genetic processes
Transcription. RNA processing Transcription is the process of creating a complementary RNA copy of a sequence of DNA. Both RNA and DNA are nucleic acids, which use base pairs of nucleotides as a complementary language that can be converted back and forth from DNA to RNA by the action of the correct enzymes. During transcription, a DNA sequence is read by RNA
Notes made by kozi1989 email:kozi1989@yahoo.gr
31
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
polymerase, which produces a complementary, antiparallel RNA strand. As opposed to DNA replication, transcription results in an RNA complement that includes uracil (U) in all instances where thymine (T) would have occurred in a DNA complement. Transcription can be explained easily in 4 or 5 steps, each moving like a wave along the DNA. 1) Helicase unwinds/"unzips" the DNA by breaking the hydrogen bonds between complementary nucleotides. 2) RNA nucleotides are paired with complementary DNA bases. 3) RNA sugar-phosphate backbone forms with assistance from RNA polymerase. 4) Hydrogen bonds of the untwisted RNA+DNA helix break, freeing the newly synthesized RNA strand. 5) If the cell has a nucleus, the RNA is further processed (addition of a 3' poly-A tail and a 5' cap) and exits through to the cytoplasm through the nuclear pore complex. Transcription is the first step leading to gene expression. The stretch of DNA transcribed into an RNA molecule is called a transcription unit and encodes at least one gene. If the gene transcribed encodes a protein, the result of transcription is messenger RNA (mRNA), which will then be used to create that protein via the process of translation. Alternatively, the transcribed gene may encode for either ribosomal RNA (rRNA) or transfer RNA (tRNA), other components of the protein-assembly process, or other ribozymes. A DNA transcription unit encoding for a protein contains not only the sequence that will eventually be directly translated into the protein (the coding sequence) but also regulatory sequences that direct and regulate the synthesis of that protein. The regulatory sequence before (upstream from) the coding sequence is called the five prime untranslated region (5'UTR), and the sequence following (downstream from) the coding sequence is called the three prime untranslated region (3'UTR). Transcription has some proofreading mechanisms, but they are fewer and less effective than the controls for copying DNA; therefore, transcription has a lower copying fidelity than DNA replication. As in DNA replication, DNA is read from 3' 5' during transcription . Meanwhile, the complementary RNA is created from the 5' 3' direction. This means its 5' end is created first in base pairing. Although DNA is arranged as two antiparallel strands in a double helix, only one of the two DNA strands, called the template strand, is used for transcription. This is because RNA is only single-stranded, as opposed to double-stranded DNA. The other DNA strand is called the coding strand, because its sequence is the same as the newly created RNA transcript (except for the substitution of uracil for thymine). The use of only the 3' 5' strand eliminates the need for the Okazaki fragments seen in DNA replication. Transcription is divided into 5 stages: pre-initiation, initiation, promoter clearance, elongation and termination. Pre-initiation
Notes made by kozi1989 email:kozi1989@yahoo.gr
32
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
In eukaryotes, RNA polymerase, and therefore the initiation of transcription, requires the presence of a core promoter sequence in the DNA. Promoters are regions of DNA that promote transcription and, in eukaryotes, are found at -30, -75, and -90 base pairs upstream from the start site of transcription. Core promoters are sequences within the promoter that are essential for transcription initiation. RNA polymerase is able to bind to core promoters in the presence of various specific transcription factors. The most common type of core promoter in eukaryotes is a short DNA sequence known as a TATA box, found 25-30 base pairs upstream from the start site of transcription.[citation needed] The TATA box, as a core promoter, is the binding site for a transcription factor known as TATA-binding protein (TBP), which is itself a subunit of another transcription factor, called Transcription Factor II D (TFIID). After TFIID binds to the TATA box via the TBP, five more transcription factors and RNA polymerase combine around the TATA box in a series of stages to form a preinitiation complex. One transcription factor, DNA helicase, has helicase activity and so is involved in the separating of opposing strands of double-stranded DNA to provide access to a single-stranded DNA template. However, only a low, or basal, rate of transcription is driven by the preinitiation complex alone. Other proteins known as activators and repressors, along with any associated coactivators or corepressors, are responsible for modulating transcription rate. Thus, preinitiation complex contains: 1) 2) 3) 4) 5) Core Promoter Sequence Transcription Factors DNA Helicase RNA Polymerase Activators and Repressors
The transcription preinitiation in archaea is, in essence, homologous to that of eukaryotes, but is much less complex.The archaeal preinitiation complex assembles at a TATA-box binding site; however, in archaea, this complex is composed of only RNA polymerase II, TBP, and TFB (the archaeal homologue of eukaryotic transcription factor II B (TFIIB)). Initiation In bacteria, transcription begins with the binding of RNA polymerase to the promoter in DNA. RNA polymerase is a core enzyme consisting of five subunits: 2 subunits, 1 subunit, 1 ' subunit, and 1 subunit. At the start of initiation, the core enzyme is associated with a sigma factor that aids in finding the appropriate -35 and -10 base pairs downstream of promoter sequences. Transcription initiation is more complex in eukaryotes. Eukaryotic RNA polymerase does not directly recognize the core promoter sequences. Instead, a collection of proteins called transcription factors mediate the binding of RNA polymerase and the initiation of transcription. Only after certain transcription factors are attached to the promoter does
Notes made by kozi1989 email:kozi1989@yahoo.gr
33
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
the RNA polymerase bind to it. The completed assembly of transcription factors and RNA polymerase bind to the promoter, forming a transcription initiation complex. Transcription in the archaea domain is similar to transcription in eukaryotes. Promoter clearance After the first bond is synthesized, the RNA polymerase must clear the promoter. During this time there is a tendency to release the RNA transcript and produce truncated transcripts. This is called abortive initiation and is common for both eukaryotes and prokaryotes. Abortive initiation continues to occur until the factor rearranges, resulting in the transcription elongation complex (which gives a 35 bp moving footprint). The factor is released before 80 nucleotides of mRNA are synthesized. Once the transcript reaches approximately 23 nucleotides, it no longer slips and elongation can occur. This, like most of the remainder of transcription, is an energy-dependent process, consuming adenosine triphosphate (ATP). Promoter clearance coincides with phosphorylation of serine 5 on the carboxy terminal domain of RNA Pol in eukaryotes, which is phosphorylated by TFIIH. Elongation One strand of the DNA, the template strand (or noncoding strand), is used as a template for RNA synthesis. As transcription proceeds, RNA polymerase traverses the template strand and uses base pairing complementarity with the DNA template to create an RNA copy. Although RNA polymerase traverses the template strand from 3' 5', the coding (non-template) strand and newly-formed RNA can also be used as reference points, so transcription can be described as occurring 5' 3'. This produces an RNA molecule from 5' 3', an exact copy of the coding strand (except that thymines are replaced with uracils, and the nucleotides are composed of a ribose (5-carbon) sugar where DNA has deoxyribose (one less oxygen atom) in its sugar-phosphate backbone). Unlike DNA replication, mRNA transcription can involve multiple RNA polymerases on a single DNA template and multiple rounds of transcription (amplification of particular mRNA), so many mRNA molecules can be rapidly produced from a single copy of a gene. Elongation also involves a proofreading mechanism that can replace incorrectly incorporated bases. In eukaryotes, this may correspond with short pauses during transcription that allow appropriate RNA editing factors to bind. These pauses may be intrinsic to the RNA polymerase or due to chromatin structure. Termination Bacteria use two different strategies for transcription termination. In Rhoindependent transcription termination, RNA transcription stops when the newly synthesized RNA molecule forms a G-C-rich hairpin loop followed by a run of Us. When the hairpin forms, the mechanical stress breaks the weak rU-dA bonds, now filling the DNA-RNA hybrid. This pulls the poly-U transcript out of the active site of the RNA polymerase, in effect,
Notes made by kozi1989 email:kozi1989@yahoo.gr
34
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
terminating transcription. In the "Rho-dependent" type of termination, a protein factor called "Rho" destabilizes the interaction between the template and the mRNA, thus releasing the newly synthesized mRNA from the elongation complex. Transcription termination in eukaryotes is less understood but involves cleavage of the new transcript followed by template-independent addition of As at its new 3' end, in a process called polyadenylation. Post-transcriptional modification is a process in cell biology by which, in eukaryotic cells, primary transcript RNA is converted into mature RNA. A notable example is the conversion of precursor messenger RNA into mature messenger RNA (mRNA) , which includes splicing and occurs prior to protein synthesis. This process is vital for the correct translation of the genomes of eukaryotes as the human primary RNA transcript that is produced as a result of transcription contains both exons, which are coding sections of the primary RNA transcript and introns, which are the non coding sections of the primary RNA transcript. The pre-mRNA molecule undergoes three main modifications. These modifications are 5' capping, 3' polyadenylation, and RNA splicing, which occur in the cell nucleus before the RNA is translated. 5' Processing Capping Capping of the pre-mRNA involves the addition of 7-methylguanosine (m7G) to the 5' end. To achieve this, the terminal 5' phosphate requires removal, which is done with the aid of a phosphatase enzyme. The enzyme guanosyl transferase then catalyses the reaction, which produces the diphosphate 5' end. The diphosphate 5' prime end then attacks the phosphorus atom of a GTP molecule in order to add the guanine residue in a 5'5' triphosphate link. The enzyme (guanine-N7-)-methyltransferase ("cap MTase") transfers a methyl group from S-adenosyl methionine to the guanine ring. This type of cap, with just the (m7G) in position is called a cap 0 structure. The ribose of the adjacent nucleotide may also be methylated to give a cap 1. Methylation of nucleotides downstream of the RNA molecule produce cap 2, cap 3 structures and so on. In these cases the methyl groups are added to the 2' OH groups of the ribose sugar. The cap protects the 5' end of the primary RNA transcript from attack by ribonucleases that have specificity to the 3'5' phosphodiester bonds. 3' Processing Cleavage and Polyadenylation The pre-mRNA processing at the 3' end of the RNA molecule involves cleavage of its 3' end and then the addition of about 200 adenine residues to form a poly(A) tail. The cleavage and adenylation reactions occur if a polyadenylation signal sequence (5'- AAUAAA3') is located near the 3' end of the pre-mRNA molecule, which is followed by another
Notes made by kozi1989 email:kozi1989@yahoo.gr
35
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
sequence, which is usually (5'-CA-3'). The second signal is the site of cleavage. A GU-rich sequence is also usually present further downstream on the pre-mRNA molecule. After the synthesis of the sequence elements, two multisubunit proteins called cleavage and polyadenylation specificity factor (CPSF) and cleavage stimulation factor (CStF) are transferred from RNA Polymerase II to the RNA molecule. The two factors bind to the sequence elements. A protein complex forms that contains additional cleavage factors and the enzyme Polyadenylate Polymerase (PAP). This complex cleaves the RNA between the polyadenylation sequence and the GU-rich sequence at the cleavage site marked by the (5'CA-3') sequences. Poly(A) polymerase then adds about 200 adenine units to the new 3' end of the RNA molecule using ATP as a precursor. As the poly(A) tails is synthesised, it binds multiple copies of poly(A) binding protein, which protects the 3'end from ribonuclease digestion. RNA splicing is the process by which introns, regions of RNA that do not code for protein, are removed from the pre-mRNA and the remaining exons connected to re-form a single continuous molecule. Although most RNA splicing occurs after the complete synthesis and end-capping of the pre-mRNA, transcripts with many exons can be spliced cotranscriptionally. The splicing reaction is catalyzed by a large protein complex called the spliceosome assembled from proteins and small nuclear RNA molecules that recognize splice sites in the pre-mRNA sequence. Many pre-mRNAs, including those encoding antibodies, can be spliced in multiple ways to produce different mature mRNAs that encode different protein sequences. This process is known as alternative splicing, and allows production of a large variety of proteins from a limited amount of DNA.
11. Molecular mechanisms of the basic genetic processes
Genetic code. Translation. Protein sorting into cellular organelles. The genetic code is the set of rules by which information encoded in genetic material (DNA or mRNA sequences) is translated into proteins (amino acid sequences) by living cells. The code defines how sequences of three nucleotides, called codons, specify which amino acid will be added next during protein synthesis. With some exceptions, a threenucleotide codon in a nucleic acid sequence specifies a single amino acid. Because the vast majority of genes are encoded with exactly the same code (see the RNA codon table), this particular code is often referred to as the canonical or standard genetic code, or simply the genetic code, though in fact there are many variant codes. For example, protein synthesis in human mitochondria relies on a genetic code that differs from the standard genetic code. Not all genetic information is stored using the genetic code. All organisms DNA contains regulatory sequences, intergenic segments, chromosomal structural areas, and other non-coding DNA that can contribute greatly to phenotype. Those elements operate under sets of rules that are distinct from the codon-to-amino acid paradigm underlying the genetic code. The genome of an organism is inscribed in DNA, or in the case of some viruses, RNA.
Notes made by kozi1989 email:kozi1989@yahoo.gr
36
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The portion of the genome that codes for a protein or an RNA is called a gene . Those genes that code for proteins are composed of tri-nucleotide units called codons, each coding for a single amino acid. Each nucleotide sub-unit consists of a phosphate, a deoxyribose sugar and one of the four nitrogenous nucleobases. The purine bases adenine (A) and guanine (G) are larger and consist of two aromatic rings. The pyrimidine bases cytosine (C) and thymine (T) are smaller and consist of only one aromatic ring. In the double-helix configuration, two strands of DNA are joined to each other by hydrogen bonds in an arrangement known as base pairing. These bonds almost always form between an adenine base on one strand and a thymine base on the other strand, or between a cytosine base on one strand and a guanine base on the other. This means that the number of A and T bases will be the same in a given double helix, as will the number of G and C bases.]:102117 In RNA, thymine (T) is replaced by uracil (U), and the deoxyribose is substituted by ribose. Each protein-coding gene is transcribed into a molecule of the related polymer RNA. In prokaryotes, this RNA functions as messenger RNA or mRNA; in eukaryotes, the transcript needs to be processed to produce a mature mRNA. The mRNA is, in turn, translated on the ribosome into an amino acid chain or polypeptide. The process of translation requires transfer RNAs specific for individual amino acids with the amino acids covalently attached to them, guanosine triphosphate as an energy source, and a number of translation factors. tRNAs have anticodons complementary to the codons in mRNA and can be "charged" covalently with amino acids at their 3' terminal CCA ends. Individual tRNAs are charged with specific amino acids by enzymes known as aminoacyl tRNA synthetases, which have high specificity for both their cognate amino acids and tRNAs. The high specificity of these enzymes is a major reason why the fidelity of protein translation is maintained. There are 4 = 64 different codon combinations possible with a triplet codon of three nucleotides; all 64 codons are assigned for either amino acids or stop signals during translation. If, for example, an RNA sequence, UUUAAACCC is considered and the reading frame starts with the first U (by convention, 5' to 3'), there are three codons, namely, UUU, AAA and CCC, each of which specifies one amino acid. This RNA sequence will be translated into an amino acid sequence, three amino acids long.:521539 A given amino acid may be encoded by between one and six different codon sequences. A comparison may be made with computer science, where the codon is similar to a word, which is the standard "chunk" for handling data (like one amino acid of a protein), and a nucleotide is similar to a bit, in that it is the smallest unit. A codon is defined by the initial nucleotide from which translation starts. For example, the string GGGAAACCC, if read from the first position, contains the codons GGG, AAA and CCC; and, if read from the second position, it contains the codons GGA and AAC; if read starting from the third position, GAA and ACC. Every sequence can thus be read in three reading frames, each of which will produce a different amino acid sequence (in the given example, Gly-Lys-Pro, Gly-Asn, or Glu-Thr, respectively). With double-stranded DNA there are six possible reading frames, three in the forward orientation on one strand and three
Notes made by kozi1989 email:kozi1989@yahoo.gr
37
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
reverse on the opposite strand. The actual frame in which a protein sequence is translated is defined by a start codon, usually the first AUG codon in the mRNA sequence. Start/stop codons Translation starts with a chain initiation codon (start codon). Unlike stop codons, the codon alone is not sufficient to begin the process. Nearby sequences (such as the ShineDalgarno sequence in E. coli) and initiation factors are also required to start translation. The most common start codon is AUG which is read as methionine or, in bacteria, as formylmethionine. Alternative start codons (depending on the organism), include "GUG" or "UUG"; these codons normally represent valine and leucine, respectively, but as a start codon, they are translated as methionine or formylmethionine. The three stop codons have been given names: UAG is amber, UGA is opal (sometimes also called umber), and UAA is ochre. Stop codons are also called "termination" or "nonsense" codons. They signal release of the nascent polypeptide from the ribosome because there is no cognate tRNA that has anticodons complementary to these stop signals, and so a release factor binds to the ribosome instead. Translation is the third stage of protein biosynthesis (part of the overall process of gene expression). In translation, messenger RNA (mRNA) produced by transcription is decoded by the ribosome to produce a specific amino acid chain, or polypeptide, that will later fold into an active protein. In Bacteria, translation occurs in the cell's cytoplasm, where the large and small subunits of the ribosome are located, and bind to the mRNA. In Eukaryotes, translation occurs across the membrane of the endoplasmic reticulum in a process called vectorial synthesis. The ribosome facilitates decoding by inducing the binding of tRNAs with complementary anticodon sequences to that of the mRNA. The tRNAs carry specific amino acids that are chained together into a polypeptide as the mRNA passes through and is "read" by the ribosome in a fashion reminiscent to that of a stock ticker and ticker tape. In many instances, the entire ribosome/mRNA complex will bind to the outer membrane of the rough endoplasmic reticulum and release the nascent protein polypeptide inside for later vesicle transport and secretion outside of the cell. Many types of transcribed RNA, such as transfer RNA, ribosomal RNA, and small nuclear RNA, do not undergo translation into proteins. Translation proceeds in four phases: activation, initiation, elongation and termination (all describing the growth of the amino acid chain, or polypeptide that is the product of translation). Amino acids are brought to ribosomes and assembled into proteins. In activation, the correct amino acid is covalently bonded to the correct transfer RNA (tRNA). The amino acid is joined by its carboxyl group to the 3' OH of the tRNA by an ester bond. When the tRNA has an amino acid linked to it, it is termed "charged". Initiation involves the small subunit of the ribosome binding to the 5' end of mRNA with the help of initiation factors (IF). Termination of the polypeptide happens when the A site of the
Notes made by kozi1989 email:kozi1989@yahoo.gr
38
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
ribosome faces a stop codon (UAA, UAG, or UGA). No tRNA can recognize or bind to this codon. Instead, the stop codon induces the binding of a release factor protein that prompts the disassembly of the entire ribosome/mRNA complex. The mRNA carries genetic information encoded as a ribonucleotide sequence from the chromosomes to the ribosomes. The ribonucleotides are "read" by translational machinery in a sequence of nucleotide triplets called codons. Each of those triplets codes for a specific amino acid. The ribosome molecules translate this code to a specific sequence of amino acids. The ribosome is a multisubunit structure containing rRNA and proteins. It is the "factory" where amino acids are assembled into proteins. tRNAs are small noncoding RNA chains (7493 nucleotides) that transport amino acids to the ribosome. tRNAs have a site for amino acid attachment, and a site called an anticodon. The anticodon is an RNA triplet complementary to the mRNA triplet that codes for their cargo amino acid. Aminoacyl tRNA synthetase (an enzyme) catalyzes the bonding between specific tRNAs and the amino acids that their anticodon sequences call for. The product of this reaction is an aminoacyl-tRNA molecule. This aminoacyl-tRNA travels inside the ribosome, where mRNA codons are matched through complementary base pairing to specific tRNA anticodons. The amino acids that the tRNAs carry are then used to assemble a protein. After the new amino acid is added to the chain, the energy provided by the hydrolysis of a GTP bound to the translocase EF-G (in prokaryotes) and eEF-2 (in eukaryotes) moves the ribosome down one codon towards the 3' end. The energy required for translation of proteins is significant. For a protein containing n amino acids, the number of high-energy Phosphate bonds required to translate it is 4n+1. The rate of translation varies. it is significantly higher in prokaryotic cells (up to 17-21 amino acid residues per second) than in eukaryotic cells (up to 6-9 amino acid residues per second). Protein targeting or protein sorting is the mechanism by which a cell transports proteins to the appropriate positions in the cell or outside of it. Sorting targets can be the inner space of an organelle, any of several interior membranes, the cell's outer membrane, or its exterior via secretion. This delivery process is carried out based on information contained in the protein itself. Correct sorting is crucial for the cell; errors can lead to diseases. Signal peptide Targeting signals are the pieces of information that enable the cellular transport machinery to correctly position a protein inside or outside the cell. This information is contained in the polypeptide chain or in the folded protein. The continuous stretch of amino acid residues in the chain that enables targeting are called signal peptides or targeting peptides. There are two types of targeting peptides, the presequences and the internal targeting peptides. The presequences of the targeting peptide are often found at the Nterminal extension and is composed of between 6-136 basic and hydrophobic amino acids.In case of peroxisomes the targeting sequence is on the C-terminal extension mostly. Other
Notes made by kozi1989 email:kozi1989@yahoo.gr
39
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
signals are composed by parts which are separate in the primary sequence. To function, these components have to come together on the protein surface by folding. They are called signal patches. In addition, protein modifications like glycosylations can induce targeting. Protein translocation Many proteins have a signal sequence, that is, a short amino acid sequence at one end that functions like a postal code for the target organelle. The translation of mRNA into protein by a ribosome takes place within the cytosol. If the synthesized proteins "belong" in a different organelle, they can be transported there in either of two ways, depending on the protein. Cotranslational translocation The N-terminal signal sequence of the protein is recognized by a signal recognition particle (SRP) while the protein is still being synthesized on the ribosome. The synthesis pauses while the ribosome-protein complex is transferred to a SRP receptor on the endoplasmic reticulum (ER), a membrane-enclosed organelle. There, the nascent protein is inserted into the Sec61 translocation complex (also known as the translocon) that passes through the ER membrane. The signal sequence is immediately cleaved from the polypeptide once it has been translocated into the ER by signal peptidase in secretory proteins. This signal sequence processing differs for some ER transmembrane proteins. Within the ER, the protein is first covered by a chaperone protein to protect it from the high concentration of other proteins in the ER, giving it time to fold correctly. Once folded, the protein is modified as needed (for example, by glycosylation), then transported to the Golgi apparatus for further processing and goes to its target organelles or is retained in the ER by various ER retention mechanisms. Posttranslational translocation Even though most proteins are cotranslationally translocated, some are translated in the cytosol and later transported to their destination. This occurs for proteins that go to a mitochondrion, a chloroplast, or a peroxisome (proteins that go to the latter have their signal sequence at the C terminus). Also, proteins targeted for the nucleus are translocated post-translation. They pass through the nuclear envelope via nuclear pores. Transmembrane proteins The amino acid chain of transmembrane proteins, which often are transmembrane receptors, passes through a membrane one or several times. They are inserted into the membrane by translocation, until the process is interrupted by a stop-transfer sequence, also called a membrane anchor sequence. These complex membrane proteins are at the moment mostly understood using the same model of targeting that has been developed for secretory proteins. However, many complex multi-transmembrane proteins contain structural aspects that do not fit the model. Seven transmembrane G-protein coupled
Notes made by kozi1989 email:kozi1989@yahoo.gr
40
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
receptors (which represent about 5% of the genome of humans) mostly do not have an amino-terminal signal sequence. In contrast to secretory proteins, the first transmembrane domain acts as the first signal sequence, which targets them to the ER membrane. This also results in the translocation of the amino terminus of the protein into the ER membrane lumen. This would seem to break the rule of "co-translational" translocation which has always held for mammalian proteins targeted to the ER. This has been demonstrated with opsin with in vitro experiments.
12. Molecular mechanisms of the basic genetic processes
Posttranslational modification of proteins. Senescence and degradation of proteins. Posttranslational modification (PTM) is the chemical modification of a protein after its translation. It is one of the later steps in protein biosynthesis, and thus gene expression, for many proteins. A protein (also called a polypeptide) is a chain of amino acids. During protein synthesis, 20 different amino acids can be incorporated to become a protein. After translation, the posttranslational modification of amino acids extends the range of functions of the protein by attaching it to other biochemical functional groups such as acetate, phosphate, various lipids and carbohydrates, by changing the chemical nature of an amino acid (e.g. citrullination) or by making structural changes, like the formation of disulfide bridges. Also, enzymes may remove amino acids from the amino end of the protein, or cut the peptide chain in the middle. For instance, the peptide hormone insulin is cut twice after disulfide bonds are formed, and a propeptide is removed from the middle of the chain; the resulting protein consists of two polypeptide chains connected by disulfide bonds. Also, most nascent polypeptides start with the amino acid methionine because the "start" codon on mRNA also codes for this amino acid. This amino acid is usually taken off during posttranslational modification. Other modifications, like phosphorylation, are part of common mechanisms for controlling the behavior of a protein, for instance activating or inactivating an enzyme. Proteolysis is the directed degradation (digestion) of proteins by cellular enzymes called proteases or by intramolecular digestion. Purposes Proteolysis is used by the cell for several purposes. They include: 1) Removal of N-terminal methionine residues after translation. 2) Removal of the signal sequence of peptides after their transport through a membrane 3) Separation of viral proteins that were translated from a polycistronic mRNA 4) Digestion of proteins from foods as a source of amino acids
Notes made by kozi1989 email:kozi1989@yahoo.gr
41
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
5) Conversion of predecessor-proteins (proenzymes, zymogens, prehormones) into their final structures. 6) Degradation of unneeded or damaged proteins for example cyclins at different stages of the cell cycle. Biodegradation is the chemical dissolution of materials by bacteria or other biological means. The term is often used in relation to ecology, waste management, biomedicine, and the natural environment (bioremediation) and is now commonly associated with environmentally friendly products that are capable of decomposing back into natural elements. Organic material can be degraded aerobically with oxygen, or anaerobically, without oxygen. A term related to biodegradation is biomineralisation, in which organic matter is converted into minerals. Biosurfactant, an extracellular surfactant secreted by microorganisms, enhances the biodegradation process.
13. Organization of the prokaryotic genome. Regulation of gene expression in
procaryotic cells The prokaryotes - bacteria and archaea - typically have a single circular chromosome. Most bacteria have a single circular chromosome.Prokaryotes chromosomes have less sequence-based structure than eukaryotes. Bacteria typically have a single point (the origin of replication) from which replication starts, while some archaea contain multiple replication origins. The genes in prokaryotes are often organized in operons, and do not contain introns, unlike eukaryotes. 1) 2) 3) 4) Usually circular Smaller than eukaryotic genome Found in the nucleoid region Less elaborately structured and folded
Compacting the DNA involves supercoiling, or further twisting the twisted chromosome. The chromosomes fifty or so DNA domains are held together by a scaffold of RNA and protein, and the entire nucleoid is attached to the cell membrane. This membrane attachment aids in the segregation of the chromosomes after they replicate in preparation for cell division. Bacteria lack the histone proteins that are found bound to the DNA and that form the nucleosomes of eukaryotic chromosomes. However, it is believed that polyamines (organic molecules with multiple _NH2 amine groups) such as spermidine, as well as some basic proteins, aid in compacting the bacterial chromosome. These basic proteins have a net positive charge that bind them to the negative charge of the phosphates in the DNA backbone. Replication of the circular chromosome begins at a single point, called OriC, and proceeds in both directions around the circle, until the two replicationforks meet up. The result is two identical loops. Replication takes approximately forty minutes.
Notes made by kozi1989 email:kozi1989@yahoo.gr
42
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Regulation of gene expression (or gene regulation) includes the processes that cells and viruses use to regulate the way that the information in genes is turned into gene products. Although a functional gene product may be an RNA or a protein, the majority of known mechanisms regulate protein coding genes. Any step of the gene's expression may be modulated, from DNA-RNA transcription to the post-translational modification of a protein. Gene regulation is essential for viruses, prokaryotes and eukaryotes as it increases the versatility and adaptability of an organism by allowing the cell to express protein when needed. Gene expression in many bacteria is regulated through the existence of operons. An operon is a cluster of genes whose protein products have related functions. For instance, the lac operon includes one gene that transports lactose sugar into the cell and another that breaks it into two parts. These genes are under the control of the same promoter, and so are transcribed and translated into protein at the same time. RNA polymerase can only reach the promoter if a repressor is not blocking it; the lac repressor is dislodged by lactose. In this way, the bacterium uses its resources to make lactose-digesting enzymes only when lactose is available. Other genes are expressed constantly at low levels; their protein products are required for housekeeping functions such as membrane synthesis and DNA repair. One such enzyme is DNA gyrase, which relieves strain in the double helix during replication and repair. DNA gyrase is the target for the antibiotic ciproflaxin (sold under the name Cipro), effective against Bacillus anthracis, the cause of anthrax. Since eukaryotes do not have this type of DNA gyrase, they are not harmed by the action of this antibiotic. As in eukaryotes, translation (protein synthesis) occurs on the ribosome. Without a nucleus to exclude it, the ribosome can attach to the messengerRNA even while the RNA is still attached to the DNA. Multiple ribosomes can attach to the same mRNA, making multiple copies of the same protein. The ribosomes of eubacteria are similar in structure to those in eukaryotes and archaea, but differ in molecular detail. This has two important consequences.
14. Organization of the eukaryotic genome. Levels of regulation of gene expression in
eukaryotic cells Molecular biology of the gene. Types of DNA sequences. DNA-binding proteins The vast majority of living organisms encode their genes in long strands of DNA. DNA (deoxyribonucleic acid) consists of a chain made from four types of nucleotide subunits, each composed of: a five-carbon sugar (2'-deoxyribose), a phosphate group, and one of the four bases adenine, cytosine, guanine, and thymine. The most common form of DNA in a cell is in a double helix structure, in which two individual DNA strands twist around each other in a right-handed spiral. In this structure, the base pairing rules specify that guanine pairs with cytosine and adenine pairs with thymine. The base pairing between guanine and cytosine forms three hydrogen bonds, whereas the base pairing between adenine and thymine forms two hydrogen bonds. The two strands in a double helix must therefore be complementary, that is, their bases must align such that the adenines of one strand are
Notes made by kozi1989 email:kozi1989@yahoo.gr
43
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
paired with the thymines of the other strand, and so on. Due to the chemical composition of the pentose residues of the bases, DNA strands have directionality. One end of a DNA polymer contains an exposed hydroxyl group on the deoxyribose; this is known as the 3' end of the molecule. The other end contains an exposed phosphate group; this is the 5' end. The directionality of DNA is vitally important to many cellular processes, since double helices are necessarily directional (a strand running 5'-3' pairs with a complementary strand running 3'-5'), and processes such as DNA replication occur in only one direction. All nucleic acid synthesis in a cell occurs in the 5'-3' direction, because new monomers are added via a dehydration reaction that uses the exposed 3' hydroxyl as a nucleophile. The expression of genes encoded in DNA begins by transcribing the gene into RNA, a second type of nucleic acid that is very similar to DNA, but whose monomers contain the sugar ribose rather than deoxyribose. RNA also contains the base uracil in place of thymine. RNA molecules are less stable than DNA and are typically single-stranded. Genes that encode proteins are composed of a series of three-nucleotide sequences called codons, which serve as the words in the genetic language. The genetic code specifies the correspondence during protein translation between codons and amino acids. The genetic code is nearly the same for all known organisms. All genes have regulatory regions in addition to regions that explicitly code for a protein or RNA product. A regulatory region shared by almost all genes is known as the promoter, which provides a position that is recognized by the transcription machinery when a gene is about to be transcribed and expressed. A gene can have more than one promoter, resulting in RNAs that differ in how far they extend in the 5' end. Although promoter regions have a consensus sequence that is the most common sequence at this position, some genes have "strong" promoters that bind the transcription machinery well, and others have "weak" promoters that bind poorly. These weak promoters usually permit a lower rate of transcription than the strong promoters, because the transcription machinery binds to them and initiates transcription less frequently. Other possible regulatory regions include enhancers, which can compensate for a weak promoter. Most regulatory regions are "upstream"that is, before or toward the 5' end of the transcription initiation site. Eukaryotic promoter regions are much more complex and difficult to identify than prokaryotic promoters. Many prokaryotic genes are organized into operons, or groups of genes whose products have related functions and which are transcribed as a unit. By contrast, eukaryotic genes are transcribed only one at a time, but may include long stretches of DNA called introns which are transcribed but never translated into protein (they are spliced out before translation). Splicing can also occur in prokaryotic genes, but is less common than in eukaryotes. Eukaryotes (cells with nuclei such as plants, yeast, and animals) possess multiple large linear chromosomes contained in the cell's nucleus. Each chromosome has one centromere, with one or two arms projecting from the centromere, although under most
Notes made by kozi1989 email:kozi1989@yahoo.gr
44
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
circumstances these arms are not visible as much. In addition most eukaryotes have a small circular mitochondrial genome, and some eukaryotes may have additional small circular or linear cytoplasmic chromosomes. In the nuclear chromosomes of eukaryotes, the uncondensed DNA exists in a semiordered structure, where it is wrapped around histones (structural proteins), forming a composite material called chromatin. 1) Complexed with a large amount of protein to form chromatin 2) Highly extended and tangled during interphase 3) Found in the nucleus Regulation can occur at any point in this pathway; specifically, it occurs at the levels of transcription, RNA processing, mRNA lifetime (longevity), and translation. The organizational structure of an eukaryotic cell determines the mode of gene regulation : 1) Chromatin packaging into nucleosomes and other organized structures possible control at the chromatin structure level 2) Compartmentalization of the cell need of internal signaling system to communicate between different compartments 3) Multicellular organism need of intercellular communication system 4) Differentiation of a totipotent cell into different cell types during body formation spatial and temporal regulation Gene expression must be controlled on a long-term basis during cellular differentiation, the divergence in form and function as cells in a multicellular organism specialize. A typical human cell probably expresses about 20% of its genes at any given time. Highly specialized cells, such as nerves or muscles, express only a tiny fraction of their genes. Although all the cells in an organism contain an identical genome, the subset of genes expressed in the cells of each type is unique. The differences between cell types are due to differential gene expression, the expression of different genes by cells with the same genome. The genomes of eukaryotes may contain tens of thousands of genes. For quite a few species, only a small amount of the DNA1.5% in humanscodes for protein. Of the remaining DNA, a very small fraction consists of genes for rRNA and tRNA. Most of the rest of the DNA seems to be largely noncoding, although researchers have found that a significant amount of it is transcribed into RNAs of unknown function.
Notes made by kozi1989 email:kozi1989@yahoo.gr
45
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Problems with gene expression and control can lead to imbalance and diseases, including cancers. Our understanding of the mechanisms controlling gene expression in eukaryotes has been enhanced by new research methods, including advances in DNA technology. In all organisms, the expression of specific genes is most commonly regulated at transcription, often in response to signals coming from outside the cell. The term gene expression is often equated with transcription. With their greater complexity, eukaryotes have opportunities for controlling gene expression at additional stages. Each stage in the entire process of gene expression provides a potential control point where gene expression can be turned on or off, sped up or slowed down. A web of control connects different genes and their products. These levels of control include chromatin packing, transcription, RNA processing, translation, and various alterations to the protein product. Control levels of gene expression Control at the level of chromatin and genome structure Control at the level of transcription initiation Control at the level of post-transcription initiation including transcription elongation, mRNA stability, alternative splicing Translational control Post-translational control
After transcription, the RNA must be processed before it can be translated. As described elsewhere, RNA processing involves addition of a 5' cap, addition of a 3' poly (A) tail, and removal of introns. This processing represents another level of regulation of gene expression, particularly in regard to splicing out of introns. Regulation can be of two types: whether an RNA gets processed; and which exons are retained in the mRNA. The first type of regulation can determine whether or not an mRNA gets translated. If an RNA is not processed, it will not be transported out of the nucleus, and will not be translated. The second type of regulation can affect the function of the protein produced . Some genes have exons that can be exchanged in a process known as exon shuffling. Regulation of RNA Longevity Imagine two mRNA molecules: one lasts for five minutes in the cytoplasm before being degraded, while the other one manages to linger for an hour before being degraded. If both are translated continually while they exist, it is obvious that more of the second polypeptide
Notes made by kozi1989 email:kozi1989@yahoo.gr
46
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
will be produced than the first. This is the principle behind regulation of RNA longevity. mRNAs from different genes have their approximate lifespan encoded in them; this serves to help regulate how much of each polypeptide is produced. The information for lifespan is found in the 3' UTR. The sequence AUUUA, when found in the 3' UTR, is a signal for early degradation (and therefore short lifetime). The more times the sequence is present, the shorter the lifespan of the mRNA. Because it is encoded in the nucleotide sequence, this is a set property of each different mRNA; the longevity of an mRNA can't be varied. Regulation of Translation Whether or not an mRNA molecule is translated can be regulated as well. The various mechanisms of translational regulation are incompletely understood, but there are many documented examples (particularly in embryonic development) of mRNA molecules that are present routinely, but are only translated under certain circumstances. For example, many animals sequester large amounts of mRNA in their eggs, and those mRNA molecules are not translated unless the egg is fertilized. Regulation of Transcription Whether or not a gene is transcribed is the major way that gene expression is regulated in eukaryotes, as it was in prokaryotes. There are some major differences between transcriptional regulation in prokaryotes and eukaryotes. For one thing, because of the complexity of eukaryotic patterns of gene expression, each eukaryotic gene needs its own promoter. In other words, eukaryotic genes are not organized into operons. Another difference is that prokaryotic genes are regulated primarily by repressors. Although repressors occasionally play a role in eukaryotes, eukaryotic genes are primarily regulated by transcriptional activators. These activators are transcription factors. The sequence or primary structure of a nucleic acid is the composition of atoms that make up the nucleic acid and the chemical bonds that those atoms. Because nucleic acids, such as DNA and RNA, are unbranched polymers, this specification is equivalent to specifying the sequence of nucleotides that comprise the molecule. This sequence is written as a succession of letters representing a real or hypothetical nucleic acid molecule or strand. By convention, the primary structure of a DNA or RNA molecule is reported from the 5' end to the 3' end. The sequence has capacity to represent information. Biological DNA represents the information which directs the functions of a living thing. In that context, the term genetic sequence is often used. Sequences can be read from the biological raw material through DNA sequencing methods. Nucleic acids also have a secondary structure and tertiary structure. Primary structure is sometimes mistakenly referred to as primary sequence. Conversely, there is no parallel concept of secondary or tertiary sequence.
Notes made by kozi1989 email:kozi1989@yahoo.gr
47
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
DNA-binding proteins are proteins that are composed of DNA-binding domains and thus have a specific or general affinity for either single or double stranded DNA. Sequencespecific DNA-binding proteins generally interact with the major groove of B-DNA, because it exposes more functional groups that identify a base pair. However there are some known minor groove DNA-binding ligands such as Netropsin, Distamycin, Hoechst 33258, Pentamidine and others. DNA-binding proteins include transcription factors which modulate the process of transcription, various polymerases, nucleases which cleave DNA molecules, and histones which are involved in chromosome packaging in the cell nucleus. DNA-binding proteins can incorporate such domains as the zinc finger, the helix-turn-helix, and the leucine zipper (among many others) that facilitate binding to nucleic acid. Structural proteins that bind DNA are well-understood examples of non-specific DNA-protein interactions. Within chromosomes, DNA is held in complexes with structural proteins. These proteins organize the DNA into a compact structure called chromatin. In eukaryotes this structure involves DNA binding to a complex of small basic proteins called histones, while in prokaryotes multiple types of proteins are involved. The histones form a disk-shaped complex called a nucleosome, which contains two complete turns of doublestranded DNA wrapped around its surface. These non-specific interactions are formed through basic residues in the histones making ionic bonds to the acidic sugar-phosphate backbone of the DNA, and are therefore largely independent of the base sequence.[10] Chemical modifications of these basic amino acid residues include methylation, phosphorylation and acetylation.[11] These chemical changes alter the strength of the interaction between the DNA and the histones, making the DNA more or less accessible to transcription factors and changing the rate of transcription.[12] Other non-specific DNAbinding proteins in chromatin include the high-mobility group proteins, which bind to bent or distorted DNA.[13] These proteins are important in bending arrays of nucleosomes and arranging them into the larger structures that make up chromosomes. A distinct group of DNA-binding proteins are the DNA-binding proteins that specifically bind single-stranded DNA. In humans, replication protein A is the bestunderstood member of this family and is used in processes where the double helix is separated, including DNA replication, recombination and DNA repair.[15] These binding proteins seem to stabilize single-stranded DNA and protect it from forming stem-loops or being degraded by nucleases.
15. Submicroscopic structure of chromosomes. Cytoplasmic heredity
In eukaryotes, nuclear chromosomes are packaged by proteins into a condensed structure called chromatin. This allows the very long DNA molecules to fit into the cell nucleus. The structure of chromosomes and chromatin varies through the cell cycle. Chromosomes are the essential unit for cellular division and must be replicated, divided, and passed successfully to their daughter cells so as to ensure the genetic diversity and survival of their progeny. Chromosomes may exist as either duplicated or unduplicated.
Notes made by kozi1989 email:kozi1989@yahoo.gr
48
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Unduplicated chromosomes are single linear strands, whereas duplicated chromosomes (copied during synthesis phase) contain two copies joined by a centromere. Extranuclear inheritance (also known as cytoplasmic inheritance) is a form of nonMendelian inheritance first discovered by Carl Correns in 1908. Cytoplasmic heredity is when the DNA is present in the cell outside of the nucleus. Mitochondrial DNA, and chloroplasts are the two organelles where you can find DNA outside of the nucleus. Mitochondrial DNA contains 37 genes,13 of which are involved in oxidative phosphoylation. Oxidative phospholation is process that uses oxygen and simple sugar to create ATP. The other genes provide for making tRNAs and rRNAs. Mitochondrial DNA is typically a circular structure with genes and regulatory regions. Mostly all mitochondrial DNA are inherited from the mothers ovum. According to the endosymbiont theory, mitochondria and chloroplasts were once free living organisms that were each taken up by a eukaryotic cell .[5] Over time, mitochondria and chloroplasts formed a symbiotic relationship with their eukaryotic hosts. Although the transfer of a number of genes from these organelles to the nucleus prevents them from living independently, each still possesses genetic material in the form of double stranded DNA. It is the transmission of this organellar DNA that is responsible for the phenomenon of extranuclear inheritance. Both chloroplasts and mitochondria are present in the cytoplasm of maternal gametes only. Paternal gametes (sperm for example) do not have cytoplasmic mitochondria. Thus, the phenotype of traits linked to genes found in either chloroplasts or mitochondria are determined exclusively by the maternal parent.
16. Microscopic structure of chromosomes. Human karyotype. Medhods of
karyotyping and chromosomal analysis. Evolution of karyotype karyotype is the number and appearance of chromosomes in the nucleus of a eukaryotic cell. The term is also used for the complete set of chromosomes in a species, or an individual organism. Karyotypes describe the number of chromosomes, and what they look like under a light microscope. Attention is paid to their length, the position of the centromeres, banding pattern, any differences between the sex chromosomes, and any other physical characteristics. The preparation and study of karyotypes is part of cytogenetics. The study of whole sets of chromosomes is sometimes known as karyology. The chromosomes are depicted (by rearranging a microphotograph) in a standard format known as a karyogram or idiogram: in pairs, ordered by size and position of centromere for chromosomes of the same size. The basic number of chromosomes in the somatic cells of an individual or a species is called the somatic number and is designated 2n. Thus, in humans 2n = 46. In the germline (the sex cells) the chromosome number is n (humans: n = 23).
Notes made by kozi1989 email:kozi1989@yahoo.gr
49
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
So, in normal diploid organisms, autosomal chromosomes are present in two copies. There may, or may not, be sex chromosomes. Polyploid cells have multiple copies of chromosomes and haploid cells have single copies. Karyotypes can be used for many purposes; such as, to study chromosomal aberrations, cellular function, taxonomic relationships, and to gather information about past evolutionary events. Although the replication and transcription of DNA is highly standardized in eukaryotes, the same cannot be said for their karyotypes, which are highly variable. There is variation between species in chromosome number, and in detailed organization, despite their construction from the same macromolecules. This variation provides the basis for a range of studies in evolutionary cytology. Routine chromosome analysis (Karyotyping) refers to analysis of metaphase chromosomes which have been banded using trypsin followed by Giemsa, Leishmanns, or a mixture of the two. This creates unique banding patterns on the chromosomes. The molecular mechanism and reason for these patterns is unknown, although it likely related to replication timing and chromatin packing. Cytogenetics employs several techniques to visualize different aspects of chromosomes: G-banding is obtained with Giemsa stain following digestion of chromosomes with trypsin. It yields a series of lightly and darkly stained bands - the dark regions tend to be heterochromatic, late-replicating and AT rich. The light regions tend to be euchromatic, early-replicating and GC rich. This method will normally produce 300-400 bands in a normal, human genome. R-banding is the reverse of G-banding (the R stands for "reverse"). The dark regions are euchromatic (guanine-cytosine rich regions) and the bright regions are heterochromatic (thymine-adenine rich regions). C-banding: Giemsa binds to constitutive heterochromatin, so it stains centromeres. Q-banding is a fluorescent pattern obtained using quinacrine for staining. The pattern of bands is very similar to that seen in G-banding. T-banding: visualize telomeres. Silver staining: Silver nitrate stains the nucleolar organization region-associated protein. This yields a dark region where the silver is deposited, denoting the activity of rRNA genes within the NOR. Classic karyotype cytogenetics In the "classic" (depicted) karyotype, a dye, often Giemsa (G-banding), less frequently Quinacrine, is used to stain bands on the chromosomes. Giemsa is specific for the phosphate groups of DNA. Quinacrine binds to the adenine-thymine-rich regions. Each chromosome has a characteristic banding pattern that helps to identify them; both chromosomes in a pair will have the same banding pattern.
Notes made by kozi1989 email:kozi1989@yahoo.gr
50
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Karyotypes are arranged with the short arm of the chromosome on top, and the long arm on the bottom. Some karyotypes call the short and long arms p and q, respectively. In addition, the differently stained regions and sub-regions are given numerical designations from proximal to distal on the chromosome arms. For example, Cri du chat syndrome involves a deletion on the short arm of chromosome 5. It is written as 46,XX,5p-. The critical region for this syndrome is deletion of 15.2, which is written as 46,XX,del. Spectral karyogram of a human female Spectral karyotyping is a molecular cytogenetic technique used to simultaneously visualize all the pairs of chromosomes in an organism in different colors. Fluorescently labeled probes for each chromosome are made by labeling chromosome-specific DNA with different fluorophores. Because there are a limited number of spectrally-distinct fluorophores, a combinatorial labeling method is used to generate many different colors. Spectral differences generated by combinatorial labeling are captured and analyzed by using an interferometer attached to a fluorescence microscope. Image processing software then assigns a pseudo color to each spectrally different combination, allowing the visualization of the individually colored chromosomes. Spectral human karyotype This technique is used to identify structural chromosome aberrations in cancer cells and other disease conditions when Giemsa banding or other techniques are not accurate enough. Digital karyotyping Digital karyotyping is a technique used to quantify the DNA copy number on a genomic scale. Short sequences of DNA from specific loci all over the genome are isolated and enumerated. This method is also known as virtual karyotyping. Analysis of banded chromosomes is done at a microscope by a clinical laboratory specialist in cytogenetics (CLSp(CG)). Generally 20 cells are analyzed which is enough to rule out mosaicism to an acceptable level. The results are summarized and given to a boardcertified cytogeneticist for review, and to write an interpretation taking into account the patients previous history and other clinical findings. Changes during development Instead of the usual gene repression, some organisms go in for large-scale elimination of heterochromatin, or other kinds of visible adjustment to the karyotype. Chromosome elimination. In some species, as in many sciarid flies, entire chromosomes are eliminated during development. Chromatin diminution (founding father: Theodor Boveri). In this process, found in some copepods and roundworms such as Ascaris suum, portions of the chromosomes are
Notes made by kozi1989 email:kozi1989@yahoo.gr
51
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
cast away in particular cells. This process is a carefully organised genome rearrangement where new telomeres are constructed and certain heterochromatin regions are lost. In A. suum, all the somatic cell precursors undergo chromatin diminution. X-inactivation. The inactivation of one X chromosome takes place during the early development of mammals (see Barr body and dosage compensation). In placental mammals, the inactivation is random as between the two Xs; thus the mammalian female is a mosaic in respect of her X chromosomes. In marsupials it is always the paternal X which is inactivated. In human females some 15% of somatic cells escape inactivation. Number of chromosomes in a set A spectacular example of variability between closely related species is the muntjac, which was investigated by Kurt Benirschke and his colleague Doris Wurster. The diploid number of the Chinese muntjac, Muntiacus reevesi, was found to be 46, all telocentric. When they looked at the karyotype of the closely related Indian muntjac, Muntiacus muntjak, they were astonished to find it had female = 6, male = 7 chromosomes. The number of chromosomes in the karyotype between (relatively) unrelated species is hugely variable. The low record is held by the nematode Parascaris univalens, where the haploid n = 1; the high record would be somewhere amongst the ferns, with the Adder's Tongue Fern Ophioglossum ahead with an average of 1262 chromosomes.[26] Top score for animals might be the shortnose sturgeon Acipenser brevirostrum at a mere 372 chromosomes.[27] The existence of supernumerary or B chromosomes means that chromosome number can vary even within one interbreeding population; and aneuploids are another example, though in this case they would not be regarded as normal members of the population. Fundamental number The fundamental number, FN, of a karyotype is the number of visible major chromosomal arms per set of chromosomes. Thus, FN 2n, the difference depending on the number of chromosomes considered single-armed (acrocentric or telocentric) present. Humans have FN = 82,[30] due to the presence of five acrocentric chromosome pairs (13, 14, 15, 21 and 22). Ploidy Ploidy is the number of complete sets of chromosomes in a cell. Polyploidy, where there are more than two sets of homologous chromosomes in the cells, occurs mainly in plants. It has been of major significance in plant evolution according to Stebbins. The proportion of flowering plants which are polyploid was estimated by Stebbins to be 30-35%, but in grasses the average is much higher, about 70%. Polyploidy in lower plants (ferns, horsetails and psilotales) is also common, and some species of ferns have reached levels of polyploidy far in excess of the highest levels known in flowering plants. Polyploidy in animals is much less common, but it has been significant in
Notes made by kozi1989 email:kozi1989@yahoo.gr
52
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
some groups. Polyploid series in related species which consist entirely of multiples of a single basic number are known as euploid. Haplo-diploidy, where one sex is diploid, and the other haploid. It is a common arrangement in the Hymenoptera, and in some other groups. Endopolyploidy occurs when in adult differentiated tissues the cells have ceased to divide by mitosis, but the nuclei contain more than the original somatic number of chromosomes. In the endocycle (endomitosis or endoreduplication) chromosomes in a 'resting' nucleus undergo reduplication, the daughter chromosomes separating from each other inside an intact nuclear membrane. In many instances, endopolyploid nuclei contain tens of thousands of chromosomes (which cannot be exactly counted). The cells do not always contain exact multiples (powers of two), which is why the simple definition 'an increase in the number of chromosome sets caused by replication without cell division' is not quite accurate. Aneuploidy Aneuploidy is the condition in which the chromosome number in the cells is not the typical number for the species. This would give rise to a chromosome abnormality such as an extra chromosome or one or more chromosomes lost. Abnormalities in chromosome number usually cause a defect in development. Down syndrome and Turner syndrome are examples of this. Aneuploidy may also occur within a group of closely related species. Classic examples in plants are the genus Crepis, where the gametic (= haploid) numbers form the series x = 3, 4, 5, 6, and 7; and Crocus, where every number from x = 3 to x = 15 is represented by at least one species. Evidence of various kinds shows that that trends of evolution have gone in different directions in different groups. Closer to home, the great apes have 24x2 chromosomes whereas humans have 23x2. Human chromosome 2 was formed by a merger of ancestral chromosomes, reducing the number. Chromosomal polymorphism Some animal species are polymorphic for chromosome fusions or dissociations. When this happens, the chromosome number is variable from one individual to another. Well-researched examples are the ladybird beetle Chilocorus stigma, some mantids of the genus Ameles, the European shrew Sorex araneus. There is some evidence from the case of the mollusc Thais lapillus (the dog whelk) on the Brittany coast, that the two chromosome morphs are adapted to different habitats.
17. Laws of heredity
Allelic form of the genes An allele is one of two or more forms of a gene. Sometimes, different alleles can result in different traits, such as color. Other times, different alleles will have the same result in the expression of a gene.
Notes made by kozi1989 email:kozi1989@yahoo.gr
53
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Most multicellular organisms have two sets of chromosomes, that is, they are diploid. These chromosomes are referred to as homologous chromosomes. Diploid organisms have one copy of each gene (and one allele) on each chromosome. If both alleles are the same, they are homozygotes. If the alleles are different, they are heterozygotes. A population or species of organisms typically includes multiple alleles at each locus among various individuals. Allelic variation at a locus is measurable as the number of alleles (polymorphism) present, or the proportion of heterozygotes in the population. For example, at the gene locus for ABO blood type proteins in humans, classical genetics recognizes three alleles, IA, IB, and IO, that determines compatibility of blood transfusions. Any individual has one of six possible genotypes (AA, AO, BB, BO, AB, and OO) that produce one of four possible phenotypes: "A" (produced by AA homozygous and AO heterozygous genotypes), "B" (produced by BB homozygous and BO heterozygous genotypes), "AB" heterozygotes, and "O" homozygotes. It is now known that each of the A, B, and O alleles is actually a class of multiple alleles with different DNA sequences that produce proteins with identical properties: more than 70 alleles are known at the ABO locus. An individual with "Type A" blood may be a AO heterozygote, an AA homozygote, or an A'A heterozygote with two different 'A' alleles. The word "allele" is a short form of allelomorph ('other form'), which was used in the early days of genetics to describe variant forms of a gene detected as different phenotypes. It derives from the Greek word , allelos, meaning "each other"
18. Laws of heredity
Interactions between genes. Penetrance and expressivity Penetrance in genetics is the proportion of individuals carrying a particular variation of a gene (allele or genotype) that also express an associated trait (phenotype). In medical genetics, the penetrance of a disease-causing mutation is the proportion of individuals with the mutation who exhibit clinical symptoms. For example, if a mutation in the gene responsible for a particular autosomal dominant disorder has 95% penetrance, then 95% of those with the mutation will develop the disease, while 5% will not. Common examples used to show degrees of penetrance are often highly penetrant. There are several reasons for this: Highly penetrant alleles, and highly heritable symptoms, are easier to demonstrate, because if the allele is present, the phenotype is generally expressed. Mendelian genetic concepts such as recessiveness, dominance, and co-dominance are fairly simple additions to this principle. Alleles which are highly penetrant are more likely to be noticed by clinicians and geneticists, and alleles for symptoms which are highly heritable are more likely to be inferred to exist, and then are more easily tracked down. Penetrance only considers whether individuals express the trait or not. 1) Complete penetrance. The allele is said to have complete penetrance if all individuals who have the disease-causing mutation have clinical symptoms of the disease.
Notes made by kozi1989 email:kozi1989@yahoo.gr
54
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
2) Highly penetrant. If an allele is highly penetrant, then the trait it produces will almost always be apparent in an individual carrying the allele. 3) ncomplete penetrance or reduced penetrance. Penetrance is said to be reduced or incomplete when some individuals fail to express the trait, even though they carry the allele. 4) Low penetrance. An allele with low penetrance will only sometimes produce the symptom or trait with which it has been associated at a detectable level. In cases of low penetrance, it is difficult to distinguish environmental from genetic factors. Expressivity is a term used in genetics to refer to variations in a phenotype among individuals carrying a particular genotype. The term can be used to characterize qualitatively or quantitatively the extent of phenotypic variation given a particular genotype. The term is analogous to the severity of a condition in clinical medicine. For example, the amount of blood ejected from the pumping heart with each contraction can be quantitated by echocardiography and is called the ejection fraction. If a specific genotype is associated with the development of congestive heart failure, the expressivity would be represented by the range of ejection fractions seen in patients that have that genotype. As a more qualitative example, the "blue" gene might have an expressivity of 25% for individuals that express the "blue" gene and appear light blue, and 75% for individuals that express the "blue" gene and appear dark blue. Expressivity is measured only when there is 100% penetrance. This differs from penetrance, which refers to the likelihood of the gene generating its associated phenotype at all, as determined by the proportion of individuals with a given genotype who also possess the associated phenotype. In contrast, expressivity refers to the influence of an expressed gene at the level of particular individuals. Variable expressivity occurs when a phenotype is expressed to a different degree among individuals with the same genotype. For example, individuals with the same allele for a gene involved in a quantitative trait like body height might have large variance (some are taller than others), making prediction of the phenotype from a particular genotype alone difficult. The expression of a phenotype may be modified by the effects of aging, other genetic loci, or environmental factors. In genetics, epistasis is the phenomenon where the effects of one gene are modified by one or several other genes, which are sometimes called modifier genes. The gene whose phenotype is expressed is called epistatic, while the phenotype altered or suppressed is called hypostatic. Epistasis can be contrasted with dominance, which is an interaction between alleles at the same gene locus. Epistasis is often studied in relation to Quantitative Trait Loci (QTL) and polygenic inheritance. In general, the fitness increment of any one allele depends in a complicated way on many other alleles; but, because of the way that the science of population genetics was developed. Epistasis and genetic interaction refer to different aspects of the same phenomenon. The term epistasis is widely used in population genetics and refers especially to the statistical properties of the phenomenon, and does not necessarily imply biochemical interaction between gene products. However, in general epistasis is used to denote the
Notes made by kozi1989 email:kozi1989@yahoo.gr
55
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
departure from 'independence' of the effects of different genetic loci . Confusion often arises due to the varied interpretation of 'independence between different branches of biology. Examples of tightly linked genes having epistatic effects on fitness are found in supergenes and the human major histocompatibility complex genes. The effect can occur directly at the genomic level, where one gene could code for a protein preventing transcription of the other gene. Alternatively, the effect can occur at the phenotypic level. For example, the gene causing albinism would hide the gene controlling color of a person's hair. In another example, a gene coding for a widow's peak would be hidden by a gene causing baldness. Fitness epistasis (where the affected trait is fitness) is one cause of linkage disequilibrium. Studying genetic interactions can reveal gene function, the nature of the mutations, functional redundancy, and protein interactions. Because protein complexes are responsible for most biological functions, genetic interactions are a powerful tool.
19. Laws of heredity
Basic types of inheritance. Inheritance of independently combining genes Single-locus inheritance Mendelian trait is one that is controlled by a single locus and shows a simple Mendelian inheritance pattern. Dominant-recessive Black and brown provide a clear example of a dominant-recessive relationship among alleles. Every dog has two genes at the black/brown locus. If both genes are for black, or if one is for black and one is for brown, the dog is black, most readily identified by nose color. If both genes are for brown, the dog is brown, again most readily identified by nose color. BB cannot be distinguished from Bb without genetic tests or breeding tests. Many genetic diseases, especially those that can be traced to an inactive or wrongly active form of a particular protein, are inherited in a simple recessive fashion. van Willebrand's disease (vWD) for instance, is inherited as a simple recessive within the Shetland Sheepdog breed. Intermediate Although this type of inheritance is common, it has a variety of names (incomplete dominance and overdominance are two common ones) some of which are also used for other things entirely. Here I will use it to refer to the type of inheritance in which the animal carrying two identical alleles shows one phenotype, the animal carrying two different identical alleles shows a different phenotype, and the animal carrying one copy of each of the alleles shows a third phenotype, usually intermediate between the two extremes but clearly distinguishable from either. In
Notes made by kozi1989 email:kozi1989@yahoo.gr
56
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
dogs, merle color is a good example of this type of inheritance. If we define M as merle and m as non-merle, we find we have three genotypes: 1) Mm non-merle, with normal intense color 2) Mm merle, with normal color diluted in a rather patchy fashion 3) Mm homozygous merle, extreme dilution, dog mostly white if a white-spotting gene is also present, and often with anomalies in hearing, vision and/or fertility. Note that there is really a continuum between dominant-recessive and intermediate inheritance. In Shetland Sheepdogs, for instance, sables carrying one gene for tan-point have on average more dark shading than dogs with two sable genes. However, the darkest shading on dogs pure for sable is probably darker than the lightest shading on dogs carrying a gene for tan-point. In practice, intermediate inheritance is often treated as if it were a special case of dominant-recessive inheritance, as can be seen by the symbols used for merle and non-merle - usually the capital letter refers to a dominant gene and the lower-case letter refers to a recessive gene. I think a separate name is justified because it could be equally well argued that homozygous merle is an undesirable recessive for which the merle color is a marker that the dog carries the merle gene. Many of the standard color genes normally treated as dominant-recessive do in fact have intermediate inheritance, the heterozygote generally much more similar to one homozygote than the other, between at least some alleles in the series. Coat color gene loci with at least some allele pairs leaning toward intermediate inheritance include A (agouti, patterning of black and tan), C (color, intensity of color), and S (white spotting). I suspect the same is true for T (ticking), G (graying) and even D (dilution) if another diluting gene, such as merle, is present. This may be much more generally true than is recognized. Co-dominant The dividing line between intermediate inheritance and co-dominant inheritance is fuzzy. Co-dominance is more likely to be used when biochemistry is concerned, as in blood types. Co-dominance means that both alleles at a locus are expressed. Co-dominance in Xlinked genes is a special case that will be treated under sex-linked inheritance. Sex-limited A classic example of a sex-limited trait in dogs is unilateral or bilateral cryptorchidism, in which one or both testicles cannot be found in their usual position in the scrotum. Since a bitch has no testicles, she cannot be a cryptorchid - but she can carry the gene(s) for cryptorchidism, and pass them to her sons. Likewise, genes affecting milk production are not normally expressed in a male. The main problem with sex-limited inheritance is that it is impossible to know even the phenotypes of the unaffected sex in a pedigree, which makes it difficult to determine the mode of inheritance. In sex-influenced inheritance, the genes behave differently in the two sexes, probably because the sex hormones provide different cellular environments in males and females. A
Notes made by kozi1989 email:kozi1989@yahoo.gr
57
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
classic example in people is male early-onset pattern baldness. The gene for baldness behaves as a dominant in males but as a recessive in females. Heterozygous males are bald and will pass the gene to about 50% of their offspring of either sex. However, only the males will normally be bald unless the mother also carries the pattern baldness gene without showing it (female heterozygote.) If the mother is affected with baldness (homozygous) but the father is not, all of the sons will be affected and all of the daughters will be non-affected carriers. A bald man may get pattern baldness from either parent; a bald woman must have received the gene from both parents. Sex-linked Every mammal has a number of paired chromosomes, that are similar in appearance and line up with each other during gamete production (sperm and eggs). In addition, each mammal has two chromosomes that determine sex. These are generally called X and Y in mammals. Normal pairing of chromosomes during the production of gametes will put one or the other in each sperm or ovum. In mammals, XY develops testicles which secrete male sex hormones and the fetus develops into a male. An XX fetus develops into a female. Thus sperm can be either X or Y; ova are always X. Sex linked inheritance involves genes located on either the X or the Y chromosome. Females can be homozygous or heterozygous for genes carried on the X chromosome; males can only be hemizygous.
20. Laws of heredity
Inheritance of linked genes. Crossing over. Molecular mechanisms of crossing over. Groups of linked genes in the human Genetic linkage is the tendency of certain loci or alleles to be inherited together . Genetic loci that are physically close to one another on the same chromosome tend to stay together during meiosis, and are thus genetically linked. At the beginning of normal meiosis, a homologous chromosome pair (called a bivalent, made up of a chromosome from the mother and a chromosome from the father) interwine and exchange sections or fragments of chromosome. The pair then breaks apart to form two chromosomes with a new combination of genes that differs from the combination supplied by the parents. Through this process of recombining genes, organisms can produce offspring with new combinations of maternal and paternal traits that may contribute to or enhance survival. This recombination of genes, called the crossing over of DNA , can cause alleles previously on the same chromosome to be separated and end up in different daughter cells. The farther the two alleles are apart, the greater the chance that a cross-over event may occur between them, and the greater the chance that the alleles are separated. relative distance between two genes can be calculated by taking the offspring of an organism showing two linked genetic traits, and finding the percentage of the offspring
Notes made by kozi1989 email:kozi1989@yahoo.gr
58
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
where the two traits do not run together. The higher the percentage of descendants that does not show both traits, the farther apart on the chromosome the two genes are. Genes for which this percentage is lower than 50% are typically thought to be linked. Genetic linkage can also be understood by looking at the relationships among phenotypes. Among individuals of an experimental population or species, some phenotypes or traits can occur randomly with respect to one another, or with some correlation with respect to one another.The former is known as independent assortment. Independent assortment occurs when the genes affecting the phenotypes are found on different chromosomes or separated by a great enough distance on the same chromosome that recombination occurs at least half of the time. The latter is known as genetic linkage. This occurs as an exception to independent assortment, and develops when genes appear near one another on the same chromosome. This phenomenon causes the genes to usually be inherited as a single unit. Genes inherited in this way are said to be linked, and are referred to as "linkage groups". For example, in fruit flies, the genes affecting eye color and wing length are inherited together because they appear on the same chromosome. Chromosomal crossover (or crossing over) is an exchange of genetic material between homologous chromosomes. It is one of the final phases of genetic recombination, which occurs during prophase I of meiosis (pachytene) in a process called synapsis. Synapsis begins before the synaptonemal complex develops, and is not completed until near the end of prophase I. Crossover usually occurs when matching regions on matching chromosomes break and then reconnect to the other chromosome. The pairing of homologues at the beginning of meiosis I ensures that each gamete receives one member of each pair. Homologues contact each other along much of their length and are held together by a special protein structure called the synaptonemal complex. The synaptonemal complex is a protein structure that forms between two pairs of sister chromatids during meiosis and that is thought to mediate chromosome pairing, synapsis, and recombination (crossing-over). This association of the homologues may persist from hours to days. The association of the two chromosomes is called a bivalent, and because there are four chromatids involved it is also called a tetrad. The points of attachment are called chiasmata (singular, chiasma). The pairing of homologues brings together the near-identical sequences found on each chromosome, and this sets the stage for crossing over. Theexact mechanism by which crossing over occurs is not known. Crossing overis controlled by a very large protein complex called a recombination nodule. Some of the proteins involved also play roles in DNA replication and repair, which is not surprising, considering that all three processes require breaking and reforming the DNA double helix. One plausible model supported by available evidence suggests that crossing over begins when one chromatid is cut through, making a break in the double-stranded DNA (recall that each DNA strand is a double helix of nucleotides). A nuclease enzyme then
Notes made by kozi1989 email:kozi1989@yahoo.gr
59
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
removes nucleotides from each side of the DNA strand, but in opposite directions, leaving each side with a single-stranded tail, perhaps 600 to 800 nucleotides long. One tail is then thought to insert itself along the length of one of the nonsister chromatids, aligning with its complementary sequence (i.e., if the tail sequence is ATCCGG, it aligns with TAGGCC on the nonsister strand). If a match is made, the tail pairs with this strand of the nonsister chromatid. This displaces the original paired strand on the nonsister chromatid, which is then freed to pair with the other single-stranded tail. The gaps are filled by a DNA polymerase enzyme. Finally, the two chromatids must be separated from each other, which requires cutting all the strands and rejoining the cut ends. *The term chiasma is linked if not identical to chromosomal crossover. *X chromosomes
21. Heredity and environment. Reaction norm. Phenocopies and genocopies
Genotype variation. Recombinational variation A phenocopy is an individual whose phenotype (generally referring to a single trait), under a particular environmental condition, is identical to the one of another individual whose phenotype is determined by the genotype. In other words the phenocopy environmental condition mimics the phenotype produced by a gene. A genocopy is an individual whose genotype, under a particular environment condition. Genotype Variations: The genotype is the genetic constitution of a cell, an organism, or an individual, that is the specific allele makeup of the individual, usually with reference to a specific character under consideration. For instance, the human albino gene has two allelic forms, dominant A and recessive a, and there are three possible genotypes- AA (homozygous dominant), Aa (heterozygous), and aa (homozygous recessive). Recombinational Variation: Genetic recombination is the process by which a strand of genetic material (usually DNA; but can also be RNA) is broken and then joined to a different DNA molecule. In eukaryotes recombination commonly occurs during meiosis as chromosomal crossover between paired chromosomes. This process leads to offspring having different combinations of genes from their parents and can produce new alleles. In molecular biology "recombination" can also refer to artificial and deliberate recombination of disparate pieces of DNA, often from different organisms, creating what is called recombinant DNA.
22. Mutational variation gene, chromosomal and genome mutations
Mutations and disease. Causes for mutations (mutagenic factors)
Notes made by kozi1989 email:kozi1989@yahoo.gr
60
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Gene Mutations are changes to the nucleotide sequence of the genetic material of an organism. Mutations can be caused by copying errors in the genetic material during cell division, by exposure to ultraviolet or ionizing radiation, chemical mutagens, or viruses. In multicellular organisms, mutations can be subdivided into germ line mutations, which can be passed on to descendants, and somatic mutations. The source of the mutation is unrelated to the consequence, although the consequences are related to which cells are affected. Mutations create variations in the gene pool. Less favorable (or deleterious) mutations can be reduced in frequency in the gene pool by natural selection, while more favorable (beneficial or advantageous) mutations may accumulate and result in adaptive evolutionary changes. Chromosomal Mutation: A mutation involving a long segment of DNA. These mutations can involve deletions, insertions, or inversions of sections of DNA. In some cases, deleted sections may attach to other chromosomes, disrupting both the chromosomes that loses the DNA and the one that gains it. Also referred to as a chromosomal rearrangement, which is very similar genome mutation. Mutations of the gene, chromosomes or genome cause diseases. Mutational disease can also be called genetic disorder. Genetic disorder is a condition caused by abnormalities in genes or chromosomes. While some diseases, such as cancer, are due to genetic abnormalities acquired in a few cells during life, the term "genetic disease" most commonly refers to diseases present in all cells of the body and present since conception. Some genetic disorders are caused by chromosomal abnormalities due to errors in meiosis, the process which produces reproductive cells such as sperm and eggs. Defective genes (mutation has already occur) may also be inherited intact from the parents. This can often happen unexpectedly when two healthy carriers of a defective recessive gene reproduce, but can also happen when the defective gene is dominant. There are few variations of mutation: Autosomal dominant: Only one mutated copy of the gene will be necessary for a person to be affected by an autosomal dominant disorder. Autosomal recessive: Two copies of the gene must be mutated for a person to be affected by an autosomal recessive disorder. An affected person usually has unaffected parents who each carry a single copy of the mutated gene (and are referred to as carriers). X-linked dominant: X-linked dominant disorders are caused by mutations in genes on the X chromosome. X-linked recessive
Notes made by kozi1989 email:kozi1989@yahoo.gr
61
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
X-linked recessive disorders are also caused by mutations in genes on the X chromosome. Males are more frequently affected than females, and the chance of passing on the disorder differs between men and women. Y-linked: Y-linked disorders are caused by mutations on the Y chromosome. Only males can get them, and all of the sons of an affected father are affected. Causes for the occurance of mutations Base substitution- spontaneous pairing errors or polymerase mistakes may occur during DNA replication Chemical modification- A base may be chemically altered by a mutagenic chemical DNA breaks- Ionizing radiation can cause double strand breaks in DNA Slipped mispairing-Deletions may result from deletion of a codon. Triplet expansion- When a 3-base sequence is repeated several times causing the formation of a mutant gene. Mutagenic factors are factors that could change the DNA sequence that makes up the gene.this factors could be hereditary or enviromental. Hereditary includes insertion deletion. Enviromental could be exposure to high intensity of ultraviolet rays high penetarting rays gamma rays toxic chemicals such as hydroxylamine infections
23. Genetic engineering
Genetic engineering at the level of population, organism and cell Genetic engineering, also called genetic modification, is the direct human manipulation of an organism's genome using modern DNA technology. It involves the introduction of foreign DNA or synthetic genes into the organism of interest. The introduction of new DNA does not require the use of classical genetic methods, however traditional breeding methods are typically used for the propagation of recombinant organisms. An organism that is generated through the introduction of recombinant DNA is considered to be a genetically modified organism. The first organisms genetically engineered were bacteria in 1973 and then mice in 1974. Insulin-producing bacteria were commercialized in 1982 and genetically modified food has been sold since 1994. The most common form of genetic engineering involves the insertion of new genetic material at an unspecified location in the host genome. This is accomplished by isolating and copying the genetic material of interest using molecular cloning methods to generate a DNA sequence containing the required genetic elements for expression, and then inserting this construct into the host organism. Other forms of genetic engineering include gene targeting
Notes made by kozi1989 email:kozi1989@yahoo.gr
62
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
and knocking out specific genes via engineered nucleases such as zinc finger nucleases or engineered homing endonucleases. Genetic engineering alters the genetic makeup of an organism using techniques that introduce heritable material prepared outside the organism either directly into the host or into a cell that is then fused or hybridized with the host .[1] This involves using recombinant nucleic acid (DNA or RNA) techniques to form new combinations of heritable genetic material followed by the incorporation of that material either indirectly through a vector system or directly through micro-injection, macro-injection and micro-encapsulation techniques. Genetic engineering does not include traditional animal and plant breeding, in vitro fertilisation, induction of polyploidy, mutagenesis and cell fusion techniques that do not use recombinant nucleic acids or a genetically modified organism in the process.[1] Cloning and stem cell research, although not considered genetic engineering,[2] are closely related and genetic engineering can be used within them.[3] Synthetic biology is an emerging discipline that takes genetic engineering a step further by introducing artificially synthesized genetic material from raw materials into an organism.[4] If genetic material from another species is added to the host, the resulting organism is called transgenic. If genetic material from the same species or a species that can naturally breed with the host is used the resulting organism is called cisgenic.[5] Genetic engineering can also be used to remove genetic material from the target organism, creating a knock out organism.[6] In Europe genetic modification is synonymous with genetic engineering while within the United States of America it can also refer to conventional breeding methods.[7] Within the scientific community, the term genetic engineering is not commonly used; more specific terms such as transgenic are preferred.
24. Genetic engineering
Genetic engineering at subcellular level, cloning by transplantation of somatic cell nuclei into oocytes In genetics and developmental biology, somatic cell nuclear transfer (SCNT) is a laboratory technique for creating a clonal embryo, using an ovum with a donor nucleus (see process below). It can be used in embryonic stem cell research, or, potentially, in regenerative medicine where it is sometimes referred to as "therapeutic cloning". It can also be used as the first step in the process of reproductive cloning. The process In SCNT the nucleus, which contains the organism's DNA, of a somatic cell (a body cell other than a sperm or egg cell) is removed and the rest of the cell discarded. At the same time, the nucleus of an egg cell is removed. The nucleus of the somatic cell is then inserted into the denucleated egg cell. After being inserted into the egg, the somatic cell nucleus is reprogrammed by the host cell. The egg, now containing the nucleus of a somatic cell, is stimulated with a shock and will begin to divide. After many mitotic divisions in
Notes made by kozi1989 email:kozi1989@yahoo.gr
63
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
culture, this single cell forms a blastocyst (an early stage embryo with about 100 cells) with almost identical DNA to the original organism. The technique of transferring a nucleus from a somatic cell into an egg that produced Dolly was an extension of experiments that had been ongoing for over 40 years. In the simplest terms, the technique used to produce Dolly the sheep - somatic cell nuclear transplantation cloning - involves removing the nucleus of an egg and replacing it with the diploid nucleus of a somatic cell. SCNT in reproductive cloning This technique is currently the basis for cloning animals (such as the famous Dolly the sheep), and in theory could be used to clone humans. However, most researchers believe that in the foreseeable future it will not be possible to use this technique to produce a human clone that will develop to term. However, it is still a possibility and can become more probable in the future as it will probably need a few more adjustments to work for humans. Limitations In SCNT, not all of the donor cell's genetic information is transferred, as the donor cell's mitochondria that contain their own mitochondrial DNA are left behind. The resulting hybrid cells retain those mitochondrial structures which originally belonged to the egg. As a consequence, clones such as Dolly that are born from SCNT are not perfect copies of the donor of the nucleus.
25. Recombinant DNA techniques and gene engineering
Specific DNA cleavage, most important enzymes in recombinant DNA techniques. DNA sequencing Recombinant DNA (rDNA) molecules are DNA sequences that result from the use of laboratory methods (molecular cloning) to bring together genetic material from multiple sources, creating sequences that would not otherwise be found in biological organisms. Recombinant DNA is possible because DNA molecules from all organisms share the same chemical structure; they differ only in the sequence of nucleotides within that identical overall structure. Consequently, when DNA from a foreign source is linked to host sequences that can drive DNA replication and then introduced into a host organism, the foreign DNA is replicated along with the host DNA. Recombinant DNA molecules are sometimes called chimeric DNA, because they are usually made of material from two different species, like the mythological chimera. The DNA sequences used in the construction of recombinant DNA molecules can originate from any species. For example, plant DNA may be joined to bacterial DNA, or human DNA may be joined with fungal DNA. In addition, DNA sequences that do not occur anywhere in nature may be created by the chemical synthesis of DNA, and incorporated into recombinant molecules. Using recombinant DNA technology and synthetic DNA, literally any DNA sequence may be created and introduced into any of a very wide range of living organisms.
Notes made by kozi1989 email:kozi1989@yahoo.gr
64
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Proteins that result from the expression of recombinant DNA within living cells are termed recombinant proteins. When recombinant DNA encoding a protein is introduced into a host organism, the recombinant protein will not necessarily be produced. Expression of foreign proteins requires the use of specialized expression vectors and often necessitates significant restructuring of the foreign coding sequence. It is important to note that recombinant DNA differs from genetic recombination in that the former results from artificial methods in the test tube, while the latter is a normal biological process that results in the remixing of existing DNA sequences in essentially all organisms. Acid nucleics cleavage can occur mainly by two processes: 1) Nucleophilic Cleavage: Transesterification + Hydrolysis 2) Oxidative cleavage Photoinduced Oxidative Cleavage of Nucleic Acids Firstly we must differentiate between photonucleases and photocleavage agents: Photonucleases are compounds that react in a photoexcited electronic state with a nucleic acid producing chain cleavage. Photocleavage agents are compounds whose photoexcited electronic state can initiate a serie of chemical reactions that finally produce the cleavage of the nucleic acid. Usually, the processes of DNA cleavage are oxidative (although some of them occurs in anaerobic conditions) and can be modulated in function of the irradiation time. Molecular cloning is the laboratory process used to create recombinant DNA. It is one of two widely-used methods (along with polymerase chain reaction, abbr. PCR) used to direct the replication of any specific DNA sequence chosen by the experimentalist. The fundamental difference between the two methods is that molecular cloning involves replication of the DNA within a living cell, while PCR replicates DNA in the test tube, free of living cells. Formation of recombinant DNA requires a cloning vector, a DNA molecule that will replicate within a living cell. Vectors are generally derived from plasmids or viruses, and represent relatively small segments of DNA that contain necessary genetic signals for replication, as well as additional elements for convenience in inserting foreign DNA, identifying cells that contain recombinant DNA, and, where appropriate, expressing the foreign DNA. The choice of vector for molecular cloning depends on the choice of host organism, the size of the DNA to be cloned, and whether and how the foreign DNA is to be expressed.[5] In standard cloning protocols, the cloning of any DNA fragment essentially involves seven steps: (1) Choice of host organism and cloning vector, (2) Preparation of vector DNA, (3) Preparation of DNA to be cloned, (4) Creation of recombinant DNA, (5) Introduction of recombinant DNA into the host organism, (6) Selection of organisms containing recombinant
Notes made by kozi1989 email:kozi1989@yahoo.gr
65
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
DNA, (7) Screening for clones with desired DNA inserts and biological properties.[4] These steps are described in some detail in a related article (molecular cloning). Recombinant DNA is made possible by two important enzymes. Restriction enzymes and DNA ligase are the two principal tools, first used by Paul Berg in 1972, employed to alter DNA. Restriction enzymes are used to "cut" DNA at a specific location. These are used if the DNA must be removed from the entire strand of DNA (as in the first method for gene isolation above) and also to open the section of DNA into which the isolated gene will be inserted. DNA ligase is used to "glue" two sections of DNA together. However, the isolated gene cannot be directly inserted into the target DNA. A cloning vector must be used. In the Cohen and Boyer experiment, a plasmid was used. A plasmid is a circular section of DNA which can be inserted into a cell. The cell then takes on the characteristics of this DNA. This technique is easy, however it does not work on larger animals. The other cloning vector is a virus. The isolated gene is inserted into the DNA of a virus and "glued" using DNA ligase. The virus then injects the gene into the cells main DNA. The cell then begins producing the desired protein. Cloning is looked upon as a much easier, cost effective, and reliable way of mass producing genetically altered larger organisms. DNA sequencing includes several methods and technologies that are used for determining the order of the nucleotide basesadenine, guanine, cytosine, and thymine in a molecule of DNA. Knowledge of DNA sequences has become indispensable for basic biological research, other research branches utilizing DNA sequencing, and in numerous applied fields such as diagnostic, biotechnology, forensic biology and biological systematics. The advent of DNA sequencing has significantly accelerated biological research and discovery. The rapid speed of sequencing attained with modern DNA sequencing technology has been instrumental in the sequencing of the human genome, in the Human Genome Project. Related projects, often by scientific collaboration across continents, have generated the complete DNA sequences of many animal, plant, and microbial genomes. The first DNA sequences were obtained in the early 1970s by academic researchers using laborious methods based on two-dimensional chromatography. Following the development of dye-based sequencing methods with automated analysis, DNA sequencing has become easier and orders of magnitude faster.
26. Recombinant DNA techniques and gene engineering. DNA cloning. Polymerase
chain reaction. Gene engineering and gene therapy. DNA cloning refers to the procedure of isolating a defined DNA sequence and obtaining multiple copies of it in vivo. Cloning is frequently employed to amplify DNA fragments containing genes, but it can be used to amplify any DNA sequence such as promoters, non-coding sequences, chemically synthesised oligonucleotides and randomly fragmented DNA. Cloning is utilized in a wide array of biological experiments and technological applications such as large scale protein production. A novel procedure of
Notes made by kozi1989 email:kozi1989@yahoo.gr
66
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
cloning or subcloning of any DNA fragment by inserting the special DNA fragment of interest into a special area of target DNA through interchange of the relevant DNA fragments. This is a one-step reaction: simple, efficient, facilitating high throughput or automatic cloning and/or subcloning. In molecular biology, a library is a collection of DNA fragments that is stored and propagated in a population of micro-organisms through the process of molecular cloning. There are different types of DNA libraries, including cDNA libraries (formed from reversetranscribed RNA) and genomic libraries (formed from genomic DNA). DNA library technology is a mainstay of current molecular biology, and the applications of these libraries depends on the source of the original DNA fragments. There are differences in the cloning vectors and techniques used in library preparation, but in general each DNA fragment is uniquely inserted into a cloning vector and the pool of recombinant DNA molecules is then transferred into a population of bacteria or yeast such that each organism contains on average one construct (vector + insert). As the population of organisms is grown in culture, the DNA molecules contained within them are copied and propagated (thus, "cloned"). The term "library" can refer to a population of organisms, each of which carries a DNA molecule inserted into a cloning vector, or alternatively to the collection of all of the cloned vector molecules. cDNA libraries A cDNA library represents a sample of the mRNA purified from a particular source (either a collection of cells, a particular tissue, or an entire organism), which has been converted back to a DNA template by the use of the enzyme reverse transcriptase. It thus represents the genes that were being actively transcribed in that particular source under the physiological, developmental, or environmental conditions that existed when the mRNA was purified. cDNA libraries can be generated using techniques that promote "full-length" clones or under conditions that generate shorter fragments used for the identification of "expressed sequence tags". cDNA libraries are useful in reverse genetics, but they only represent a very small (less than 1%) portion of the overall genome in a given organism. The polymerase chain reaction (PCR) is a scientific technique in molecular biology to amplify a single or a few copies of a piece of DNA across several orders of magnitude, generating thousands to millions of copies of a particular DNA sequence. Developed in 1983 by Kary Mullis,[1] PCR is now a common and often indispensable technique used in medical and biological research labs for a variety of applications.[2][3] These include DNA cloning for sequencing, DNA-based phylogeny, or functional analysis of genes; the diagnosis of hereditary diseases; the identification of genetic fingerprints (used in forensic sciences and paternity testing); and the detection and diagnosis of infectious diseases. In 1993, Mullis was awarded the Nobel Prize in Chemistry along with Michael Smith for his work on PCR.[4]
Notes made by kozi1989 email:kozi1989@yahoo.gr
67
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The method relies on thermal cycling, consisting of cycles of repeated heating and cooling of the reaction for DNA melting and enzymatic replication of the DNA. Primers (short DNA fragments) containing sequences complementary to the target region along with a DNA polymerase (after which the method is named) are key components to enable selective and repeated amplification. As PCR progresses, the DNA generated is itself used as a template for replication, setting in motion a chain reaction in which the DNA template is exponentially amplified. PCR can be extensively modified to perform a wide array of genetic manipulations. Almost all PCR applications employ a heat-stable DNA polymerase, such as Taq polymerase, an enzyme originally isolated from the bacterium Thermus aquaticus. This DNA polymerase enzymatically assembles a new DNA strand from DNA building-blocks, the nucleotides, by using single-stranded DNA as a template and DNA oligonucleotides (also called DNA primers), which are required for initiation of DNA synthesis. The vast majority of PCR methods use thermal cycling, i.e., alternately heating and cooling the PCR sample to a defined series of temperature steps. These thermal cycling steps are necessary first to physically separate the two strands in a DNA double helix at a high temperature in a process called DNA melting. At a lower temperature, each strand is then used as the template in DNA synthesis by the DNA polymerase to selectively amplify the target DNA. The selectivity of PCR results from the use of primers that are complementary to the DNA region targeted for amplification under specific thermal cycling conditions. Genetic engineering, also called genetic modification, is the direct human manipulation of an organism's genome using modern DNA technology. It involves the introduction of foreign DNA or synthetic genes into the organism of interest. The introduction of new DNA does not require the use of classical genetic methods, however traditional breeding methods are typically used for the propagation of recombinant organisms. An organism that is generated through the introduction of recombinant DNA is considered to be a genetically modified organism. The first organisms genetically engineered were bacteria in 1973 and then mice in 1974. Insulin-producing bacteria were commercialized in 1982 and genetically modified food has been sold since 1994. The most common form of genetic engineering involves the insertion of new genetic material at an unspecified location in the host genome. This is accomplished by isolating and copying the genetic material of interest using molecular cloning methods to generate a DNA sequence containing the required genetic elements for expression, and then inserting this construct into the host organism. Other forms of genetic engineering include gene targeting and knocking out specific genes via engineered nucleases such as zinc finger nucleases or engineered homing endonucleases. Isolating the Gene Elements of genetic engineering
Notes made by kozi1989 email:kozi1989@yahoo.gr
68
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
First, the gene to be inserted into the genetically modified organism must be chosen and isolated. Presently, most genes transferred into plants provide protection against insects or tolerance to herbicides.[25] In animals the majority of genes used are growth hormone genes.[26] Once chosen the genes must be isolated. This typically involves multiplying the gene using polymerase chain reaction (PCR). If the chosen gene or the donor organism's genome has been well studied it may be present in a genetic library. If the DNA sequence is known, but no copies of the gene are available, it can be artificially synthesized. Once isolated, the gene is inserted into a bacterial plasmid. Constructs The gene to be inserted into the genetically modified organism must be combined with other genetic elements in order for it to work properly. The gene can also be modified at this stage for better expression or effectiveness. As well as the gene to be inserted most constructs contain a promoter and terminator region as well as a selectable marker gene. The promoter region initiates transcription of the gene and can be used to control the location and level of gene expression, while the terminator region ends transcription. The selectable marker, which in most cases confers antibiotic resistance to the organism it is expressed in, is needed to determine which cells are transformed with the new gene. The constructs are made using recombinant DNA techniques, such as restriction digests, ligations and molecular cloning. Gene Targeting The most common form of genetic engineering involves inserting new genetic material randomly within the host genome. Other techniques allow new genetic material to be inserted at a specific location in the host genome or generate mutations at desired genomic loci capable of knocking out endogenous genes. The technique of gene targeting uses homologous recombination to target desired changes to a specific endogenous gene. This tends to occur at a relatively low frequency in plants and animals and generally requires the use of selectable markers. The frequency of gene targeting can be greatly enhanced with the use of engineered nucleases such as zinc finger nucleases, engineered homing endonucleases, or nucleases created from TAL effectors. In addition to enhancing gene targeting, engineered nucleases can also be used to introduce mutations at endogenous genes that generate a gene knockout. Proccesses Transformation About 1% of bacteria are naturally able to take up foreign DNA but it can also be induced in other bacteria. Stressing the bacteria for example, with a heat shock or an electric shock, can make the cell membrane permeable to DNA that may then incorporate into their genome or exist as extrachromosomal DNA. DNA is generally inserted into animal
Notes made by kozi1989 email:kozi1989@yahoo.gr
69
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
cells using microinjection, where it can be injected through the cells nuclear envelope directly into the nucleus or through the use of viral vectors. In plants the DNA is generally inserted using Agrobacterium-mediated recombination or biolistics. In Agrobacterium-mediated recombination the plasmid construct must also contain T-DNA. Agrobacterium naturally inserts DNA from a tumor inducing plasmid into any susceptible plant's genome it infects, causing crown gall disease. The T-DNA region of this plasmid is responsible for insertion of the DNA. The genes to be inserted are cloned into a binary vector, which contains T-DNA and can be grown in both E. Coli and Agrobacterium. Once the binary vector is constructed the plasmid is transformed into Agrobacterium containing no plasmids and plant cells are infected. The Agrobacterium will then naturally insert the genetic material into the plant cells. In biolistics particles of gold or tungsten are coated with DNA and then shot into young plant cells or plant embryos. Some genetic material will enter the cells and transform them. This method can be used on plants that are not susceptible to Agrobacterium infection and also allows transformation of plant plastids. Another transformation method for plant and animal cells is electroporation. Electroporation involves subjecting the plant or animal cell to an electric shock, which can make the cell membrane permeable to plasmid DNA. In some cases the electroporated cells will incorporate the DNA into their genome. Due to the damage caused to the cells and DNA the transformation efficiency of biolistics and electroporation is lower than agrobacterial mediated transformation and microinjection.[39] Selection Not all the organism's cells will be transformed with the new genetic material; in most cases a selectable marker is used to differentiate transformed from untransformed cells. If a cell has been successfully transformed with the DNA it will also contain the marker gene. By growing the cells in the presence of an antibiotic or chemical that selects or marks the cells expressing that gene it is possible to separate the transgenic events from the nontransgenic. Another method of screening involves using a DNA probe that will only stick to the inserted gene. A number of strategies have been developed that can remove the selectable marker from the mature transgenic plant.[40] Regeneration As often only a single cell is transformed with genetic material the organism must be regrown from that single cell. As bacteria consist of a single cell and reproduce clonally regeneration is not necessary. In plants this is accomplished through the use of tissue culture. Each plant species has different requirements for successful regeneration through tissue culture. If successful an adult plant is produced that contains the transgene in every cell. In animals it is necessary to ensure that the inserted DNA is present in the embryonic stem cells. When the offspring is produced they can be screened for the presence of the gene. All offspring from the first generation will be heterozygous for the inserted gene and must be mated together to produce a homozygous animal.
Notes made by kozi1989 email:kozi1989@yahoo.gr
70
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Confirmation The finding that a recombinant organism contains the inserted genes is not usually sufficient to ensure that the genes will be expressed in an appropriate manner in the intended tissues of the recombinant organism. To examine the presence of the gene, further analysis frequently uses PCR, Southern hybridization, and DNA sequencing, which serve to determine the chromosomal location and copy number of the inserted gene. To examine expression of the trans-gene, an extensive analysis of transcription, RNA processing patterns, and the expression and localization of the protein product(s) is usually necessary, using methods including northern hybridization, quantitative RT-PCR, Western blot, immunofluorescence and phenotypic analysis. When appropriate, the organism's offspring are studied to confirm that the trans-gene and associated phenotype are stably inherited. Gene therapy is the insertion, alteration, or removal of genes within an individual's cells and biological tissues to treat disease. It is a technique for correcting defective genes that are responsible for disease development.[1] The most common form of gene therapy involves the insertion of functional genes into an unspecified genomic location in order to replace a mutated gene, but other forms involve directly correcting the mutation or modifying normal gene that enables a viral infection. Although the technology is still in its infancy, it has been used with some success. The most common form of genetic engineering involves the insertion of a functional gene at an unspecified location in the host genome.This is accomplished by isolating and copying the gene of interest, generating a construct containing all the genetic elements for correct expression, and then inserting this construct into a random location in the host organism. Other forms of genetic engineering include gene targeting and knocking out specific genes via engineered nucleases such as zinc finger nucleases, engineered I-CreI homing endonucleases, or nucleases generated from TAL effectors. An example of geneknockout mediated gene therapy is the knockout of the human CCR5 gene in T-cells in order to control HIV infection. This approach is currently being used in several human clinical trials. The biology of human gene therapy remains complex and many techniques need further development. Many diseases and their strict genetic link need to be understood more fully before gene therapy can be used appropriately. The public policy debate surrounding the possible use of genetically engineered material in human subjects has been equally complex. Major participants in the debate have come from the fields of biology, government, law, medicine, philosophy, politics, and religion, each bringing different views to the discussion. Gene therapy may be classified into the two following types: Germ line gene therapy In the case of germ line gene therapy, Germ cells, i.e., sperm or eggs, are modified by the introduction of functional genes, which are integrated into their genomes.
Notes made by kozi1989 email:kozi1989@yahoo.gr
71
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Therefore, the change due to therapy would be heritable and would be passed on to later generations. This new approach, theoretically, should be highly effective in counteracting genetic disorders and hereditary diseases. However, many jurisdictions prohibit this for application in human beings, at least for the present, for a variety of technical and ethical reasons. Somatic gene therapy In the case of somatic gene therapy, the therapeutic genes are transferred into the somatic cells of a patient. Any modifications and effects will be restricted to the individual patient only, and will not be inherited by the patient's offspring or later generations.
27. The organism as a unified system. Immunological homeostasis. Organs of the
immune system. Immunological homeostasis Maintaining the internal environment at a constant level or between narrow limits for Bodys defense system against foreign invaders and for Specific-antigen/antibody Ex-blood pH, gas concentrations, blood glucose, body temperature, water balance. Homeostasis is achived by regulated pathways and lymphocyte death and inactivation. The immune system is a tightly regulated network that is able to maintain a balance of immune homeostasis under normal physiological conditions. Normally, when challenged with foreign antigen, specific appropriate responses are initiated that are aimed at restoring homeostasis. However under particular circumstances, this balance is not maintained and immune responses either under or over react. Cancer is an example of a situation where the immune response can be inefficient or unresponsive, resulting in uncontrolled growth of the cancer cells. Effective immunological homeostasis relies on a continual balance between a number of factors, including helper T (Th) cell activation and regulatory T cell (Treg) suppression. When homeostasis is disrupted and the immune system tips in favor of activation, the host becomes susceptible to autoimmunity. The organs of the immune system either make the cells that participate in the immune response or act as sites for immune function. These organs include the lymphatic vessels, lymph nodes, tonsils, thymus, Peyer's patch, and spleen. The lymph nodes are small aggregations of tissues interspersed throughout the lymphatic system. White blood cells (lymphocytes) that function in the immune response are concentrated in the lymph nodes. Lymphatic fluid circulates through the lymph nodes via the lymphatic vessels. As the lymph filters through the lymph nodes, foreign cells of microorganisms are detected and overpowered. Primary lymphoid organs
Notes made by kozi1989 email:kozi1989@yahoo.gr
72
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The tonsils contain large numbers of lymphocytes. Located at the back of the throat and under the tongue, the tonsils filter out potentially harmful bacteria that may enter the body via the nose and mouth. Peyer's patches are lymphatic tissues which perform this same function in the digestive system. Peyer's patches are scattered throughout the small intestine and the appendix. They are also filled with lymphocytes that are activated when they encounter disease-causing microorganisms. The thymus gland is another site of lymphocyte production. Located within the upper chest region, the thymus gland is most active during childhood when it makes large numbers of lymphocytes. The lymphocytes made here do not stay in the thymus, however; they migrate to other parts of the body and concentrate in the lymph nodes. The thymus gland continues to grow until puberty; during adulthood, however, the thymus shrinks in size until it is sometimes impossible to detect in x-rays. Bone marrow, found within the bones, also produces lymphocytes. These lymphocytes migrate out of the bone marrow to other sites in the body. Because bone marrow is an integral part of the immune system, certain bone cancer treatments that require the destruction of bone marrow are extremely risky, because without bone marrow, a person cannot make lymphocytes. People undergoing bone marrow replacement must be kept in strict isolation to prevent exposure to viruses or bacteria. Secondary lymphoid organs Lymph nodes are present at the junctions of lymphatic vessels and serve as the first organized structures to encounter most antigens. The major function of the lymph nodes is to filter antigen from the lymph. The spleen acts as a reservoir for blood and any rupture to the spleen can cause dangerous internal bleeding, a potentially fatal condition. The spleen also destroys wornout red blood cells. Moreover, the spleen is also a site for immune function, since it contains lymphatic tissue and produces lymphocytes.
28. Antigen as inducers of the immune response. Haptens.
An antigen is a substance/molecule that, when introduced into the body, triggers the production of an antibody by the immune system, which will then kill or neutralize the antigen that is recognized as a foreign and potentially harmful invader. These invaders can be molecules such as pollen or cells such as bacteria. The term originally came from antibody generator and was a molecule that binds specifically to an antibody, but the term now also refers to any molecule or molecular fragment that can be bound by a major histocompatibility complex (MHC) and presented to a T-cell receptor. "Self" antigens are usually tolerated by the immune system; whereas "Non-self" antigens are identified as intruders and attacked by the immune system. Autoimmune disorders arise from the immune system reacting to its own antigens. In similar manner, an immunogen is a specific type of antigen. An immunogen is
Notes made by kozi1989 email:kozi1989@yahoo.gr
73
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
defined as a substance that is able to provoke an adaptive immune response if injected on its own. Stated another way, an immunogen is able to induce an immune response, whereas an antigen is able to combine with the products of an immune response once they are made. The overlapping concepts of immunogenicity and antigenicity are, therefore, subtly different. Immunogenicity is the ability to induce a humoral and/or cell-mediated immune response. Antigenicity is the ability to combine specifically with the final products of the immune response (i.e. secreted antibodies and/or surface receptors on T-cells). Although all molecules that have the property of immunogenicity also have the property of antigenicity, the reverse is not true." At the molecular level, an antigen is characterized by its ability to be "bound" at the antigen-binding site of an antibody. Note also that antibodies tend to discriminate between the specific molecular structures presented on the surface of the antigen (as illustrated in the Figure). Antigens are usually proteins or polysaccharides. This includes parts (coats, capsules, cell walls, flagella, fimbrae, and toxins) of bacteria, viruses, and other microorganisms. Lipids and nucleic acids are antigenic only when combined with proteins and polysaccharides. Non-microbial exogenous (non-self) antigens can include pollen, egg white, and proteins from transplanted tissues and organs or on the surface of transfused blood cells. Vaccines are examples of immunogenic antigens intentionally administered to induce acquired immunity in the recipient. Cells present their immunogenic-antigens to the immune system via a histocompatibility molecule. Depending on the antigen presented and the type of the histocompatibility molecule, several types of immune cells can become activated. Epitope - The distinct molecular surface features of an antigen capable of being bound by an antibody (a.k.a. antigenic determinant). Antigenic molecules, normally being "large" biological polymers, usually present several surface features that can act as points of interaction for specific antibodies. Any such distinct molecular feature constitutes an epitope. Therefore, most antigens have the potential to be bound by several distinct antibodies, each of which is specific to a particular epitope. Using the "lock and key" metaphor, the antigen itself can be seen as a string of keys - any epitope being a "key" - each of which can match a different lock. Different antibody idiotypes, each having distinctly formed complementarity determining regions, correspond to the various "locks" that can match "the keys" (epitopes) presented on the antigen molecule. Allergen - A substance capable of causing an allergic reaction. The (detrimental) reaction may result after exposure via ingestion, inhalation, injection, or contact with skin. Superantigen - A class of antigens that cause non-specific activation of T-cells, resulting in polyclonal T cell activation and massive cytokine release.
Notes made by kozi1989 email:kozi1989@yahoo.gr
74
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Tolerogen - A substance that invokes a specific immune non-responsiveness due to its molecular form. If its molecular form is changed, a tolerogen can become an immunogen. Immunoglobulin binding protein - These proteins are capable of binding to antibodies at positions outside of the antigen-binding site. That is, whereas antigens are the "target" of antibodies, immunoglobulin-binding proteins "attack" antibodies. Protein A, protein G, and protein L are examples of proteins that strongly bind to various antibody isotypes. Antigens can be classified in order of their class. Exogenous antigens Exogenous antigens are antigens that have entered the body from the outside, for example by inhalation, ingestion, or injection. The immune system's response to exogenous antigens is often subclinical. By endocytosis or phagocytosis, exogenous antigens are taken into the antigen-presenting cells (APCs) and processed into fragments. APCs then present the fragments to T helper cells (CD4+) by the use of class II histocompatibility molecules on their surface. Some T cells are specific for the peptide: MHC complex. They become activated and start to secrete cytokines. Cytokines are substances that can activate cytotoxic T lymphocytes (CTL), antibody-secreting B cells, macrophages, and other particles. Some antigens start out as exogenous antigens, and later become endogenous (for example, intracellular viruses). Intracellular antigens can be released back into circulation upon the destruction of the infected cell, again. Endogenous antigens Endogenous antigens are antigens that have been generated within previouslynormal cells as a result of normal cell metabolism, or because of viral or intracellular bacterial infection. The fragments are then presented on the cell surface in the complex with MHC class I molecules. If activated cytotoxic CD8+ T cells recognize them, the T cells begin to secrete various toxins that cause the lysis or apoptosis of the infected cell. In order to keep the cytotoxic cells from killing cells just for presenting self-proteins, self-reactive T cells are deleted from the repertoire as a result of tolerance (also known as negative selection). Endogenous antigens include xenogenic (heterologous), autologous and idiotypic or allogenic (homologous) antigens. An autoantigen is usually a normal protein or complex of proteins (and sometimes DNA or RNA) that is recognized by the immune system of patients suffering from a specific autoimmune disease. These antigens should, under normal conditions, not be the target of the immune system, but, due to mainly genetic and environmental factors, the normal immunological tolerance for such an antigen has been lost in these patients. Hapten is a small molecule that can elicit an immune response only when attached to a large carrier such as a protein; the carrier may be one that also does not elicit an
Notes made by kozi1989 email:kozi1989@yahoo.gr
75
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
immune response by itself. (In general, only large molecules, infectious agents, or insoluble foreign matter can elicit an immune response in the body.) Once the body has generated antibodies to a hapten-carrier adduct, the small-molecule hapten may also be able to bind to the antibody, but it will usually not initiate an immune response; usually only the haptencarrier adduct can do this. Sometimes the small-molecule hapten can even block immune response to the hapten-carrier adduct by preventing the adduct from binding to the antibody, a process called hapten inhibition. The mechanisms of absence of immune response may vary and involve complex immunological mechanisms, but can include absent or insufficient co-stimulatory signals from antigen-presenting cells. The first haptens used were aniline and its carboxyl derivatives (o-, m-, and paminobenzoic acid). A well-known example of a hapten is urushiol, which is the toxin found in poison ivy. When absorbed through the skin from a poison ivy plant, urushiol undergoes oxidation in the skin cells to generate the actual hapten, a reactive molecule called a quinone, which then reacts with skin proteins to form hapten adducts. Usually, the first exposure causes only sensitization, in which there is a proliferation of effector T-cells. After a second exposure later, the proliferated T cells can become activated, generating an immune reaction, producing the typical blisters of poison ivy exposure. Some haptens can induce autoimmune disease. An example is hydralazine, a blood pressure-lowering drug that occasionally can produce drug-induced lupus erythematosus in certain individuals. This also appears to be the mechanism by which the anaesthetic gas halothane can cause a life-threatening hepatitis, as well as the mechanism by which penicillin-class drugs cause autoimmune hemolytic anemia. Other haptens that are commonly used in molecular biology applications include fluorescein, biotin, digoxigenin, and dinitrophenol.
29. Antibodies. Structure and function of antibodies, antigenic determination of
antibodies. Antigen-antibody reaction. An antibody, also known as an immunoglobulin, is a large Y-shaped protein used by the immune system to identify and neutralize foreign objects like bacteria and viruses. The antibody recognizes a unique part of the foreign target, termed an antigen. Each tip of the "Y" of an antibody contains a paratope (a structure analogous to a lock) that is specific for one particular epitope (similarly analogous to a key) on an antigen, allowing these two structures to bind together with precision. Using this binding mechanism, an antibody can tag a microbe or an infected cell for attack by other parts of the immune system, or can neutralize its target directly (for example, by blocking a part of a microbe that is essential for its invasion and survival). The production of antibodies is the main function of the humoral immune system. Antibodies are produced by a type of white blood cell called a plasma cell . Antibodies can occur in two physical forms, a soluble form that is secreted from the cell,
Notes made by kozi1989 email:kozi1989@yahoo.gr
76
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
and a membrane-bound form that is attached to the surface of a B cell and is referred to as the B cell receptor (BCR). The BCR is only found on the surface of B cells and facilitates the activation of these cells and their subsequent differentiation into either antibody factories called plasma cells, or memory B cells that will survive in the body and remember that same antigen so the B cells can respond faster upon future exposure.[4] In most cases, interaction of the B cell with a T helper cell is necessary to produce full activation of the B cell and, therefore, antibody generation following antigen binding.[5] Soluble antibodies are released into the blood and tissue fluids, as well as many secretions to continue to survey for invading microorganisms. Antibodies are glycoproteins belonging to the immunoglobulin superfamily ; the terms antibody and immunoglobulin are often used interchangeably.[6] Antibodies are typically made of basic structural unitseach with two large heavy chains and two small light chains. There are several different types of antibody heavy chains, and several different kinds of antibodies, which are grouped into different isotypes based on which heavy chain they possess. Five different antibody isotypes are known in mammals, which perform different roles, and help direct the appropriate immune response for each different type of foreign object they encounter. Though the general structure of all antibodies is very similar, a small region at the tip of the protein is extremely variable, allowing millions of antibodies with slightly different tip structures, or antigen binding sites, to exist. This region is known as the hypervariable region. Each of these variants can bind to a different target, known as an antigen.[1] This enormous diversity of antibodies allows the immune system to recognize an equally wide variety of antigens.[6] The large and diverse population of antibodies is generated by random combinations of a set of gene segments that encode different antigen binding sites (or paratopes), followed by random mutations in this area of the antibody gene, which create further diversity.[7][8] Antibody genes also re-organize in a process called class switching that changes the base of the heavy chain to another, creating a different isotype of the antibody that retains the antigen specific variable region. This allows a single antibody to be used by several different parts of the immune system. An epitope, also known as antigenic determinant, is the part of an antigen that is recognized by the immune system, specifically by antibodies, B cells, or T cells. The part of an antibody that recognizes the epitope is called a paratope. Although epitopes are usually thought to be derived from non-self proteins, sequences derived from the host that can be recognized are also classified as epitopes. The epitopes of protein antigens are divided into two categories, conformational epitopes and linear epitopes, based on their structure and interaction with the paratope.[1] A conformational epitope is composed of discontinuous sections of the antigen's amino acid sequence. These epitopes interact with the paratope based on the 3-D surface features and shape or tertiary structure of the antigen. Most epitopes are conformational. In contrast linear epitopes interact with the paratope based on their primary
Notes made by kozi1989 email:kozi1989@yahoo.gr
77
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
structure. The amino acids that make up a linear epitope are a continuous sequence of amino acids from the antigen. T cell epitopes are presented on the surface of an antigen-presenting cell, where they are bound to MHC molecules. T cell epitopes presented by MHC class I molecules are typically peptides between 8 and 11 amino acids in length, whereas MHC class II molecules present longer peptides, and non-classical MHC molecules also present non-peptidic epitopes such as glycolipids. Cross-reactivity Epitopes are sometimes cross-reactive. This property is exploited by the immune system in regulation by anti-idiotypic antibodies (originally proposed by Nobel laureate Niels Kaj Jerne). If an antibody binds to an antigen's epitope, the paratope could become the epitope for another antibody that will then bind to it. If this second antibody is of IgM class, its binding can upregulate the immune response; if the second antibody is of IgG class, its binding can downregulate the immune response. A reaction that occurs when an antigen combines with a corresponding antibody to produce an immune complex. A substance that induces the immune system to form a corresponding antibody is called an immunogen. All immunogens are also antigens because they react with corresponding antibodies; however, an antigen may not be able to induce the formation of an antibody and therefore may not be an immunogen. For instance, lipids and all low-molecular-weight substances are not immunogenic. However, many such substances, termed haptens, can be attached to immunogens, called carriers, and the complex then acts as a new immunogen. A molecule of antibody has two identical binding sites for one antigen or more, depending on its class. Each site is quite small and can bind only a comparably small portion of the surface of the antigen, which is termed an epitope. The specificity of an antibody for an antigen depends entirely upon the possession of the appropriate epitope by an antigen. The binding site on the antibody and the epitope on the antigen are complementary regions on the surface of the respective molecules which interlock in the antigen-antibody reaction. The intensity with which an antibody binds to the antigen depends on the exactitude of the fit between the respective binding site and epitope, as well as some inherent characteristics of the reacting molecules and factors in the environment. The epitope must be continuous spatially, but not structurally: in other words, if the molecule of the antigen consists of several chains, then an epitope may be formed by adjacent regions on two different chains, as well as by adjacent regions on the same chain. If the epitope is now modified either chemically (for example, by altering the hapten) or physically (for example, by causing the chains to separate), then its fit in the binding site will be altered or abolished, and the antigen will react with the antibody either less strongly or not at all. The immune complex formed in the reaction consists of closely apposed, but still discrete, molecules of antigen and antibody. Therefore, the immune complex can dissociate into the original molecules. The proportion of the dissociated, individual molecules of
Notes made by kozi1989 email:kozi1989@yahoo.gr
78
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
antigen and antibody to those of the immune complex clearly depends on the intensity of the binding. These proportions can be measured in a standardized procedure, so that the concentration of antigen [Ag], antibody [Ab], and the immune complex [AgAb] becomes known. A fraction is then calculated and called either the dissociation constant or the association constant. The magnitude of either of these constants can be used subsequently to assess the intensity of the antigen-antibody reaction. See also Immunoassay. Only one epitope of its kind generally occurs on each molecule of antigen, other than that which consists of multiple, identical units, though many epitopes of different configuration are possible. The reaction between antigen and antibody is followed by a structural change in the remainder of the antibody molecule. The change results in the appearance of previously hidden regions of the molecule. Some of these hidden regions have specific functions, such as binding complement. Fixation of complement by immune complexes has been used to detect and measure antigen-antibody reactions.
30. Complement system
The complement system helps or complements the ability of antibodies and phagocytic cells to clear pathogens from an organism. It is part of the immune system called the innate immune system that is not adaptable and does not change over the course of an individual's lifetime. However, it can be recruited and brought into action by the adaptive immune system. The complement system consists of a number of small proteins found in the blood, generally synthesized by the liver, and normally circulating as inactive precursors (proproteins). When stimulated by one of several triggers, proteases in the system cleave specific proteins to release cytokines and initiate an amplifying cascade of further cleavages. The end-result of this activation cascade is massive amplification of the response and activation of the cell-killing membrane attack complex. Over 25 proteins and protein fragments make up the complement system, including serum proteins, serosal proteins, and cell membrane receptors. They account for about 5% of the globulin fraction of blood serum. Three biochemical pathways activate the complement system: 1) The classical complement pathway, 2) The alternative complement pathway, 3) And the mannose-binding lectin pathway. The proteins and glycoproteins that constitute the complement system are synthesized by the liver hepatocytes. But significant amounts are also produced by tissue macrophages, blood monocytes, and epithelial cells of the genitourinal tract and gastrointestinal tract. The three pathways of activation all generate homologous variants of the protease C3-convertase. The classical complement pathway typically requires antigen:
Notes made by kozi1989 email:kozi1989@yahoo.gr
79
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
antibody complexes for activation (specific immune response), whereas the alternative and mannose-binding lectin pathways can be activated by C3 hydrolysis or antigens without the presence of antibodies (non-specific immune response). In all three pathways, a C3convertase cleaves and activates component C3, creating C3a and C3b, and causing a cascade of further cleavage and activation events. C3b binds to the surface of pathogens, leading to greater internalization by phagocytic cells by opsonization. C5a is an important chemotactic protein, helping recruit inflammatory cells. C3a is the precusor of an important cytokine (adipokine) named ASP and is usually rapidly cleaved by carboxypeptidase B. Both C3a and C5a have anaphylatoxin activity, directly triggering degranulation of mast cells as well as increasing vascular permeability and smooth muscle contraction. C5b initiates the membrane attack pathway, which results in the membrane attack complex (MAC), consisting of C5b, C6, C7, C8, and polymeric C9.[2] MAC is the cytolytic endproduct of the complement cascade; it forms a transmembrane channel, which causes osmotic lysis of the target cell. Kupffer cells and other macrophage cell types help clear complement-coated pathogens. As part of the innate immune system, elements of the complement cascade can be found in species earlier than vertebrates; most recently in the protostome horseshoe crab species, putting the origins of the system back further than was previously thought. The following are the basic functions of the complement: 1) 2) 3) 4) 5) Opsonization - enhancing phagocytosis of antigens Chemotaxis - attracting macrophages and neutrophils Lysis - rupturing membranes of foreign cells Clumping of antigen-bearing agents Altering the molecular structure of viruses
It has three pathways: 1) Classical pathway 2) Alternative pathway 3) Lectin pathway
31. Cells involved in the immune response. T lymphocytes, B lymphocytes,
phagocytic and antigen-presenting cells. T cells or T lymphocytes belong to a group of white blood cells known as lymphocytes, and play a central role in cell-mediated immunity. They can be distinguished from other lymphocyte types, such as B cells and natural killer cells (NK cells) by the presence of a special receptor on their cell surface called T cell receptors (TCR). The abbreviation T, in T cell, stands for thymus, since this is the principal organ responsible for the T cell's maturation. Several different subsets of T cells have been discovered, each with a distinct function. T helper cell (TH cells) assist other white blood cells in immunologic processes, including maturation of B cells into plasma cells and B memory cells, and activation of
Notes made by kozi1989 email:kozi1989@yahoo.gr
80
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
cytotoxic T cells and macrophages, among other functions. These cells are also known as CD4+ T cells because they express the CD4 protein on their surface. Helper T cells become activated when they are presented with peptide antigens by MHC class II molecules that are expressed on the surface of Antigen Presenting Cells (APCs). Once activated, they divide rapidly and secrete small proteins called cytokines that regulate or assist in the active immune response. These cells can differentiate into one of several subtypes, including TH1, TH2, TH3, TH17, or TFH, which secrete different cytokines to facilitate a different type of immune response. The mechanism by which T cells are directed into a particular subtype is poorly understood, though signalling patterns from the APC are thought to play an important role. Cytotoxic T cells (TC cells, or CTLs) destroy virally infected cells and tumor cells, and are also implicated in transplant rejection. These cells are also known as CD8+ T cells since they express the CD8 glycoprotein at their surface. These cells recognize their targets by binding to antigen associated with MHC class I, which is present on the surface of nearly every cell of the body. Through IL-10, adenosine and other molecules secreted by regulatory T cells, the CD8+ cells can be inactivated to an anergic state, which prevent autoimmune diseases such as experimental autoimmune encephalomyelitis. Memory T cells are a subset of antigen-specific T cells that persist long-term after an infection has resolved. They quickly expand to large numbers of effector T cells upon reexposure to their cognate antigen, thus providing the immune system with "memory" against past infections. Memory T cells comprise two subtypes: central memory T cells (TCM cells) and effector memory T cells (TEM cells). Memory cells may be either CD4+ or CD8+. Memory T cells typically express the cell surface protein CD45RO. Regulatory T cells (Treg cells), formerly known as suppressor T cells, are crucial for the maintenance of immunological tolerance. Their major role is to shut down T cellmediated immunity toward the end of an immune reaction and to suppress auto-reactive T cells that escaped the process of negative selection in the thymus. Two major classes of CD4+ regulatory T cells have been described, including the naturally occurring Treg cells and the adaptive Treg cells. Naturally occurring Treg cells (also known as CD4+CD25+FoxP3+ Treg cells) arise in the thymus and have been linked to interactions between developing T cells with both myeloid (CD11c+) and plasmacytoid (CD123+) dendritic cells that have been activated with TSLP.[3][4] Whereas the adaptive Treg cells (also known as Tr1 cells or Th3 cells) may originate during a normal immune response. Naturally occurring Treg cells can be distinguished from other T cells by the presence of an intracellular molecule called FoxP3. Mutations of the FOXP3 gene can prevent regulatory T cell development, causing the fatal autoimmune disease IPEX. Natural killer T cells (NKT cells) are a special kind of lymphocyte that bridges the adaptive immune system with the innate immune system. Unlike conventional T cells that recognize peptide antigen presented by major histocompatibility complex (MHC) molecules, NKT cells recognize glycolipid antigen presented by a molecule called CD1d. Once activated, these cells can perform functions ascribed to both Th and Tc cells (i.e.,
Notes made by kozi1989 email:kozi1989@yahoo.gr
81
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
cytokine production and release of cytolytic/cell killing molecules). They are also able to recognize and eliminate some tumor cells and cells infected with herpes viruses. T cells (gamma delta T cells) represent a small subset of T cells that possess a distinct T cell receptor (TCR) on their surface. A majority of T cells have a TCR composed of two glycoprotein chains called - and - TCR chains. However, in T cells, the TCR is made up of one -chain and one -chain. This group of T cells is much less common (2% of total T cells) than the T cells, but are found at their highest abundance in the gut mucosa, within a population of lymphocytes known as intraepithelial lymphocytes (IELs). The antigenic molecules that activate T cells are still widely unknown. However, T cells are not MHC restricted and seem to be able to recognize whole proteins rather than requiring peptides to be presented by MHC molecules on antigen presenting cells. Some murine T cells recognize MHC class IB molecules though. Human V9/V2 T cells, which constitute the major T cell population in peripheral blood, are unique in that they specifically and rapidly respond to a set of non-peptidic phosphorylated isoprenoid precursors, collectively named phosphoantigens. Phosphoantigens are produced by virtually all living cells. The most common phosphoantigens from animal and human cells (including cancer cells) are isopentenyl pyrophosphate (IPP) and its isomer dimethylallyl pyrophosphate (DMAPP). Many microbes produce the highly active compound hydroxy-DMAPP (HMB-PP) and corresponding mononucleotide conjugates, in addition to IPP and DMAPP. Plant cells produce both types of phosphoantigens. Drugs activating human V9/V2 T cells comprise synthetic phosphoantigens and aminobisphosphonates, which up-regulate endogenous IPP/DMAPP. All T cells originate from haematopoietic stem cells in the bone marrow . Haematopoietic progenitors derived from haematopoietic stem cells populate the thymus and expand by cell division to generate a large population of immature thymocytes. The earliest thymocytes express neither CD4 nor CD8, and are therefore classed as doublenegative (CD4-CD8-) cells. As they progress through their development they become doublepositive thymocytes (CD4+CD8+), and finally mature to single-positive (CD4+CD8- or CD4CD8+) thymocytes that are then released from the thymus to peripheral tissues. B cells are lymphocytes that play a large role in the humoral immune response (as opposed to the cell-mediated immune response, which is governed by T cells). The principal functions of B cells are to make antibodies against antigens, perform the role of antigen-presenting cells (APCs) and eventually develop into memory B cells after activation by antigen interaction. B cells are an essential component of the adaptive immune system.The abbreviation "B", in B cell, comes from the bursa of Fabricius in birds, where they mature. In mammals, immature B cells are formed in the bone marrow, which is used as a backronym for the cells' name. Plasma B cells (also known as plasma cells) are large B cells that have been exposed to antigen and produce and secrete large amounts of antibodies, which assist in the destruction of microbes by binding to them and making them easier targets for phagocytes and activation of the complement system. They are sometimes referred to as antibody factories. An electron micrograph of these cells reveals large amounts of rough endoplasmic
Notes made by kozi1989 email:kozi1989@yahoo.gr
82
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
reticulum, responsible for synthesizing the antibody, in the cell's cytoplasm. These are short lived cells and undergo apoptosis when the inciting agent that induced immune response is eliminated. This occurs because of cessation of continuous exposure to various colony stimulating factors required for survival. Memory B cells are formed from activated B cells that are specific to the antigen encountered during the primary immune response. These cells are able to live for a long time, and can respond quickly following a second exposure to the same antigen. B-1 cells express IgM in greater quantities than IgG and their receptors show polyspecificity, meaning that they have low affinities for many different antigens. Polyspecific immunoglobulins often have a preference for other immunoglobulins, self antigens and common bacterial polysaccharides. B-1 cells are present in low numbers in the lymph nodes and spleen and are instead found predominantly in the peritoneal and pleural cavities. B-2 cells are the conventional B cells most texts refer to. Marginal-zone B cells Follicular B Cells Phagocytes are the white blood cells that protect the body by ingesting (phagocytosing) harmful foreign particles, bacteria, and dead or dying cells. Their name comes from the Greek phagein, "to eat" or "devour", and "-cyte", the suffix in biology denoting "cell", from the Greek kutos, "hollow vessel". They are essential for fighting infections and for subsequent immunity. Phagocytes are important throughout the animal kingdom and are highly developed within vertebrates. One litre of human blood contains about six billion phagocytes. Phagocytes occur in many species; some amoebae behave like macrophage phagocytes, which suggests that phagocytes appeared early in the evolution of life. Phagocytes of humans and other animals are called "professional" or "nonprofessional" depending on how effective they are at phagocytosis. The professional phagocytes include cells called neutrophils, monocytes, macrophages, dendritic cells, and mast cells. The main difference between professional and non-professional phagocytes is that the professional phagocytes have molecules called receptors on their surfaces that can detect harmful objects, such as bacteria, that are not normally found in the body. Phagocytes are crucial in fighting infections, as well as in maintaining healthy tissues by removing dead and dying cells that have reached the end of their lifespan. During an infection, chemical signals attract phagocytes to places where the pathogen has invaded the body. These chemicals may come from bacteria or from other phagocytes already present. The phagocytes move by a method called chemotaxis. When phagocytes come into contact with bacteria, the receptors on the phagocyte's surface will bind to them. This binding will lead to the engulfing of the bacteria by the phagocyte. Some phagocytes kill the ingested pathogen with oxidants and nitric oxide. After phagocytosis, macrophages and dendritic cells can also participate in antigen presentation, a process in
Notes made by kozi1989 email:kozi1989@yahoo.gr
83
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
which a phagocyte moves parts of the ingested material back to its surface. This material is then displayed to other cells of the immune system. Some phagocytes then travel to the body's lymph nodes and display the material to white blood cells called lymphocytes. This process is important in building immunity. However, many pathogens have evolved methods to evade attacks by phagocytes. An antigen-presenting cell (APC) or accessory cell is a cell that displays foreign antigen complexes with major histocompatibility complex (MHC) on their surfaces. T-cells may recognize these complexes using their T-cell receptors (TCRs). These cells process antigens and present them to T-cells. T cells cannot recognise, and therefore react to, 'free' antigen. T cells can only 'see' antigen that has been processed and presented by cells via an MHC molecule. Most cells in the body can present antigen to CD8+ T cells via MHC class I molecules and, thus, act as "APCs"; however, the term is often limited to those specialized cells that can prime T cells (i.e., activate a T cell that has not been exposed to antigen, termed a naive T cell). These cells, in general, express MHC class II as well as MHC class I molecules, and can stimulate CD4+ ("helper") cells as well as CD8+ ("cytotoxic") T cells, respectively.To help distinguish between the two types of APCs, those that express MHC class II molecules are often called professional antigen-presenting cells. Professional APCs are very efficient at internalizing antigen, either by phagocytosis or by receptor-mediated endocytosis, and then displaying a fragment of the antigen, bound to a class II MHC molecule, on their membrane. The T cell recognizes and interacts with the antigen-class II MHC molecule complex on the membrane of the antigen-presenting cell. An additional co-stimulatory signal is then produced by the antigen-presenting cell, leading to activation of the T cell. The expression of co-stimulatory molecules is a defining feature of professional APCs.There are three main types of professional antigen-presenting cell: 1) Dendritic cells (DCs), which have the broadest range of antigen presentation, and are probably the most important APC. Activated DCs are especially potent TH cell activators because, as part of their composition, they express co-stimulatory molecules such as B7. 2) Macrophages, which are also CD4+ and are therefore also susceptible to infection by HIV. 3) B-cells, which express (as B cell receptor) and secrete a specific antibody, can internalize the antigen, which bind to its BCR and present it incorporated to MHC II molecule, but are inefficient APC for most other antigens. A non-professional APC does not constitutively express the Major Histocompatibility Complex class II (MHC class II) proteins required for interaction with naive T cells; these are expressed only upon stimulation of the non-professional APC by certain cytokines such as IFN-. Non-professional APCs include:
Notes made by kozi1989 email:kozi1989@yahoo.gr
84
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
1) 2) 3) 4) 5) 6)
Fibroblasts (skin) Thymic epithelial cells Thyroid epithelial cells Glial cells (brain) Pancreatic beta cells Vascular endothelial cells
32. Human alloantigens. ABO(H) system. Lewis system. Lectins. Biosynthesis of the
A, B, H and Lewis antigens.
Notes made by kozi1989 email:kozi1989@yahoo.gr
85
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The ABO blood group system is the most important blood type system (or blood group system) in human blood transfusion. The associated anti-A antibodies and anti-B antibodies are usually IgM antibodies, which are usually produced in the first years of life by sensitization to environmental substances such as food, bacteria, and viruses. ABO blood types are also present in some animals, for example apes such as chimpanzees, bonobos, and gorillas. The Lewis antigen system is a human blood group system based upon genes on chromosome 19 q13.3, (FUT2) and 19p13.3 (FUT3), which both have fucosyltransferase activity. There are two main types of Lewis antigens, Lewis a and Lewis b. They are red cell antigens which are not produced by the erythrocyte itself. Instead, Lewis antigens are a component of exocrine epithelial secretions, and are subsequently adsorbed onto the surface of the erythrocyte. However, absorption can only take place if the Se (secreter) gene is also present. It is possible to have any combination of Lewis a, Lewis b, both or neither antigens. The most common are Lewis a negative and Lewis b positive Le(a-b+). Having both antigens
present on red cells is extremely rare and is thought to be caused by a weak expression of the secretor gene. The link between the Lewis blood group and secretion of the ABO blood group antigens was possibly the first example of multiple effects of a human gene.[2] The presence of fucosyltransferase converts the Lewis a antigen to Lewis b. This is the same fucosyltransferase which can convert membrane bound A, B or H antigens into soluble A, B and H, allowing the person to secrete them into body fluids. Therefore, people with Lewis a antigens will not secrete the A, B or H antigens (and are called ABH non-secretors), and the presence of the Lewis b antigen is only found in secreters. Lewis negative (Le a-, Le b-) can be either secretors or non-secretors. Lectins are sugar-binding proteins (not to be confused with glycoproteins, which are proteins containing sugar chains or residues) that are highly specific for their sugar moieties. They play a role in biological recognition phenomena involving cells and
Notes made by kozi1989 email:kozi1989@yahoo.gr
86
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
proteins. For example, some viruses use lectins to attach themselves to the cells of the host organism during infection. Lectins may be disabled by specific mono- and oligosaccharides, which bind to them and prevent their attachment to cell membranes. Lectins serve many different biological functions in animals, from the regulation of cell adhesion to glycoprotein synthesis and the control of protein levels in the blood. They may also bind soluble extracellular and intercellular glycoproteins. Some lectins are found on the surface of mammalian liver cells that specifically recognize galactose residues. It is believed that these cell-surface receptors are responsible for the removal of certain glycoproteins from the circulatory system. Another lectin is a receptor that recognizes hydrolytic enzymes containing mannose-6-phosphate, and targets these proteins for delivery to the lysosomes. I-cell disease is one type of defect in this particular system. Lectins are also known to play important roles in the immune system by recognizing carbohydrates that are found exclusively on pathogens, or that are inaccessible on host cells. Examples are the lectin complement activation pathway and mannose-binding lectin. The production of A, B and H antigens are controlled by the action of transferases. These transferases are enzymes that catalyze (or control) addition of specific sugars to the oligosaccharide chain. The H, A, or B genes each produce a different transferase, which adds a different specific sugar to the oligosaccharide chain.
33. Human alloantigens.Rhesus system. Immunological conflict between mother and fetus.
Origing and biological importance of alloantigens. The Rh (Rhesus) blood group system (including the Rh factor) is one of the currently 30 human blood group systems. It is clinically the most important blood group system after ABO. The Rh blood group system currently consists of 50 defined blood-group antigens, among which the 5 antigens D, C, c, E, and e are the most important ones . The commonlyused terms Rh factor, Rh positive and Rh negative refer to the D antigen only. Besides its role in blood transfusion, the Rh blood group system, in particular the D antigen, is a relevant cause of the hemolytic disease of the newborn or erythroblastosis fetalis for which prevention is key. An individual either has, or does not have, the "Rhesus factor" on the surface of their red blood cells. This term strictly refers only to the most immunogenic D antigen of the Rh blood group system, or the Rh- blood cell system. The status is usually indicated by Rh positive (Rh+, does have the D antigen) or Rh negative (Rh-, does not have the D antigen) suffix to the ABO blood type. However, other antigens of this blood group system are also clinically relevant. These antigens are listed separately (see below: Rh nomenclature). In contrast to the ABO blood group, immunization against Rh can generally only occur through blood transfusion or placental exposure during pregnancy. This situation occurs not so rare. The reason for it - the incompatibility of blood between mother and fetus of the so-called Rhesus factor. Rhesus factor (in another way -Rh-antigen or Rh-antigen) is a substance which is in the surface of red blood cells
Notes made by kozi1989 email:kozi1989@yahoo.gr
87
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
(erythrocytes). The meaning of Rh-antigen that He serves as the identification mark for the immune system, a kind "identity card". Some people have rhesus antigens (these are called Rh-positive), y Some - no (respectively - Rh-negative individuals). Let us for convenience and brevity, we denote the first Rh (+), and second - Rh (-). Is man Rh (+) or Rh (-) - depends on a combination of genes, referred to it by parents. Such genes three and their inheritance obeys classical laws of Mendel. Remember classic example of a school biology textbook? If not, no problem: physician-geneticist explain to you the these patterns if necessary. Threat Rh-conflict pregnancy exists only if the woman Rh-negative, and her husband (the father of the unborn child) - Rh-positive. When This fetus must necessarily inherit the Rh-antigen from the father. Although this situation does not always lead to rhesus conflict. All other combinations Rh-affiliation of the spouses in this regard are completely safe. Should that the first pregnancy of Rh negative women Rh (+)-often the result all ends well. In subsequent pregnancies the risk of similar conflict increases. Why is this happening, will become clear after the explanation mechanism of Rh-conflict. Essence Rhesus-conflict is as follows: during pregnancy, fetal erythrocytes pass through the placenta into the blood of the mother. Rhesus antigen, located on erythrocytes of the fetus is the "wrong ID" for the immune system of the mother. Fetal cells are recognized and subjected to alien destruction with the help of special proteins antibodies. Because fetal erythrocytes continue to flow in the maternal circulation, continues to develop destructive antibodies. Antibodies are "looking for" source of income "alien" cells, ie fetus. In their way stands the placenta. If antibodies able to break through her barrier, then they begin to destroy red blood cells fetus is in its own vessels. Thus there is a large number substance called bilirubin. Bilirubin can damage the brain of the fetus, paint his skin yellow (jaundice). Since erythrocytes the fetus is continuously destroyed, his liver and spleen are trying to accelerate development of new red blood cells, while increasing in size. Eventually and they can not cope. There comes a strong anoxia and starts a new round of heavy disturbances in the body of the fetus. In the worst cases, it ends with congenital fetal dropsy, which can lead to his death. In milder cases of Rh-conflict manifested after birth jaundice or anemia in the newborn. All these states are united medical term "hemolytic disease of the fetus and newborn. Treatment of this disease difficult, complex, sometimes you want to replace transfusion baby. The first pregnancy, the expectant mother's immune system meets with "outsiders" (Rh + fetal erythrocytes) for the first time. Therefore, antibodies produced not so much: about as long as necessary for the destruction of red blood cells entering the fetus. Rate of formation of antibodies also not too high (this is in fact the first meeting!). But after childbirth in a woman's body remained "memory cells", which in subsequent pregnancies will be able to "organize" a fast and powerful development of antibodies against the Rh factor. Therefore, the reaction of female immune system on fruit Rh antigen in the second
Notes made by kozi1989 email:kozi1989@yahoo.gr
88
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
and third pregnancy much faster than the first. Accordingly, the above and the risk of the fetus. (Same like above) Hemolytic disease of the newborn, also known as Hemolytic disease of the fetus and newborn, HDN, HDFN, or Erythroblastosis fetalis,[1] is an alloimmune condition that develops in a fetus, when the IgG molecules (one of the five main types of antibodies) produced by the mother pass through the placenta. Among these antibodies are some which attack the red blood cells in the fetal circulation; the red cells are broken down and the fetus can develop reticulocytosis and anaemia. This fetal disease ranges from mild to very severe, and fetal death from heart failure (hydrops fetalis) can occur. When the disease is moderate or severe, many erythroblasts are present in the fetal blood and so these forms of the disease can be called erythroblastosis fetalis (or erythroblastosis foetalis). An alloantigen is an antigen that is a part of an animal's self-recognition system. e.g., Major histocompatibility complex molecules. When injected into another animal, they trigger an immune response aimed at eliminating them. Therefore, it can be thought of as an antigen that is present in some members of the same species, but is not common to all members of that species. If an alloantigen is presented to a member of the same species that does not have the alloantigen, it will be recognized as foreign. They are the products of polymorphic genes Alloimmunity can occur 1) In the recipient after transfusions of fluids such as blood or plasma. 2) In the recipient after allografts (grafts). 3) In the fetus after maternal antibodies have passed through the placenta into the fetus, as in haemolytic disease of the newborn and fetomaternal alloimmune thrombocytopenia. Alloimmunity can be regulated by neonatal B cells.
34. Cellular interactions during the immune response. Activation of antigenpresenting cells. Activation of T lymphocytes. Activation of B lymphocytes. Antigen-presenting cells are cells that express surface MHC (Major histocompatibility complex) class II molecules. MHC class II molecules preferentially bind to T cell receptors associated with the CD4 surface molecule. Thus, APCs present antigen to the CD4+ T cells, the helper/inducer subset. Class I MHC molecules preferentially bind to T cell receptors associated with the CD8 surface molecule. Class I MHC molecules are present on the surface of all nucleated cells, thus allowing cells to present their internal antigens to cytotoxic T cells. This mechanism is critically important in host defense against intracellular pathogens such as viruses. T-cell activation requires two signals:
Notes made by kozi1989 email:kozi1989@yahoo.gr
89
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Engagement of the T-cell receptor by the antigen/MHC complex A second signals, usually transduced by direct cognate interaction (touching) between constimulatory cell surface molecules on the APC and T cell. T-cell activation resulting from the two signals leads to production of cytoplasmic and nuclear factors, resulting in gene activation and new DNA synthesis. Activation of CD4+ T cells occurs through the engagement of both the T cell receptor and CD28 on the T cell by the major histocompatibility complex (MHC) peptide and B7 family members on the APC, respectively. Both are required for production of an effective immune response; in the absence of CD28 co-stimulation, T-cell receptor signalling alone results in anergy. The signalling pathways downstream from both CD28 and the T cell receptor involve many proteins. The first signal is provided by binding of the T cell receptor to a short peptide presented by the major histocompatibility complex (MHC) on another cell . This ensures that only a T cell with a TCR specific to that peptide is activated. The partner cell is usually a professional antigen presenting cell (APC), usually a dendritic cell in the case of nave responses, although B cells and macrophages can be important APCs. The peptides presented to CD8+ T cells by MHC class I molecules are 8-9 amino acids in length; the peptides presented to CD4+ cells by MHC class II molecules are longer, as the ends of the binding cleft of the MHC class II molecule are open. The second signal comes from co-stimulation, in which surface receptors on the APC are induced by a relatively small number of stimuli, usually products of pathogens, but sometimes breakdown products of cells, such as necrotic-bodies or heat shock proteins. The only co-stimulatory receptor expressed constitutively by nave T cells is CD28, so costimulation for these cells comes from the CD80 and CD86 proteins, which together constitute the B7 protein, (B7.1 and B7.2 respectively) on the APC. Other receptors are expressed upon activation of the T cell, such as OX40 and ICOS, but these largely depend upon CD28 for their expression. The second signal licenses the T cell to respond to an antigen. Without it, the T cell becomes anergic, and it becomes more difficult for it to activate in future. This mechanism prevents inappropriate responses to self, as self-peptides will not usually be presented with suitable co-stimulation. The T cell receptor exists as a complex of several proteins. The actual T cell receptor is composed of two separate peptide chains, which are produced from the independent T cell receptor alpha and beta (TCR and TCR) genes. The other proteins in the complex are the CD3 proteins: CD3 and CD3 heterodimers and, most important, a CD3 homodimer, which has a total of six ITAM motifs. The ITAM motifs on the CD3 can be phosphorylated by Lck and in turn recruit ZAP-70. Lck and/or ZAP-70 can also phosphorylate the tyrosines on many other molecules, not least CD28, LAT and SLP-76, which allows the aggregation of signalling complexes around these proteins. Phosphorylated LAT recruits SLP-76 to the membrane, where it can then bring in PLC, VAV1, Itk and potentially PI3K. Both PLC- and PI3K act on PI(4,5)P2 on the inner leaflet of the membrane to create the active intermediaries diacylglycerol (DAG), inositol-1,4,5Notes made by kozi1989 email:kozi1989@yahoo.gr
90
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
trisphosphate (IP3), and phosphatidlyinositol-3,4,5-trisphosphate (PIP3). DAG binds and activates some PKCs, most important, in T cells PKC, a process important for activating the transcription factors NF-B and AP-1. IP3 is released from the membrane by PLC- and diffuses rapidly to activate receptors on the ER, which induce the release of calcium. The released calcium then activates calcineurin, and calcineurin in turn activates NFAT, which then translocates to the nucleus. NFAT is a transcription factor, which activates the transcription of a pleiotropic set of genes, most notable, IL-2, a cytokine that promotes long term proliferation of activated T cells. Vitamin D and its active metabolite calcitriol also plays a role in the activation of Tcells. Naive T cells have very low expression of the vitamin D receptor (VDR) and PLC-1. However activated T cell receptor signaling through a "non-classical" p38 pathway upregulates VDR expression and calcitriol activated VDR in turn up-regulates PLC-1 expression.[7] Furthermore activation of T cells first requires activation through the nonclassical pathway to increase expression of VDR and PLC-1 before activation through the classical pathway can proceed. This provides a delayed response mechanism where by the innate immune system is allowed time (~48 hrs) to clear an infection before the inflammatory T cell mediated adaptive immune response kicks in.[7] Hence the activation of naive T cells is dependent on adequate calcitriol levels. Finally T cells express CYP27B1, the enzyme required to convert calcidiol into calcitriol. B cell recognition of antigen is not the only element necessary for B cell activation (a combination of clonal proliferation and terminal differentiation into plasma cells). B cells that have not been exposed to antigen, also known as nave B cells, can be activated in a T cell-dependent or -independent manner. T cell-dependent activation Once a pathogen is ingested by an antigen-presenting cell such as a macrophage or dendritic cell, the pathogen's proteins are then digested to peptides and attached to a class II MHC protein. This complex is then moved to the outside of the cell membrane. The macrophage is now activated to deliver multiple signals to a specific T cell that recognizes the peptide presented. The T cell is then stimulated to produce autocrines (Refer to Autocrine signalling), resulting in the proliferation and differentiation to effector and memory T cells. Helper T cells (i.e. CD4+ T cells) then activate specific B cells through a phenomenon known as an Immunological synapse. Activated B cells subsequently produce antibodies which assist in inhibiting pathogens until phagocytes (i.e. macrophages, neutrophils) or the complement system for example clears the host of the pathogen(s). Most antigens are T-dependent, meaning T cell help is required for maximal antibody production. With a T-dependent antigen, the first signal comes from antigen cross linking the B cell receptor (BCR) and the second signal comes from co-stimulation provided by a T cell. T dependent antigens contain proteins that are presented on B cell Class II MHC to a special subtype of T cell called a Th2 cell. When a B cell processes and presents the same antigen to
Notes made by kozi1989 email:kozi1989@yahoo.gr
91
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
the primed Th cell, the T cell secretes cytokines that activate the B cell. These cytokines trigger B cell proliferation and differentiation into plasma cells. Isotype switching to IgG, IgA, and IgE and memory cell generation occur in response to T-dependent antigens. This isotype switching is known as Class Switch Recombination (CSR). Once this switch has occurred, that particular B cell will usually no longer make the earlier isotypes, IgM or IgD. T cell-independent activation Many antigens are T cell-independent in that they can deliver both of the signals to the B cell. Mice without a thymus (nude or athymic mice that do not produce any T cells) can respond to T independent antigens. Many bacteria have repeating carbohydrate epitopes that stimulate B cells, by cross-linking the IgM antigen receptors in the B cell, responding with IgM synthesis in the absence of T cell help. Conjugate vaccines are made to provide a stronger immune response against these foreign molecules. There are two types of T cell independent activation; Type 1 T cell-independent (polyclonal) activation, and type 2 T cellindependent activation (in which macrophages present several of the same antigen in a way that causes cross-linking of antibodies on the surface of B cells).
35. Phases in differentiation of immunocompetent cells. Primary and secondary
immune response. Major histocompatibility complex (MHC). Primary immune response: The response that occurs after a harmful antigen has been encountered for the first time. Slowly as the primary immune response progresses the quality of antibodies improve. The antibodies that are more effective in killing the invading antigens become predominant. Secondary immune response: Is a much quicker and more effective response that occurs after an previously encountered antigen reappears. The secondary immune response occurs after an antigen that has already been encountered reappears. For example, if someone had recovered from the flu and later encounters the same strain of that virus antibodies that were made specifically for that antigen will rise dramatically with almost no lag period. The major histocompatibility complex (MHC) is a large genomic region or gene family found in most vertebrates that encodes MHC molecules. MHC molecules play an important role in the immune system and autoimmunity. Proteins are continually synthesized and destroyed in the cell. These include normal proteins (self) and microbial pathogens (nonself). The MHC proteins act as "signposts" that serve to alert the immune system if foreign material is present inside a cell. They achieve this by displaying fragmented pieces or antigens on the host cell's surface. These antigens may be self or nonself. (The protein fragment is sometimes compared to a hot dog, and the MHC protein to the bun.[1]) The constitutive presentation of MHC:peptide on cell surfaces allows for pathogen surveillance by immune cells, usually a T cell or natural killer (NK) cell. If
Notes made by kozi1989 email:kozi1989@yahoo.gr
92
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
activating T or NK cell surface receptors recognize MHC:peptide through binding interactions, it can activate the immune cell and lead to the development of an immune response against the presented antigen. Because MHC genes must defend against a great diversity of microbes in the environment, the MHC molecules(coded for by the MHC genes) must be able to present a wide range of peptides. MHC genes achieve this through several mechanisms: (1) the MHC locus is polygenic, (2) MHC genes are highly polymorphic and numerous alleles have been described, and (3) several MHC genes are codominantly expressed. There are two general classes of MHC molecules: Class I and Class II. Class I MHC molecules are found on all nucleated cells and present peptides to cytotoxic T cells. Class II MHC molecules are found on certain immune cells themselves, chiefly macrophages, B cells and dendritic cells, collectively known as professional antigen-presenting cells (APCs). These APCs specialize in the uptake of pathogens and subsequent processing into peptide fragments within phagosomes. The Class II MHC molecules on APCs present these fragments to helper T cells, which stimulate an immune reaction from other cells. The best-known genes in the MHC region are the subset that encodes antigenpresenting proteins on the cell surface. In humans, these genes are referred to as human leukocyte antigen (HLA) genes; however people often use the abbreviation MHC to refer to HLA gene products. To clarify the usage, some of the biomedical literature uses HLA to refer specifically to the HLA protein molecules and reserves MHC for the region of the genome that encodes for this molecule.
36. Genetic control of the antibody synthesis and the intensity of the immune
response. Studies of the molecular biology of lymphoid cells have markedly increased our understanding of how millions of different antibodies can be synthesized by a single animal. To date, the most detailed understanding has been achieved for the mouse, primarily because of the relatively greater experimental availability of this species. These studies, as well as those involving other species, have shown that the complete genes for antibody polypeptide chains are assembled from disparate genetic elements which are originally widely separated in the genome. The assembly process itself, together with the coding information present in the germ line genetic elements, contributes to the diversity of structure (and thus combining specificities) shown by mature antibody molecules. Specifically, the diversity of structure characteristic of antibody variable regions is due to three distinct mechanisms: Innate variability of germ line genes Mismatching of individual gene segments during their somatic rearrangement leading to junctional diversity And somatic mutation in variable region genetic material during or after the rearrangement.
Notes made by kozi1989 email:kozi1989@yahoo.gr
93
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
These processes lead to the wide array of combining specificities that permit the humoral immune system of a mature animal to interact with essentially any non-self antigen which it encounters. Complex genetic rearrangements are also responsible for the class switching phenomenon long known to be characteristic of the humoral immune response. A form of homologous recombination between constant region genes, possibly mediated by specific "switching" enzymes, is now believed to be involved in this phenomenon. It is also currently believed that the restriction of gene rearrangement processes to one of the two possible chromosomes of a diploid pair in each cell is responsible for the phenomenon of allelic exclusion that has long been associated with the normal functioning of mammalian Bcells.
37. Transplantation immunity. Host versus graft reaction. Types of transplantation.
Primary and secondary rejection. Organ transplantation is the moving of an organ from one body to another or from a donor site on the patient's own body, for the purpose of replacing the recipient's damaged or absent organ. The emerging field of Regenerative medicine is allowing scientists and engineers to create organs to be re-grown from the patient's own cells (stem cells, or cells extracted from the failing organs). Organs and/or tissues that are transplanted within the same person's body are called autografts. Transplants that are performed between two subjects of the same species are called allografts. Allografts can either be from a living or cadaveric source. Organs that can be transplanted are the heart, kidneys, liver, lungs, pancreas, intestine, and thymus. Tissues include bones, tendons (both referred to as musculoskeletal grafts), cornea, skin, heart valves, and veins. Worldwide, the kidneys are the most commonly transplanted organs, followed closely by the liver and then the heart. The cornea and musculoskeletal grafts are the most commonly transplanted tissues; these outnumber organ transplants by more than tenfold. Organ donors may be living, or brain dead. Tissue may be recovered from donors who are cardiac dead - up to 24 hours past the cessation of heartbeat. Unlike organs, most tissues (with the exception of corneas) can be preserved and stored for up to five years, meaning they can be "banked". Types of transplant Autograft Transplant of tissue to the same person. Sometimes this is done with surplus tissue, or tissue that can regenerate, or tissues more desperately needed elsewhere (examples include skin grafts, vein extraction for CABG, etc.) Sometimes an autograft is done to remove the tissue and then treat it or the person, before returning it (examples include stem cell autograft and storing blood in advance of surgery). In a rotationplasty a distal joint is use to replace a more proximal one, typically a foot and ankle joint is used to replace a knee joint.
Notes made by kozi1989 email:kozi1989@yahoo.gr
94
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The a patient's foot is severed and reversed, the knee removed, and the tibia joined with the femur. Allograft and allotransplantation An allograft is a transplant of an organ or tissue between two genetically nonidentical members of the same species. Most human tissue and organ transplants are allografts. Due to the genetic difference between the organ and the recipient, the recipient's immune system will identify the organ as foreign and attempt to destroy it, causing transplant rejection. Isograft A subset of allografts in which organs or tissues are transplanted from a donor to a genetically identical recipient (such as an identical twin). Isografts are differentiated from other types of transplants because while they are anatomically identical to allografts, they do not trigger an immune response. Xenograft and xenotransplantation A transplant of organs or tissue from one species to another. An example are porcine heart valve transplants, which are quite common and successful. Another example is attempted piscine-primate (fish to non-human primate) transplant of islet (i.e. pancreatic or insular tissue) tissue. The latter research study was intended to pave the way for potential human use, if successful. However, xenotransplantion is often an extremely dangerous type of transplant because of the increased risk of non-compatibility, rejection, and disease carried in the tissue. This is a very serious type of transplant. Split transplants Sometimes a deceased-donor organ, usually a liver, may be divided between two recipients, especially an adult and a child. This is not usually a preferred option because the transplantation of a whole organ is more successful. Domino transplants This operation is usually performed on patients with cystic fibrosis because both lungs need to be replaced and it is a technically easier operation to replace the heart and lungs at the same time. As the recipient's native heart is usually healthy, it can be transplanted into someone else needing a heart transplant. That term is also used for a special form of liver transplant in which the recipient suffers from familial amyloidotic polyneuropathy, a disease where the liver slowly produces a protein that damages other organs. This patient's liver can be transplanted into an older patient who is likely to die from other causes before a problem arises. This term also refers to a series of living donor transplants in which one donor
Notes made by kozi1989 email:kozi1989@yahoo.gr
95
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
donates to the highest recipient on the waiting list and the transplant center utilizes that donation to facilitate multiple transplants. These other transplants are otherwise impossible due to blood-type or antibody barriers to transplantation. The "Good Samaritan" kidney is transplanted into one of the other recipients, whose donor in turn donates his or her kidney to an unrelated recipient. Depending on the patients on the waiting list, this has sometimes been repeated for up to six pairs, with the final donor donating to the patient at the top of the list. This method allows all organ recipients to get a transplant even if their living donor is not a match to them. This further benefits patients below any of these recipients on waiting lists, as they move closer to the top of the list for a deceased-donor organ. Johns Hopkins Medical Center in Baltimore and Northwestern University's Northwestern Memorial Hospital have received significant attention for pioneering transplants of this kind. Transplant rejection occurs when a transplanted organ or tissue is not accepted by the body of the transplant recipient. This is explained by the concept that the immune system of the recipient attacks the transplanted organ or tissue. This is expected to happen, because the immune system's purpose is to distinguish foreign material within the body and attempt to destroy it, just as it attempts to destroy infecting organisms such as bacteria and viruses. When possible, transplant rejection can be reduced through serotyping to determine the most appropriate donor-recipient match and through the use of immunosuppressant drugs. Hyperacute rejection Hyperacute rejection is a complement-mediated response in recipients with preexisting antibodies to the donor (for example, ABO blood type antibodies). Hyperacute rejection occurs within minutes after the transplant and must be immediately removed to prevent a severe systemic inflammatory response. Rapid agglutination of the blood occurs. This is a particular risk in kidney transplants, and so a prospective cytotoxic crossmatch is performed prior to kidney transplantation to ensure that antibodies to the donor are not present. Hyperacute rejection is analogous to a blood transfusion reaction as it is a humoralmediated immune response. For other organs, hyperacute rejection is prevented by transplanting only ABO-compatible grafts. Hyperacute rejection is not significant in liver allografts and cellular transplants because these tissues have remarkable regenerative abilities. Hyperacute rejection is the outcome of xenotransplanted organ in nonimmunosuppressed recipients. Acute rejection Acute rejection may begin as early as one week after transplantation (as opposed to hyperacute rejection, which is immediate). The risk of acute rejection is highest in the first three months after transplantation. However, acute rejection can also occur months to years after transplantation. A single episode of acute rejection is not a cause for concern if recognized and treated promptly, and rarely leads to organ failure. But recurrent episodes
Notes made by kozi1989 email:kozi1989@yahoo.gr
96
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
are associated with chronic rejection (see below). Acute rejection occurs to some degree in all transplants (except those between identical twins) unless the immune response is altered through the use of immunosuppressive drugs. It is caused by mismatched HLA, which are present on all cells of the body. There are a large number of different alleles of each HLA, so a perfect match between all HLA in the donor tissue and the recipient's body is extremely rare. Tissues such as the kidney or the liver which are highly vascularized (rich in blood vessels), are often the earliest victims of acute rejection. In fact, episodes of acute rejection occur in around 10-30% of all kidney transplants, and 50 to 60% of liver transplants. Damage to the endothelial lining of blood vessels is an early predictor of irreversible acute transplant rejection. The reason that acute rejection usually begins one week after transplantation is the delay in activation of the involved T-cells. Often transplanted organs are acquired from a cadaveric source (e.g. trauma victim) and as a result of ischemia and/or trauma are already in a state of inflammation. The inflammatory response results in donor-derived dendritic cells migrating to the secondary lymphoid tissues (e.g. lymph node) of the recipient. There they present self-antigen derived from the donated organ to recipient T-cells. T-cells that interact with allogeneic HLA complexes have the potential to become activated and develop an immune response against the 1.) self-peptide, 2.) the allogeneic HLA molecule itself, or 3.) a combination of both. These T-cells must differentiate before the alloreaction begins and the tissue is rejected. The alloreactive T-cells cause cells in the transplanted tissue to lyse, or produce cytokines that cause necrosis of the transplanted tissue. This process can take days, or even weeks to manifest. The diagnosis of acute rejection relies on clinical data, including patient signs and symptoms, laboratory testing and ultimately a tissue biopsy. The biopsy is interpreted by a pathologist who notes changes in the tissue that suggest rejection. Generally the pathologist looks for three main histological features. First, the presence of T-cells infiltrating the transplanted tissue; these may be accompanied by a heterogeneous collection of other cell types including eosinophils, plasma cells and neutrophils. (The proportions of these cell types may be helpful in diagnosing the exact type of rejection.) Secondly, evidence of structural injury to the transplanted tissue; the characteristics of this injury will depend on the type of tissue being transplanted. Lastly, injury to the blood vessels in the transplanted tissue. Recent technological advancements have led to genetic expression testing in the form of a blood test. These tests, such as AlloMap Molecular Expression Testing have a high negative predictive value help manage the ACR rejection in transplant patients. These genetic expression tests are specific to the transplanted organ type. Chronic rejection The term "chronic rejection" was initially a term used to describe a long-term loss of function in transplanted organs, associated with fibrosis of the internal blood vessels of
Notes made by kozi1989 email:kozi1989@yahoo.gr
97
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
the transplanted tissue. But this pathology is now termed chronic allograft vasculopathy. The term chronic rejection is reserved for cases of transplant rejection where the rejection is due to a poorly understood chronic inflammatory and immune response against the transplanted tissue. Primary rejection: a rejection occurring more than 7 days after transplantation, mainly from a cellular immune response. Secondary rejection is the failure of an organ or tissue graft in a host who is already immune to the histocompatibility antigens of the graft because of a previous graft with the same antigens.
38. Graft versus host reaction. Anti-tumor immunity. Runt disease, homologous
wasting disease. Tumor antigens and anti-tumor defence. Graft-versus-host disease (GVHD) is a common complication of allogeneic bone marrow transplantation in which functional immune cells in the transplanted marrow recognize the recipient as "foreign" and mount an immunologic attack. It can also take place in a blood transfusion under certain circumstances. Causes According to the 1966 Billingham Criteria, 3 criteria must be met in order for GVHD to occur.[1] 1) An immunocompetent graft is administered, with viable and functional immune cells. 2) The recipient is immunologically disparate - histoincompatible. 3) The recipient is immunocompromised and therefore cannot destroy or inactivate the transplanted cells. After bone marrow transplantation, T cells present in the graft, either as contaminants or intentionally introduced into the host, attack the tissues of the transplant recipient after perceiving host tissues as antigenically foreign. The T cells produce an excess of cytokines, including TNF- and interferon-gamma (IFN). A wide range of host antigens can initiate graft-versus-host-disease, among them the human leukocyte antigens (HLAs). However, graft-versus-host disease can occur even when HLA-identical siblings are the donors. HLAidentical siblings or HLA-identical unrelated donors often have genetically different proteins (called minor histocompatibility antigens) that can be presented by MHC molecules to the donor's T-cells, which see these antigens as foreign and so mount an immune response. While donor T-cells are undesirable as effector cells of graft-versus-host-disease, they are valuable for engraftment by preventing the recipient's residual immune system from rejecting the bone marrow graft (host-versus-graft). In addition, as bone marrow transplantation is frequently used to treat cancer, mainly leukemias, donor T-cells have
Notes made by kozi1989 email:kozi1989@yahoo.gr
98
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
proven to have a valuable graft-versus-tumor effect. A great deal of current research on allogeneic bone marrow transplantation involves attempts to separate the undesirable graftvs-host-disease aspects of T-cell physiology from the desirable graft-versus-tumor effect. Types In the clinical setting, graft-versus-host-disease is divided into acute and chronic forms. The acute or fulminant form of the disease (aGVHD) is normally observed within the first 100 days post-transplant,[2] and is a major challenge to transplants owing to associated morbidity and mortality. The chronic form of graft-versus-host-disease (cGVHD) normally occurs after 100 days. The appearance of moderate to severe cases of cGVHD adversely influences long-term survival. This distinction is not arbitrary: Acute and chronic graft-versus-host-disease appear to involve different immune cell subsets, different cytokine profiles, somewhat different host targets, and respond differently to treatment. Homologous wasting disease: A term used to describe the disease state resulting from a graft versus host reaction. Graft versus host reaction occurs when the immune system of a transplant patient attacks the transplanted tissue but in homologous wasting disease the immune cells in the transplanted tissue actually attacks the host tissues. The condition occurs most often after a bone marrow transplant. Tumor antigen is an antigenic substance produced in tumor cells, i.e., it triggers an immune response in the host. Tumor antigens are useful in identifying tumor cells and are potential candidates for use in cancer therapy. Normal proteins in the body are not antigenic because of self-tolerance, a process in which self-reacting cytotoxic T lymphocytes (CTLs) and autoantibody-producing B lymphocytes are culled "centrally" in primary lymphatic tissue (BM) and "peripherally" in secondary lymphatic tissue (mostly thymus for T-cells and spleen/lymph nodes for B cells). Thus any protein that is not exposed to the immune system triggers an immune response. This may include normal proteins that are well sequestered from the immune system, proteins that are normally produced in extremely small quantities, proteins that are normally produced only in certain stages of development, or proteins whose structure is modified due to mutation. Classification of tumor antigens Initially they were broadly classified into two categories based on their pattern of expression: Tumor-Specific Antigens (TSA), which are present only on tumor cells and not on any other cell and Tumor-Associated Antigens (TAA), which are present on some tumor cells and also some normal cells.
Notes made by kozi1989 email:kozi1989@yahoo.gr
99
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
This classification, however, is imperfect because many antigens thought to be tumor-specific turned out to be expressed on some normal cells as well. The modern classification of tumor antigens is based on their molecular structure and source. Accordingly they can be classified as: 1) Products of Mutated Oncogenes and Tumor Suppressor Genes 2) Products of Other Mutated Genes Overexpressed or Aberrantly Expressed Cellular Proteins 3) Tumor Antigens Produced by Oncogenic Viruses 4) Oncofetal Antigens 5) Altered Cell Surface Glycolipids and Glycoproteins 6) Cell Type-Specific Differentiation Antigens Types Any protein produced in a tumor cell that has an abnormal structure due to mutation can act as a tumor antigen. Such abnormal proteins are produced due to mutation of the concerned gene. Mutation of protooncogenes and tumor suppressors which lead to abnormal protein production are the cause of the tumor and thus such abnormal proteins are called tumor-specific antigens. Examples of tumor-specific antigens include the abnormal products of ras and p53 genes. In contrast, mutation of other genes unrelated to the tumor formation may lead to synthesis of abnormal proteins which are called tumor-associated antigens. Proteins that are normally produced in very low quantities but whose production is dramatically increased in tumor cells, trigger an immune response. An example of such a protein is the enzyme tyrosinase, which is required for melanin production. Normally tyrosinase is produced in minute quantities but its levels are very much elevated in melanoma cells. Oncofetal antigens are another important class of tumor antigens. Examples are alphafetoprotein (AFP) and carcinoembryonic antigen (CEA). These proteins are normally produced in the early stages of embryonic development and disappear by the time the immune system is fully developed. Thus self-tolerance does not develop against these antigens. Abnormal proteins are also produced by cells infected with oncoviruses, e.g. EBV and HPV. Cells infected by these viruses contain latent viral DNA which is transcribed and the resulting protein produces an immune response. In addition to proteins, other substances like cell surface glycolipids and glycoproteins may also have an abnormal structure in tumor cells and could thus be targets of the immune system. Another important role of the immune system is to identify and eliminate tumors. The transformed cells of tumors express antigens that are not found on normal cells. To the
Notes made by kozi1989 email:kozi1989@yahoo.gr
100
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
immune system, these antigens appear foreign, and their presence causes immune cells to attack the transformed tumor cells. The antigens expressed by tumors have several sources;[83] some are derived from oncogenic viruses like human papillomavirus, which causes cervical cancer,[84] while others are the organism's own proteins that occur at low levels in normal cells but reach high levels in tumor cells. One example is an enzyme called tyrosinase that, when expressed at high levels, transforms certain skin cells (e.g. melanocytes) into tumors called melanomas.[85][86] A third possible source of tumor antigens are proteins normally important for regulating cell growth and survival, that commonly mutate into cancer inducing molecules called oncogenes.[83][87][88] The main response of the immune system to tumors is to destroy the abnormal cells using killer T cells, sometimes with the assistance of helper T cells.[ 86][89] Tumor antigens are presented on MHC class I molecules in a similar way to viral antigens. This allows killer T cells to recognize the tumor cell as abnormal.[90] NK cells also kill tumorous cells in a similar way, especially if the tumor cells have fewer MHC class I molecules on their surface than normal; this is a common phenomenon with tumors.[91] Sometimes antibodies are generated against tumor cells allowing for their destruction by the complement system. Clearly, some tumors evade the immune system and go on to become cancers.[92] Tumor cells often have a reduced number of MHC class I molecules on their surface, thus avoiding detection by killer T cells.[90] Some tumor cells also release products that inhibit the immune response; for example by secreting the cytokine TGF-, which suppresses the activity of macrophages and lymphocytes.[93] In addition, immunological tolerance may develop against tumor antigens, so the immune system no longer attacks the tumor cells.[92] Paradoxically, macrophages can promote tumor growth when tumor cells send out cytokines that attract macrophages, which then generate cytokines and growth factors that nurture tumor development. In addition, a combination of hypoxia in the tumor and a cytokine produced by macrophages induces tumor cells to decrease production of a protein that blocks metastasis and thereby assists spread of cancer cells.
39. Regulation and control of the immune processes. Immune tolerance.
An immune system is a system of biological structures and processes within an organism that protects against disease by identifying and killing pathogens and tumor cells. It detects a wide variety of agents, from viruses to parasitic worms, and needs to distinguish them from the organism's own healthy cells and tissues in order to function properly. Detection is complicated as pathogens can evolve rapidly, and adapt to avoid the immune system and allow the pathogens to successfully infect their hosts. Hormones can act as immunomodulators, altering the sensitivity of the immune system. For example, female sex hormones are known immunostimulators of both adaptive[95] and innate immune responses.[96] Some autoimmune diseases such as lupus erythematosus strike women preferentially, and their onset often coincides with puberty. By
Notes made by kozi1989 email:kozi1989@yahoo.gr
101
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
contrast, male sex hormones such as testosterone seem to be immunosuppressive.[97] Other hormones appear to regulate the immune system as well, most notably prolactin, growth hormone and vitamin D. When a T-cell encounters a foreign pathogen, it extends a vitamin D receptor. This is essentially a signaling device that allows the T-cell to bind to the active form of vitamin D, the steroid hormone calcitriol. T-cells have a symbiotic relationship with vitamin D. Not only does the T-cell extend a vitamin D receptor, in essence asking to bind to the steroid hormone version of vitamin D, calcitriol, but the T-cell expresses the gene CYP27B1, which is the gene responsible for converting the pre-hormone version of vitamin D, calcidiol into the steroid hormone version, calcitriol. Only after binding to calcitriol can T-cells perform their intended function. Other immune system cells that are known to express CYP27B1 and thus activate vitamin D calcidiol, are dendritic cells, keratinocytes and macrophages. It is conjectured that a progressive decline in hormone levels with age is partially responsible for weakened immune responses in aging individuals.[102] Conversely, some hormones are regulated by the immune system, notably thyroid hormone activity.[103] The age-related decline in immune function is also related to dropping vitamin D levels in the elderly. As people age, two things happen that negatively affect their vitamin D levels. First, they stay indoors more due to decreased activity levels. This means that they get less sun and therefore produce less cholecalciferol via UVB radiation. Second, as a person ages the skin becomes less adept at producing vitamin D.[104] The immune system is affected by sleep and rest, and sleep deprivation is detrimental to immune function.[106] Complex feedback loops involving cytokines, such as interleukin-1 and tumor necrosis factor- produced in response to infection, appear to also play a role in the regulation of non-rapid eye movement (REM) sleep.[107] Thus the immune response to infection may result in changes to the sleep cycle, including an increase in slowwave sleep relative to REM sleep.[108] Immune tolerance or immunological tolerance is the process by which the immune system does not attack an antigen. It can be either 'natural' or 'self tolerance', where the body does not mount an immune response to self antigens, or 'induced tolerance', where tolerance to external antigens can be created by manipulating the immune system . It occurs in three forms: central tolerance, peripheral tolerance and acquired tolerance. Genetic defects in these processes lead to autoimmunity, such as in Autoimmune polyendocrine syndrome type 1 (APS-1) and immunodysregulation polyendocrinopathy enteropathy X-linked syndrome (IPEX). Central tolerance Central tolerance occurs during lymphocyte development and operates in the thymus and bone marrow. Here, T and B lymphocytes that recognize self antigens are deleted before they develop into fully immunocompetent cells, preventing autoimmunity. This process is most active in fetal life, but continues throughout life as immature
Notes made by kozi1989 email:kozi1989@yahoo.gr
102
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
lymphocytes are generated. In mammals the process occurs in the thymus (T cells)[1][2] and bone marrow (B cells), when maturing lymphocytes are exposed to self antigens. Self antigens are present in both organs due to endogenous expression within the organ and importation of antigen due to circulation from peripheral sites. In the case of T cell central tolerance, additional sources of antigen are made available in the thymus by the action of the transcription factor AIRE. Positive selection occurs first when naive T-cells are exposed to antigens in the thymus. T-cells which have receptors with sufficient affinity for self-MHC molecules are selected. Other cells that do not show sufficient affinity to self-antigens will undergo a deletion process known as death by neglect which involves apoptosis of the cells. The positive selection is a classical example of the importance of some degree of autorreactiveness. This does not occur in B-cells. Negative selection of T-cells with a very high affinity of self-MHC molecules are induced to anergy, or lineage divergence to form T-regulatory cells. Peripheral tolerance Peripheral tolerance is immunological tolerance developed after T and B cells mature and enter the periphery. Acquired tolerance Acquired or induced tolerance refers to the immune system's adaptation to external antigens characterized by a specific non-reactivity of the lymphoid tissues to a given antigen that in other circumstances would likely induce cell-mediated or humoral immunity. One of the most important natural kinds of acquired tolerance is immune tolerance in pregnancy, where the fetus and the placenta must be tolerated by the maternal immune system. In adults, tolerance may be induced by repeated administration of very large doses of antigen, or of small doses that are below the threshold required for stimulation of an immune response (reference needed). Tolerance is most readily induced by soluble antigens administered either intravenously or sublingually, but specially orally (reference needed). Immunosuppression also facilitates the induction of tolerance (reference needed). In clinical practice, acquired immunity is important in organ transplantation, when the body must be forced to accept an organ from another individual. The failure of the body to accept an organ is known as transplant rejection. To prevent rejection, a variety of medicines are used to produce induced tolerance. One of the most important forms of acquired tolerance is oral tolerance.[3] Oral tolerance, the specific suppression of cellular and/or humoral immune reactivity to an antigen by prior administration of the antigen by the oral route, probably evolved to prevent hypersensitivity reactions to food proteins and bacterial antigens present in the mucosal flora.[4] It is of immense immunological importance, since it is a continuous natural immunologic event driven by exogenous antigen. Due to their privileged access to the
Notes made by kozi1989 email:kozi1989@yahoo.gr
103
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
internal milieu, antigens that continuously contact the mucosa represent a frontier between foreign and self components. Oral tolerance evolved to treat external agents that gain access to the body via a natural route as internal components without danger signals, which then become part of self. Failure of oral tolerance is attributed to the development and pathogenesis of several immunologically based diseases, including inflammatory bowel disease (Crohn's disease and ulcerative colitis).
40. Immunity theories. Evolution of immunity.
Immune systems are generally characterized by their ability to distinguish between self and non-self cells, tissues, or molecules, and to eliminate the non-self. The mammalian immune system is an incredibly complex and intricate system that can recognize non-self and provide protection from a wide variety of pathogens. While there is a high degree of interconnectivity between its components, the immune system can be loosely divided into two subsystems, the innate and the adaptive immune systems. In general, innate immunity is a non-specific, inducible response to pathogens. It is immediate in action, yet short-lived. On the other hand, the adaptive immune system is much more specific, but takes longer to activate. It also features immunological memory, and can augment itself to respond more quickly and with greater specificity to future infections of similar pathogens. Both systems work together to provide protection against a diverse and rapidly-evolving array of pathogens. While much is known about the evolution of the immune system, the details of its origin remain to be elucidated. The innate immune system is the more ancient of the two systems, with roots deep in the deuterostome branch of the bilaterians, roughly one billion years ago. Immunity and the immune system is often talked about but is always a very vague subject involving various organisms that the average person cannot see let alone examine for themselves. Invariably, in technical discussions antibodies and similar organisms are discussed in great detail. No one will dispute that in our blood streams there course many substances and organisms borne of our body that help to protect us from the foreign intrusion of intrusive or even dangerous organisms that could be injurious to our bodies. *White Blood Cells *Macrophages, *T- Cells, These definitely do their work to protect us and are part of the body by design. No one had to put them there and no one can buy these organisms in a product and ingest them to strengthen their effect. In fact where similar organisms to be borrowed from another body and placed into our own they would likely be killed off by the bodies own
Notes made by kozi1989 email:kozi1989@yahoo.gr
104
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
organisms of defense. There is no specific organ called the immune organ. If these organisms are the army within the body that defends it they can then in isolation be called the immune system! These however do not function in isolation.
41. Reproductive biology. Sex determination and differentiation.
Sex-determination system is a biological system that determines the development of sexual characteristics in an organism. Most sexual organisms have two sexes. In many cases, sex determination is genetic: males and females have different alleles or even different genes that specify their sexual morphology. In animals, this is often accompanied by chromosomal differences. In other cases, sex is determined by environmental variables (such as temperature) or social variables (the size of an organism relative to other members of its population). The details of some sex-determination systems are not yet fully understood. The XX/XY sex-determination system is the most familiar sex-determination systems, as it is found in humans. In the system, females have two of the same kind of sex chromosome (XX), while males have two distinct sex chromosomes (XY). Some species (including humans) have a gene SRY on the Y chromosome that determines maleness; others (such as the fruit fly) use the presence of two X chromosomes to determine femaleness. The XY sex chromosomes are different in shape and size from each other, unlike the autosomes, and are termed allosomes. The XX/XY system is also found in most other mammals, as well as some insects. However, at least one monotreme, the platypus, presents a particular sex determination scheme that in some ways resembles that of the ZW sex chromosomes of birds, and also lacks the SRY gene, whereas some rodents, such as several Arvicolinae (voles and lemmings), are also noted for their unusual sex determination systems. The platypus has ten sex chromosomes; males have an XYXYXYXYXY pattern while females have ten X chromosomes. Although it is an XY system, the platypus' sex chromosomes share no homologues with eutherian sex chromosomes.Instead, homologues with eutherian sex chromosomes lie on the platypus chromosome 6, which means that the eutherian sex chromosomes were autosomes at the time that the monotremes diverged from the therian mammals (marsupials and eutherian mammals). Sexual differentiation is the process of development of the differences between males and females from an undifferentiated zygote (fertilized egg). As male and female individuals develop from zygotes into fetuses, into infants, children, adolescents, and eventually into adults, sex and gender differences at many levels develop: Genes, Chromosomes, Gonads, Hormones,
Notes made by kozi1989 email:kozi1989@yahoo.gr
105
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Anatomy, Psyche, Social behaviors. Sex differences range from nearly absolute to simply statistical. Sex-dichotomous differences are developments which are wholly characteristic of one sex only. Examples of sex-dichotomous differences include aspects of the sex-specific genital organs such as ovaries, a uterus or a phallic urethra. In contrast, sex-dimorphic differences are matters of degree (e.g., size of phallus). Some of these (e.g., stature, behaviors) are mainly statistical, with much overlap between male and female populations. Nevertheless, even the sex-dichotomous differences are not absolute in the human population, and there are individuals who are exceptions (e.g., males with a uterus, or females with an XY karyotype), or who exhibit biological and/or behavioral characteristics of both sexes. Sex differences may be induced by specific genes, by hormones, by anatomy, or by social learning. Some of the differences are entirely physical (e.g., presence of a uterus) and some differences are just as obviously purely a matter of social learning and custom (e.g., relative hair length). Many differences, though, such as gender identity, appear to be influenced by both biological and social factors ("nature" and "nurture"). The early stages of human differentiation appear to be quite similar to the same biological processes in other mammals and the interaction of genes, hormones and body structures is fairly well understood. In the first weeks of life, a fetus has no anatomic or hormonal sex, and only a karyotype distinguishes male from female. Specific genes induce gonadal differences, which produce hormonal differences, which cause anatomic differences, leading to psychological and behavioral differences, some of which are innate and some induced by the social environment. The various ways that genes, hormones, and upbringing affect different human behaviors and mental traits are difficult to test experimentally and charged with political conflict.
42. Reproductive biology. Haploid-diploid cycle. Cytological basis of sexual
reproduction. Meiosis. Sexual reproduction involves both haploid and diploid cells. Cells in higher eucaryotic organisms proliferate in the diploid phase to form a multicellular organism; only the gametes (the egg and the sperm) are haploid. These gametes reunite at fertilization to generate a diploid zygote, which will develop into a diploid organism. For simplicity, only one chromosome is shown for each gamete, and the sperm cell has been greatly enlarged. Sexual reproduction is the creation of a new organism by combining the genetic material of two organisms. The two main processes are: meiosis, involving the halving of
Notes made by kozi1989 email:kozi1989@yahoo.gr
106
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
the number of chromosomes; and fertilization, involving the fusion of two gametes and the restoration of the original number of chromosomes. During meiosis, the chromosomes of each pair usually cross over to achieve homologous recombination. Meiosis (pronounced /maoss/ ( listen)) is a special type of cell division necessary for sexual reproduction. In animals, meiosis produces gametes like sperm and egg cells, while in other organisms like fungi it generates spores. Meiosis begins with one diploid cell containing two copies of each chromosomeone from the organism's mother and one from its fatherand produces four haploid cells containing one copy of each chromosome. Each of the resulting chromosomes in the gamete cells is a unique mixture of maternal and paternal DNA, ensuring that offspring are genetically distinct from either parent. This gives rise to genetic diversity in sexually reproducing populations, which enables them to adapt during the course of evolution. Before meiosis, the cell's chromosomes are duplicated by a round of DNA replication. This leaves the maternal and paternal versions of each chromosome, called homologs, composed of two exact copies called sister chromatids and attached at the centromere region. In the beginning of meiosis, the maternal and paternal homologs pair to each other. Then they typically exchange parts by homologous recombination, leading to crossovers of DNA from the maternal version of the chromosome to the paternal version and vice versa. Spindle fibers bind to the centromeres of each pair of homologs and arrange the pairs at the spindle equator. Then the fibers pull the recombined homologs to opposite poles of the cell. As the chromosomes move away from the center, the cell divides into two daughter cells, each containing a haploid number of chromosomes composed of two chromatids. After the recombined maternal and paternal homologs have separated into the two daughter cells, a second round of cell division occurs. There meiosis ends as the two sister chromatids making up each homolog are separated and move into one of the four resulting gamete cells. Upon fertilization, for example when a sperm enters an egg cell, two gamete cells produced by meiosis fuse. The gamete from the mother and the gamete from the father each contribute half to the set of chromosomes that make up the new offsping's genome. Meiosis uses many of the same mechanisms as mitosis, a type of cell division used by eukaryotes like plants and animals to split one cell into two identical daughter cells. In all plants, and in many protists, meiosis results in the formation of spores, haploid cells that can divide vegetatively without undergoing fertilization. Some eukaryotes, like Bdelloid rotifers, have lost the ability to carry out meiosis and have acquired the ability to reproduce by parthenogenesis. Meiosis does not occur in archaea or bacteria, which reproduce via asexual processes such as binary fission. Process Because meiosis is a "one-way" process, it cannot be said to engage in a cell cycle as mitosis does. However, the preparatory steps that lead up to meiosis are identical in pattern and name to the interphase of the mitotic cell cycle.[citation needed]
Notes made by kozi1989 email:kozi1989@yahoo.gr
107
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Interphase is divided into three phases: Growth 1 (G1) phase: This is a very active period, where the cell synthesizes its vast array of proteins, including the enzymes and structural proteins it will need for growth . In G1 stage each of the chromosomes consists of a single (very long) molecule of DNA. In humans, at this point cells are 46 chromosomes, 2N, identical to somatic cells. Synthesis (S) phase: The genetic material is replicated: each of its chromosomes duplicates, so that each of the 46 chromosomes becomes a complex of two identical sister chromatids. The cell is still considered diploid because it still contains the same number of centromeres. The identical sister chromatids have not yet condensed into the densely packaged chromosomes visible with the light microscope. This will take place during prophase I in meiosis.[citation needed] Growth 2 (G2) phase: G2 phase as seen before mitosis is not present in Meiosis. Actually, the first four stages of prophase I in many respects correspond to the G2 phase of mitotic cell cycle. Interphase is followed by meiosis I and then meiosis II . Meiosis I consists of separating the pairs of homologous chromosome, each made up of two sister chromatids, into two cells. One entire haploid content of chromosomes is contained in each of the resulting daughter cells; the first meiotic division therefore reduces the ploidy of the original cell by a factor of 2. Meiosis II consists of decoupling each chromosome's sister strands (chromatids), and segregating the individual chromatids into haploid daughter cells. The two cells resulting from meiosis I divide during meiosis II, creating 4 haploid daughter cells. Meiosis I and II are each divided into prophase, metaphase, anaphase, and telophase stages, similar in purpose to their analogous subphases in the mitotic cell cycle. Therefore, meiosis includes the stages of meiosis I (prophase I, metaphase I, anaphase I, telophase I), and meiosis II (prophase II, metaphase II, anaphase II, telophase II). Meiosis generates genetic diversity in two ways: (1) independent alignment and subsequent separation of homologous chromosome pairs during the first meiotic division allows a random and independent selection of each chromosome segregates into each gamete; and (2) physical exchange of homologous chromosomal regions by homologous recombination during prophase I results in new combinations of DNA within chromosomes. Phases Meiosis is divided into meiosis I and meiosis II which are further divided into Karyokinesis I and Cytokinesis I & Karyokinesis II and Cytokinesis II respectively. Meiosis I separates homologous chromosomes, producing two haploid cells (N chromosomes, 23 in humans), so meiosis I is referred to as a reductional division. A regular diploid human cell contains 46 chromosomes and is considered 2N because it contains 23 pairs of homologous chromosomes. However, after meiosis I, although the cell contains 46
Notes made by kozi1989 email:kozi1989@yahoo.gr
108
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
chromatids, it is only considered as being N, with 23 chromosomes. This is because later, in Anaphase I, the sister chromatids will remain together as the spindle fibres pull the pair toward the pole of the new cell. In meiosis II, an equational division similar to mitosis will occur whereby the sister chromatids are finally split, creating a total of 4 haploid cells (23 chromosomes, N) - two from each daughter cell from the first division. Prophase I During prophase I, DNA is exchanged between homologous chromosomes in a process called homologous recombination. This often results in chromosomal crossover. The new combinations of DNA created during crossover are a significant source of genetic variation, and may result in beneficial new combinations of alleles. The paired and replicated chromosomes are called bivalents or tetrads, which have two chromosomes and four chromatids, with one chromosome coming from each parent. At this stage, non-sister chromatids may cross-over at points called chiasmata (plural; singular chiasma). Leptotene The first stage of prophase I is the leptotene stage, also known as leptonema, from Greek words meaning "thin threads". In this stage of prophase I, individual chromosomeseach consisting of two sister chromatidschange from the diffuse state they exist in during the cell's period of growth and gene expression, and condense into visible strands within the nucleus. However the two sister chromatids are still so tightly bound that they are indistinguishable from one another. During leptotene, lateral elements of the synaptonemal complex assemble.Leptotene is of very short duration and progressive condensation and coiling of chromosome fibers takes place. Chromosome assume a long thread like shape,they contract and become thick.At the beginning chromosomes are present in diploid number as in mitotic prophase.Each chromosome is made up of only one chromatid and half of the total chromosome are paternal and half maternal.For every paternal chromosome there is a corresponding maternal chromosome similar in size,shape and nature of inherited characters and are called homologous chromosome. Zygotene The zygotene stage, also known as zygonema, from Greek words meaning "paired threads", occurs as the chromosomes approximately line up with each other into homologous chromosome pairs. This is called the bouquet stage because of the way the telomeres cluster at one end of the nucleus. At this stage, the synapsis (pairing/coming together) of homologous chromosomes takes place, facilitated by assembly of central element of the synaptonemal complex.Pairing is brought about by a zipper like fashion and may start at the centromere(procentric),at the chromosome ends(proterminal),or at any other portion(intermediate).Individuals of a pair are equal in length and in position of centromere. Thus pairing is highly specific and exact.The paired chromosomes are called Bivalent or tetrad chromosome.
Notes made by kozi1989 email:kozi1989@yahoo.gr
109
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Pachytene The pachytene stage, also known as pachynema, from Greek words meaning "thick threads", is the stage when chromosomal crossover (crossing over) occurs. Nonsister chromatids of homologous chromosomes randomly exchange segments over regions of homology. Sex chromosomes, however, are not wholly identical, and only exchange information over a small region of homology. At the sites where exchange happens, chiasmata form. The exchange of information between the non-sister chromatids results in a recombination of information; each chromosome has the complete set of information it had before, and there are no gaps formed as a result of the process. Because the chromosomes cannot be distinguished in the synaptonemal complex, the actual act of crossing over is not perceivable through the microscope, and chiasmata are not visible until the next stage. Diplotene During the diplotene stage, also known as diplonema, from Greek words meaning "two threads", the synaptonemal complex degrades and homologous chromosomes separate from one another a little. The chromosomes themselves uncoil a bit, allowing some transcription of DNA. However, the homologous chromosomes of each bivalent remain tightly bound at chiasmata, the regions where crossing-over occurred. The chiasmata remain on the chromosomes until they are severed in anaphase I. In human fetal oogenesis all developing oocytes develop to this stage and stop before birth. This suspended state is referred to as the dictyotene stage and remains so until puberty. Diakinesis Chromosomes condense further during the diakinesis stage, from Greek words meaning "moving through". This is the first point in meiosis where the four parts of the tetrads are actually visible. Sites of crossing over entangle together, effectively overlapping, making chiasmata clearly visible. Other than this observation, the rest of the stage closely resembles prometaphase of mitosis; the nucleoli disappear, the nuclear membrane disintegrates into vesicles, and the meiotic spindle begins to form. Synchronous processes During these stages, two centrosomes, containing a pair of centrioles in animal cells, migrate to the two poles of the cell. These centrosomes, which were duplicated during S-phase, function as microtubule organizing centers nucleating microtubules, which are essentially cellular ropes and poles. The microtubules invade the nuclear region after the nuclear envelope disintegrates, attaching to the chromosomes at the kinetochore. The kinetochore functions as a motor, pulling the chromosome along the attached microtubule toward the originating centriole, like a train on a track. There are four kinetochores on each tetrad, but the pair of kinetochores on each sister chromatid fuses and functions as a unit during meiosis I.
Notes made by kozi1989 email:kozi1989@yahoo.gr
110
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Microtubules that attach to the kinetochores are known as kinetochore microtubules. Other microtubules will interact with microtubules from the opposite centriole: these are called nonkinetochore microtubules or polar microtubules. A third type of microtubules, the aster microtubules, radiates from the centrosome into the cytoplasm or contacts components of the membrane skeleton. Metaphase I Homologous pairs move together along the metaphase plate: As kinetochore microtubules from both centrioles attach to their respective kinetochores, the homologous chromosomes align along an equatorial plane that bisects the spindle, due to continuous counterbalancing forces exerted on the bivalents by the microtubules emanating from the two kinetochores of homologous chromosomes. The physical basis of the independent assortment of chromosomes is the random orientation of each bivalent along the metaphase plate, with respect to the orientation of the other bivalents along the same equatorial line. Anaphase I Kinetochore (bipolar spindles) microtubules shorten, severing the recombination nodules and pulling homologous chromosomes apart. Since each chromosome has only one functional unit of a pair of kinetochores,[4] whole chromosomes are pulled toward opposing poles, forming two haploid sets. Each chromosome still contains a pair of sister chromatids. Nonkinetochore microtubules lengthen, pushing the centrioles farther apart. The cell elongates in preparation for division down the center. Telophase I The last meiotic division effectively ends when the chromosomes arrive at the poles. Each daughter cell now has half the number of chromosomes but each chromosome consists of a pair of chromatids. The microtubules that make up the spindle network disappear, and a new nuclear membrane surrounds each haploid set. The chromosomes uncoil back into chromatin. Cytokinesis, the pinching of the cell membrane in animal cells or the formation of the cell wall in plant cells, occurs, completing the creation of two daughter cells. Sister chromatids remain attached during telophase I. Cells may enter a period of rest known as interkinesis or interphase II. No DNA replication occurs during this stage. Meiosis II Meiosis II is the second part of the meiotic process . Much of the process is similar to mitosis. The end result is production of four haploid cells (23 chromosomes, N in humans) from the two haploid cells (22 chromosomes, N * each of the chromosomes consisting of two sister chromatids) produced in meiosis I. The four main steps of Meiosis II are:
Notes made by kozi1989 email:kozi1989@yahoo.gr
111
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Prophase II, Metaphase II, Anaphase II, and Telophase II. In prophase II we see the disappearance of the nucleoli and the nuclear envelope again as well as the shortening and thickening of the chromatids. Centrioles move to the polar regions and arrange spindle fibers for the second meiotic division. In metaphase II, the centromeres contain two kinetochores that attach to spindle fibers from the centrosomes (centrioles) at each pole. The new equatorial metaphase plate is rotated by 90 degrees when compared to meiosis I, perpendicular to the previous plat. This is followed by anaphase II, where the centromeres are cleaved, allowing microtubules attached to the kinetochores to pull the sister chromatids apart. The sister chromatids by convention are now called sister chromosomes as they move toward opposing poles. The process ends with telophase II, which is similar to telophase I, and is marked by uncoiling and lengthening of the chromosomes and the disappearance of the spindle. Nuclear envelopes reform and cleavage or cell wall formation eventually produces a total of four daughter cells, each with a haploid set of chromosomes. Meiosis is now complete and ends up with four new daughter cells.
43. Reproductive biology. Gametogenesis. Spermatogenesis. Characteristic features
of male gametes. Origin of germ cells. Gametogenesis is a biological process by which diploid or haploid precursor cells undergo cell division and differentiation to form mature haploid gametes. Depending on the biological life cycle of the organism, gametogenesis occurs by meiotic division of diploid gametocytes into various gametes, or by mitotic division of haploid gametogenous cells. For example, plants produce gametes through mitosis in gametophytes. The gametophytes grow from haploid spores after sporic meiosis. The existence of a multicellular, haploid phase in the life cycle between meiosis and gametogenesis is also referred to as alternation of generations. Spermatogenesis is the process by which male primary germ cells undergo division, and produce a number of cells termed spermatogonia, from which the primary spermatocytes are derived. Each primary spermatocyte divides into two secondary spermatocytes, and each secondary spermatocyte into two spermatids or young spermatozoa. These develop into mature spermatozoa, also known as sperm cells. Thus, the primary spermatocyte gives rise to two cells, the secondary spermatocytes, and the two secondary spermatocytes by their subdivision produce four spermatozoa. Spermatozoa are the mature male gametes in many sexually reproducing organisms. Thus, spermatogenesis is the male version of gametogenesis. In mammals it occurs in the male testes and epididymis in a stepwise fashion, and for humans takes approximately 64 days.[2] Spermatogenesis is highly dependent upon optimal conditions for the process to occur correctly, and is essential for sexual reproduction. It starts at puberty
Notes made by kozi1989 email:kozi1989@yahoo.gr
112
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
and usually continues uninterrupted until death, although a slight decrease can be discerned in the quantity of produced sperm with increase in age. Stages Spermatocytogenesis Spermatocytogenesis is the male form of gametocytogenesis and results in the formation of spermatocytes possessing half the normal complement of genetic material. In spermatocytogenesis, a diploid spermatogonium which resides in the basal compartment of seminiferous tubules, divides mitotically to produce two diploid intermediate cells called primary spermatocytes. Each primary spermatocyte then moves into the adluminal compartment of the seminiferous tubules and duplicates its DNA and subsequently undergoes meiosis I to produce two diploid secondary spermatocytes, which will later divide once more into haploid cells. This division implicates sources of genetic variation, such as random inclusion of either parental chromosomes, and chromosomal crossover, to increase the genetic variability of the gamete. ` Each cell division from a spermatogonium to a spermatid is incomplete; the cells remain connected to one another by bridges of cytoplasm to allow synchronous development. It should also be noted that not all spermatogonia divide to produce spermatocytes, otherwise the supply would run out. Instead, certain types of spermatogonia divide to produce copies of themselves, thereby ensuring a constant supply of gametogonia to fuel spermatogenesis. Spermatidogenesis Spermatidogenesis is the creation of spermatids from secondary spermatocytes. Secondary spermatocytes produced earlier rapidly enter meiosis II and divide to produce haploid spermatids. The brevity of this stage means that secondary spermatocytes are rarely seen in histological preparations. Spermiogenesis During spermiogenesis, the spermatids begin to grow a tail, and develop a thickened mid-piece, where the mitochondria gather and form an axoneme. Spermatid DNA also undergoes packaging, becoming highly condensed. The DNA is packaged firstly with specific nuclear basic proteins, which are subsequently replaced with protamines during spermatid elongation. The resultant tightly packed chromatin is transcriptionally inactive. The Golgi apparatus surrounds the now condensed nucleus, becoming the acrosome. One of the centrioles of the cell elongates to become the tail of the sperm. Maturation then takes place under the influence of testosterone, which removes the remaining unnecessary cytoplasm and organelles. The excess cytoplasm, known as residual bodies, is phagocytosed by surrounding Sertoli cells in the testes. The resulting spermatozoa are now mature but lack motility, rendering them sterile. The mature
Notes made by kozi1989 email:kozi1989@yahoo.gr
113
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
spermatozoa are released from the protective Sertoli cells into the lumen of the seminiferous tubule in a process called spermiation. The non-motile spermatozoa are transported to the epididymis in testicular fluid secreted by the Sertoli cells with the aid of peristaltic contraction. While in the epididymis the spermatozoa gain motility and become capable of fertilization. However, transport of the mature spermatozoa through the remainder of the male reproductive system is achieved via muscle contraction rather than the spermatozoon's recently acquired motility. A spermatozoon (alternate spellings spermatozoan, spermatozon; plural spermatozoa) is a motile sperm cell, or moving form of the haploid cell that is the male gamete. (A non-motile sperm cell is called a spermatium.) A spermatozoon joins an ovum to form a zygote. (A zygote is a single cell, with a complete set of chromosomes, that normally develops into an embryo.) The term spermatozoon comes from the ancient Greek word (seed) and (living being). Sperm cells contribute approximately half of the nuclear genetic information to the diploid offspring. In mammals, the sex of the offspring is determined by the sperm cell: a spermatozoon bearing a Y-chromosome will lead to a male (XY) offspring, while one bearing an X-chromosome will lead to a female (XX) offspring (the ovum always provides an Xchromosome). Sperm cells were first observed by Anton van Leeuwenhoek in 1677.[1] In male humans, sperm cells consists of a head 5 m by 3 m and a tail 41 m long. The tail flagellates, which propels the sperm cell (at about 13 mm/minute in humans) by whipping in an elliptical cone.[2] Semen has an alkaline nature, and they do not reach full motility (hypermotility) until they reach the vagina where the alkaline pH is neutralized by acidic vaginal fluids. This gradual process takes 2030 minutes. In this time, fibrinogen from the seminal vesicles forms a clot, securing and protecting the sperm. Just as they become hypermotile, fibrinolysin from the prostate dissolves the clot, allowing the sperm to progress optimally. The spermatozoon is characterized by a minimum of cytoplasm and the most densely packed DNA known in eukaryotes. Compared to mitotic chromosomes in somatic cells, sperm DNA is at least sixfold more highly condensed.[3] The specimen contributes with DNA/chromatin, a centriole and perhaps also an oocyte-activating factor (OAF).[4] It may also contribute with paternal Messenger RNA (mRNA), also contributing to embryonic development.[4] A germ cell is any biological cell that gives rise to the gametes of an organism that reproduces sexually. In many animals, the germ cells originate near the gut of an embryo and migrate to the developing gonads. There, they undergo cell division of two types, mitosis and meiosis, followed by cellular differentiation into mature gametes, either eggs or sperm. Unlike animals, plants do not have germ cells set aside in early development. Instead, germ cells can come from somatic cells in the adult floral meristem.
Notes made by kozi1989 email:kozi1989@yahoo.gr
114
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
44. Reproductive biology. Oogenesis. Molecular mechanisms of oocyte maturation.
Characteristic features of mature ova. Oogenesis or ogenesis is the creation of an ovum (egg cell). It is the female form of gametogenesis. The male equivalent is spermatogenesis. It involves the development of the various stages of the immature ovum. In mammals, the first part of oogenesis starts in the germinal epithelium, which gives rise to the development of ovarian follicles, the functional unit of the ovary. Note that this process, important to all animal life cycles yet unlike all other instances of cell division, occurs completely without the aid of oo spindle-coordinating centrosomes.[citation needed] Oogenesis consists of several sub-processes: oocytogenesis, ootidogenesis, and finally maturation to form an ovum (oogenesis proper). Folliculogenesis is a separate subprocess that accompanies and supports all three oogenetic sub-processes. Oogonium --(Oocytogenesis)--> Primary Oocyte --(Meiosis I)-->First Polar Body (Discarded afterward) + Secondary oocyte --(Meiosis II)--> Secondary Polar Body (Discarded afterward) + Ovum The Creation of Oogonia The creation of oogonia traditionally doesn't belong to oogenesis proper but, instead, to the common process of gametogenesis, which in the female human begins with the processes of folliculogenesis, oocytogenesis, and ootidogenesis. Human Oogenesis At the start of the menstrual cycle, some 12-20 primary follicles begin to develop under the influence of elevated FSH to form secondary follicles. The primary follicles have formed from primordial follicles, which developed in the ovary at around 10 30 weeks after conception. By around day 9 of the cycle, only one healthy secondary follicle remains, with the rest having undergone cellular atresia. The remaining follicle is called the dominant follicle and is responsible for producing large amounts of oestradiol during the late follicular phase. Oestradiol production depends upon co-operation between the theca and granulosa cells. On day 14 of the cycle, an LH surge occurs, which itself is triggered by the positive feedback of oestradiol. This causes the secondary follicle to develop into a tertiary follicle, which then ovulates some 2436 hours later. An important event in the development of the tertiary follicle occurs when the primary oocyte completes the first meiotic division, resulting in the formation of a polar body and a secondary oocyte. The empty follicle then forms a corpus luteum.[citation needed] Oocytogenesis Oogenesis starts with the process of developing oogonia, which occurs via the transformation of primordial follicles into primary oocytes, a process called oocytogenesis.[2] Oocytogenesis is complete either before or shortly after birth.
Notes made by kozi1989 email:kozi1989@yahoo.gr
115
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Ootidogenesis The succeeding phase of ootidogenesis occurs when the primary oocyte develops into an ootid. This is achieved by the process of meiosis. In fact, a primary oocyte is, by its biological definition, a cell whose primary function is to divide by the process of meiosis.[6] However, although this process begins at prenatal age, it stops at prophase I. In late fetal life, all oocytes, still primary oocytes, have halted at this stage of development, called the dictyate. After menarche, these cells then continue to develop, although only a few do so every menstrual cycle. Meiosis I Meiosis I of ootidogenesis begins during embryonic development, but halts in the diplotene stage of prophase I until puberty. For those primary oocytes that continue to develop in each menstrual cycle, however, synapsis occurs and tetrads form, enabling Chromosomal crossover to occur. As a result of meiosis I, the primary oocyte has now developed into the secondary oocyte and the first polar body. Meiosis II Immediately after meiosis I, the haploid secondary oocyte initiates meiosis II. However, this process is also halted at the metaphase II stage until fertilization, if such should ever occur. When meiosis II has completed, an ootid and another polar body have now been created. Folliculogenesis Synchronously with ootidogenesis, the ovarian follicle surrounding the ootid has developed from a primordial follicle to a preovulatory one. Maturation into ovum Both polar bodies disintegrate at the end of Meiosis II, leaving only the ootid, which then eventually undergoes maturation into a mature ovum. The function of forming polar bodies is to discard the extra haploid sets of chromosomes that have resulted as a consequence of meiosis. The primary oocyte In the first week of the cycle the maturation of the oocyte in its associated follicle depends on the progress of the maturation of the surrounding follicle cells. The fittest follicle with its oocyte becomes the dominant follicle in the second cycle week and, later, a graafian follicle Up to just under two days before ovulation, the maturation of the oocyte consists in its ingestion of substances (growth of the yolk) that are supplied by the surrounding granulosa cells. This exchange of substances is mediated through cytoplasma processes of
Notes made by kozi1989 email:kozi1989@yahoo.gr
116
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
the granulosa cells that are anchored through the pellucid zone at the oocyte surface (Fig. 10). The oocyte nucleus [2n, 4C] is also matured in the last days before the LH peak. Up to that point it was arrested in the extremely elongated prophase (= dictyotene) of the first meiosis (the arrested condition that has existed since the fetal period). Through the maturation the nucleus changes in the diakinesis (of the prophase) and prepares itself for the completion of the first meiosis, which is triggered by the LH peak. The spindle apparatus for dividing the chromosomes has formed and oriented itself radially to the cellular surface. The first polar body will arise at the spot where the spindle apparatus is anchored on the cellular surface. Further, the processes of the granulosa cells have retracted from the oocyte surface into the pellucid zone. They have released themselves from the oocyte and this leads to the formation of the perivitelline space. In this space the ejection of the first polar body takes place as a sign that the first meiosis has ended. The secondary oocyte Through the effects of LH on the granulosa cells, these have begun to loosen their cellular bonds and to multiply. They now also produce progesterone that is released into the follicle fluid. Through the separation of the homologous chromosomes in the first meiosis a haploid (reduplicated) set of chromosomes (1n, 2C) is now to be found in the secondary oocyte. The first polar body also contains 1n, 2C. Via a fine cytoplasmic connection the polar body and oocyte remain bound together following the meiotic division, similar to what takes place when male gametes are formed. The follicle that is about to rupture Besides the hormones, the granulosa cells also secrete an extra-cellular matrix, mainly hyaluronic acid, into the follicle fluid. Before ovulation the follicle fluid volume increases markedly. The cumulus cell bonds loosen further. In this way, together with the enclosed oocyte, they free themselves from where they were attached to the follicular wall and now swim in the follicle fluid. The wreath of granulosa cells that enclose the oocyte is called the corona radiata. The oocyte has now ended all the steps of maturation that were set into motion by the LH peak. The molecular and structural preparations for the time following the penetration by the sperm cell have now been made in the cytoplasma. A spindle apparatus (2nd meiosis) has again been able to form with the chromosomes in the equatorial level (metaphase plate). The spindle is once more anchored radially to the cell membrane near the polar body. The same processes of spindle formation also take place in the polar body. The second meiosis is arrested in this position. The final steps of the maturation, namely the freeing for the second meiosis, are first completed by the secondary oocyte when the spermatozoon has penetrated the oocyte.
Notes made by kozi1989 email:kozi1989@yahoo.gr
117
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
An ovum (plural ova, from the Latin word ovum meaning egg or egg cell) is a haploid female reproductive cell or gamete. Both animals and embryophytes have ova. The term ovule is used for the young ovum of an animal, as well as the plant structure that carries the female gametophyte and egg cell and develops into a seed after fertilization. In lower plants and algae, the ovum is also often called oosphere.
45. Reproductive biology. Fertilization. External fertilization and mechanisms for
blocking polyspermy. Completion of fertilization. Fertilisation (also known as conception, fecundation and syngamy) is the fusion of gametes to produce a new organism. In animals, the process involves the fusion of an ovum with a sperm, which eventually leads to the development of an embryo. Depending on the animal species, the process can occur within the body of the female in internal fertilisation, or outside (external fertilisation). The entire process of development of new individuals is called reproduction. External fertilization is a form of fertilization in which a sperm cell is united with an egg cell external to the bodies of the reproducing individuals. In contrast, internal fertilization takes place inside the female after insemination through copulation. In sexual reproduction, there must be some way of getting the sperm to the egg. Since sperm are designed to be mobile in a watery environment (they have tails and are streamlined), aquatic animals can make use of the water in which they live. Many plants make use of external fertilization, especially ones without bright flowers or other means of attracting animals. In many aquatic animals such as coral or hydra, eggs and sperm are simultaneously shed into the water, and the sperm swim through the water to fertilize the egg in a process known as broadcast fertilization. In many fish species, including salmon, the female will deposit unfertilized eggs in the substrate and the male will swim by and fertilize them. External fertilization uses or needs thousands of sperm cells In biology, polyspermy describes an egg that has been fertilized by more than one sperm. Diploid organisms normally contain two copies of each chromosome, one from each parent. The cell resulting from polyspermy, on the other hand, contains three or more copies of each chromosome -- one from the egg and one each from multiple sperm. Usually, the result is an inviable zygote. This may occur because sperm are too efficient at reaching and fertilizing eggs due to the selective pressures of sperm competition. Such a situation is often deleterious to the female: in other words, the male-male competition among sperm spills over to create sexual conflict. Fast block of polyspermy The eggs of sexually reproducing organisms are adapted to avoid this situation.[2] The defenses are particularly well characterized in the sea urchin, which responds to the acceptance of one sperm by inhibiting the successful penetration of the egg by subsequent sperm. Similar defenses exist in other eukaryotes.
Notes made by kozi1989 email:kozi1989@yahoo.gr
118
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The prevention of polyspermy in sea urchins depends on a change in the electrical charge across the surface of the egg, which is caused by the fusion of the first sperm with the egg. [3] Unfertilized sea urchin eggs have a negative charge inside, but the charge becomes positive upon fertilization. When sea urchin sperm encounter an egg with a positive charge, sperm-egg fusion is blocked. Thus, after the first sperm contacts the egg and causes the change, subsequent sperm are prevented from fusing. This "electrical polyspermy block" is thought to result because a positively charged molecule in the sperm surface membrane is repelled by the positive charge at the egg surface.[4] Electrical polyspermy blocks operate in many animal species, including frogs, clams, and marine worms, but not in the several mammals that have been studied (hamster, rabbit, mouse). [5] In species without an electrical block, polyspermy is usually prevented by secretion of materials that establish a mechanical barrier to polyspermy. Animals such as a sea urchins have a two-step polyspermy prevention strategy, with the fast, but transient, electrical block superseded after the first minute or so by a more slowly developing permanent mechanical block. It is thought that electrical blocks evolved in those species where a very fast block to polyspermy is needed, due to the presence of many sperm arriving simultaneously at the egg surface, as occurs in animals such as sea urchins. In sea urchins, fertilization occurs externally in the ocean, such that hundreds of sperm can encounter the egg within several seconds. Slow block of polyspermy In mammals, in which fertilization occurs internally, fewer sperm reach the fertilization site in the oviduct. This may be the result of the female genital tract being adapted to minimize the number of sperm reaching the egg. [6] Nevertheless, polyspermy preventing mechanisms are essential in mammals; a secretion reaction, the "cortical reaction" modifies the extracellular coat of the egg (the zona pellucida), and additional mechanisms that are not well understood modify the egg's plasma membrane. A zygote (from Greek zygtos "joined" or "yoked", from zygoun "to join" or "to yoke"),[1] or zygocyte, is the initial cell formed when two gamete cells are joined by means of sexual reproduction. It is the earliest developmental stage of the embryo. A zygote is always synthesized from the union of two gametes, and constitutes the first stage in a unique organism's development. Zygotes are usually produced by a fertilization event between two haploid cellsan ovum from a female and a sperm cell from a malewhich combine to form the single diploid cell. Such zygotes contain DNA derived from both the mother and the father, and this provides all the genetic information necessary to form a new individual. The term zygote is also used more loosely to refer to the group of cells formed by the first few cell divisions, although this is properly referred to as a morula. In mammalian reproduction, after fertilization has taken place the zygote travels down the fallopian tube, while dividing to form more cells[3] without the zygote actually increasing in size. This cell division is mitotic, and is known as cleavage.[4] All mammals go
Notes made by kozi1989 email:kozi1989@yahoo.gr
119
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
through the zygote stage of life. Mammalian zygotes eventually develop into a blastocyst, after which they are more generally termed an embryo, and then a fetus
46. Reproductive biology. Fertilization. Internal fertilization in mammals.
Differences between external and internal fertilization. In mammals, internal fertilization is done through copulation, which involves the insertion of the penis into the vagina. Some other higher vertebrate animals (most reptiles, most birds, and some fish) reproduce internally, but their fertilization is cloacal. The union of spermatozoa of the parent organism. At some point, the growing egg or offspring must be expelled. There are three general ways of doing this: Oviparous organisms, including most insects and reptiles, monotremes, and all birds lay eggs that continue to develop after being laid, and hatch later. Viviparous organisms, including almost all mammals (such as whales, kangaroos and humans) bear their young live. The developing young spend proportionately more time within the female's reproductive tract. The young are later released to survive on their own, with varying amounts of help from the parent (s) on the species. Ovoviviparous organisms, like the garter snake, and the Madagascar hissing cockroach, have eggs (with shells) that hatch as they are laid, making it look like "live birth". Most species of land animals reproduce by internal fertilization. For example: All reptiles, such as the snake and turtle reproduce by internal fertilizations. Males and females usually have an opening called the cloaca through which semen, urin and feces can be released. During mating, the male and female join their cloacas. The male releases semen into the female's cloaca. The spermatozoe then travel up a canal to reach the ova. External fertilization 1) During external fertilization, many gametes are released into the water by each sex at the same time and place. 2) This type of fertilization requires water because animal sperm must swim to the eggs. Water also protects the gametes from drying out. 3) Species which have external fertilization are either aquatic or return to water for reproduction. ex frogs Internal fertilization is practiced by species that lay shelled eggs or have a period of internal embryonic development. 1) This type of fertilization enables animals to reproduce in a terrestrial environment because it enables sperm to swim to the egg and it prevents gametes from drying out. 2) The male often has a copulatory organ (a penis) for transferring sperm. Males of some aquatic animals (ex: sharks, skates, rays) have specialized pelvic fins which allow the sperm to be passed to the female.
Notes made by kozi1989 email:kozi1989@yahoo.gr
120
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
EXTERNAL Lots of eggs produced (between thousands and millions). Many eggs remain unfertilised. Parent's job is done after fertilisation. Smaller percentage of offspring survive to maturity. Offspring very immature when they hatch.
Year:
INTERNAL Far fewer eggs produced (between one and several hundred). Most eggs are fertilised. To some extent the parent (female) is still responsible. Greater proportion of offspring survive to maturity. Offspring more mature when they hatch.
47. Reproductive biology. In vitro fertilization. ICSI method. Atypical forms of
reproduction. In vitro fertilisation (IVF) is a process by which egg cells are fertilised by sperm outside the body: in vitro. IVF is a major treatment in infertility when other methods of assisted reproductive technology have failed. The process involves hormonally controlling the ovulatory process, removing ova (eggs) from the woman's ovaries and letting sperm fertilise them in a fluid medium. The fertilised egg (zygote) is then transferred to the patient's uterus with the intent to establish a successful pregnancy. The first successful birth of a "test tube baby", Louise Brown, occurred in 1978. Method Theoretically, in vitro fertilisation could be performed by collecting contents from a woman's fallopian tubes or uterus after natural ovulation, mix it with semen from a man and reinsert into the uterus. However, without additional techniques, the chances of pregnancy would be extremely small. Such additional techniques that are routinely used in IVF include ovarian hyperstimulation to retrieve multiple eggs, ultrasound-guided transvaginal oocyte retrieval directly from the ovaries, egg and sperm preparation, as well as culture and selection of resultant embryos before embryo transfer back into the uterus. Ovarian hyperstimulation There are two main protocols for stimulating the ovaries for IVF treatment. The long protocol involves downregulation (suppression or exhaustion) of the pituitary ovarian axis by the prolonged use of a GnRH agonist. Subsequent ovarian hyperstimulation, typically using follicle stimulating hormone (FSH), starts once the process of downregulation is complete, generally after 10 to 14 days. The short protocol skips the downregulation part, and consist of a regimen of fertility medications to stimulate the development of multiple follicles of the ovaries. In most patients, injectable gonadotropins (usually FSH analogues) are used under close monitoring. Such monitoring frequently checks the estradiol level and, by means of gynecologic
Notes made by kozi1989 email:kozi1989@yahoo.gr
121
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
ultrasonography, follicular growth. Typically approximately 10 days of injections will be necessary. Spontaneous ovulation during the cycle is typically prevented by the use of GnRH antagonists that are used just during the last days of stimulation to block the natural surge of luteinising hormone (LH) and allow the physician to start the ovulation process by using medication, usually injectable human chorionic gonadotropins. Ovarian stimulation carries the risk of excessive stimulation. This complication is lifethreatening and ovarian stimulation using gonadotrophins must only be carried out under strict medical supervision Egg retrieval When the ovarian follicles have reached a certain degree of development, final maturation is induced, generally by an injection of human chorionic gonadotropin (hCG). Commonly, this is known as the "trigger shot."[2] hCG acts as an analogue of luteinising hormone, and ovulation would occur between 38 and 40 hours after a single HCG injection,[3] but the egg retrieval is performed at a time usually between 34 and 36 hours after hCG injection, that is, just prior to when the follicles would rupture. This avails for scheduling the egg retrieval procedure at a time where the eggs are fully mature. HCG injection confers a risk of ovarian hyperstimulation syndrome. Using a GnRH agonist instead of hCG eliminates the risk of ovarian hyperstimulation syndrome, but with a delivery rate of approximately 6% less than with hCG. The eggs are retrieved from the patient using a transvaginal technique called transvaginal oocyte retrieval, involving an ultrasound-guided needle piercing the vaginal wall to reach the ovaries. Through this needle follicles can be aspirated, and the follicular fluid is handed to the IVF laboratory to identify ova. It is common to remove between ten and thirty eggs. The retrieval procedure takes about 20 minutes and is usually done under conscious sedation or general anaesthesia. Egg and sperm preparation In the laboratory, the identified eggs are stripped of surrounding cells and prepared for fertilisation. An oocyte selection may be performed prior to fertilisation to select eggs with optimial chances of successful pregnancy. In the meantime, semen is prepared for fertilisation by removing inactive cells and seminal fluid in a process called sperm washing. If semen is being provided by a sperm donor, it will usually have been prepared for treatment before being frozen and quarantined, and it will be thawed ready for use. Fertilisation The sperm and the egg are incubated together at a ratio of about 75,000:1 in the culture media for about 18 hours. In most cases, the egg will be fertilised by that time and the fertilised egg will show two pronuclei. In certain situations, such as low sperm count or motility, a single sperm may be injected directly into the egg using intracytoplasmic sperm injection (ICSI). The fertilised egg is passed to a special growth medium and left for about 48
Notes made by kozi1989 email:kozi1989@yahoo.gr
122
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
hours until the egg consists of six to eight cells. In gamete intrafallopian transfer, eggs are removed from the woman and placed in one of the fallopian tubes, along with the man's sperm. This allows fertilisation to take place inside the woman's body. Therefore, this variation is actually an in vivo fertilisation, not an in vitro fertilisation. Embryo culture Typically, embryos are cultured until having reached the 6 8 cell stage three days after retrieval. In many Canadian, American and Australian programmes[citation needed], however, embryos are placed into an extended culture system with a transfer done at the blastocyst stage at around five days after retrieval, especially if many good-quality embryos are still available on day 3. Blastocyst stage transfers have been shown to result in higher pregnancy rates.[5] In Europe, transfers after 2 days are common. Culture of embryos can either be performed in an artificial culture medium or in an autologous endometrial coculture (on top of a layer of cells from the woman's own uterine lining). With artificial culture medium, there can either be the same culture medium throughout the period, or a sequential system can be used, in which the embryo is sequentially placed in different media. For example, when culturing to the blastocyst stage, one medium may be used for culture to day 3, and a second medium is used for culture thereafter.[6] Single or sequential medium are equally effective for the culture of human embryos to the blastocyst stage.[7] Artificial embryo culture media basically contain glucose, pyruvate, and energy-providing components, but addition of amino acids, nucleotides, vitamins, and cholesterol improve the performance of embryonic growth and development.[8] Methods to permit dynamic embryo culture with fluid flow and embryo movement are also available.[9] Embryo selection Laboratories have developed grading methods to judge oocyte and embryo quality. In order to optimise pregnancy rates, there is significant evidence that a morphological scoring system is the best strategy for the selection of embryos.[10] However, presence of soluble HLA-G might be considered as a second parameter if a choice has to be made between embryos of morphologically equal quality.[10] Also, two-pronuclear zygotes (2PN) transitioning through 1PN or 3PN states tend to develop into poorer-quality embryos than those who constantly remain 2PN.[11] In addition to tests that optimise pregnancy chances, Preimplantation genetic diagnosis (PGD) or screening may be performed prior to transfer in order to avoid inheritable diseases.[12] Methods are emerging in making comprehensive analyses of transcriptomes of embryos in order to assess embryo quality.[13] Embryo transfer Embryos are graded by the embryologist based on the number of cells, evenness of growth and degree of fragmentation. The number to be transferred depends on the
Notes made by kozi1989 email:kozi1989@yahoo.gr
123
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
number available, the age of the woman and other health and diagnostic factors. In countries such as Canada, the UK, Australia and New Zealand, a maximum of two embryos are transferred except in unusual circumstances. In the UK and according to HFEA regulations, a woman over 40 may have up to three embryos transferred, whereas in the USA, younger women may have many embryos transferred based on individual fertility diagnosis. Most clinics and country regulatory bodies seek to minimise the risk of pregnancies carrying multiples. As it is not uncommon for more implantations to take than desired, the next step faced by the expectant mother is that of selective abortion. The embryos judged to be the "best" are transferred to the patient's uterus through a thin, plastic catheter, which goes through her vagina and cervix. Several embryos may be passed into the uterus to improve chances of implantation and pregnancy. Intracytoplasmic sperm injection (ICSI, pronounced "eeksee"[1] or "icksy"[2]) is an in vitro fertilization procedure in which a single sperm is injected directly into an egg. This procedure is most commonly used to overcome male infertility problems, although it may also be used where eggs cannot easily be penetrated by sperm, and occasionally in addition to sperm donation. It can be used in teratozoospermia, because once the egg is fertilized, abnormal sperm morphology does not appear to influence blastocyst development or blastocyst morphology.[3] Even with severe teratozoospermia, microscopy can still detect the few sperm cells that have a "normal" morphology, allowing for optimal success rate.[3] Procedure ICSI is generally performed in addition an in vitro fertilisation procedure to extract often several oocytes from a woman. The procedure is done under a microscope using multiple micromanipulation devices (micromanipulator, microinjectors and micropipettes). A holding pipette (on the left of picture) stabilizes the mature oocyte with gentle suction applied by a microinjector. From the opposite side a thin, hollow glass micropipette is used to collect a single sperm, having immobilised it by cutting its tail with the point of the micropipette. The micropipette is pierced through the oolemma and into the inner part of the oocyte (cytoplasm). The sperm is then released into the oocyte. The pictured oocyte has an extruded polar body at about 12 o'clock indicating its maturity. After the procedure, the oocyte will be placed into cell culture and checked on the following day for signs of fertilization. In contrast, in natural fertilization sperm compete and when the first sperm penetrates the oolemma, the oolemma hardens to block the entry of any other sperm. Concern has been raised that in ICSI this sperm selection process is bypassed and the sperm is selected by the embryologist without any specific testing. However, in mid 2006 the FDA cleared a device that allows embryologists to select mature sperm for ICSI based on sperm binding to hyaluronan, the main constituent of the gel layer (cumulus oophorus) surrounding the oocyte. The device provides microscopic droplets of hyaluronan hydrogel attached to the
Notes made by kozi1989 email:kozi1989@yahoo.gr
124
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
culture dish. The embryologist places the prepared sperm on the microdot, selects and captures sperm that bind to the dot. Basic research on the maturation of sperm shows that hyaluronan-binding sperm are more mature and show fewer DNA strand breaks and significantly lower levels of aneuploidy than the sperm population from which they were selected. A brand name for one such sperm selection device is PICSI. Production of offsprings by a single parent without the formation of gametes is called asexual reproduction. It is also called as somatogenic reproduction. Asexual reproduction is a mode of reproduction by which offspring arise from a single parent, and inherit the genes of that parent only, it is reproduction which does not involve meiosis, ploidy reduction, or fertilization. A more stringent definition is agamogenesis which is reproduction without the fusion of gametes. Asexual reproduction is the primary form of reproduction for single-celled organisms such as the archaea, bacteria, and protists. Many plants and fungi reproduce asexually as well. While all prokaryotes reproduce asexually (without the formation and fusion of gametes), mechanisms for lateral gene transfer such as conjugation, transformation and transduction are sometimes likened to sexual reproduction.[1] A complete lack of sexual reproduction is relatively rare among multicellular organisms, particularly animals. It is not entirely understood why the ability to reproduce sexually is so common among them. Current hypotheses [2] suggest that asexual reproduction may have short term benefits when rapid population growth is important or in stable environments, while sexual reproduction offers a net advantage by allowing more rapid generation of genetic diversity, allowing adaptation to changing environments. Developmental constraints[3] may underlie why few animals have relinquished sexual reproduction completely in their life-cycles. Types Binary fission In binary fission the parent organism is replaced by two daughter organisms, because it literally divides in two. Many single-celled organisms, both prokaryotes (the archaea and the bacteria), and eukaryotes (such as protists and unicellular fungi), reproduce asexually through binary fission; most of these are also capable of sexual reproduction. Some single-celled organisms rely on one or more host organisms in order to reproduce. Budding Some cells split via budding (for example baker's yeast), resulting in a 'mother' and 'daughter' cell. The offspring organism is smaller than the parent. Budding is also known on a multicellular level; an animal example is the hydra, which reproduces by budding. The buds grow into fully matured individuals which eventually break away from the parent organism. Vegetative reproduction
Notes made by kozi1989 email:kozi1989@yahoo.gr
125
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Vegetative reproduction is a type of asexual reproduction found in plants where new individuals are formed without the production of seeds or spores by meiosis or syngamy.[4] Examples of vegetative reproduction include the formation of miniaturized plants called plantlets on specialized leaves (for example in kalanchoe) and some produce new plants out of rhizomes or stolon (for example in strawberry). Other plants reproduce by forming bulbs or tubers (for example tulip bulbs and dahlia tubers). Some plants produce adventitious shoots and suckers that form along their lateral roots. Plants that reproduce vegetatively may form a clonal colony, where all the individuals are clones, and the clones may cover a large area.[5] Spore formation Many multicellular organisms form spores during their biological life cycle in a process called sporogenesis. Exceptions are animals and some protists, who undergo gametic meiosis immediately followed by fertilization. Plants and many algae on the other hand undergo sporic meiosis where meiosis leads to the formation of haploid spores rather than gametes. These spores grow into multicellular individuals (called gametophytes in the case of plants) without a fertilization event. These haploid individuals give rise to gametes through mitosis. Meiosis and gamete formation therefore occur in separate generations or "phases" of the life cycle, referred to as alternation of generations. Since sexual reproduction is often more narrowly defined as the fusion of gametes (fertilization), spore formation in plant sporophytes and algae might be considered a form of asexual reproduction (agamogenesis) despite being the result of meiosis and undergoing a reduction in ploidy. However, both events (spore formation and fertilization) are necessary to complete sexual reproduction in the plant life cycle. Fungi and some algae can also utilize true asexual spore formation, which involves mitosis giving rise to reproductive cells called mitospores that develop into a new organism after dispersal. This method of reproduction is found for example in conidial fungi and the red alga Polysiphonia, and involves sporogenesis without meiosis. Thus the chromosome number of the spore cell is the same as that of the parent producing the spores. However, mitotic sporogenesis is an exception and most spores, such as those of plants, most Basidiomycota, and many algae, are produced by meiosis. Fragmentation Fragmentation is a form of asexual reproduction where a new organism grows from a fragment of the parent. Each fragment develops into a mature, fully grown individual. Fragmentation is seen in many organisms such as animals (some annelid worms and sea stars), fungi, and plants. Some plants have specialized structures for reproduction via fragmentation, such as gemmae in liverworts. Most lichens, which are a symbiotic union of a fungus and photosynthetic algae or bacteria, reproduce through fragmentation to ensure that new individuals contain both symbionts. These fragments can take the form of soredia, dust-like particles consisting of fungal hyphae wrapped around photobiont cells.
Notes made by kozi1989 email:kozi1989@yahoo.gr
126
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Agamogenesis Agamogenesis is any form of reproduction that does not involve a male gamete. Examples are parthenogenesis and apomixis. Parthenogenesis Parthenogenesis is a form of agamogenesis in which an unfertilized egg develops into a new individual. Parthenogenesis occurs naturally in many plants, invertebrates (e.g. water fleas, aphids, stick insects, some ants, bees and parasitic wasps), and vertebrates (e.g. some reptiles, amphibians, fish, very rarely birds). In plants, apomixis may or may not involve parthenogenesis. Apomixis and nucellar embryony Apomixis in plants is the formation of a new sporophyte without fertilization. It is important in ferns and in flowering plants, but is very rare in other seed plants. In flowering plants, the term "apomixis" is now most often used for agamospermy, the formation of seeds without fertilization, but was once used to include vegetative reproduction. An example of an apomictic plant would be the triploid European dandelion. Apomixis mainly occurs in two forms: In gametophytic apomixis, the embryo arises from an unfertilized egg within a diploid embryo sac that was formed without completing meiosis. In nucellar embryony, the embryo is formed from the diploid nucellus tissue surrounding the embryo sac. Nucellar embryony occurs in some citrus seeds. Male apomixis can occur in rare cases, such as the Saharan Cypress where the genetic material of the embryo are derived entirely from pollen. The term "apomixis" is also used for asexual reproduction in some animals, notably water-fleas, Daphnia.
48. Individual development. Embryonic period. Body plan formation, grey crescent,
cleavage. Regulative and mosaic types of development. Week 1 Fertilization of ovum to form zygote The zygote undergoes mitotic cellular divisions, but does not increase in size. This mitosis is also known as cleavage. A hollow cavity forms marking the blastocyst stage The blastocyst hatches from its protein shell (zona pellucida) and performs implantation onto the endometrial lining of the mother's uterus Week 2 Trophoblast cells surrounding the embryonic cells proliferate and invade deeper into the uterine lining. They will eventually form the placenta and embryonic membranes. The blastocyst is fully implanted day 7-12 of fertilization Formation of the yolk sac
Notes made by kozi1989 email:kozi1989@yahoo.gr
127
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Week 3 Week 4
The embryonic cells flatten into a disk, two-cells thick Primitive streak develops and Primary stem villi appear
A notochord forms in the center of the embryonic disk and gastrulation occurs A neural groove (future spinal cord) forms over the notochord with a brain bulge at one end. Neuromeres appear Somites, the divisions of the future vertebra, form Primitive heart tube is forming. Vasculature begins to develop in embryonic disc
The heart bulges, further develops, and begins to beat in a regular rhythm. Septum primum appear Branchial arches, grooves which will form structures of the face and neck, form. The neural tube closes The ears begin to form as otic pits Arm buds and a tail are visible Pulmonary primordium, the first traits of the lung appear Hepatic plate, the first traits of the liver appear Buccopharyngeal membrane ruptures. This is the future mouth Cystic diverticulum, which will become the gallbladder, and dorsal pancreatic bud, which will become the pancreas appear Urorectal septum begins to form. Thus, the rectal and urinary passageways become separated Anterior and posterior horns differentiate in the spinal cord Spleen appears Ureteric buds appear Week 5 Lens pits and optic cups form the start of the developing eye. Nasal pits form. The brain divides into 5 vesicles, including the early telencephalon. Leg buds form and hands form as flat paddles on the arms. Rudimentary blood moves through primitive vessels connecting to the yolk sac and chorionic membranes.
Week 6 Lungs begin to form. The brain continues to develop. Arms and legs have lengthened with foot and hand areas distinguishable.
Notes made by kozi1989 email:kozi1989@yahoo.gr
128
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The hands and feet have digits, but may still be webbed. The gonadal ridge begins to be perceptible. The lymphatic system begins to develop. Week 7 Nipples and hair follicles begin to form. Location of the elbows and toes are visible. Spontaneous limb movements may be detected by ultrasound. All essential organs have at least begun formation.
Grey crescent A cortical region of the newly fertilised egg of frogs and some salamanders that forms just after fertilization on the side opposite sperm penetration. In embryology, cleavage is the division of cells in the early embryo. The zygotes of many species undergo rapid cell cycles with no significant growth, producing a cluster of cells the same size as the original zygote. The different cells derived from cleavage are called blastomeres and form a compact mass called the morula. Cleavage ends with the formation of the blastula. Depending mostly on the amount of yolk in the egg, the cleavage can be holoblastic (total or entire cleavage) or meroblastic (partial cleavage). The pole of the egg with the highest concentration of yolk is referred to as the vegetal pole while the opposite is referred to as the animal pole. Cleavage differs from other forms of cell division in that it increases the number of cells without increasing the mass. This means that with each successive subdivision, the ratio of nuclear to cytoplasmic material increases. Mechanism The rapid cell cycles are facilitated by maintaining high levels of proteins that control cell cycle progression such as the cyclins and their associated cyclin-dependent kinases (cdk). The complex CyclinB/cdc2 a.k.a. MPF (maturation promoting factor) promotes entry into mitosis. The processes of karyokinesis (mitosis) and cytokinesis work together to result in cleavage. The mitotic apparatus is made up of a central spindle and polar asters made up of polymers of tubulin protein called microtubules. The asters are nucleated by centrosomes and the centrosomes are organized by centrioles brought into the egg by the sperm as basal bodies. Cytokinesis is mediated by the contractile ring made up of polymers of actin protein called microfilaments. Karyokinesis and cytokinesis are independent but spatially and temporally coordinated processes. While mitosis can occur in the absence of cytokinesis, cytokinesis requires the mitotic apparatus. The end of cleavage coincides with the beginning of zygotic transcription. This point
Notes made by kozi1989 email:kozi1989@yahoo.gr
129
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
is referred to as the midblastula transition and appears to be controlled by the nuclear:cytoplasmic ratio (about 1/6). Regulative development A type of embryonic development in which the fertilized ovum undergoes indeterminate cleavage, producing blastomeres that have similar developmental potencies and are each capable of giving rise to a single embryo. Determination of the particular organs and parts of the embryo occurs during later stages of development and is influenced by inductors and intercellular interaction. Damage or destruction of various cells during the early stages of development results in readjustments and substitutions so that a normal organism is formed. Mosaic development A kind of embryonic development occurring in the blastocyst. The fertilized ovum undergoes determinate cleavage, developing according to a precise, unalterable plan in which each blastomere has a characteristic position and limited developmental potency and is a precursor of a specific part of the embryo. Damage to or destruction of these cells results in a defective organism.
49. Individual development. Formation of the blastula. Gastrulation. Cellular and
molecular mechanisms of gastrulation. The blastula (from Greek (blastos), meaning "sprout") is a solid sphere of cells formed during an early stage of embryonic development in animals. The blastula is created when the zygote undergoes the cell division process known as cleavage. [1] The blastula is preceded by the morula and is followed by the gastrula in the developmental sequence.[citation needed A common feature of a vertebrate blastula is that it consists of a layer of blastomeres, known as the blastoderm, which surrounds an interior central cavity known as the blastocoel.[2] [3] In mammals, blastulation leads to the formation of the blastocyst, which must not be confused with the blastula. The blastocyst contains an embryoblast, which is homologous to the blastula. However, it also includes the trophoblast, which goes on to form the extraembryonic tissues.[citation needed] After the cleavage has produced over 100 cells, the embryo is called a blastula .[3] The blastula is usually a spherical layer of cells (the blastoderm) surrounding a fluid-filled or yolk-filled cavity (the blastocoel). Mammals at this stage form a structure called the blastocyst,[4] characterized by an inner cell mass that is distinct from the surrounding blastula. The blastocyst must not be confused with the blastula; even though they are similar in structure, their cells have different fates. Before gastrulation, the cells of the trophoblast become differentiated into two strata: the outer stratum forms a syncytium (i.e. a layer of protoplasm studded with nuclei,
Notes made by kozi1989 email:kozi1989@yahoo.gr
130
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
but showing no evidence of subdivision into cells), termed the syncytiotrophoblast; while the inner layer, the cytotrophoblast or "Layer of Langhans," consists of well-defined cells. As already stated, the cells of the trophoblast do not contribute to the formation of the embryo proper; they form the ectoderm of the chorion and play an important part in the development of the placenta. On the deep surface of the inner cell mass, a layer of flattened cells, called the entoderm, is differentiated and quickly assumes the form of a small sac, called the yolk sac. Spaces appear between the remaining cells of the mass and, by the enlargement and coalescence of these spaces, a cavity called the amniotic cavity is gradually developed. The floor of this cavity is formed by the embryonic disk, which is composed of a layer of prismatic cells, the embryonic ectoderm, derived from the inner cell mass and lying in apposition with the entoderm. Gastrulation is a phase early in the embryonic development of most animals, during which the single-layered blastula is reorganized into a trilaminar ("three-layered") structure known as the gastrula. These three germ layers are known as the ectoderm, mesoderm, and endoderm. Gastrulation takes place after cleavage and the formation of the blastula and primitive streak. Gastrulation is followed by organogenesis, when individual organs develop within the newly formed germ layers.[3] Each layer gives rise to specific tissues and organs in the developing embryo. The ectoderm gives rise to epidermis, and to the neural crest and other tissues that will later form the nervous system. The mesoderm is found between the ectoderm and the endoderm and gives rise to somites, which form muscle; the cartilage of the ribs and vertebrae; the dermis, the notochord, blood and blood vessels, bone, and connective tissue. The endoderm gives rise to the epithelium of the digestive system and respiratory system, and organs associated with the digestive system, such as the liver and pancreas.[4] Following gastrulation, cells in the body are either organized into sheets of connected cells (as in epithelia), or as a mesh of isolated cells, such as mesenchyme.[2][5] The molecular mechanism and timing of gastrulation is different in different organisms. However some common features of gastrulation across triploblastic organisms include: 1) A change in the topological structure of the embryo, from a simply connected surface (sphere-like), to a non-simply connected surface (torus-like); 2) the differentiation of cells into one of three types (endodermal, mesodermal, and ectodermal); and 3) the digestive function of a large number of endodermal cells.
50. Individual development. Embryonic indection. Embryonic stem cells. Elements
of organogenesis. Embryonic induction
Notes made by kozi1989 email:kozi1989@yahoo.gr
131
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The organization of the embryo as a whole appears to be determined to a large extent during gastrulation, by which process different regions of the blastoderm are displaced and brought into new spatial relationships to each other. Groups of cells that were distant from each other in the blastula come into close contact, which increases possibilities for interaction between materials of different origin. In the development of vertebrates in particular, the sliding of cells (presumptive mesoderm) into the interior and their placement on the dorsal side of the archenteron (in the archenteric roof), in immediate contact with the overlying ectoderm, is of major importance in development and subsequent differentiation. Experiments have shown that, at the start of gastrulation, ectoderm is incapable of progressive development of any kind; that only after invagination, with chordamesoderm lying directly underneath it, does ectoderm acquire the ability for progressive development. The dorsal mesoderm, which later differentiates into notochord, prechordal mesoderm, and somites, causes the overlying ectoderm to differentiate as neural plate. Lateral mesoderm causes overlying ectoderm to differentiate as skin. The influence exercised by parts of the embryo, which causes groups of cells to proceed along a particular path of development, is called embryonic induction. Though induction requires that the interacting parts come into close proximity, actual contact is not necessary. The inducing influencewhatever it might beis a diffusible substance emitted by the activating cells (the inductor). The inducing substance of the mesoderm is a large molecule, probably a protein or a nucleoprotein, which presumably penetrates reacting cells, though direct and unequivocal proof of such penetration is still unavailable. Inducing substances are active on vertebrates belonging to many different classes; e.g., inductions of primary organs have been obtained by transplanting mammalian tissues into frog embryos or by transplanting tissues of a chick embryo into the embryo of a rabbit. Induction is responsible not only for the subdivision of ectoderm into neural plate and epidermis but also for the development of a large number organ rudiments in vertebrates. The notochord is a source of induction for the development of the adjoining somites and nephrotomes; the latter appear jointly to induce development of limb rudiments from the lateral plate mesoderm. Embryonic stem cells (ES cells) are pluripotent stem cells derived from the inner cell mass of the blastocyst, an early-stage embryo.[1] Human embryos reach the blastocyst stage 45 days post fertilization, at which time they consist of 50 150 cells. Isolating the embryoblast or inner cell mass (ICM) results in destruction of the fertilized human embryo, which raises ethical issues. Embryonic stem cells are distinguished by two distinctive properties: 1) their pluripotency, and 2) their ability to replicate indefinitely ES cells are pluripotent, that is, they are able to differentiate into all derivatives of the three primary germ layers: ectoderm, endoderm, and mesoderm. These include each of
Notes made by kozi1989 email:kozi1989@yahoo.gr
132
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
the more than 220 cell types in the adult body. Pluripotency distinguishes embryonic stem cells from adult stem cells found in adults; while embryonic stem cells can generate all cell types in the body, adult stem cells are multipotent and can only produce a limited number of cell types. Additionally, under defined conditions, embryonic stem cells are capable of propagating themselves indefinitely. This allows embryonic stem cells to be employed as useful tools for both research and regenerative medicine, because they can produce limitless numbers of themselves for continued research or clinical use. Because of their plasticity and potentially unlimited capacity for self-renewal, ES cell therapies have been proposed for regenerative medicine and tissue replacement after injury or disease. Diseases that could potentially be treated by pluripotent stem cells include a number of blood and immune-system related genetic diseases, cancers, and disorders; juvenile diabetes; Parkinson's; blindness and spinal cord injuries. Besides the ethical concerns of stem cell therapy (see stem cell controversy), there is a technical problem of graft-versus-host disease associated with allogeneic stem cell transplantation. However, these problems associated with histocompatibility may be solved using autologous donor adult stem cells, therapeutic cloning, stem cell banks or more recently by reprogramming of somatic cells with defined factors (e.g. induced pluripotent stem cells). Other potential uses of embryonic stem cells include investigation of early human development, study of genetic disease and as in vitro systems for toxicology testing. At some point after the different germ layers are defined, organogenesis begins. The first stage in vertebrates is called neurulation, where the neural plate folds forming the neural tube. Other common organs or structures which arise at this time include the heart and somites (also above), but from now on embryogenesis follows no common pattern among the different taxa of the animal kingdom. In most animals organogenesis along with morphogenesis will result in a larva. The hatching of the larva, which must then undergo metamorphosis, marks the end of embryonic development.
51. Individual development
Embryonic development of mammals and human. Differentiation, differences between cells, cell memory. In developmental biology, cellular differentiation is the process by which a less specialized cell becomes a more specialized cell type. Differentiation occurs numerous times during the development of a multicellular organism as the organism changes from a simple zygote to a complex system of tissues and cell types. Differentiation is a common process in adults as well: adult stem cells divide and create fully-differentiated daughter cells during tissue repair and during normal cell turnover. Differentiation dramatically changes a cell's size, shape, membrane potential, metabolic activity, and responsiveness to signals. These changes are largely due to highly-controlled modifications in gene expression. With a
Notes made by kozi1989 email:kozi1989@yahoo.gr
133
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
few exceptions, cellular differentiation almost never involves a change in the DNA sequence itself. Thus, different cells can have very different physical characteristics despite having the same genome. A cell that is able to differentiate into all cell types of the adult organism is known as pluripotent. Such cells are called embryonic stem cells in animals and meristematic cells in higher plants. A cell that is able to differentiate into all cell types, including the placental tissue, is known as totipotent. In mammals, only the zygote and subsequent blastomeres are totipotent, while in plants many differentiated cells can become totipotent with simple laboratory techniques. In cytopathology, the level of cellular differentiation is used as a measure of cancer progression. "Grade" is a marker of how differentiated a cell in a tumor is. Three basic categories of cells make up the mammalian body: germ cells, somatic cells, and stem cells. Each of the approximately 100 trillion (1014) cells in an adult human has its own copy or copies of the genome except certain cell types, such as red blood cells, that lack nuclei in their fully differentiated state. Most cells are diploid; they have two copies of each chromosome. Such cells, called somatic cells, make up most of the human body, such as skin and muscle cells. Cells differentiate to specialize for different functions. Germ line cells are any line of cells that give rise to gametes eggs and spermand thus are continuous through the generations. Stem cells, on the other hand, have the ability to divide for indefinite periods and to give rise to specialized cells. They are best described in the context of normal human development. Development begins when a sperm fertilizes an egg and creates a single cell that has the potential to form an entire organism. In the first hours after fertilization, this cell divides into identical cells. In humans, approximately four days after fertilization and after several cycles of cell division, these cells begin to specialize, forming a hollow sphere of cells, called a blastocyst. The blastocyst has an outer layer of cells, and inside this hollow sphere, there is a cluster of cells called the inner cell mass. The cells of the inner cell mass go on to form virtually all of the tissues of the human body. Although the cells of the inner cell mass can form virtually every type of cell found in the human body, they cannot form an organism. These cells are referred to as pluripotent. Pluripotent stem cells undergo further specialization into multipotent progenitor cells that then give rise to functional cells. Examples of stem and progenitor cells includ 1) Hematopoietic stem cells (adult stem cells) from the bone marrow that give rise to red blood cells, white blood cells, and platelets 2) Mesenchymal stem cells (adult stem cells) from the bone marrow that give rise to stromal cells, fat cells, and types of bone cells 3) Epithelial stem cells (progenitor cells) that give rise to the various types of skin cells 4) Muscle satellite cells (progenitor cells) that contribute to differentiated muscle tissue
Notes made by kozi1989 email:kozi1989@yahoo.gr
134
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Since each cell, regardless of cell type, possesses the same genome, determination of cell type must occur at the level of gene expression. While the regulation of gene expression can occur through cis- and trans-regulatory elements including a genes promoter and enhancers, the problem arises to how this expression pattern is maintained over numerous generations of cell division. As it turns out, epigenetic processes play a crucial role in regulating the decision to adopt a stem, progenitor, or mature cell fate. This section will focus primarily on mammalian stem cells. Body memory is a hypothesis that the body itself is capable of storing memories, as opposed to only the brain. This is used to explain having memories for events where the brain was not in a position to store memories and is sometimes a catalyst for repressed memories recovery. These memories are often characterised with phantom pain in a part or parts of the body the body appearing to remember the past trauma. The idea of body memory is a belief frequently associated with the idea of repressed memories, in which memories of incest or sexual abuse can be retained and recovered through physical sensations.[1] This is considered pseudoscientific as there is no known means by which tissues other than the brain are capable of storing memories. Cellular memory Cellular memory is an additional hypothesis that memories can be stored outside the brain. However, unlike body memory, the cellular memory hypothesis states that these memories are stored in all the cells of human bodies, not in the bodies organs.*5] The idea that non-brain tissues can have memories is also believed by some individuals who have received organ transplants, though this is also considered impossible.
52. Individual development
Postembryonic period.Life span, ageing and death. Postembryonic period is the period in the ontogenesis of multicellular animals that follows the period of embryonic development and usually ends with the onset of sexual maturity and, in most animals, the cessation of growth. Postembryonic development begins after the emergence of the embryo from the egg and embryonic membranes, when the organism becomes capable of active feeding and locomotion. Upon transition to the postembryonic state the organism either immediately possesses the principal pubertal morphological characteristics (direct development) or essentially differs from the pubertal form, in which case the larva that hatches out of the egg must undergo a metamorphosis before it reaches its adult state. Growth continues during the period of postembryonic development and further organogenesis and histogenesis occur. The functions of the developing organism become more complex; establishment of the final proportions of the body is especially characteristic. In some animals postembryonic development constitutes the greater part of their life span. Among insects, in a number of cicadas the larva lives 17 years, while the pubertal
Notes made by kozi1989 email:kozi1989@yahoo.gr
135
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
insect lives one summer; the mayfly larva lives to three years, but often in its pubertal form the mayfly lives only one day. The necessity for defense against enemies and active food procurement are ensured during postembryonic development by a number of adaptations that make the animal selfsufficient, sometimes for its entire life span. With indirect, or larval, development, the animal, having larval organs, is self-sufficient only during the postembryonic period. Life expectancy is the expected (in the statistical sense) number of years of life remaining at a given age. It is denoted by ex, which means the average number of subsequent years of life for someone now aged x, according to a particular mortality experience. (In technical literature, this symbol means the average number of complete years of life remaining, excluding fractions of a year. The corresponding statistic including fractions of a year, the normal meaning of life expectancy, has a symbol with a small circle over the e.) The term that is known as life expectancy is most often used in the context of human populations, but is also used in plant or animal ecology;[2] it is calculated by the analysis of life tables (also known as actuarial tables). The term life expectancy may also be used in the context of manufactured objects[3] although the related term shelf life is used for consumer products and the terms "mean time to breakdown" (MTTB) and "mean time before failures" (MTBF) are used in engineering literature. Life span is the time between birth and death. It ranges from a mayfly's day to certain trees' thousands of years. Its limit appears to depend on heredity, but such factors as (in humans) disease, natural disasters, war, diet, and habits such as smoking reduce it. Maximum life span is theoretical. Aging is a series of biological changes that follow a natural progression from birth through maturity to old age and death. For most people, advancing age is characterized by graying or thinning hair, loss of height, wrinkling of the skin, and decreased muscular strength. Still, an active lifestyle, including both exercise and sound nutrition, can contribute greatly to achieving a long and productive life. Genetics (inherited physical characteristics) also appears to play a role in the process of aging and death. People whose parents or grandparents live to old age seem to have a better chance of living long lives themselves. Great strides have been made in increasing the average life expectancy in humans in the United States. This is due largely to the elimination of many diseases of early childhood and young adulthood and to the advanced methods of treating diseases that in the past would have resulted in early death. However, the maximum potential life span (how long an organism can exist) of humans appears to remain the sameabout 85 years regardless of efforts to expand it. Every species has a different normal life span. In most species, death occurs not long after the reproductive phase of life ends. This is obviously not the case for humans. However, women do experience physical changes when they are past their childbearing
Notes made by kozi1989 email:kozi1989@yahoo.gr
136
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
years. Levels of the hormone estrogen begin to fall, which results in the gradual cessation, or stopping, of menstruation. (Hormones are chemicals produced by the body that regulate various bodily functions. Menstruation is the monthly shedding of the lining of the uterus [womb] in nonpregnant females.) After menopause (when menstruation ceases and childbearing is no longer possible), women produce less facial skin oil (which leads to wrinkling) and are at greater risk of developing osteoporosis (brittle bones). Men continue to produce nearly the same levels of facial oils and are thus less prone to early wrinkling. As we age, our cells become less efficient and our bodies become less able to carry out their normal functions. Muscles lose strength, hearing and vision become less acute, reflex times slow down, lung capacity decreases, and the heart's ability to pump blood may be affected. In addition, the immune system weakens, making it less able to fight infection and disease. No single theory on how and why people age is able to account for all aspects of aging, but most take one of two approaches: (1) that humans are genetically programmed to age and die, and (2) that natural wear and tear causes aging and eventual death. Arguments in favor of genetic programming cite hormonal control and/or limited cell division (reproduction) as the mechanisms involved in the aging process. One hormonal approach focuses on the hypothalamus (at the base of the forebrain), which controls the production of growth hormones (which influence growth and development) in the pituitary gland. It is thought thatpossibly due to the action of a hormonal clockthe hypothalamus either slows down normal hormonal function or that it becomes more error-prone with time, leading to physiological aging. Repeated experiments have shown that human cells grown in tissue culture in the laboratory will divide only about 50 times before they die. (Exceptions are cancer cells, which have unlimited growth, and brain and muscle cells, which do not divide after birth). This suggests a genetic clock that limits how many time cells can divide. It is thought that as cell division decreases, as it does in older people, the functioning of the body also begins to slow down, resulting in aging and eventual death. The wear-and-tear theory of aging and death suggests that genes (the molecules that carry instructions for passing on specific traits from one generation to another) are altered by random mutations (changes in form) that accumulate over time, gradually leading to aging and disease. Environmental agents, such as X rays, ultraviolet radiation, and chemical toxins, can contribute to this mutation process. All cells have the ability to repair damaged DNA (deoxyribonucleic acid, the molecular basis of heredity), but when these repair mechanisms fail, mutations can accumulate. This failure is also thought to be a factor in the development of cancer.
53. Population genetics and biology
Population.Phenotype, genotype and gene frequencies.Types of mating.Hardy Weinberg law.
Notes made by kozi1989 email:kozi1989@yahoo.gr
137
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Population genetics is the study of allele frequency distribution and change under the influence of the four main evolutionary processes: natural selection, genetic drift, mutation and gene flow. It also takes into account the factors of recombination, population subdivision and population structure. It attempts to explain such phenomena as adaptation and speciation. A population is all the organisms that both belong to the same species and live in the same geographical area. The area that is used to define the population is such that inter-breeding is possible between any pair within the area and more probable than crossbreeding with individuals from other areas. Normally breeding is substantially more common within the area than across the border. A phenotype is an organism's observable characteristics or traits: such as its morphology, development, biochemical or physiological properties , behavior, and products of behavior (such as a bird's nest). Phenotypes result from the expression of an organism's genes as well as the influence of environmental factors and the interactions between the two. The genotype of an organism is the inherited instructions it carries within its genetic code. The genotype is the genetic makeup of a cell, an organism, or an individual (i.e. the specific allele makeup of the individual) usually with reference to a specific character under consideration. Not all organisms with the same genotype look or act the same way because appearance and behavior are modified by environmental and developmental conditions. Similarly, not all organisms that look alike necessarily have the same genotype. This genotype-phenotype distinction was proposed by Wilhelm Johannsen in 1911 to make clear the difference between an organism's heredity and what that heredity produces Phenotypic variation (due to underlying heritable genetic variation) is a fundamental prerequisite for evolution by natural selection. It is the living organism as a whole that contributes (or not) to the next generation, so natural selection affects the genetic structure of a population indirectly via the contribution of phenotypes. Without phenotypic variation, there would be no evolution by natural selection. The interaction between genotype and phenotype has often been conceptualized by the following relationship: Genotype + environment phenotype A slightly more nuanced version of the relationships is: Genotype + environment + random-variation phenotype Allele frequency is the proportion of all copies of a gene that is made up of a particular gene variant (allele). In other words, it is the number of copies of a particular allele divided by the number of copies of all alleles at the genetic place (locus) in a population. It can be expressed for example as a percentage. In population genetics, allele frequencies are used to depict the amount of genetic diversity at the individual, population, and species level. It is also the relative proportion of all alleles of a gene that are of a designed type.
Notes made by kozi1989 email:kozi1989@yahoo.gr
138
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Given the following: 1) A particular locus on a chromosome and the gene occupying that locus 2) A population of n individuals carrying n loci in each of their somatic cells (e.g. Two loci in the cells of diploid species, which contain two sets of chromosomes) 3) Different alleles of the gene exist 4) One allele exists in a copies Then the allele frequency is the fraction or percentage of all the occurrences of that locus that is occupied by a given allele and the frequency of one of the alleles is a/(n*N). A mating system is a way in which a group is structured in relation to sexual behaviour. The precise meaning depends upon the context. With respect to higher animals, it specifies which males mate with which females, under which circumstances; recognised animal mating systems include monogamy, polygamy (which includes polygyny, polyandry, and polygynandry) and promiscuity. In plants, it refers to the degree and circumstances of outcrossing. The HardyWeinberg principle (also known by a variety of names: HWP, Hardy Weinberg equilibrium, HardyWeinberg Theorem, HWE, or HardyWeinberg law) states that both allele and genotype frequencies in a population remain constant that is, they are in equilibriumfrom generation to generation unless specific disturbing influences are introduced. Those disturbing influences include: 1) Non-random mating, 2) Mutations, selection, 3) Limited population size, 4) "overlapping generations", 5) Random genetic drift, 6) Gene flow 7) Meiotic drive. It is important to understand that outside the lab, one or more of these "disturbing influences" are always in effect. That is, HardyWeinberg equilibrium is impossible in nature. Genetic equilibrium is an ideal state that provides a baseline against which to measure change. Static allele frequencies in a population across generations assume: no mutation (the alleles don't change), no migration or emigration (no exchange of alleles between populations), infinitely large population size, and no selective pressure for or against any genotypes. Genotype frequencies will also be static when mating is random. In the simplest case of a single locus with two alleles: the dominant allele is denoted A and the recessive a and their frequencies are denoted by p and q; freq(A) = p; freq(a) = q; p + q = 1. If the population is in equilibrium, then we will have freq(AA) = p2 for the AA homozygotes in the population, freq(aa) = q2 for the aa homozygotes, and freq(Aa) = 2pq for the heterozygotes. This concept was named after G. H. Hardy and Wilhelm Weinberg.
Notes made by kozi1989 email:kozi1989@yahoo.gr
139
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
A better, but equivalent, probabilistic description for the HWP is that the alleles for the next generation for any given individual are chosen randomly and independent of each other. Consider two alleles, A and a, with frequencies p and q, respectively, in the population. The different ways to form new genotypes can be derived using a Punnett square, where the fraction in each is equal to the product of the row and column probabilities.
Punnett square for HardyWeinberg equilibrium Females A a (q ) A A a (pq) A a 2 a (q )
( p) Males (p ) a (q )
2
A A (p ) a (pq)
2
The formula is sometimes written as (p ) + (2pq) + (q2)= 1, representing the trivial fact that frequencies must add up to one. The final three possible genotypic frequencies in the offspring become:
These frequencies are called HardyWeinberg frequencies (or HardyWeinberg proportions). This is achieved in one generation, and only requires the assumption of random mating with an infinite population size. Sometimes, a population is created by bringing together males and females with different allele frequencies. In this case, the assumption of a single population is violated until after the first generation, so the first generation will not have HardyWeinberg equilibrium. Successive generations will have HardyWeinberg equilibrium.
54. Population genetics and biology
Factors influencing gene frequencies.Mutations.Migration. Several factors can disturb the Hardy-Weinberg equilibrium by influencing either the distribution of genes in the population or by altering the gene frequencies. These include the following: 1) Non Random Mating Random mating (Panmixis) is the selection of a partner irrespective of his/her genotype. Non-random mating can result in an increase in the frequency of homozygous state i.e. by Assortative mating - the tendency to chose partners who share similar characteristics of heights, racial originate.
Notes made by kozi1989 email:kozi1989@yahoo.gr
140
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Consanguinity - marriage between blood relatives who have at least one common ancestor. Consanguinity in a community results in a relative increase in frequency of affected homozygotes with a relative decrease in the heterozygote frequency. 2) Mutation rate: If a locus shows a higher mutation rate, then the H-W equilibrium will be destroyed and there will be a steady increase in mutant allele. In a population found in H-W equilibrium fitness of affected individuals is balanced by the occurrance of new mutations. The spontaneous mutation rate () varies with different loci. 3) Selection In an `ideal' population there is no selection for or against any particular genotype. Selection is a powerful tool for altering gene frequency. It may reduce (negative selection) or increase (positive selection) a particular phenotype and hence its genotype. Genetic selection acts on the individual phenotype and either hinders or favours reproduction and thus propogation of the individuals genotype. 4) Small population size In a large population the number of children produced by individuals with different genotypes, assuming no alterations in fitness for any particular genotype, will tend to balance out, so the gene frequency will remain stable. However, in small populations, one allele could be transmitted to a higher proportion of offspring by chance, thus resulting in marked changes in allele frequency from one generation to the next, thus disturbing HW equilibrium. This phenomenon is called "random genetic drift". If one allele is lost it is said to be "extinguished" and the other allele is described as having become "fixed". Small population may become "genetic isolates" due to religious, geographical or other reasons. In such populations due to admixing, recessive and late-onset dominant traits increase in frequency, then in the general population. Table 2 gives examples of some genetic isolates with high frequencies of certain autosomal disorders. 5) Gene flow (Migration) When new alleles are introduced into a population as a consequence of migration, and subsequent inter-marriages, this alters allele frequencies. The slow diffusion of alleles across a racial or geographical boundry is known as gene flow e.g. Blood group B is thought to have originated in Asia and slowly spread to other populations, particularly westward due to invasions. In molecular biology and genetics, mutations are changes in a genomic sequence: the DNA sequence of a cell's genome or the DNA or RNA sequence. They can be defined as
Notes made by kozi1989 email:kozi1989@yahoo.gr
141
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
sudden and spontaneous changes in the cell. Mutations are caused by radiation, viruses, transposons and mutagenic chemicals, as well as errors that occur during meiosis or DNA replication. They can also be induced by the organism itself, by cellular processes such as hypermutation. Mutation can result in several different types of change in sequences;(DNA) these can either have no effect, alter the product of a gene, or prevent the gene from functioning properly or completely. In genetics, the mutation rate is the chance of a mutation occurring in an organism or gene in each generation (or, in the case of multicellular organisms, cell division). The mutation frequency is the number of individuals in a population with a particular mutation, and tends to be reported more often as it is easier to measure (for instance, there is no need to restrict the population to experiencing only one generation, as needed to measure mutation rate). In evolutionary biology, mutations can have a neutral, favorable or unfavorable effect on the organism, with respect to the present environment. The effect of a low mutation rate on a population is that few variations are available to respond to sudden environmental change. This means the species is slower to adapt. A higher mutation rate damages more individuals, but by increasing variation in the population could increase the speed at which the population can adapt to changing circumstances. The majority of mutations in a multi-cellular organism's genome are neutral and do not harm the organism. Spontaneous mutation Spontaneous mutations on the molecular level can be caused by: 1) Tautomerism A base is changed by the repositioning of a hydrogen atom, altering the hydrogen bonding pattern of that base resulting in incorrect base pairing during replication. 2) Depurination Loss of a purine base (A or G) to form an apurinic site (AP site). 3) Deamination Hydrolysis changes a normal base to an atypical base containing a keto group in place of the original amine group. Examples include C U and A HX (hypoxanthine), which can be corrected by DNA repair mechanisms; and 5MeC (5methylcytosine) T, which is less likely to be detected as a mutation because thymine is a normal DNA base. 4) Slipped strand mispairing - Denaturation of the new strand from the template during replication, followed by renaturation in a different spot ("slipping"). This can lead to insertions or deletions. Induced mutation 1) Induced mutations on the molecular level can be caused by: 2) Chemicals Hydroxylamine NH2OH 3) Base analogs (e.g. BrdU)
Notes made by kozi1989 email:kozi1989@yahoo.gr
142
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
4) Alkylating agents (e.g. N-ethyl-N-nitrosourea) These agents can mutate both replicating and non-replicating DNA. In contrast, a base analog can only mutate the DNA when the analog is incorporated in replicating the DNA. Each of these classes of chemical mutagens has certain effects that then lead to transitions, transversions, or deletions. 5) Agents that form DNA adducts (e.g. ochratoxin A metabolites)[25] 6) DNA intercalating agents (e.g. ethidium bromide) 7) DNA crosslinkers 8) Oxidative damage Nitrous acid converts amine groups on A and C to diazo groups, altering their hydrogen bonding patterns which leads to incorrect base pairing during replication.
55. Population genetics and biology
Natural selection.Balanced polymorphism.Genetic homeostasis.Isolation. Natural selection is the process by which biologic traits become more or less common in a population due to consistent effects upon the survival or reproduction of their bearers. It is a key mechanism of evolution. The genetic variation within a population of organisms may cause some individuals to survive and reproduce more successfully than others. Factors which affect reproductive success are also important, an issue which Charles Darwin developed in his ideas on sexual selection. Natural selection acts on the phenotype, or the observable characteristics of an organism, but the genetic (heritable) basis of any phenotype which gives a reproductive advantage will become more common in a population (see allele frequency). Over time, this process can result in adaptations that specialize populations for particular ecological niches and may eventually result in the emergence of new species. In other words, natural selection is an important process (though not the only process) by which evolution takes place within a population of organisms. As opposed to artificial selection, in which humans favor specific traits, in natural selection the environment acts as a sieve through which only certain variations can pass. Natural selection is one of the cornerstones of modern biology. The term was introduced by Darwin in his influential 1859 book On the Origin of Species, in which natural selection was described as analogous to artificial selection, a process by which animals and plants with traits considered desirable by human breeders are systematically favored for reproduction. The concept of natural selection was originally developed in the absence of a valid theory of heredity; at the time of Darwin's writing, nothing was known of modern genetics. The union of traditional Darwinian evolution with subsequent discoveries in classical and molecular genetics is termed the modern evolutionary synthesis. Natural selection remains the primary explanation for adaptive evolution. Natural selection can act on any heritable phenotypic trait, and selective pressure
Notes made by kozi1989 email:kozi1989@yahoo.gr
143
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
can be produced by any aspect of the environment, including sexual selection and competition with members of the same or other species. However, this does not imply that natural selection is always directional and results in adaptive evolution; natural selection often results in the maintenance of the status quo by eliminating less fit variants. Polymorphism in biology occurs when two or more clearly different phenotypes exist in the same population of a species in other words, the occurrence of more than one form or morph. In order to be classified as such, morphs must occupy the same habitat at the same time and belong to a panmictic population (one with random mating). Polymorphism is common in nature; it is related to biodiversity, genetic variation and adaptation; it usually functions to retain variety of form in a population living in a varied environment. The most common example is sexual dimorphism, which occurs in many organisms. Other examples are mimetic forms of butterflies (see mimicry), and human haemoglobin and blood types. Polymorphism results from evolutionary processes, as does any aspect of a species. It is heritable, and is modified by natural selection. In polyphenism, an individual's genetic make-up allows for different morphs, and the switch mechanism that determines which morph is shown is environmental. In genetic polymorphism the genetic make-up determines the morph. Ants exhibit both types in a single population. Polymorphism as described here involves morphs of the phenotype. Balancing selection refers to a number of selective processes by which multiple alleles (different versions of a gene) are actively maintained in the gene pool of a population at frequencies above that of gene mutation. This usually happens when the heterozygotes for the alleles under consideration have a higher adaptive value than the homozygote. In this way genetic polymorphism is conserved. There are three main types of natural selection: In directional selection the allele frequency for a trait continuously shifts in one direction. In stabilizing selection the frequency of the alleles of lower fitness decreases until they vanish. Balancing selection is similar but not identical to disruptive selection where individuals of extreme trait values are favored against those with average trait values. These terms are used for quantitative characters controlled by a number of genes. Evidence for balancing selection can be found in the number of alleles in a population which are maintained above mutation rate frequencies. All modern research has shown that this significant genetic variation is ubiquitous in panmictic populations. It is a genetic expression of the field experience of Darwin, Wallace and others, that natural populations in the wild are extraordinarily varied (though not all such variation is of genetic origin). There are several mechanisms (which are not exclusive within any given population)
Notes made by kozi1989 email:kozi1989@yahoo.gr
144
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
by which balancing selection works to maintain polymorphism. The two major and most studied are heterozygote advantage and frequency-dependent selection.
56. Population genetics and biology
Quantitative traits examples, distribution curve Quantitative traits refer to phenotypes (characteristics) that vary in degree and can be attributed to polygenic effects, i.e., product of two or more genes, and their environment. Quantitative trait loci (QTLs) are stretches of DNA containing or linked to the genes that underlie a quantitative trait. Mapping regions of the genome that contain genes involved in specifying a quantitative trait is done using molecular tags such as AFLP or, more commonly SNPs . This is an early step in identifying and sequencing the actual genes underlying trait variation. Polygenic inheritance, also known as quantitative or multifactorial inheritance refers to inheritance of a phenotypic characteristic (trait) that is attributable to two or more genes, or the interaction with the environment, or both. Unlike monogenic traits, polygenic traits do not follow patterns of Mendelian inheritance (separated traits). Instead, their phenotypes typically vary along a continuous gradient depicted by a bell curve. An example of a polygenic trait ishuman skin color . Many genes factor into determining a person's natural skin color, so modifying only one of those genes changes the color only slightly. Many disorders with genetic components are polygenic, including autism, cancer, diabetes and numerous others. Most phenotypic characteristics are the result of the interaction of multiple genes. Examples of disease processes generally considered to be results of multifactorial etiology: Congenital malformation Cleft palate Congenital dislocation of the hip Congenital heart defects Neural tube defects Pyloric stenosis Talipes
Adult onset diseases Diabetes Mellitus Cancer
Notes made by kozi1989 email:kozi1989@yahoo.gr
145
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Epilepsy Glaucoma Hypertension Ischaemic heart disease Manic depression Schizophrenia
Multifactorially inherited diseases are said to constitute the majority of genetic disorders affecting humans which will result in hospitalization or special care of some kind.
57. Theory of evolution
Species.Speciation.Speciation mechanisms In biology, a species is one of the basic units of biological classification and a taxonomic rank. A species is often defined as a group of organisms capable of interbreeding and producing fertile offspring. While in many cases this definition is adequate, more precise or differing measures are often used, such as similarity of DNA, morphology or ecological niche. Presence of specific locally adapted traits may further subdivide species into subspecies. The commonly used names for plant and animal taxa sometimes correspond to species: for example, "lion," "walrus," and "Camphor tree" each refers to a species. In
other cases common names do not: for example, "deer" refers to a family of 34 species, including Eld's Deer, Red Deer and Elk (Wapiti). The last two species were once considered a single species, illustrating how species boundaries may be revised with increased scientific knowledge. Species that are believed to have the same ancestors are grouped together, and this group is called a genus. A species can only belong to one genus that it was grouped into. The belief is best checked by a similarity of their DNA, but for practical reasons, other similar
Notes made by kozi1989 email:kozi1989@yahoo.gr
146
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
properties are used. For plants similarities of flowers are used. All species are given a two part name (called a "binomial name" - "bi" for two, "nomial" for name). The first part of a binomial name is the generic name, the genus of the species. The second part is either the specific name (a term used only in zoology, never in botany, for the second part of a binomial) or the specific epithet (the term always used in botany, which can also be used in zoology). For example, Boa constrictor, which is commonly called by its bionomial name, and is one of five species of the Boa genus. The first part of the name is capitalized, and the second part has a lower case. The two part name is written in italics. A usable definition of the word "species" and reliable methods of identifying particular species are essential for stating and testing biological theories and for measuring biodiversity. Traditionally, multiple examples of a proposed species must be studied for unifying characters before it can be regarded as a species. Extinct species known only from fossils are generally difficult to assign precise taxonomic rankings. Speciation is the evolutionary process by which new biological species arise . The biologist Orator F. Cook seems to have been the first to coin the term 'speciation' for the splitting of lineages or 'cladogenesis,' as opposed to 'anagenesis' or 'phyletic evolution' occurring within lineages.[1][2] Whether genetic drift is a minor or major contributor to speciation is the subject matter of much ongoing discussion. There are four geographic modes of speciation in nature, based on the extent to which speciating populations are geographically isolated from one another: allopatric, peripatric, parapatric, and sympatric. Speciation may also be induced artificially, through animal husbandry or laboratory experiments. Observed examples of each kind of speciation are provided throughout.[3] Natural speciation All forms of natural speciation have taken place over the course of evolution ; however it still remains a subject of debate as to the relative importance of each mechanism in driving biodiversity. One example of natural speciation is the diversity of the three-spined stickleback, a marine fish that, after the last ice age, has undergone speciation into new freshwater colonies in isolated lakes and streams. Over an estimated 10,000 generations, the sticklebacks show structural differences that are greater than those seen between different genera of fish including variations in fins, changes in the number or size of their bony plates, variable jaw structure, and color differences.[5] Allopatric During allopatric (from the ancient Greek allos, "other" + Greek patr, "fatherland") speciation, a population splits into two geographically isolated populations (for example, by habitat fragmentation due to geographical change such as mountain building or social change such as emigration). The isolated populations then undergo
Notes made by kozi1989 email:kozi1989@yahoo.gr
147
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
genotypic and/or phenotypic divergence as: (a) they become subjected to dissimilar selective pressures; (b) they independently undergo genetic drift; (c) different mutations arise in the two populations. When the populations come back into contact, they have evolved such that they are reproductively isolated and are no longer capable of exchanging genes. Island genetics, the tendency of small, isolated genetic pools to produce unusual traits, has been observed in many circumstances, including insular dwarfism and the radical changes among certain famous island chains, for example on Komodo. The Galpagos islands are particularly famous for their influence on Charles Darwin. During his five weeks there he heard that Galpagos tortoises could be identified by island, and noticed that Mockingbirds differed from one island to another, but it was only nine months later that he reflected that such facts could show that species were changeable. When he returned to England, his speculation on evolution deepened after experts informed him that these were separate species, not just varieties, and famously that other differing Galpagos birds were all species of finches. Though the finches were less important for Darwin, more recent research has shown the birds now known as Darwin's finches to be a classic case of adaptive evolutionary radiation. Peripatric In peripatric speciation, a subform of allopatric speciation, new species are formed in isolated, smaller peripheral populations that are prevented from exchanging genes with the main population. It is related to the concept of a founder effect, since small populations often undergo bottlenecks. Genetic drift is often proposed to play a significant role in peripatric speciation. The London Underground mosquito is a variant of the mosquito Culex pipiens that entered in the London Underground in the nineteenth century. Evidence for its speciation include genetic divergence, behavioral differences, and difficulty in mating.[7] Parapatric In parapatric speciation, there is only partial separation of the zones of two diverging populations afforded by geography; individuals of each species may come in contact or cross habitats from time to time, but reduced fitness of the heterozygote leads to selection for behaviours or mechanisms that prevent their inter-breeding. Parapatric speciation is modelled on continuous variation within a 'single', connected habitat acting as a source of natural selection rather than the effects of isolation of habitats produced in peripatric and allopatric speciation. Sympatric Sympatric speciation refers to the formation of two or more descendant species from a single ancestral species all occupying the same geographic location. In sympatric speciation, species diverge while inhabiting the same place. Often-cited
Notes made by kozi1989 email:kozi1989@yahoo.gr
148
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
examples of sympatric speciation are found in insects that become dependent on different host plants in the same area.[8][9] However, the existence of sympatric speciation as a mechanism of speciation is still hotly contested. People have argued that the evidences of sympatric speciation are in fact examples of micro-allopatric, or heteropatric speciation. The most widely accepted example of sympatric speciation is that of the cichlids of Lake Nabugabo in East Africa, which is thought to be due to sexual selection. Until recently, there has a been a dearth of strong evidence that supports this form of speciation, with a general feeling that interbreeding would soon eliminate any genetic differences that might appear. But there has been at least one recent study that suggests that sympatric speciation has occurred in Tennessee cave salamanders. Sympatric speciation driven by ecological factors may also account for the extraordinary diversity of crustaceans living in the depths of Siberia's Lake Baikal. Speciation via polyploidization Polyploidy is a mechanism that has caused many rapid speciation events in sympatry because offspring of, for example, tetraploid x diploid matings often result in triploid sterile progeny.[12] However, not all polyploids are reproductively isolated from their parental plants, and gene flow may still occur for example through triploid hybrid x diploid matings that produce tetraploids, or matings between meiotically unreduced gametes from diploids and gametes from tetraploids (see also hybrid speciation). It has been suggested that many of the existing plant and most animal species have undergone an event of polyploidization in their evolutionary history.[13][14] Reproduction of successful polyploid species is sometimes asexual, by parthenogenesis or apomixis, as for unknown reasons many asexual organisms are polyploid. Rare instances of polyploid mammals are known, but most often result in prenatal death.
58. Theory of evolution
Evolution of higher ranks (macroevolution). Origin of high-rank systematic groups. Direction of evolution. Macroevolution is evolution on a scale of separated gene pools. Macroevolutionary studies focus on change that occurs at or above the level of species, in contrast with microevolution,[2] which refers to smaller evolutionary changes (typically described as changes in allele frequencies) within a species or population. The process of speciation may fall within the purview of either, depending on the forces thought to drive it. Paleontology, evolutionary developmental biology, comparative genomics and genomic phylostratigraphy contribute most of the evidence for the patterns and processes that can be classified as macroevolution. An example of macroevolution is the appearance of feathers during the evolution of birds from theropod dinosaurs. Abrupt transformations from one biologic system to another, for example the passing of life from water into land or the transition from invertebrates to vertebrates, are rare. Few major biological types have emerged during the evolutionary history of life and most of
Notes made by kozi1989 email:kozi1989@yahoo.gr
149
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
them survive till today. When lifeforms take such giant leaps, they meet little to no competition and are able to exploit a plethora of available niches, following a pattern of adaptive radiation. This can lead to convergent evolution, where unrelated populations display similar adaptations. The evolutionary course of Equidae (wide family including all horses and related animals) is often viewed as a typical example of macroevolution. The earliest known genus, Hyracotherium (now reclassified as a palaeothere), was a herbivore animal resembling a dog that lived in the early cenozoic. As its habitat transformed into an open arid grassland, selective pressure required that the animal become a fast grazer. Thus elongation of legs and head as well as reduction of toes gradually occurred, producing the only extant genus of Equidae, Equus. Direction of evolution Highly energetic chemistry is believed to have produced a self-replicating molecule around 4 billion years ago and half a billion years later the last common ancestor of all life existed.[235] The current scientific consensus is that the complex biochemistry that makes up life came from simpler chemical reactions.[236] The beginning of life may have included self-replicating molecules such as RNA,[237] and the assembly of simple cells.[238] Common descent The hominoids are descendants of a common ancestor. All organisms on Earth are descended from a common ancestor or ancestral gene pool. Current species are a stage in the process of evolution, with their diversity the product of a long series of speciation and extinction events. The common descent of organisms was first deduced from four simple facts about organisms: First, they have geographic distributions that cannot be explained by local adaptation. Second, the diversity of life is not a set of completely unique organisms, but organisms that share morphological similarities. Third, vestigial traits with no clear purpose resemble functional ancestral traits and finally, that organisms can be classified using these similarities into a hierarchy of nested groups similar to a family tree. However, modern research has suggested that, due to horizontal gene transfer, this "tree of life" may be more complicated than a simple branching tree since some genes have spread independently between distantly related species. Past species have also left records of their evolutionary history. Fossils, along with the comparative anatomy of present-day organisms, constitute the morphological, or anatomical, record. By comparing the anatomies of both modern and extinct species, paleontologists can infer the lineages of those species. However, this approach is most successful for organisms that had hard body parts, such as shells, bones or teeth. Further, as prokaryotes such as bacteria and archaea share a limited set of common morphologies, their fossils do not provide information on their ancestry. More recently, evidence for common descent has come from the study of biochemical similarities between organisms. For example, all living cells use the same basic
Notes made by kozi1989 email:kozi1989@yahoo.gr
150
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
set of nucleotides and amino acids.[245] The development of molecular genetics has revealed the record of evolution left in organisms' genomes: dating when species diverged through the molecular clock produced by mutations.[246] For example, these DNA sequence comparisons have revealed that humans and chimpanzees share 96% of their genomes and analyzing the few areas where they differ helps shed light on when the common ancestor of these species existed.
59. Theory of evolution
Molecular evolution. Pre-cellular evolution. Phylogenetic relationship between organisms.Genetic differentiation. Molecular evolution is the process of evolution at the scale of DNA, RNA, and proteins. Molecular evolution emerged as a scientific field in the 1960s as researchers from molecular biology, evolutionary biology and population genetics sought to understand recent discoveries on the structure and function of nucleic acids and protein. Some of the key topics that spurred development of the field have been the evolution of enzyme function, the use of nucleic acid divergence as a "molecular clock" to study species divergence, and the origin of noncoding DNA. In natural science, abiogenesis (pronounced /eba.dnss/ ay-by-oh-jen--siss) or biopoesis is the study of how biological life arises from inorganic matter through natural processes, and the method by which life on Earth arose. Most amino acids, often called "the building blocks of life", can form via natural chemical reactions unrelated to life, as demonstrated in the MillerUrey experiment and similar experiments that involved simulating some of the conditions of the early Earth in a laboratory.[1] In all living things, these amino acids are organized into proteins, and the construction of these proteins is mediated by nucleic acids, that are themselves synthesized through biochemical pathways catalysed by proteins. Which of these organic molecules first arose and how they formed the first life is the focus of abiogenesis. In any theory of abiogenesis, two aspects of life have to be accounted for: replication and metabolism. The question of which came first gave rise to different types of theories. In the beginning, metabolism-first theories (Oparin coacervate) were proposed, and only later thinking gave rise to the modern, replication-first approach. In modern, still somewhat limited understanding, the first living things on Earth are thought to be single cell prokaryotes (which lack a cell nucleus), perhaps evolved from protobionts (organic molecules surrounded by a membrane-like structure).[2] The oldest ancient fossil microbe-like objects are dated to be 3.5 Ga (billion years old), approximately one billion years after the formation of the Earth itself.[3][4] By 2.4 Ga, the ratio of stable isotopes of carbon, iron and sulfur shows the action of living things on inorganic minerals and sediments[5][6] and molecular biomarkers indicate photosynthesis, demonstrating that life on Earth was widespread by this time.
Notes made by kozi1989 email:kozi1989@yahoo.gr
151
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
In biology, phylogenetics is the study of evolutionary relatedness among groups of organisms (e.g. species, populations), which is discovered through molecular sequencing data and morphological data matrices. The term phylogenetics derives from the Greek terms phyle () and phylon (), denoting tribe and race; and the term genetikos (), denoting relative to birth, from genesis () birth. Taxonomy, the classification, identification, and naming of organisms, is richly informed by phylogenetics, but remains methodologically and logically distinct. [1] The fields of phylogenetics and taxonomy overlap in the science of phylogenetic systematics one methodology, cladism (also cladistics) shared derived characters (synapomorphies) used to create ancestor-descendant trees (cladograms) and delimit taxa (clades).[2][3] In biological systematics as a whole, phylogenetic analyses have become essential in researching the evolutionary tree of life. Grouping of organisms Phylogenetic groups, or taxa, can be monophyletic, paraphyletic, or polyphyletic. There are some terms that describe the nature of a grouping in such trees. For instance, all birds and reptiles are believed to have descended from a single common ancestor, so this taxonomic grouping (yellow in the diagram below) is called monophyletic. "Modern reptile" (cyan in the diagram) is a grouping that contains a common ancestor, but does not contain all descendants of that ancestor (birds are excluded). This is an example of a paraphyletic group. A grouping such as warm-blooded animals would include only mammals and birds (red/orange in the diagram) and is called polyphyletic because the members of this grouping do not include the most recent common ancestor. Human genetic variation refers to genetic differences both within and among populations. There may be multiple variants of any given gene in the human population (alleles), leading to polymorphism. Many genes are not polymorphic, meaning that only a single allele is present in the population: that allele is then said to be fixed. No two humans are genetically identical. Even monozygotic twins, who develop from one zygote, have infrequent genetic differences due to mutations occurring during development and gene copy number variation has been observed.[2] Differences between individuals, even closely related individuals, are the key to techniques such as genetic fingerprinting. Alleles occur at different frequencies in different human populations, with populations that are more geographically and ancestrally remote tending to differ more. Causes of differences between individuals include the exchange of genes during meiosis and various mutational events. There are at least two reasons why genetic variation exists between populations. Natural selection may confer an adaptive advantage to individuals in a specific environment if an allele provides a competitive advantage. Alleles under selection are likely to occur only in those geographic regions where they confer an advantage. The second main cause of genetic variation is due to the high degree of neutrality of most mutations. Most mutations do not appear to have any selective effect one way or the other on the organism. The main cause is genetic drift, this is the effect of
Notes made by kozi1989 email:kozi1989@yahoo.gr
152
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
random changes in the gene pool. In humans, founder effect and past small population size (increasing the likelihood of genetic drift) may have had an important influence in neutral differences between populations.
60. Human evolution
Anthropogenesis. Human races. Origin of the human races. Factors of racial diversification. Racism. Human evolution is the phenotypic history of the genus Homo, including the emergence of Homo sapiens as a distinct species and as a unique category of hominids ("great apes") and mammals. The study of human evolution uses many scientific disciplines, including physical anthropology, primatology, archaeology, linguistics and genetics. The term "human" in the context of human evolution refers to the genus Homo, but studies of human evolution usually include other hominids, such as the Australopithecines, from which the genus Homo had diverged by about 2.3 to 2.4 million years ago in Africa.[2][3] Scientists have estimated that humans branched off from their common ancestor with chimpanzees about 57 million years ago. Several species and subspecies of Homo evolved and are now extinct, introgressed or extant. Examples include Homo erectus (which inhabited Asia, Africa, and Europe) and Neanderthals (either Homo neanderthalensis or Homo sapiens neanderthalensis) (which inhabited Europe and Asia). Archaic Homo sapiens evolved between 400,000 and 250,000 years ago. A human race is defined as a group of people with certain common inherited features that distinguish them from other groups of people. All men of whatever race are currently classified by the anthropologist or biologist as belonging to the one species, Homo sapiens. This is another way of saying that the differences between human races are not great, even though they may appear so, i.e. black vs white skin. All races of mankind in the world can interbreed because they have so much in common. Most anthropologists recognize 3 or 4 basic races of man in existence today. These races can be further subdivided into as many as 30 subgroups. The Australoid or Australian Aborigine, is sometimes regarded as a sub-group of the Caucasoid as they have many features in common with this group despite their dark skin. The American Indian is usually classed with the Mongoloid division. The most significant difference between these races is: Language (b) hair and facial features (c) skin color. This distinguishing feature is primarily due to difference in amount of the pigment, melanin, in the skin. This melanin protects the body by absorbing ultraviolet (UV) radiation from the sunlight which falls on the skin. UV radiation can damage the skin and produce skin cancer if it is not filtered out in this way. The shades of color in human skin can be represented by the Von Luschan Scale and ranges from almost pure white to pure black. Differences such as hair, skin and facial features are almost certainly inherited, but the way in which their inheritance is controlled is little understood, and definitely not
Notes made by kozi1989 email:kozi1989@yahoo.gr
153
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
simple. In skin color, for instance, 2 to 4 pairs of genes are involved. (Genes are the units in each cell of the body that carry inherited information) It is known that some of these genes produce lightness and some darkness. The brown skinned person has genes for both dark and light. The darker person has more genes for darkness and the lighter person has more genes for lightness. Scientific study of human evolution is concerned, primarily, with the development of the genus Homo, but usually involves studying other hominids and hominines as well, such as Australopithecus. "Modern humans" are defined as the Homo sapiens species, of which the only extant subspecies is known as Homo sapiens sapiens. Homo sapiens idaltu (roughly translated as "elder wise human"), the other known subspecies, is now extinct.[16] Homo neanderthalensis, which became extinct 30,000 years ago, has sometimes been classified as a subspecies, "Homo sapiens neanderthalensis"; genetic studies now suggest that the functional DNA of modern humans and Neanderthals diverged 500,000 years ago.[17] Similarly, the discovered specimens of the Homo rhodesiensis species have been classified by some as a subspecies, but this classification is not widely accepted. Anatomically modern humans first appear in the fossil record in Africa about 195,000 years ago, and studies of molecular biology give evidence that the approximate time of divergence from the common ancestor of all modern human populations was 200,000 years ago.[18][19][20][21][22] The broad study of African genetic diversity headed by Dr. Sarah Tishkoff found the San people to express the greatest genetic diversity among the 113 distinct populations sampled, making them one of 14 "ancestral population clusters". The research also located the origin of modern human migration in south-western Africa, near the coastal border of Namibia and Angola. The evolutionary history of primates can be traced back 65 million years. Primates are one of the oldest of all surviving placental mammal groups. The oldest known primatelike mammal species (those of the genus Plesiadapis) come from North America, but inhabited Eurasia and Africa on a wide scale during the tropical conditions of the Paleocene and Eocene. Molecular evidence suggests that the last common ancestor between humans and the remaining great apes diverged 48 million years ago. The gorillas were the first group to split, then the chimpanzees (genus Pan) split off from the line leading to the humans. The functional portion of human DNA is approximately 98.4% identical to that of chimpanzees when comparing single nucleotide polymorphisms (see human evolutionary genetics). Therefore, the closest living relatives of humans are gorillas and chimpanzees, as they share a relatively recent common ancestor.[24] Humans are probably most closely related to two chimpanzee species: the Common Chimpanzee and the Bonobo.[24] Full genome sequencing has resulted in the conclusion that "after 6.5 [million] years of separate evolution, the differences between chimpanzee and human are ten times greater than those between two unrelated people and ten times less than those between rats and mice".[attribution needed] Current estimates of suggested concurrence between functional human and chimpanzee DNA sequences range between 95% and 99%. Early estimates indicated that the human lineage may have diverged from
Notes made by kozi1989 email:kozi1989@yahoo.gr
154
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
that of chimpanzees about five million years ago, and from that of gorillas about eight million years ago. However, a hominid skull discovered in Chad in 2001, classified as Sahelanthropus tchadensis, is approximately seven million years old, and may be evidence of an earlier divergence. Human evolution is characterised by a number of important changes morphological, developmental, physiological, and behaviouralwhich have taken place since the split between the last common ancestor of humans and chimpanzees. The first major morphological change was the evolution of a bipedal locomotor adaptation from an arboreal or semi-arboreal one,[30] with all its attendant adaptations (a valgus knee, low intermembral index (long legs relative to the arms), reduced upper-body strength). The human species developed a much larger brain than that of other primates typically 1,400 cm in modern humans, over twice the size of that of a chimpanzee or gorilla. The pattern of human postnatal brain growth differs from that of other apes (heterochrony), and allows for extended periods of social learning and language acquisition in juvenile humans. Physical anthropologists[who?] argue that the differences between the structure of human brains and those of other apes are even more significant than their differences in size. Racism is the belief that there are inherent differences in people's traits and capacities that are entirely due to their race, however defined, and that, as a consequence, justify the different treatment of those people, both socially and legally. Moreover, racism is the practice of the different treatment of certain a group or groups, which is then justified by recourse to racial stereotyping or pseudo-science. Those who disagree with the proposition that there are races or that there are such inherent (i.e., non-personal, social, or cultural) differences regard any differences in treatment of people on the basis of those criteria as being racial discrimination. Some of those who argue that there are such inherent differences also argue that one race is inferior to another race.[1][2][3] In the case of institutional racism, certain racial groups may be denied rights or benefits, or receive preferential treatment. Racial discrimination typically points out taxonomic differences between different groups of people, although anyone may be discriminated against on an ethnic or cultural basis, independent of their somatic differences. According to the United Nations conventions, there is no distinction between the term racial discrimination and ethnicity discrimination. There is some evidence that the meaning of the term has changed over time, and that earlier definitions of racism involved the simple belief that human populations are divided into separate races.[4] Many biologists, anthropologists, and sociologists reject this taxonomy in favor of more specific and/or empirically verifiable criteria, such as geography, ethnicity, or a history of endogamy.
61. Ecology. Interaction between humans and environment
Populations, communities, ecosystems.The overpopulation problem.
Notes made by kozi1989 email:kozi1989@yahoo.gr
155
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
A population is all the organisms that both belong to the same species and live in the same geographical area. The area that is used to define the population is such that inter-breeding is possible between any pair within the area and more probable than cross-breeding with individuals from other areas. Normally breeding is substantially more common within the area than across the border. In ecology, a community is an assemblage of two or more populations of different species occupying the same geographical area. The term community has a variety of uses. In its simplest form it refers to groups of organisms in a specific place and/or time, for example, "the fish community of Lake Ontario before industrialization". Community ecologists study the interactions between species in communities on many spatial and temporal scales, including the distribution, structure, abundance, demography, and interactions between coexisting populations.[1] The primary focus of community ecology is on the interactions between populations as determined by specific genotypic and phenotypic characteristics. An ecosystem is a biological environment consisting of all the organisms living in a particular area, as well as all the nonliving, physical components of the environment with which the organisms interact, such as air, soil, water and sunlight. [1] It is all the organisms in a given area, along with the nonliving (abiotic) factors with which they interact; a biological community and its physical environment. The entire array of organisms inhabiting a particular ecosystem is called a community.[1] In a typical ecosystem, plants and other photosynthetic organisms are the producers that provide the food.[1] Ecosystems can be permanent or temporary. Ecosystems usually form a number of food webs. Ecosystems are functional units consisting of living things in a given area, non-living chemical and physical factors of their environment, linked together through nutrient cycle and energy flow. 1) 2) Natural Terrestrial ecosystem Aquatic ecosystem Artificial, ecosystems created by humans.
Central to the ecosystem concept is the idea that living organisms interact with every other element in their local environment. Eugene Odum, a founder of ecology, stated: "Any unit that includes all of the organisms (ie: the "community") in a given area interacting with the physical environment so that a flow of energy leads to clearly defined trophic structure, biotic diversity, and material cycles (i.e.: exchange of materials between living and nonliving parts) within the system is an ecosystem." Overpopulation is a condition where an organism's numbers exceed the carrying capacity of its habitat. The term often refers to the relationship between the human population and its environment, the Earth.
Notes made by kozi1989 email:kozi1989@yahoo.gr
156
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Without farming, the world population would probably have reached half a million by now."*2+ The worlds population has significantly increased in the last 50 years, mainly due to medical advancements and substantial increases in agricultural productivity. Overpopulation does not depend only on the size or density of the population, but on the ratio of population to available sustainable resources. It also depends on the way resources are used and distributed throughout the population. Overpopulation can result from an increase in births, a decline in mortality rates due to medical advances, from an increase in immigration, or from an unsustainable biome and depletion of resources. It is possible for very sparsely populated areas to be overpopulated, as the area in question may have a meager or non-existent capability to sustain human life (e.g. a desert). The resources to be considered when evaluating whether an ecological niche is overpopulated include clean water, clean air, food, shelter, warmth, and other resources necessary to sustain life. If the quality of human life is addressed, there may be additional resources considered, such as medical care, education, proper sewage treatment and waste disposal. Overpopulation places competitive stress on the basic life sustaining resources,[9] leading to a diminished quality of life.
62. Behaviour genetic basis and adaptive value
Components of behaviour. Social organization and sociobiology.Strategy of natural section. Behavioural genetics is the field of study that examines the role of genetics in animal (including human) behaviour. Often associated with the "nature versus nurture" debate, behavioural genetics is highly interdisciplinary, involving contributions from biology, genetics, ethology, psychology, and statistics. Behavioural geneticists study the inheritance of behavioural traits. In humans, this information is often gathered through the use of the twin study or adoption study. In animal studies, breeding, transgenesis, and gene knockout techniques are common; psychiatric genetics is a closely related field. The primary methods of modern human behavioural genetics are twin and adoption studies. These techniques have been expanded upon in recent decades to include multivariate genetic analysis, extremes analysis, and more recently genomic techniques. Sociobiology is a field of scientific study which is based on the assumption that social behavior has resulted from evolution and attempts to explain and examine social behavior within that context. Often considered a branch of biology and sociology, it also draws from ethology, anthropology, evolution, zoology, archaeology, population genetics, and other disciplines. Within the study of human societies, sociobiology is very closely allied to the fields of human behavioral ecology and evolutionary psychology. Sociobiology investigates social behaviors, such as mating patterns, territorial fights, pack hunting, and the hive society of social insects. It argues that just as selection pressure led to animals evolving useful ways of interacting with the natural environment, it led to the genetic evolution of advantageous social behavior.
Notes made by kozi1989 email:kozi1989@yahoo.gr
157
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
While the term "sociobiology" can be traced to the 1940s, the concept didn't gain major recognition until 1975 with the publication of Edward O. Wilson's book, Sociobiology: The New Synthesis. The new field quickly became the subject of heated controversy. Criticism, most notably made by Richard Lewontin and Stephen Jay Gould, centered on sociobiology's contention that genes play an ultimate role in human behavior and that traits such as aggressiveness can be explained by biology rather than a person's social environment. Sociobiologists generally responded to the criticism by pointing to the complex relationship between nature and nurture. Sociology Sociol the formation of a stable structure of relations inside a group, which provides a basis for order and patterns relationships for new members Sociology is the study of society.[1] It is a social sciencea term with which it is sometimes synonymouswhich uses various methods of empirical investigation[2] and critical analysis[3] to develop and refine a body of knowledge about human social activity.
Natural selection is the process by which biologic traits become more or less common in a population due to consistent effects upon the survival or reproduction of their bearers. It is a key mechanism of evolution. The genetic variation within a population of organisms may cause some individuals to survive and reproduce more successfully than others. Factors which affect reproductive success are also important, an issue which Charles Darwin developed in his ideas on sexual selection. Natural selection acts on the phenotype, or the observable characteristics of an organism, but the genetic (heritable) basis of any phenotype which gives a reproductive advantage will become more common in a population (see allele frequency). Over time, this process can result in adaptations that specialize populations for particular ecological niches and may eventually result in the emergence of new species. In other words, natural selection is an important process (though not the only process) by which evolution takes place within a population of organisms. As opposed to artificial selection, in which humans favor specific traits, in natural selection the environment acts as a sieve through which only certain variations can pass Natural selection can act on any heritable phenotypic trait, and selective pressure can be produced by any aspect of the environment, including sexual selection and competition with members of the same or other species. However, this does not imply that natural selection is always directional and results in adaptive evolution; natural selection often results in the maintenance of the status quo by eliminating less fit variants The unit of selection can be the individual or it can be another level within the hierarchy of biological organisation, such as genes, cells, and kin groups. There is still debate about whether natural selection acts at the level of groups or species to produce
Notes made by kozi1989 email:kozi1989@yahoo.gr
158
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
adaptations that benefit a larger, non-kin group. Likewise, there is debate as to whether selection at the molecular level prior to gene mutations and fertilization of the zygote should be ascribed to conventional natural selection because traditionally natural selection is an environmental and exterior force that acts upon a phenotype typically after birth. Some science journalists distinguish natural selection from gene selection by informally referencing selection of mutations as "pre-selection." Selection at a different level such as the gene can result in an increase in fitness for that gene, while at the same time reducing the fitness of the individuals carrying that gene, in a process called intragenomic conflict. Overall, the combined effect of all selection pressures at various levels determines the overall fitness of an individual, and hence the outcome of natural selection. Natural selection occurs at every life stage of an individual. An individual organism must survive until adulthood before it can reproduce, and selection of those that reach this stage is called viability selection. In many species, adults must compete with each other for mates via sexual selection, and success in this competition determines who will parent the next generation. When individuals can reproduce more than once, a longer survival in the reproductive phase increases the number of offspring, called survival selection.
63. Ecology of parasitism
Parasitism as a biological phenomenon. Parasites and hosts Parasitism is a type of symbiotic relationship between organisms of different species where one organism, the parasite, benefits at the expense of the other, the host. Traditionally parasite referred to organisms with lifestages that went beyond one host (e.g. Taenia solium), which are now called macroparasites (typically protozoa and helminths). Parasites can now also refer to microparasites, which are typically smaller, such as viruses and bacteria and can be directly transmitted between hosts of one species. Unlike predators, parasites are generally much smaller than their host, although both are special cases of consumer-resource interactions.[1] Parasite show a high degree of specialization for their mode of life, and reproduce at a faster rate than their hosts. Classic examples of parasitism include interactions between vertebrate hosts and diverse animals such as tapeworms, flukes, the Plasmodium species, and fleas. Parasitism is differentiated from the parasitoid relationship, though not sharply, by the fact that parasitoids generally kill or sterilise their hosts. Parasitoidy occurs in about as many classes of organism as parasitism does. The harm and benefit in parasitic interactions concern the biological fitness of the organisms involved. Parasites reduce host fitness in many ways, ranging from general or specialized pathology (such as parasitic castration), impairment of secondary sex characteristics, to the modification of host behaviour. Parasites increase their fitness by exploiting hosts for resources necessary for the parasite's survival: (i.e. food, water, heat, habitat, and dispersal). Although the concept of parasitism applies unambiguously to many cases in nature, it
Notes made by kozi1989 email:kozi1989@yahoo.gr
159
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
is best considered part of a continuum of types of interactions between species, rather than an exclusive category. Particular interactions between species may satisfy some but not all parts of the definition. In many cases, it is difficult to demonstrate that the host is harmed. In others, there may be no apparent specialization on the part of the parasite, or the interaction between the organisms may be short-lived. In medicine, only eukaryotic organisms are considered parasites, with the exclusion of bacteria and viruses. Some branches of biology, however, regard members of these groups as parasitic. In biology, a host is an organism that harbors a parasite (that is, a virus, a bacterium, a protozoan, or a fungus), or a mutual or commensal symbiont, typically providing nourishment and shelter. In botany, a host plant is one that supplies food resources and substrate for certain insects or other fauna. Examples of such interactions include a cell being host to a virus, a legume plant hosting helpful nitrogen-fixing bacteria, and animals as hosts to parasitic worms, e.g. nematodes A primary host or definitive host is a host in which the parasite reaches maturity and, if applicable, reproduces sexually. A secondary host or intermediate host is a host that harbors the parasite only for a short transition period, during which (usually) some developmental stage is completed. A parasite is an organism that grows, feeds, and is sheltered on or in a different organism while contributing nothing to the survival of its host.
64. Ecology of parasitism
Origin of parasitism, parasite adaptation to the host. Host parasite interactions Parasites infect hosts that exist within their same geographical area (sympatric) more effectively. This phenomenon supports the "Red Queen hypothesis - which states that interactions between species (such as host and parasites) lead to constant natural selection for adaptation and counter adaptation. The parasites track the locally common host phenotypes, therefore the parasites are less infective to allopatric (from different geographical region) hosts.Parasites are classified based on their interactions with their hosts and on their life cycles. Parasites that live on the surface of the host are called ectoparasites (e.g. some mites) and those that live inside the host are called endoparasites (including all parasitic worms). Endoparasites can exist in one of two forms: intercellular (inhabiting spaces in the hosts body) or intracellular (inhabiting cells in the hosts body). Intracellular parasites, such as bacteria or viruses, tend to rely on a third organism which is generally known as the carrier or vector. The vector does the job of transmitting them to the host. An example of this interaction is the transmission of malaria, caused by a protozoan of the genus Plasmodium, to humans by the bite of an anopheline mosquito. An epiparasite is one that feeds on another parasite. This relationship is also sometimes referred to as hyperparasitism which may be exemplified by a protozoan (the hyperparasite) living in the digestive tract of a flea living on a dog.
Notes made by kozi1989 email:kozi1989@yahoo.gr
160
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Social parasites take advantage of interactions between members of social organisms such as ants or termites. In kleptoparasitism, parasites appropriate food gathered by the host. An example is the brood parasitism practiced by many species of cuckoo and cowbird, which do not build nests of their own but rather deposit their eggs in nests of other species and abandon them there. The host behaves as a "babysitter" as they raise the young as their own. If the host removes the cuckoo's eggs, some cuckoos will return and attack the nest to compel host birds to remain subject to this parasitism. The cowbirds parasitism does not necessarily harm its hosts brood; however, the cuckoo may remove one or more host eggs to avoid detection, and furthermore the young cuckoo may heave the hosts eggs and nestlings from the nest. Parasitism can take the form of isolated cheating or exploitation among more generalized mutualistic interactions. For example, broad classes of plants and fungi exchange carbon and nutrients in common mutualistic mycorrhizal relationships; however, some plant species known as myco-heterotrophs "cheat" by taking carbon from a fungus rather than donating it. Parasitoids are organisms whose larval development occurs inside or on the surface of another organism, resulting in the death of the host. This means that the interaction between the parasitoid and the host is fundamentally different from that of a true parasite and shares some of the characteristics of predation.
Parasitology
65. Characteristics of Protozoa. Phylum Sarcomastigophora. Subphylum
Mastigophora (Flagellata). Leismania donovani.Leismania tropica. Protozoa (from the Greek words , proto, meaning first, and , zoa, meaning animals) are a diverse group of single-cell eukaryotic organisms, many of which are motile. In general, protozoans are referred to as animal-like protists because of movement (motile). However, both protozoa and protists are paraphyletic groups (not including all genetic relatives of the group). The most important protozoans range usually from 10 to 52 micrometers, but can grow as large as 1 mm, and are seen easily by microscope. They were considered formerly to be part of the protista family. Protozoa exist throughout aqueous environments and soil, occupying a range of trophic levels. Motility and digestion Tulodens are one of the slow-moving form of protozoans. They move around with whip-like tails called flagella, hair-like structures called cilia, or foot-like structures called pseudopodia. Others do not move at all.
Notes made by kozi1989 email:kozi1989@yahoo.gr
161
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Protozoa may absorb food via their cell membranes, some, e.g., amoebas, surround food and engulf it, and yet others have openings or "mouth pores" into which they sweep food. All protozoa digest their food in stomach-like compartments called vacuoles. As components of the micro- and meiofauna, protozoa are an important food source for microinvertebrates. Thus, the ecological role of protozoa in the transfer of bacterial and algal production to successive trophic levels is important. As predators, they prey upon unicellular or filamentous algae, bacteria, and microfungi . Protozoa are both herbivores and consumers in the decomposer link of the food chain. They also control bacteria populations and biomass to some extent. Protozoa such as the malaria parasites (Plasmodium spp.), trypanosomes and leishmania, are also important as parasites and symbionts of multicellular animals. Some protozoa have life stages alternating between proliferative stages (e.g., trophozoites) and dormant cysts. As cysts, protozoa can survive harsh conditions, such as exposure to extreme temperatures or harmful chemicals, or long periods without access to nutrients, water, or oxygen for a period of time. Being a cyst enables parasitic species to survive outside of a host, and allows their transmission from one host to another. When protozoa are in the form of trophozoites (Greek, tropho = to nourish), they actively feed. The conversion of a trophozoite to cyst form is known as encystation, while the process of transforming back into a trophozoite is known as excystation. Protozoa can reproduce by binary fission or multiple fission. Some protozoa reproduce sexually, some asexually, while some use a combination, (e.g., Coccidia). An individual protozoon is hermaphroditic. Leishmania donovani: Visceral leishmaniasis (Kala-azar) is spread through an insect vector, the sandfly of the Phlebotomus genus in the Old World and the Lutzomyia genus in the New World. Sandflies are tiny creatures, 3-6 millimeters long by 1.5-3 millimeters in diameter, and found in tropical or temperate regions throughout the world. Sandfly larvae grow in warm, moist organic matter, such as old trees, house walls or waste making them hard to eradicate. The adult female sand fly is a bloodsucker, usually feeding at night on sleeping prey. When the fly bites an animal infected with L. donovani, the pathogen is ingested along with the preys blood. At this point the protozoan is in the smaller of its two forms, called an amastigote round, non-motile, and only three to seven micrometers in diameter. Life cycle Taken into the stomach of the sandfly, the amastigotes quickly transform into a second L. donovani form, called the promastigote.This form is spindle-shaped, triple the size of the amastigote, and has a single flagellum that allows for motility. The promastigotes live extracellularly in the sandflys alimentary canal, reproducing asexually, then migrate to the proximal end of the gut where they become poised for a regurgitational transmission . This is their means of transmission back into a mammalian host, as the fly injects its saliva into prey when it bites. The promastigotes are introduced locally at the bite site along with
Notes made by kozi1989 email:kozi1989@yahoo.gr
162
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
the flys saliva. Once inside the new host, promastigotes invade macrophages . Once inside, they transform back into the smaller amastigote form. As an amastigote, L. donovani can only reproduce intracellularly and the amastigotes replicate in the most hostile part of the macrophage cell, inside the phagolysosome, whose normal defensive response they are able to prevent. After they have reproduced to a certain extent, the L. donovani lyse their host cell by sheer pressure of mass, but there is some recent speculation that they are able to leave the cell by triggering the exocytosis response of the macrophage. The daughter cell protozoans then migrate through the bloodstream to find new macrophage hosts. In time, L. donovani becomes a systemic infection, spreading to all the hosts organs, particularly the spleen and liver. Leishmania tropica is a species of flagellate parasites that infects humans and rodents. It can cause a disease called oriental sore which is a form of cutaneous leishmaniasis.Leishmania tropica is a single-celled trypanosome parasite responsible for causing cutaneous Leishmaniasis. Its prevalence in several countries around the world is apparent by the many names it has adopted: "Baghdad Boil", "Jericho buttons", and "Oriental sore", to name a few. Cutaneous Leishmaniasis develops at the site of a sand fly bite. The sand fly, which carries the promastigote stage of L. tropica in its blood, is the vector. Life cycle Leishmaniasis is transmitted by the bite of infected female phlebotomine sandflies. The life cycle of Leishmania Tropica is identical to that of other related parasites of the same genus and includes both an amastigote and a promastigote stage. The sand flies inject the infective stage of promastigote. The promastigote stage is considered to be part of the infective stage, in which a sand fly infects a host with the parasite through feeding. The amastigote is part of the tissue stage in which the parasite transforms after being engulfed by a macrophage.
66. Phylum Sacromastigophora. Lamblia intestinalis. (Giardia lamblia)
Giardia lamblia (synonymous with Lamblia intestinalis and Giardia duodenalis) is a Flagellated protozoan parasite that Colonizes and reproduces in the small intestine, Causing giardiasis.
The giardia parasite attaches to the epithelium by a ventral adhesive disc, and reproduces via binary fission. Giardiasis does not spread via the bloodstream, nor does it spread to other parts of the gastro-intestinal tract, but remains confined to the lumen of the small intestine. Giardia trophozoites absorb their nutrients from the lumen of the small intestine, and are anaerobes. If the organism is split and stained, it has a very characteristic pattern that resembles a familiar "smiley face" symbol. Chief pathways of human infection include ingestion of untreated sewage, a phenomenon particularly common in many
Notes made by kozi1989 email:kozi1989@yahoo.gr
163
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
developing countries, contamination of natural waters also occurs in watersheds where intensive grazing occurs. Giardia infects humans, but is also one of the most common parasites infecting cats, dogs and birds. Mammalian hosts also include cows, beavers, deer, and sheep. Giardia infection can occur through ingestion of dormant cysts in contaminated water, food, or by the faecal-oral route (through poor hygiene practices). The Giardia cyst can survive for weeks to months in cold water, and therefore can be present in contaminated wells and water systems, especially stagnant water sources such as naturally occurring ponds, storm water storage systems, and even clean-looking mountain streams. They may also occur in city reservoirs and persist after water treatment, as the Giardia cysts are resistant to conventional water treatment methods such as chlorination and ozonolysis. Zoonotic transmission is also possible, and therefore Giardia infection is a concern for people camping in the wilderness or swimming in contaminated streams or lakes, especially the artificial lakes formed by beaver dams (hence the popular name for giardiasis, "Beaver Fever"). In addition to waterborne sources, fecal-oral transmission can also occur, for example in day care centers, where children may have poor hygiene practices. Those who work with children are also at risk of being infected, as are family members of infected individuals. Not all Giardia infections are symptomatic, and many people can unknowingly serve as carriers of the parasite. Life cycle The life cycle begins with a noninfective cyst being excreted with the feces of an infected individual. The cyst is hardy, providing protection from various degrees of heat and cold, desiccation, and infection from other organisms. A distinguishing characteristic of the cyst is four nuclei and a retracted cytoplasm. Once ingested by a host, the trophozoite emerges to an active state of feeding and motility. After the feeding stage, The trophozoite undergoes asexual replication through longitudinal binary fission. The resulting trophozoites and cysts then pass through the digestive system in the faeces. While the trophozoites may be found in the faeces, only the cysts are capable of surviving outside of the host. Distinguishing features of the trophozoites are large karyosomes and lack of peripheral chromatin, giving the two nuclei a halo appearance. Cysts are distinguished by a retracted cytoplasm. This protozoan lacks mitochondria, although the discovery of the presence of mitochodrial remnants organelles in one recent study "indicate that Giardia is not primitively amitochondrial and that it has retained a functional organelle derived from the original mitochondrial endosymbiont". This organelle is now termed a mitosome.
Notes made by kozi1989 email:kozi1989@yahoo.gr
164
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
67. Phylum Sacromastigophora. Trichomonas hominis. Trichomonas vaginalis.
Trichomonas hominis has no cystic stage. The trophozoite measures from 5-15m in length by 7-10m in width. The shape is pyriform and has an axostyle which runs from the nucleus down the centre of the body and extends from the end of the body and undulating membrane which extends the entire length of the body and projects from the body like a free flagellum. It has 4 free flagella and a single nucleus at the anterior end .Trichomonas hominis is found in the large intestine where it forms a harmless infection. Trichomonas vaginalis is an anaerobic, flagellated protozoan, a form of microorganism. The parasitic microorganism is the causative agent of trichomoniasis, and is the most common pathogenic protozoan infection of humans in industrialized countries. Infection rates between men and women are the same with women showing symptoms while infections in men are usually asymptomatic. Transmission takes place directly because the trophozoite does not have a cyst. The T. vaginalis trophozoite is oval as well as flagellated. It is slightly larger than a white blood cell, measuring 9 X 7 m. Five flagella arise near the cytostome; four of these immediately extend outside the cell together, while the fifth flagellum wraps backwards along the surface of the organism. The functionality of the fifth flagellum is not known. In addition, a conspicuous barb-like axostyle projects opposite the four-flagella bundle; the axostyle may be used for attachment to surfaces and may also cause the tissue damage noted in trichomoniasis infections. Incubation Period is typically 5 to 28 days Trichomonas vaginalis is pathogenic in the genitourinary tract In women, it lives in the vagina and the Skene's glands (of the urethra) In men, it lives in the urethra Multiplies when vaginal conditions become more basic than usual (normal pH is 3.8 to 4.2) No known cystic form
68. Phylum Sacromastigophora. Trypanosoma rhodesiense and Trypanosoma
gambience (Trypanosoma brucei). Trypanosoma equiperdum. Trypanosoma cruzi. Trypanosoma brucei is a parasitic protist species that causes African trypanosomiasis (or sleeping sickness) in humans and nagana in animals in Africa. There are 3 sub-species of T. brucei: T. b. brucei, T. b. gambiense and T. b. rhodesiense. These obligate parasites have two hosts - an insect vector and mammalian host. Due to the large difference between these hosts the trypanosome undergoes complex changes during its life cycle to facilitate its survival in the insect gut and the mammalian bloodstream. It also features a unique and notable variable surface glycoprotein (VSG) coat in order to avoid the host's immune system. The insect vector for T. brucei is the tsetse fly. The parasite lives in the midgut of the fly (procyclic form), whereupon it migrates to the salivary glands for injection to the
Notes made by kozi1989 email:kozi1989@yahoo.gr
165
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
mammalian host on biting. The parasite lives within the bloodstream (bloodstream form) where it can reinfect the fly vector after biting. Later during a T. brucei infection the parasite may migrate to other areas of the host. A T. brucei infection may be transferred human to human via bodily fluid exchange, primarily blood transfer. Life cycle The tsetse fly (genus Glossina) is a large, brown biting fly that serves as both a host and vector for the Trypanosome parasites. While taking blood from a mammalian host, an infected tsetse fly injects metacyclic trypomastigotes into skin tissue. From the bite, parasites first enter the lymphatic system and then pass into the bloodstream . Inside the mammalian host, they transform into bloodstream trypomastigotes, and are carried to other sites throughout the body, reach other blood fluids (e.g., lymph, spinal fluid), and continue to replicate by binary fission. The entire life cycle of African trypanosomes is represented by extracellular stages. A tsetse fly becomes infected with bloodstream trypomastigotes when taking a blood meal on an infected mammalian host. In the fly's midgut, the parasites transform into procyclic trypomastigotes, multiply by binary fission, leave the midgut, and transform into epimastigotes. The epimastigotes reach the fly's salivary glands and continue multiplication by binary fission. The entire life cycle in the fly takes approximately 3 weeks. There are three different sub-species of T. brucei, which cause different variants of trypanosomiasis. T. brucei gambiense - Causes slow onset chronic trypanosomiasis in humans. Most common in central and western Africa, where humans are thought to be the primary reservoir.[1] T. brucei rhodesiense - Causes fast onset acute trypanosomiasis in humans. Most common in southern and eastern Africa, where game animals and livestock are thought to be the primary reservoir.[1] T. brucei brucei - Causes animal African trypanosomiasis, along with several other species of trypanosoma. T. b. brucei is not human infective due to its susceptibility to lysis by human apolipoprotein L1. However, as it shares many features with T. b. gambiense and T. b. rhodesiense (such as antigenic variation) it is used as a model for human infections in laboratory and animal studies. Trypanosoma equiperdum is a protozoan that causes Dourine, or covering sickness, in horses and other animals in the family equidae. The donkey shows no symptoms but is a carrier of this parasite. Unlike other species of Trypanosoma, this parasite can be sexually transmitted, also mother foal transmission has been documented and does not require a vector. Can be found in genital organs and secretions.
Notes made by kozi1989 email:kozi1989@yahoo.gr
166
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Trypanosoma cruzi is a species of parasitic euglenoid trypanosomes. This species causes the trypanosomiasis diseases in humans and animals in America. Transmission occurs when the reduviid bug deposits feces on the skin surface and subsequently bites ; the human host then scratches the bite area, which facilitates penetration of the infected feces. Human American trypanosomiasis, or Chagas disease, is a potentially fatal disease of humans. The parasite has two forms, a trypomastigote found in human blood and an amastigote found in tissues. The acute form usually goes unnoticed and may present as a localized swelling at the site of entry. The chronic form may develop 10 to 20 years after infection. This form affects internal organs (e.g., the heart, the esophagus, the colon, and the peripheral nervous system). Affected people may die from heart failure. Life cycle Trypanosoma cruzi life cycle starts in an animal reservoir . These reservoirs are usually mammals, wild or domestic, and include humans. A reduviid bug serves as the vector. While taking a blood meal, it ingests T. cruzi . In the reduviid bug, they go into the epimastigote stage. This makes it possible to reproduce. After reproducing through mitosis, the epimastigotes move onto the rectal cell wall. There, they become infectious. Infectious T. cruzi are called trypomastigotes. Then, while the reduviid bug is taking a blood meal from a human, it defecates. The trypomastigotes are in the feces. Trypomastigotes are capable of swimming into the host's cells using flagella, a characteristic swimming tail dominant in the Euglenoid class of protists. The trypomastigotes enter the human host through the bite wound or by crossing mucous membranes. The host cells contain macromolecules such as laminin, thrombospondin, heparan sulphate, and fibronectin that cover the surface of the host cells. These macromolecules are essential in the adhesion between parasite and host and the invasion process by the parasite of the host. The trypomastigotes must cross a network of proteins that line the exterior of the host cells in order to make contact and invade the host cells. The molecules and proteins on the cytoskeleton of the cell also bind to the surface of the parasite and initiate host invasion. When they enter a human cell, they become amastigotes. This is another reproductive stage. After reproducing through mitosis until a large amount of amastigotes are in a cell, pseudocysts are formed in infected cells. The amastigotes then turn back into trypomastigotes, and the cell bursts. The trypomastigotes swim along to either infect other cells or get sucked up by other reduviid bugs.
69. Subphylum Sarcodina. Entamoeba histolytica.
Entamoeba histolytica is an anaerobic parasitic protozoan, part of the genus Entamoeba. Predominantly infecting humans and other primates, E. histolytica is estimated to infect about 50 million people worldwide. Mammals such as dogs and cats can become infected transiently, but are not thought to contribute significantly to transmission .There are no intermediate or reservoir hosts. The active (trophozoite) stage exists only in the host and in fresh loose feces ; cysts
Notes made by kozi1989 email:kozi1989@yahoo.gr
167
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
survive outside the host in water, in soils, and on foods, especially under moist conditions on the latter. The cysts are readily killed by heat and by freezing temperatures, and survive for only a few months outside of the host. When cysts are swallowed they cause infections by excysting (releasing the trophozoite stage) in the digestive tract. The cyst is resistant to the gastric environment and passes into small intestine where it decysts. Having been ingested, the parasites excyst, releasing up trophozoites, the active stage in Entamoeba histolytica's life cycle. On emerging from the cyst, a trophozoite with four nuclei will divide itself three times, and then each nucleus will divide itself once, so that a total of eight trophozoites are released into the new host's digestive tract, resulting in invasive infections all over the host's body. The trophozoites migrate through the host's body to the large intestines, where they ingest red blood cells, bore into intestinal walls, damaging the host's tissue in the process, and multiply by binary fission. At this stage, some of the trophozoites will go on to bore into the mucosal walls of the large intestine and invade the host's bloodstream and major organs such as the liver, heart and lungs. Others will encyst themselves, encasing themselves in a capsule cyst with four nuclei and eventually being expelled from the host's body in the form of fecal matter, beginning the a new cycle. Infection can lead to amoebic dysentery or amoebic liver abscess.Symptoms can include fulminating dysentery, bloody diarrhea, weight loss, fatigue, abdominal pain, and amoeboma. The amoeba can actually 'bore' into the intestinal wall, causing lesions and intestinal symptoms, and it may reach the blood stream. From there, it can reach different vital organs of the human body, usually the liver, but sometimes the lungs, brain, spleen, etc. A common outcome of this invasion of tissues is a liver abscess, which can be fatal if untreated. Ingested red blood cells are sometimes seen in the amoeba cell cytoplasm.
70. Phylum Sporozoa (Apicomplexa). Plasmodium vivax. Plasmodium malariae.
Plasmodium falciparum. Plasmodium vivax is a protozoal parasite and a human pathogen. The most frequent and widely distributed cause of recurring (tertian) malaria, P. vivax is one of the four species of malarial parasite that commonly infect humans. It is less virulent than Plasmodium falciparum, which is the deadliest of the four, and is seldom fatal. P. vivax is carried by the female Anopheles mosquito, since it is the only sex of the species that bites. P. vivax can reproduce both asexually and sexually, depending on its life cycle stage. Asexual forms 1) Immature trophozoites (Ring or signet-ring shaped), about 1/3 of the diameter of a RBC. 2) Mature trophozoites: Very irregular and delicate (described as amoeboid); many pseudopodial processes seen. Presence of fine grains of brown pigment (malarial
Notes made by kozi1989 email:kozi1989@yahoo.gr
168
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
pigment) or hematin probably derived from the haemoglobin of the infected red blood cell. 3) Schizonts (also called meronts): As large as a normal red cell; thus the parasitized corpuscle becomes distended and larger than normal. There are about sixteen merozoites. Sexual forms Gametocytes: Round. The gametocytes of P. vivax are commonly found in the peripheral blood at about the end of the first week of parasitemia. Life cycle The incubation period for the infection usually ranges from ten to seventeen days and sometimes up to a year. Persistent liver stages allow relapse up to five years after elimination of red blood cell stages and clinical cure. The infection of Plasmodium vivax takes place in human when an infected female anopheles mosquito sucks blood from a healthy person. During feeding, the mosquito injects saliva to prevent blood clotting and, along with the saliva, thousands of sporozoites are inoculated into human blood; within a half-hour the sporozoites reach the liver. There they enter hepatic cells, transform into the tropozoite form and feed on hepatic cells, and reproduce asexually. This process gives rise to thousands of merozoites (plasmodium daughter cells) in the circulatory system and the liver. Liver stage The P. vivax sporozoite enters a hepatocyte and begins its exoerythrocytic schizogony stage. This is characterized by multiple rounds of nuclear division without cellular segmentation. After a certain number of nuclear divisions, the parasite cell will segment and merozoites are formed. There are situations where some of the sporozoites do not immediately start to grow and divide after entering the hepatocyte, but remain in a dormant, hypnozoite stage for weeks or months. The duration of latency is variable from one hypnozoite to another and the factors that will eventually trigger growth are not known; this explains how a single infection can be responsible for a series of waves of parasitaemia or "relapses".[16] Different strains of P. vivax have their own characteristic relapse pattern and timing.[17] The earlier stage is exo-erythrocytic generation. Erythrocytic cycle P. vivax preferentially penetrates young red blood cells (reticulocytes). In order to achieve this, merozoites have two proteins at their apical pole (PvRBP-1 and PvRBP-2). The parasite uses the Duffy blood group antigens (Fy6) to penetrate red blood cells. This antigen does not occur in the majority of humans in West Africa [phenotype Fy (a-b-)]. As a result P.
Notes made by kozi1989 email:kozi1989@yahoo.gr
169
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
vivax occurs less frequently in West Africa. The parasitised red blood cell is up to twice as large as a normal red cell and Schffner's dots (also known as Schffner's stippling or Schffner's granules) is seen on the infected cell's surface, the spotted appearance of which varies in color from light pink, to red, to red-yellow, as coloured with Romanovsky stains. The parasite within it is often wildly irregular in shape (described as "amoeboid"). Schizonts of P. vivax have up to twenty merozoites within them. It is rare to see cells with more than one parasite within them. Merozoites will only attach to immature blood cell (reticulocytes) and therefore it is unusual to see more than 3% of all circulating erythrocytes parasitised. Sexual stage The sexual stage includes following processes by which P. vivax reproduces sexually: 1) 2) 3) 4) 5) 6) Transfer to mosquito Gametogenesis Microgametes - Macrogametes Fertilization Ookinite Oocyst Sporogony
Mosquito stage The life cycle in mosquitoes include: 1. Formation of gametes 2. Fertilization 3. Sporogony Plasmodium malariae is a parasitic protozoa that causes malaria in humans. It is closely related to Plasmodium falciparum and Plasmodium vivax which are responsible for most malarial infection. While found worldwide, it is a so-called "benign malaria" and is not nearly as dangerous as that produced by P. falciparum or P. vivax. P. malariae causes fevers that recur at approximately three-day intervals (a quartan fever), longer than the two-day (tertian) intervals of the other malarial parasites, hence its alternate name quartan malaria. The ring stages that are formed by the invasion of merozoites released by rupturing liver stage schizonts are the first stages that appear in the blood .The ring stages grow slowly but soon fill one-fourth to one-third of the parasitized cell. Pigment increases rapidly and the half-grown parasite may have from 30 to 50 jet-black granules. The parasite changes various shapes as it grows and stretches across the host cell to form the band form. P. malariae can be maintained at very low infection rates among a sparse and mobile population because unlike the other Plasmodium parasites, it can remain in a human host for an extended period of time and still remain infectious to mosquitoes.[6] The vector of transmission of the parasite is the female Anopheles mosquito, but many different species have been shown to transmit the parasite at least experimentally.[1] However, there are no animal reservoirs[clarification needed] for Plasmodium malariae.
Notes made by kozi1989 email:kozi1989@yahoo.gr
170
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Life cycle P. malariae is the only human malaria parasite that causes fevers that recur at approximately three-day intervals (therefore occurring evey fourth day, a quartan fever), longer than the two-day (tertian) intervals of the other malarial parasites. Liver stage In this stage, many thousands of merozoites are produced in each schizont. As the merozoites are released, they invade erythrocytes and initiate the erythrocytic cycle, where the parasite digests hemoglobin to obtain amino acids for protein synthesis. Erythrocytic cycle The total length of the intraerythrocytic development is roughly 72 hours for P. malariae. At the schizont stage, after schizogonic division, there are roughly 6 8 parasite cells in the erythrocyte. Following the erythrocytic cycle, which lasts for seventy two hours on average, six to fourteen merozoites are released to reinvade other erythrocytes.[1] Finally, some of the merozoites develop into either micro- or macrogametocytes.[1] The two types of gametocytes are taken into the mosquito during feeding and the cycle is repeated. There are no animal reservoirs for P. malariae. Mosquito stage Similar to the other human-infecting Plasmodium parasites, Plasmodium malariae has distinct developmental cycles in the Anopheles mosquito and in the human host.[1] The mosquito serves as the definitive host and the human host is the intermediate.[1] When the Anopheles mosquito takes a blood meal from an infected individual, gametocytes are ingested from the infected person.[1] A process known as exflagellation of the microgametocyte soon ensues and up to eight mobile microgametes are formed.[1] Sexual stage Following fertilization of the macrogamete, a mobile ookinete is formed, which penetrates the peritropic membrane surrounding the blood meal and travels to the outer wall of the mid-gut of the mosquito.[1] The oocyst then develops under the basal membrane and after a period of two to three weeks a variable amount of sporozoites are produced within each oocyst.[1] The number of sporozoites that are produced varies with temperature and can range from anywhere between many hundreds to a few thousand.[1] Eventually, the oocyst ruptures and the sporozoites are released into the hemocoel of the mosquito. The sporozoites are then carried by the circulation of the hemolymph to the salivary glands, where they become concentrated in the acinal cells.[1] A small number of sporozoites are introduced into the salivary duct and injected into the venules of the bitten human.[1] This initiates the cycle in the human liver.
Notes made by kozi1989 email:kozi1989@yahoo.gr
171
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Plasmodium falciparum is a protozoan parasite, one of the species of Plasmodium that cause malaria in humans. It is transmitted by the female Anopheles mosquito. P. falciparum is the most dangerous of these infections[1] as P. falciparum (or malignant) malaria has the highest rates of complications and mortality. Life cycle The life cycle of all Plasmodium species is complex. Infection in humans begins with the bite of an infected female Anopheline mosquito. Sporozoites released from the salivary glands of the mosquito enter the bloodstream during feeding quickly invade liver cells (hepatocytes). Sporozoites are cleared from the circulation within 30 minutes. During the next 14 days in the case of P. falciparum, the liver-stage parasites differentiate and undergo asexual multiplication resulting in tens of thousands of merozoites which burst from the hepatoctye. Individual merozoites invade red blood cells (erythrocytes) and undergo an additional round of multiplication producing 12-16 merozoites within a schizont. The length of this erythrocytic stage of the parasite life cycle depends on the parasite species: irregular cycle for P. falciparum, 48 hours for P. vivax, and P. ovale and 72 hours for P. malariae. The clinical manifestations of malaria, fever and chills, are associated with the synchronous rupture of the infected erythrocyte. The released merozoites go on to invade additional erythrocytes. Not all of the merozoites divide into schizonts, some differentiate into sexual forms, male and female gametocytes. These gametocytes are taken up by a female anophylean mosquito during a blood meal. Within the mosquito midgut, the male gametocyte undergoes a rapid nuclear division, producing 8 flagellated microgametes which fertilize the female macrogamete. The resulting ookinete traverses the mosquito gut wall and encysts on the exterior of the gut wall as a oocyst. Soon the oocyst ruptures, releasing hundreds of sporozoites into the mosquito body cavity where they eventually migrate to the mosquito salivary gland.
71. Phylum Sporozoa (Apicomplexa). Morphological differences in the erythrocyte
schizogonia of the genus Plasmodium. Types of malaria After entering the erythrocyte, the merozoite lose one of their membranes, the apical rings, conoid and the rhopteries. Phagotropy commences and both smooth and granular endoplasmic reticulum becomes prominent. The nucleus may become lobulated. Within the erythrocytes the merozoite grow first to a ring-shaped form and then to a larger trophozoite form. In the schizont stage, the parasite divides several times to produce new merozoites, which leave the red blood cells and travel within the bloodstream to invade new red blood cells. The parasite feeds by ingesting haemoglobin and other materials from red blood cells and serum. The feeding process damages the erythrocytes The infection is initiated when sporozoites are injected with the saliva of a feeding mosquito. Sporozoites are carried by the circulatory system to the liver and invade hepatocytes (1). Theintracellular parasite undergoes an asexual replication known as exoerythrocytic schizogony within the hepatocyte (2-4). Exoerythrocytic schizogony
Notes made by kozi1989 email:kozi1989@yahoo.gr
172
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
culminates in the production ofmerozoites which are released into the bloodstream (5). A proportion of the liver-stage parasitesfrom P. vivax and P. ovale go through a dormant period (not shown) instead of immediatelyundergoing asexual replication (i.e., stay temporarily at step 2). These hypnozoites willreactivate several weeks to months (or years) after the primary infection and are responsible forrelapses. Merozoites invade erythrocytes (6) and undergo a trophic period in which the parasite enlarges. The early trophozoite is often referred to as 'ring form' because of its morphology.Trophozoite enlargement is accompanied by an active metabolism including the ingestion ofhost cytoplasm and the proteolysis of hemoglobin into amino acids. The end of the trophic periodis manifested by multiple rounds of nuclear division without cytokinesis resulting is a schizont. Merozoites bud from the mature schizont, also called a segmenter (10), and the merozoitesare released following rupture of the infected erythrocyte (11). Invasion of erythrocytesreinitiates another round of the blood-stage replicative cycle (6-11). P. vivax preferentially penetrates young red blood cells (reticulocytes). In order to achieve this, merozoites have two proteins at their apical pole (PvRBP-1 and PvRBP-2). The parasite uses the Duffy blood group antigens (Fy6) to penetrate red blood cells. This antigen does not occur in the majority of humans in West Africa [phenotype Fy (a-b-)]. As a result P. vivax occurs less frequently in West Africa.[19] The total length of the intraerythrocytic development is roughly 72 hours for P. malariae. At the schizont stage, after schizogonic division, there are roughly 68 parasite cells in the erythrocyte. Following the erythrocytic cycle, which lasts for seventy two hours on average, six to fourteen merozoites are released to reinvade other erythrocytes.[1] Finally, some of the merozoites develop into either micro- or macrogametocytes.[1] The two types of gametocytes are taken into the mosquito during feeding and the cycle is repeated.[1] There are no animal reservoirs for P. malariae. There are four common kinds of malaria that are seen in people. The 4 common kinds of malaria that are seen include the Plasmodium Vivax type of Malaria. This is commonly known as the P.v. type. The other type of Malaria is the Plasmodium Malariae. This type is also known as P.m. The plasmodium ovale and the Plasmodium falciparum are the two other kinds of malaria that are also commonly seen. These are known as P.o and P.f respectively. The type of Malaria is mainly classified on the basis of the organism that causes the disease. For example the Plasmodium vivax type of Malaria is caused by the P. vivax organism. Though the types of Malaria may vary depending on the organism that causes the disease, there is another difference between the different types of Malaria. This difference is based on the various signs and symptoms that are associated with the Malaria. The Plasmodium Falciparum is the kind of malaria that can be fatal in many instances. This is because this kind of malaria causes a lot of severe complications
72. Phylum Sporozoa (Apicomplexa). Toxoplasma gondii.
Notes made by kozi1989 email:kozi1989@yahoo.gr
173
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Toxoplasma gondii is a species of parasitic protozoa in the genus Toxoplasma. The definitive host of T. gondii is the cat, but the parasite can be carried by many warmblooded animals (birds or mammals, including humans). Toxoplasmosis, the disease of which T. gondii is the causative agent, is usually minor and self-limiting but can have serious or even fatal effects on a fetus whose mother first contracts the disease during pregnancy or on an immunocompromised human or cat. Life cycle The life cycle of T. gondii has two phases. The sexual part of the life cycle (coccidia like) takes place only in cats, both domestic and wild (family Felidae), which makes cats the parasite's primary host. The second phase, the asexual part of the life cycle, can take place in other warmblooded animals, including cats, mice, humans, and birds. The hosts in which asexual reproduction takes place is called the intermediate host. Rodents are the typical intermediate host. T. gondii constructing daughter scaffolds within the mother cell. In both kinds of hosts, the Toxoplasma parasite invades cells and forms a space called a vacuole . Inside this specialized vacuole, called a parasitophorous vacuole, the parasite forms bradyzoites, which are the slowly replicating versions of the parasite.[3] The vacuoles containing the reproductive bradyzoites form cysts mainly in the tissues of the muscles and brain. Since the parasites are inside of cells, they are safe from the host's immune system, which does not respond to the cysts. Toxoplasma's resistance to antibiotics varies, but the cysts are very difficult to eradicate entirely. Inside the vacuoles, T. gondii replicates itself (by endodyogeny) until the infected cell fills with parasites and bursts, releasing tachyzoites, the motile, asexually reproducing form of the parasite. Unlike the bradyzoites, the free tachyzoites are usually efficiently cleared by the host's immune system, although some of them manage to infect cells and form bradyzoites, thus maintaining the infection. Tissue cysts are ingested by a cat (e.g., by feeding on an infected mouse). The cysts survive passage through the stomach of the cat and the parasites infect epithelial cells of the small intestine where they undergo sexual reproduction and oocyst formation. Oocysts are shed with the feces. Animals and humans that ingest oocysts (e.g., by eating unwashed vegetables) or tissue cysts in improperly cooked meat become infected. The parasite enters macrophages in the intestinal lining and is distributed via the blood stream throughout the body. Similar to the mechanism used in many viruses, Toxoplasma is able to dysregulate hosts cell cycle by holding cell division before mitosis (the G2/M border).[4] This dysregulation of the hosts cell cycle is caused by a heat-sensitive secretion (with a molecular mass larger than 10 kDa).[5] Infected cells secrete the factor which inhibits the cell cycle of neighboring cells. The reason for Toxoplasmas dysregulation is unknown, but studies have shown that infection is preferential to host cells in the S-phase and host cell structures with which Toxoplasma interacts may not be accessible during other stages of the cell cycle.
Notes made by kozi1989 email:kozi1989@yahoo.gr
174
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Acute stage Toxoplasma infections can be asymptomatic, but often give flu-like symptoms in the early acute stages, and like flu can become, in very rare cases, fatal. The acute stage fades in a few days to months, leading to the latent stage. Latent infection is normally asymptomatic; however, in the case of immunocompromised patients (such as those infected with HIV or transplant recipients on immunosuppressive therapy), toxoplasmosis can develop. The most notable manifestation of toxoplasmosis in immunocompromised patients is toxoplasmic encephalitis, which can be deadly. If infection with T. gondii occurs for the first time during pregnancy, the parasite can cross the placenta, possibly leading to hydrocephalus or microcephaly, intracranial calcification, and chorioretinitis, with the possibility of spontaneous abortion (miscarriage) or intrauterine death.
73. Phylum Ciliophora. Class Ciliata. Balantidium coli.
Balantidium coli is a parasitic species of ciliate protozoan that causes the disease Balantidiasis. It is the only member of the ciliate phylum known to be pathogenic to humans. Balantidium coli as seen in a wet mount of a stool specimen. The organism is surrounded by cilia.Balantidium coli has two developmental stages, a trophozoite stage and a cyst stage. In trophozoites, the two nuclei are visible. The macronucleus is long and sausage-shaped, and the spherical micronucleus is nested next to it, often hidden by the macronucleus. The opening, known as the peristome, at the pointed anterior end leads to the cytosome, or the mouth. Cysts are smaller than trophozoites and are round and have a tough, heavy cyst wall made of one or two layers. Usually only the macronucleus and sometimes cilia and contractile vacuoles are visible in the cyst. Living trophozoites and cysts are yellowish or greenish in color. Balantidium is the only ciliated protozoan known to infect humans . Balantidiasis is a zoonotic disease and is acquired by humans via the fecal-oral route from the normal host, the pig, where it is asymptomatic. Contaminated water is the most common mechanism of transmission. Balantidium coli lives in the cecum and colon of humans, pigs, rats and other mammals. It is not readily transmissible from one species of host to another because it requires a period of time to adjust to the symbiotic flora of the new host. Once it has adapted to a host species, the protozoan can become a serious pathogen, especially in humans. Trophozoites multiply and encyst due to the dehydration of feces. Infection occurs when the cysts are ingested, usually through contaminated food or water. Balantidium infection in immunocompetent individuals is not unheard of, but it rarely causes a serious disease of the gastrointestinal tract. It can thrive in the gastrointestinal tract as long as there is a balance between the protozoan and the host without causing dysenteric symptoms. Infection most likely occurs in people with malnutrition due to the low stomach acidity or people with immune compromised systems. In acute disease, explosive diarrhea may occur as often as every twenty minutes.
Notes made by kozi1989 email:kozi1989@yahoo.gr
175
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Perforation of the colon may also occur in acute infections which can lead to life-threatening situations. Life cycle when a host ingests a cyst, which usually happens during the consumption of contaminated water or food. Once the cyst is ingested, it passes through the hosts digestive system. While the cyst receives some protection from degradation by the acidic environment of the stomach through the use of its outer wall, it is likely to be destroyed at a pH lower than 5, allowing it to survive easier in the stomachs of malnourished individuals who have less stomach acid. Once the cyst reaches the small intestine, trophozoites are produced. The trophozoites then colonize the large intestine, where they live in the lumen and feed on the intestinal flora. Some trophozoites invade the wall of the colon using proteolytic enzymes and multiply, and some of them return to the lumen. In the lumen trophozoites may disintegrate or undergo encystation. Encystation is triggered by dehydration of the intestinal contents and usually occurs in the distal large intestine, but may also occur outside of the host in feces. Now in its mature cyst form, cysts are released into the environment where they can go on to infect a new host.
74. Transition of Metazoa. Phylum Coelenterata (Cnidaria). Genus Hyadra. Genus
Obelia. Coelenterata is an obsolete long term encompassing two animal phyla, the Ctenophora (comb jellies) and the Cnidaria (coral animals, true jellies, sea anemones, sea pens, and their allies). The name comes from the Greek "koilos" ("full bellied"), referring to the hollow body cavity common to these two phyla. They have very simple tissue organization, with only two layers of cells, external and internal. Cnidaria( /nadri/ with a silent c) is a phylum containing over 9,000 species of animals found exclusively in aquatic and mostly marine environments. Their distinguishing feature is cnidocytes, specialized cells that they use mainly for capturing prey. Their bodies consist of mesoglea, a non-living jelly-like substance, sandwiched between two layers of epithelium that are mostly one cell thick. They have two basic body forms: swimming medusae and sessile polyps, both of which are radially symmetrical with mouths surrounded by tentacles that bear cnidocytes. Both forms have a single orifice and body cavity that are used for digestion and respiration . Many cnidarian species produce colonies that are single organisms composed of medusa-like or polyp-like zooids, or both. Cnidarians' activities are coordinated by a decentralized nerve net and simple receptors. Several free-swimming Cubozoa and Scyphozoa possess balancesensing statocysts, and some have simple eyes. Not all cnidarians reproduce sexually. Many
Notes made by kozi1989 email:kozi1989@yahoo.gr
176
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
have complex lifecycles with asexual polyp stages and sexual medusae, but some omit either the polyp or the medusa stage. Hydra (English pronunciation: /hadr/) is a genus of simple fresh-water animal possessing radial symmetry. Hydras are predatory animals belonging to the phylum Cnidaria and the class Hydrozoa. They can be found in most unpolluted fresh-water ponds, lakes, and streams in the temperate and tropical regions and can be found by gently sweeping a collecting net through weedy areas. They are usually a few millimetres long and are best studied with a microscope. Biologists are especially interested in hydras due to their regenerative ability; and that they appear not to age or die of old age. However, presently, there is no scientific consensus on whether Hydra undergo senescence, as discussed below. Morphology Hydra has a tubular body secured by a simple adhesive foot called the basal disc. Gland cells in the basal disc secrete a sticky fluid that allows for its adhesive properties. At the free end of the body is a mouth opening surrounded by one to twelve thin, mobile tentacles. Each tentacle, or cnida (plural: cnidae), is clothed with highly specialised stinging cells called cnidocytes. Cnidocytes contain specialized structures called nematocysts, which look like miniature light bulbs with a coiled thread inside. At the narrow outer edge of the cnidocyte is a short trigger hair called a cnidocil. Upon contact with prey, the contents of the nematocyst are explosively discharged, firing a dart-like thread containing neurotoxins into whatever triggered the release which can paralyse the prey, especially if many hundreds of nematocysts are fired. Hydra has two main body layers separated by mesoglea, a gel-like substance. The outer layer is the epidermis, and the inner layer is called the gastrodermis. The cells making up these two body layers are relatively simple. Hydramacin is a bactericide recently discovered in Hydra; it protects the outer layer against infection. The nervous system of Hydra is a nerve net, which is structurally simple compared to mammalian nervous systems. Hydra does not have a recognizable brain or true muscles. Nerve nets connect sensory photoreceptors and touch-sensitive nerve cells located in the body wall and tentacles.Respiration and excretion occurs by diffusion through the epidermis. Motion and locomotion If a Hydra is alarmed or attacked, the tentacles can be retracted to small buds, and the body column itself can be retracted to a small gelatinous sphere. Hydras generally react in the same way regardless of the direction of the stimulus, and this may be due to the simplicity of the nerve net. Hydras are generally sedentary or sessile, but do occasionally move quite readily, especially when hunting. They do this by bending over and attaching themselves to the substrate with the mouth and tentacles and then release the foot, which provides the usual attachment, this process is called looping. The body then bends over and makes a new place
Notes made by kozi1989 email:kozi1989@yahoo.gr
177
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
of attachment with the foot. By this process of "inch-worming" or "somersaulting", a hydra can move several inches (c. 100 mm) in a day. Hydras may also move by amoeboid motion of their bases or by simply detaching from the substrate and floating away in the current. Reproduction and life cycle When food is plentiful, many hydras reproduce asexually by producing buds in the body wall, which grow to be miniature adults and simply break away when they are mature. When conditions are harsh, often before winter or in poor feeding conditions, sexual reproduction occurs in some hydras. Swellings in the body wall develop into either a simple ovary or testes. The testes release free-swimming gametes into the water, and these can fertilize the egg in the ovary of another individual. The fertilized eggs secrete a tough outer coating, and, as the adult dies, these resting eggs fall to the bottom of the lake or pond to await better conditions, whereupon they hatch into miniature adults. Hydras are hermaphrodites and may produce both testes and an ovary at the same time. Many members of the Hydrozoa go through a body change from a polyp to an adult form called a medusa. However, all hydras, despite being hydrozoans, remain as polyps throughout their lives. Obelia is a genus in the class Hydrozoa, which consists of mainly marine and some freshwater animal species and have both the polyp and medusa stages in their life cycle. The genus belongs to the phylum Cnidaria, which are all aquatic and mainly marine organisms that are relatively simple in structure. The Obelia are naturally found underwater in the ocean throughout the world . They are marine colonial hydrozoans that are found and hard substrata in the subtidal zones. They are usually found no deeper than 200 metres (660 ft) from the water's surface, growing in intertidal rockpools and at the extreme low water of spring tides. Structure Through its life cycle, Obelia takes two forms: polyp and medusa. They are diploblastic, with two true tissue layers an epidermis and a gastrodermis, with a jelly-like mesoglea filling the area between the two true tissue layers . They carry a nerve net with no brain or ganglia. A gastrovascular cavity is present where the digestion starts and later becomes intracellular. They have incomplete digestive tracts where the food enters, is digested, and expelled through the same opening. During the polyp stage, the mouth is situated at the top of the body, surrounded by tentacles, whereas during the medusa stage, the mouth is situated at the distal end of the main body structure. Four gonads lie in this main body structure, or manubrium. When food is taken in through the mouth, it enters the manubrium. The food is then distributed through a canal system, consisting of four radial canals and an outer ring. Defense and the capture of prey are helped by unique stinging cells called cnidocytes that contain nematocysts, which are triggered by the cnidocil. Life cycle
Notes made by kozi1989 email:kozi1989@yahoo.gr
178
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The polyp colony reproduces asexually. During this stage of life, Obelia are confined to substrate surfaces. On this mature colony there are individual hydranths called gastrozooids, which can be found expanded or contracted, to aid in the growth of this organism by feeding; the reproductive polyp gonozooids has medusa buds. Other hydranths are specialized for defense. The main stalky body of the colony is composed of a coenosarc, which is covered by a protective perisarc. The next generation of the life cycle begins when the medusae are released from these gonozooids, producing free swimming only male medusae velum with gonads, a mouth, and tentacles. The physical appearance of the male and female medusae velum, including their gonads, are indistinguishable, and the sex can only be determined by observing the inside of the gonads, which will either contain sperm or eggs. The medusae reproduce sexually, releasing sperm and eggs that fertilize to form a zygote, which later morphs into a blastula, then a ciliated swimming larva called a planula. The planulae live free-swimming for a while but eventually attach themselves to some solid surface, where they begin their reproductive phase of life. Once attached to a substrate, a planula quickly develops into one feeding polyp. As the polyp grows, it begins developing branches of other feeding individuals, thus forming a new generation of polyps by asexual budding.
75. Phylum Plathelminthes (Platyhelminthes). Class Trematoda. Fasciola hepatica
The flatworms, known in scientific literature as Platyhelminthes or Plathelminthes (from the Greek , platy, meaning "flat" and (root: -), helminth-, meaning worm)[2] are a phylum of relatively simple bilaterian, unsegmented, soft-bodied invertebrate animals. Unlike other bilaterians, they have no body cavity, and no specialized circulatory and respiratory organs, which restricts them to flattened shapes that allow oxygen and nutrients to pass through their bodies by diffusion. Over half of all known flatworm species are parasitic, and some do enormous harm to humans and their livestock. Schistosomiasis, caused by one genus of trematodes, is the second most devastating of all human diseases caused by parasites, surpassed only by malaria. Neurocysticercosis, which arises when larvae of the pork tapeworm Taenia solium penetrate the central nervous system, is the major cause of acquired epilepsy worldwide. The threat of platyhelminth parasites to humans in developed countries is rising because of organic farming, the popularity of raw or lightly cooked foods, and imports of food from high-risk areas. In less developed countries, people often cannot afford the fuel required to cook food thoroughly, and poorly designed water-supply and irrigation projects increase the dangers presented by poor sanitation and unhygienic farming. Trematoda is a class within the phylum Platyhelminthes that contains two groups of parasitic flatworms, commonly referred to as "flukes". Trematodes are flattened oval or worm-like animals, usually no more than a few centimetres in length, although species as small as 1 millimetre (0.039 in) and as large as 7
Notes made by kozi1989 email:kozi1989@yahoo.gr
179
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
metres (23 ft) are known. Their most distinctive external feature is the presence of two suckers, one close to the mouth, and the other on the underside of the animal. The body surface of trematodes comprises a tough syncitial tegument, which helps protect against digestive enzymes in those species that inhabit the gut of larger animals. It is also the surface of gas exchange; there are no respiratory organs. The mouth is located at the forward end of the animal, and opens into a muscular, pumping pharynx. The pharynx connects, via a short oesophagus, to one or two blindending caeca, which occupy most of the length of the body. In some species, the caeca are themselves branched. As in other flatworms, there is no anus, and waste material must be egested through the mouth. Although the excretion of nitrogenous waste occurs mostly through the tegument, trematodes do possess an excretory system, which is instead mainly concerned with osmoregulation. This consists of two or more protonephridia, with those on each side of the body opening into a collecting duct. The two collecting ducts typically meet up at a single bladder, opening to the exterior through one or two pores near the posterior end of the animal. The brain consists of a pair of ganglia in the head region, from which two or three pairs of nerve cords run down the length of the body. The nerve cords running along the ventral surface are always the largest, while the dorsal cords are present only in the Aspidogastrea. Trematodes generally lack any specialised sense organs, although some ectoparasitic species do possess one or two pairs of simple ocelli.[3] Reproductive system Most trematodes are simultaneous hermaphrodites, having both male and female organs. There are usually two testes, with sperm ducts that join together on the underside of the front half of the animal. This final part of the male system varies considerably in structure between species, but may include sperm storage sacs and accessory glands, in addition to the copulatory organ, which is either eversible, and termed a cirrus, or noneversible, and termed a penis. There is usually only a single ovary, which is connected, via a pair of ducts to a number of vitelline glands on either side of the body, that produce yolk cells. Eggs pass from the ovary into a glandular receptacle called the ootype or Mehlis' gland, where fertilization occurs. This opens into an elongated uterus that opens to the exterior close to the male opening. The ovary is often also associated with a storage sac for sperm, and a copulatory duct termed Laurer's canal.[3] Life cycles Almost all trematodes infect mollusks as the first host in the life cycle, and most have a complex life cycle involving other hosts. Most trematodes are monoecious and alternately reproduce sexually and asexually. The two main exceptions to this are the Aspidogastrea, which have no asexual reproduction, and the schistosomes, which are
Notes made by kozi1989 email:kozi1989@yahoo.gr
180
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
dioecious. In the definitive host, in which sexual reproduction occurs, eggs are commonly shed along with host feces. Eggs shed in water release free-swimming larval forms that are infective to the intermediate host, in which asexual reproduction occurs. A species that exemplifies the remarkable life history of the trematodes is the bird fluke, Leucochloridium paradoxum. The definitive hosts, in which the parasite multiplies, are various woodland birds, while the hosts in which the parasite grows (intermediate host) are various species of snail. The adult parasite in the bird's gut produces eggs and these eventually end up on the ground in the bird's faeces. Some very fortunate eggs get swallowed by a snail and here they hatch into tiny, transparent larva (miracidium). These larvae grow and take on a sac-like appearance. This stage is known as the sporocyst and it forms a central body in the snail's digestive gland that extends into a brood sac in the snail's head, muscular foot and eye-stalks. It is in the central body of the sporocyst where the parasite replicates itself, producing lots of tiny embryos (redia). These embryos move to the brood sac and mature into cercaria. Fasciola hepatica, also known as the common liver fluke or sheep liver fluke, is a parasitic flatworm of the class Trematoda, phylum Platyhelminthes that infects the livers of various mammals, including humans. The disease caused by the fluke is called fascioliasis (also known as fasciolosis). F. hepatica is distributed worldwide, and causes great economic losses in sheep and cattle. Life cycle To complete its life cycle, F. hepatica requires a freshwater snail as an intermediate host, such as Galba truncatula, in which the parasite can reproduce asexually. From the snail, minute cercariae emerge and swim through pools of water in pasture, and encyst as metacercariae on near-by vegetation. From here, the metacercariae are ingested by the ruminant, or in some cases, by humans eating uncooked foods such as watercress. Contact with low pH in the stomach causes the early immature juvenile to begin the process of excystment. In the duodenum, the parasite breaks free of the metacercariae and burrows through the intestinal lining into the peritoneal cavity . The newly excysted juvenile does not feed at this stage, but once it finds the liver parenchyma after a period of days, feeding will start. This immature stage in the liver tissue is the pathogenic stage, causing anaemia and clinical signs sometimes observed in infected animals. The parasite browses on liver tissue for a period of up to six weeks, and eventually finds its way to the bile duct, where it matures into an adult and begins to produce eggs. Up to 25,000 eggs per day per fluke can be produced, and in a light infection, up to 500,000 eggs per day can be deposited onto pasture by a single sheep.
76. Class Trematoda. Dicrocoelium lanceatum (Dicrocoelium dendriticum).
Notes made by kozi1989 email:kozi1989@yahoo.gr
181
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The Lancet liver fluke (Dicrocoelium dendriticum) is a parasite fluke that tends to live in cattle or other grazing mammals. Dicrocoelium dendriticum has a similar morphology to Clonorchis sinensis, the Chinese liver fluke. Dicrocoelium dendriticum is distinguished by lobed testes in the anterior of the body, as opposed to Clonorchis sinensis whose testes are located in the posterior. They both are flat and have a characteristic taper at the anterior and posterior ends. The anterior is distinguished by an oral sucker at the point, an acetabulum and the testes. The posterior is where the uterus lies. In the parasites mid-section lie the vitelline glands that are involved in egg formation. Life cycle Dicrocoelium dendriticum spends its adult life inside the liver of its host. After mating, the eggs are excreted in the feces. The first intermediate host, the terrestrial snail (Cochlicopa lubrica in the United States), eats the feces, and becomes infected by the larval parasites . The larvae (or miracidium) drill through the wall of the gut and settle in its digestive tract , where they develop into a juvenile stage. The snail tries to defend itself by walling the parasites off in cysts, which it then excretes and leaves behind in the grass. The second intermediate host, an ant (Formica fusca in the United States[5]), uses the trail of snail slime as a source of moisture. The ant then swallows a cyst loaded with hundreds of juvenile lancet flukes. The parasites enter the gut and then drift through its body. Most of the cercariae encyst in the haemocoel of the ant and mature into metacercariae, but one moves to the sub-esophageal ganglion (a cluster of nerve cells underneath the esophagus). There, the fluke takes control of the ant's actions by manipulating these nerves.[6] As evening approaches and the air cools, the infected ant is drawn away from other members of the colony and upward to the top of a blade of grass. Once there, it clamps its mandibles onto the top of the blade and stays there until dawn. Afterward, it goes back to its normal activity at the ant colony. If the host ant were to be subjected to the heat of the direct sun, it would die along with the parasite. Night after night, the ant goes back to the top of a blade of grass until a grazing animal comes along and eats the blade, ingesting the ant along with it, thus putting lancet flukes back inside their host. They live out their adult lives inside the animal, reproducing so that the cycle begins again.
77. Class Trematoda. Opisthorchis felineus. Genus Schistosoma and its species.
Opisthorchis felineus, or cat liver fluke is a trematode parasite that infects the liver in mammals. It was first discovered in 1884 in a cat's liver by Sebastiano Rivolta of Italy. In 1891, Russian scientist K.N. Vinogradov found it in a human, and named the parasite a "Siberian liver fluke". In the 1930s, helminthologist Hans Vogel of Hamburg published an article describing the life cycle of Opisthorchis felineus. The first "intermediate hosts" of the parasite are freshwater snails Bithynia inflata
Notes made by kozi1989 email:kozi1989@yahoo.gr
182
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
(synonym: Codiella inflata),[3] Bithynia troschelii[3] and Bithynia leachii.[3] The second "intermediate hosts" are freshwater fish , followed by the final host, which are fish-eating mammals such as felines and humans. It is estimated that 1.5 million people in Russia are infected with the parasite. Inhabitants of Siberia acquire the infection by consuming raw, slightly salted and frozen fish. Opisthorchiasis, the disease caused by Opisthorchis felineus, ranges in severity from asymptomatic infection to severe illness. Patient outcome is dependent on early detection and treatment. Human cases of opisthorchiasis may affect the liver, pancreas, and gall bladder . If not treated in the early stages, opisthorchiasis may cause cirrhosis of the liver and increased risk of liver cancer, but may be asymptomatic in children. Two weeks after flukes enter the body, the parasites infect the biliary tract. Symptoms of infection include fever, general malaise, skin rash, and gastrointestinal disturbances. Severe anemia and liver damage may also incapacitate the infected person for 1-2 months. A genus of trematodes, Schistosoma, commonly known as blood-flukes and bilharzia, includes flatworms which are responsible for a highly significant parasitic infection of humans by causing the disease schistosomiasis, and are considered by the World Health Organization as the second most important parasitic disease, next only to malaria, with hundreds of millions infected worldwide. Adult worms parasitize mesenteric blood vessels. They are unique among trematodes or any other flatworms in that they are dioecious with distinct sexual dimorphism between male and female. Eggs are passed through urine or feces to fresh water, where larva must pass though an intermediate snail host, before a different larval stage of the parasite emerges that can infect a new mammalian host by directly penetrating the skin. The genus Schistosoma as currently defined is paraphyletic so revisions are likely. Currently twenty one species are recognised within this genus. The genus has been divided into four groups - indicum, japonicum, haematobium and mansoni. The affinities of the remaining three species are still being clarified. Twelve species are found in Africa. Eleven of these are divided into two groups those with a lateral spine on the egg (mansoni group) and those with a terminal spine (haematobium group). The four mansoni group species are: S. edwardiense, S. hippotami, S. mansoni and S. rodhaini. The eight haematobium group species are: S. bovis, S. curassoni, S. intercalatum, S. guineensis, S. haematobium, S. leiperi, S. margrebowiei and S. matthei
78. Class Cestoda. Taenia solium.
Notes made by kozi1989 email:kozi1989@yahoo.gr
183
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Taenia solium, also called the pork tapeworm, is a cyclophyllid cestode in the family Taeniidae. It infects pigs and humans in Asia, Africa, South America, parts of Southern Europe and pockets of North America. In the larval stage, it causes cysticercosis in humans. Like all cyclophyllid cestodes, T. solium has four suckers on its scolex ("head"). T. solium also has two rows of hooks.T. solium is normally 2 m to 3 m in length, but can become very large, over 50 m long in some situations. T. solium has a very similar life cycle to Taenia saginata. Cysticerci have three morphologically distinct types. The common one is the ordinary "cellulose" cysticercus which has a fluid filled bladder that is 0.5 cm to 1.5 cm in length and an invaginated scolex. The intermediate form has a scolex while the "racemose" has no evident scolex but are believed to be larger and much more dangerous. They are 20 cm in length and have 60 ml of fluid and 13% of patients might have all three types in the brain. Humans are usually infected through eating infected pork, fostering adult tapeworms in the intestine, and passing eggs through feces, but autoinfection is also possible. In that case, a cysticercus (a larva sometimes called a "bladder worm") develops in the human and the human acts like an intermediate host . This happens if eggs get to the stomach, usually as a result of contaminated hands, but also due to retroperistalsis. Cysticerci often occur in the central nervous system, which can cause major neurological problems like hydrocephalus, paraplegy, meningitis, convulsions and even death. The condition of having cysticerci in one's body is called cysticercosis. Eggs can be diagnosed only to the family level, but if a proglottid's uterus is stained with India ink, the number of visible uterine branches can help identify the species: unlike the Taenia saginata uteri, T. solium uteri have only five to ten uterine branches on each side. Life cycle This infection is caused by ingestion of eggs shed in the feces of a human tapeworm carrier. Pigs and humans become infected by ingesting eggs or gravid proglottids. Humans are infected either by ingestion of food contaminated with feces containing eggs, or by autoinfection. In the latter case, a human infected with adult T. solium can ingest eggs produced by that tapeworm, either through fecal contamination or, possibly, from proglottids carried into the stomach by reverse peristalsis. Once eggs are ingested, oncospheres hatch in the intestine, invade the intestinal wall, and migrate to striated muscles, as well as the brain, liver, and other tissues, where they develop into cysticerci. In humans, cysts can cause serious sequelae if they localize in the brain, resulting in neurocysticercosis. The parasite life cycle is completed, resulting in human tapeworm infection, when humans ingest undercooked pork containing cysticerci. Cysts evaginate and attach to the
Notes made by kozi1989 email:kozi1989@yahoo.gr
184
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
small intestine by their scolex. Adult tapeworms develop, (up to 2 to 7 m in length and produce less than 1000 proglottids, each with approximately 50,000 eggs) and reside in the small intestine for years. Diagnosis requires biopsy of the infected tissue and examination of feces. T. solium eggs and proglottids found in feces diagnoses taeniasis and not cysticercosis
79. Class Cestoda. Taeniarhynchus saginatus (Taenia saginata)
Taenia saginata, also known as Taeniarhynchus saginata or the beef tapeworm, is a parasite of both cattle and humans, causing taeniasis in humans. Taenia saginata occurs where cattle are raised by infected humans maintaining poor hygiene, human feces are improperly disposed of, meat inspection programs are poor, and where meat is eaten without proper cooking. The disease is relatively common in Africa, some parts of Eastern Europe, Southeast Asia, and Latin America. T. saginata is normally 4 m to 10 m in length, but can become very large, over 12 m long in some situations. The body is whitish in colour, divided into the anterior scolex, followed by a short neck and a highly extended body proper called strobila. Unlike other tapeworms the scolex does not have a rostellum or scolex armature. It is composed of 4 powerful suckers. The strobila is composed a series of ribbon-like segments called proglottids. The segments are made up of mature and gravid proglottids. T. saginata is the largest of genus Taenia consisting between 1000 to 2000 proglottids and can also have a lifespan of 25 years in a host's intestine. The mature proglottid contains the uterus (unbranched), ovary, genital pore, testes, and vitelline gland . It does not have a digestive system, no mouth, no anus, or digestive tract. It is also an acoelomate, meaning that it does not have a body cavity. In the gravid proglottid, the uterus is branched and is filled with eggs. The gravid segments detach and are passed in the feces. Each of these segments can act like a worm. When they dry up, the proglottid ruptures, and the eggs are released. The egg can only infect cattle, the intermediate host. Inside the cow's duodenum the oncosphere hatches with the help of the gastric and intestinal secretions and migrates through the blood to the muscle. There it develops into infective cysticercoid cysticerci. Life cycle: The life cycle is indirect and complicated, and is completed in humans as the definitive host and cattle as the intermediate host. The adult worm inhabits the small intestine of humans. Fertilized eggs are released through the faeces along with the gravid proglottid which gets detached from the strobila. Cattle ingest the infective embryo while grazing. The digestive enzymes will break the thick shell of the egg and allow formation of the zygotes called "oncospheres". These zygotes then penetrate the mucous layer of the digestive tract and enter the circulation of the host . This is where the young larval stages form a pea-sized, fluid filled cyst, also known as Cysticercus and these cysts seem to form in the muscular fibers and are sometimes seen in specific organs like the lungs and liver. Humans acquire the infective larvae from eating undercooked meat. The digestive
Notes made by kozi1989 email:kozi1989@yahoo.gr
185
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
enzymes break down the cysticercus and the larval cyst is released and the inverted scolex is able to come out and attach to the hosts intestine. Adult tapeworm soon develop, and within three months it can reach 5 m long.
80. Class Cestoda. Diphyllobothrium latum. Hymenolepis nana
Diphyllobothrium is a genus of tapeworm which can cause Diphyllobothriasis in humans through consumption of raw or undercooked fish. The principal species causing diphyllobothriosis is Diphyllobothrium latum, known as the broad or fish tapeworm, or broad fish tapeworm. D. latum is a pseudophyllid cestode that infects fish and mammals. D. latum is native to Scandinavia, western Russia, and the Baltics, though it is now also present in North America, especially the Pacific Northwest. In Far East Russia, D. klebanovskii, having Pacific salmon as its second intermediate host, was identified. The adult worm is composed of three fairly distinct morphological segments: the scolex (head), the neck, and the lower body. Each side of the scolex has a slit-like groove, which is a bothrium (tentacle) for attachment to the intestine. The scolex attaches to the neck, or proliferative region. From the neck, grows many proglottid segments which contain the reproductive organs of the worm. D. latum is the longest tapeworm in humans, averaging ten meters long. Adults can shed up to a million eggs a day. In adults, proglottids are wider than they are long (hence the name broad tapeworm). As in all pseudophyllid cestodes, the genital pores open midventrally. Life cycle: Adult tapeworms may infect humans, canids, felines, bears, pinnipeds, and mustelids..Immature eggs are passed in feces of the mammal host (the definitive host, where the worms reproduce). After ingestion by a suitable freshwater crustacean such as a copepod (the first intermediate host), the coracidia develop into procercoid larvae. Following ingestion of the copepod by a suitable second intermediate host, typically a minnow or other small freshwater fish, the procercoid larvae are released from the crustacean and migrate into the fish's flesh where they develop into a plerocercoid larvae (sparganum). The plerocercoid larvae are the infective stage for the definitive host (including humans). Because humans do not generally eat undercooked minnows and similar small freshwater fish, these do not represent an important source of infection. Nevertheless, these small second intermediate hosts can be eaten by larger predator species, for example, trout, perch, and walleyed pike. In this case, the sparganum can migrate to the musculature of the larger predator fish and mammals can acquire the disease by eating these later intermediate infected host fish raw or undercooked. After ingestion of the infected fish, the plerocercoids develop into immature adults and then into mature adult tapeworms which will reside in the small intestine. The adults attach to the intestinal mucosa by means of the two bilateral grooves (bothria) of their scolex. The adults can reach more than 10 m (up to 30 ft) in length in some species such as D. latum, with more than 3,000 proglottids. One or several of the tape-like proglottid segments (hence the name tape-worm) regularly detach
Notes made by kozi1989 email:kozi1989@yahoo.gr
186
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
from the main body of the worm and release immature eggs in fresh water to start the cycle over again. Immature eggs are discharged from the proglottids (up to 1,000,000 eggs per day per worm) and are passed in the feces. The incubation period in humans, after which eggs begin to appear in the feces is typically 46 weeks, but can vary from as short as 2 weeks to as long as 2 years. The tapeworm can live up to 20 years Dwarf tapeworm (Hymenolepis nana, previously known as Vampirolepis nana, Hymenolepis fraterna, and Taenia nana) is a cosmopolitan species that is one of the most common cestodes of humans in the world, especially among children. It can be found throughout the world, but is usually most common in temperate zones. It is the most common cestode of humans and is found wherever there are human inhabitants. As its name implies (Greek: nanos dwarf), it is a small species, seldom exceeding 40 mm long and 1 mm wide. The scolex bears a retractable rostellum armed with a single circle of 20 to 30 hooks. The scolex also has four suckers, or a tetrad. The neck is long and slender, and the segments are wider than long. Genital pores are unilateral, and each mature segment contains three testes. After apolysis gravid segments disintegrate, releasing eggs, which measure 30 m to 47 m in diameter. The oncosphere is covered with a thin, hyaline, outer membrane and an inner, thick membrane with polar thickenings that bear several filaments. The heavy embryophores that give taeniid eggs their characteristic striated appearance are lacking in this and the other families of tapeworms infecting humans. Life cycle Infection is acquired most commonly from eggs in the feces of another infected individual, which are transferred in food, by contaminated fingers, or in sewagecontaminated drinking water. Hymenolepis nana is the only cestode that does not require an intermediate host to develop into its infective stage . When eaten by a person or rodent, eggs hatch in the duodenum, releasing oncospheres, which penetrate the mucosa and come to lie in lymph channels of the villi. Oncospheres develops into a cysticercoid which has a tail and a well formed scolex. It is made of longitudinal fibers and is spade shaped with the rest of the worm still inside the cyst. In five to six days cysticercoids emerge into the lumen of the small intestine, where they attach and mature.The direct life cycle is doubtless a recent modification of the ancestral two-host life cycle, found in other species of hymenolepidids, because cysticercoids of H. nana can still develop normally within larval fleas and beetles.
81. Class Cestoda. Echinococcus granulosus
Echinococcus granulosus, also called the Hydatid worm or Hyper Tape-worm, is a cyclophyllid cestode that parasitizes the small intestine of canids as an adult, but which has important intermediate hosts such as livestock and humans, where it causes hydatid disease. The adult tapeworm ranges in length from 2 mm to 7 mm and has three proglottids ("segments") when intact - an immature proglottid, mature proglottid and a gravid
Notes made by kozi1989 email:kozi1989@yahoo.gr
187
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
proglottid. Like all cyclophyllideans, E. granulosus has four suckers on its scolex ("head"), and E. granulosus also has a rostellum with hooks. In canids, E. granulosus causes a typical tapeworm infection, and produces eggs that are passed with the dog's feces. Intermediate hosts include herbivores such as sheep, deer, moose, kangaroos, and wallabies, and any other organism (including humans) that ingests dog feces. In the intermediate host, eggs hatch into oncosphere larvae that travel through the blood and form hydatid cysts in the host's tissues. These cysts can grow to be the size of a softball or basketball, and may contain several smaller "balloons" inside the main cyst. In the related worm Echinococcus multilocularis, the outer cyst is not present. If the outer cyst ruptures, new cysts can form at a different location in the body. Each smaller section contains several juvenile worms, and dogs may eat millions of them, resulting in very heavy infections. Hydatid cysts occur in organs like the liver, brain and lungs, not in subcutaneous tissue. Though this has never been tested experimentally, it is assumed that infected animals make easier prey for canids.
82. Phylum Nematoda. Ascaris lumbricoides
Ascaris lumbricoides is the giant roundworm of humans, belonging to the phylum Nematoda. An ascarid nematode, it is responsible for the disease ascariasis in humans, and it is the largest and most common parasitic worm in humans. A quarter of the human population is estimated to be infected by this parasite. Ascariasis is prevalent worldwide and more so in tropical and subtropical countries. Ascaris lumbricoides is characterized by its great size . Males are 24 mm in diameter and 1531 cm long. The males' posterior end is curved ventrally and has a bluntly pointed tail. Females are 36 mm wide and 2049 cm long. The vulva is located in the anterior end and accounts for about a third of its body length. Uteri may contain up to 27 million eggs at a time with 200,000 being laid per day. Fertilized eggs are oval to round in shape and are 45-75 micrometers long and 35-50 micrometers wide with a thick outer shell. Unfertilized eggs measure 88-94 micrometers long and 44 micrometers wide. Life cycle Ascaris lumbricoides, or "roundworm", infections in humans occur when an ingested infective egg releases a larval worm that penetrates the wall of the duodenum and enters the blood stream. From here, it is carried to the liver and heart, and enters pulmonary circulation to break free in the alveoli, where it grows and molts. In 3 weeks, the larvae pass from the respiratory system to be coughed up, swallowed, and thus returned to the small intestine, where they mature to adult male and female worms. Fertilization can now occur and the female produces as many as 200,000 eggs per day for a year. These fertilized eggs become infectious after 2 weeks in soil; they can persist in soil for 10 years or more. The eggs have a lipid layer, that makes them resistant to the effects of acids and alkalis as well as other chemicals. This resilience helps to explain why this nematode is such a ubiquitous parasite.
Notes made by kozi1989 email:kozi1989@yahoo.gr
188
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
83. Phylum Nematoda. Enterobius vermicularis. Trichocephalus trichiurus (Trichuris
trichiura) The pinworm (in the United States of America) (genus Enterobius), also known as threadworm (in the United Kingdom) or seatworm, is a nematode (roundworm) and a common human intestinal parasite, especially in children.[5] The medical condition associated with pinworm infestation is known as enterobiasis,[6] or less precisely as oxyuriasis in reference to the family Oxyuridae. The pinworm appears as a white, small and delicate nematode (i.e., roundworm).[12] The adult female has a sharply pointed posterior end, is 8 to 13 millimeters long, and 0.5 millimeter thick.[12] The adult male is considerably smaller, measuring 2 to 5 millimeters long and 0.2 millimeter thick, and has a curved posterior end.[12] The eggs are translucent[12] and have a surface that adheres to environmental objects.[13] The eggs measure 50 to 60 micrometers by 20 to 30 micrometers, and have a thick shell that is flattened on one side.[12] The small size and colorlessness of the eggs make them invisible to the naked eye, except in barely visible clumps of thousands of eggs.[14][15] Eggs may contain a developing embryo or a fully developed pinworm larva.[12] Inside the host, the larvae grow to 140150 micrometers in length. Life cycle The entire life cyclefrom egg to adulttakes place in the human gastrointestinal tract of a single human host. The life cycle begins with eggs being ingested.[13] The eggs hatch in the duodenum (i.e., first part of the small intestine).[20] The emerging pinworm larvae grow rapidly to a size of 140 to 150 micrometers in size,[19] and migrate through the small intestine towards the colon.[13] During this migration they moult twice and become adults.[13][17] Females survive for 5 to 13 weeks, and males about 7 weeks.[13] The male and female pinworms mate in the ileum (i.e., last part of the small intestine),[13] whereafter the male pinworms usually die,[20] and are passed out with stool.[14] The gravid female pinworms settle in the ileum, caecum (i.e., beginning of the large intestine), appendix and ascending colon,[13] where they attach themselves to the mucosa[17] and ingest colonic contents.[16] Almost the entire body of a gravid female becomes filled with eggs.[20] The estimations of the number of eggs in a gravid female pinworm ranges from about 11,000[13] to 16,000.[17] The egg-laying process begins approximately five weeks after initial ingestion of pinworm eggs by the human host.[13] The gravid female pinworms migrate through the colon towards the rectum at a rate of 12 to 14 centimeters per hour.[13] They emerge from the anus, and while moving on the skin near the anus, the female pinworms deposit eggs either through (1) contracting and expelling the eggs, (2) dying and then disintegrating, or (3) bodily rupture due to the host scratching the worm.[20] After depositing the eggs, the female becomes opaque and dies.[14] The reason the female emerges from the anus is to obtain the oxygen necessary for the maturation of the eggs.
Notes made by kozi1989 email:kozi1989@yahoo.gr
189
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Pinworms spread through human-to-human transmission, by ingesting (i.e., swallowing) infectious pinworm eggs.[17][20] The eggs are hardy and can remain viable (i.e., infectious) in a moist environment for up to three weeks. They do not tolerate heat well, but can survive in low temperatures: two-thirds of the eggs are still viable after 18 hours at 8 degrees Celsius (18 F). After the eggs have been initially deposited near the anus, they are readily transmitted to other surfaces through contamination.[20] The surface of the eggs is sticky when laid[13][14], and the eggs are readily transmitted from their initial deposit near the anus to fingernails, hands, night-clothing and bed linen.[19] From here, eggs are further transmitted to food, water, furniture, toys, bathroom fixtures and other objects.[13][17][20] Household pets often carry the eggs in their fur, while not actually being infected.[15] Dust containing eggs can become airborne and widely dispersed when dislodged from surfaces, for instance when shaking out bed clothes and linen.[17][14][15] Consequently the eggs can enter the mouth and nose through inhalation, and be swallowed later.[17][14][19][20] Although pinworms do not strictly multiply inside the body of their human host,[19] some of the pinworm larvae may hatch on the anal mucosa, and migrate up the bowel and back into the gastrointestinal tract of the original host.[17][19] This process is called retroinfection. The human whipworm (Trichuris trichiura or Trichocephalus trichiuris) is a roundworm, which causes trichuriasis when it infects a human large intestine. The name whipworm refers to the shape of the worm; they look like whips with wider "handles" at the posterior end. Trichuris trichiura has a narrow anterior esophageal end and shorter and thicker posterior anus. These pinkish-white worms are threaded through the mucosa. They attach to the host through their slender anterior end and feed on tissue secretions instead of blood. Females are larger than males; approximately 3550 mm long compared to 3045 mm.[2] The females have a bluntly round posterior end compared to their male counterparts with a coiled posterior end. Their characteristic eggs are barrel-shaped, brown, and have bipolar protuberances. Life cycle The female T. trichiura produces 2,00010,000 single celled eggs per day.[1] Eggs are deposited from human feces to soil where, after two to three weeks, they become embryonated and enter the infective stage. These embryonated infective eggs are ingested and hatch in the human small intestine. This is the location of growth and molting. The infective larvae penetrate the villi and continue to develop in the small intestine. The young worms move to the cecum and penetrate the mucosa and there they complete development to adult worms in the large intestine. The life cycle from time of ingestion of eggs to development of mature worms takes approximately three months. During this time, there may be limited signs of infection in stool samples due to lack of egg production and
Notes made by kozi1989 email:kozi1989@yahoo.gr
190
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
shedding. The female T. trichiura begin to lay eggs after three months of maturity. Worms can live up to five years, during which time females can lay up to 20,000 eggs per day.
84. PhylumNematoda. Trichinella spiralis
Trichinella spiralis is a nematode parasite, occurring in rats, pigs, bears and humans, and is responsible for the disease trichinosis. It is sometimes referred to as the "pork worm" due to it being found commonly in undercooked pork products. Trichinella species are the smallest nematode parasite of humans, have an unusual life cycle and are one of the most widespread and clinically important parasites in the world. The small adult worms mature in the intestines of an intermediate host such as a pig . Each adult female produces batches of live larvae, which bore through the intestinal wall, enter the blood (to feed on it) and lymphatic system, and are carried to striated muscle, Once in the muscle, they encyst, or become enclosed in a capsule. Humans can be infected by eating infected pork or wild carnivores such as fox, cat or bear. Males of T. spiralis measure between 1.4 mm and 1.6 mm long and are flat anteriorly than posteriorly. The anus can be found in the terminal (side) and they have a large copulatory pseudobursa on each side. he females of T. spiralis are about twice the size of the males and have an anus found terminally. The vulva is located near the esophagus. The single uterus of the female is filled with developing eggs in the posterior portion, while the anterior portion contained the fully developed juveniles. Life cycle Trichinella spiralis is a parasitic nematode that has a direct life cycle, meaning it completes all stages of development in one host. The larval forms are encapsulated as a small cystic structure within the infected host. Humans typically become infected when they eat improperly cooked pork or Trichinella infected meat. When a human eats the infected meat, the larvae are released from the nurse cell (due to stomach pH) and migrate to the intestine, where they burrow into the intestinal mucosa, mature, and reproduce. Interestingly, juveniles within nurse cells have an anaerobic or facultative anaerobic metabolism, but when they become activated, they adopt an aerobic metabolism characteristics of the adult. Female Trichinella worms live for about six weeks, and in that time can produce up to 1,500 larvae; when a spent female dies, she passes out of the host. The larvae can then gain access to the circulation and migrate around the body of the host The migration and encystment of larvae can cause fever and pain brought upon by the host inflammatory response. In some cases, migration to specific organ tissues can cause myocarditis and encephalitis that can result in death.
85. Phylum Nematoda. Strongyloides stercoralis. Ancylostoma duodenale.
Strongyloides stercoralis, also known as the threadworm, is the scientific name of a human parasitic roundworm causing the disease of strongyloidiasis. Strongyloides stercoralis is a nematode that can parasitize humans. The adult parasitic stage lives in tunnels in the mucosa of the small intestine.
Notes made by kozi1989 email:kozi1989@yahoo.gr
191
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Whereas males grow to only about 0.9 mm in length, females can be anywhere from 2.0 to 2.5 mm. Both genders also possess a tiny buccal capsule and cylindrical esophagus without a posterior bulb.[9] In the free-living stage, the esophagi of both sexes are rhabditiform. Males can be distinguished from their female counterparts by two structures: the spicules and gubernaculum. The Strongyloides' life cycle is heterogonic - it is more complex than that of most nematodes with its alternation between free-living and parasitic cycles, and its potential for autoinfection and multiplication within the host. The parasitic has a homogenic life cycle, while the free-living has a heterogonic life cycle. The heterogonic life cycle is advantageous to the parasite because it allows reproduction for one or more generations in the absence of a host. Life cycle In the free-living cycle, the rabditiform larvae passed in the stool can either molt twice and become infective filariform larvae (direct development) or molt four times and become free living adult males and females that mate and produce eggs from which rabditiform larvae hatch. In the direct development L1 (1st-stage larvae) transform into IL (infective larvae) via three molts. In the indirect route results first in the development of free-living adults which mate; the female lays eggs which hatch and then develop into IL. The direct route gives IL faster (3 days) versus the indirect route (710 days). However, the indirect route results in an increase in the number of IL produced. Speed of development of IL is traded off for increased numbers. The free-living males and females of S. stercoralis die after one generation; they do not persist in the soil. The latter in turn can either develop into a new generation of free-living adults, or into infective filariform larvae. The filariform larvae penetrate the human host skin to initiate the parasitic cycle. The infectious larvae penetrate the skin when there is contact with the soil. While S. stercoralis is attracted to chemicals such as carbon dioxide or sodium chloride, these chemicals are very non-specific. Larvae have been thought to locate their hosts via chemicals in the skin, predominantly urocanic acid, a histidine metabolite on the uppermost layer of skin that is removed by sweat or the daily skin-shedding cycle.[8] Urocanic acid concentrations can be up to five times greater in the foot than any other part of the human body. Some of them enter the superficial veins and ride the blood vessels to the lungs, where they enter the alveoli. They are then coughed up and swallowed into the gut, where they parasitise the intestinal mucosa (duodenum and jejunum). In the small intestine they molt twice and become adult female worms. The females live threaded in the epithelium of the small intestine and by parthenogenesis produce eggs, which yield rabditiform larvae. Only females will reach reproductive adulthood in the intestine. Female strongyloides reproduce through parthenogenesis. The eggs hatch in the intestine and young larvae are then excreted in the feces. It takes about two weeks to reach egg development from the
Notes made by kozi1989 email:kozi1989@yahoo.gr
192
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
initial skin penetration. By this process, S. stercoralis can cause both respiratory and gastrointestinal symptoms. The worms also participate in autoinfection, in which the rabditiform larvae become infective filariform larvae, which can penetrate either the intestinal mucosa (internal autoinfection) or the skin of the perianal area (external autoinfection); in either case, the filariform larvae may follow the previously described route, being carried successively to the lungs, the bronchial tree, the pharynx, and the small intestine where they mature into adults; or they may disseminate widely in the body. To date, occurrence of autoinfection in humans with helminthic infections is recognized only in Strongyloides stercoralis and Capillaria philippinensis infections. In the case of Strongyloides, autoinfection may explain the possibility of persistent infections for many years in persons who have not been in an endemic area and of hyperinfections in immunodepressed individuals. Ancylostoma duodenale is a species of the worm genus Ancylostoma. It is a parasitic nematode worm and commonly known as Old World hookworm. It lives in the small intestine of host such as humans, cats and dogs. Ancylostoma duodenale and Necator americanus are the two human hookworms that are normally discussed together as the cause of hookworm infection. 90% asymptomatic can lead to iron deficiency anemia. Ancylostoma duodenale, a member of the Ancylostomidae, is also commonly referred to as a "hookworm". As such, hookworms have the ability to mate and mature in the small intestine of their host. Ancylostoma duodenale has two ventral plates at the anterior margin of the buccal capsule. Each of them has two large teeth that are fused at their bases. A pair of small teeth can be found in the depths of the buccal capsule. Males are 8 mm to 11 mm long with a copulatory bursa at the posterior end. It also has needlelike spicules which are not fused and have simple tips. Females are 10 mm to 13 mm long with the vulva located at the posterior end and can lay 10,000 to 30,000 eggs per day. Their life span is one year. Life cycle When a filariform larva (infective stage) penetrates the intact skin, the larva enters the blood circulation. Then it is carried to the lungs, coughed up and swallowed back to the small intestine. The larva later matures into adult in the small intestine and female worms can lay 25,000 eggs per day. The eggs are released into the feces and resided on soil. Embryonated egg on soil will hatch into juvenile 1 stage (rhabditiform or noninfective stage) and mature into filariform larvae. The filariform larvae can then penetrate another exposed skin and begin a new cycle of human infection.
86. Phylum Nematoda. Dracunculus medinensis. Wuchereria bancrofti. Loa loa.
Dracunculiasis, also known as Guinea worm disease, is caused by the large female nematode, Dracunculus medinensis,[2] which is among the longest nematodes infecting humans.[3] The adult female is primarily larger than the adult male. The longest adult female
Notes made by kozi1989 email:kozi1989@yahoo.gr
193
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
recorded was 800 millimetres (31 in), while the adult male was only 40 mm (1.6 in).[4] Mature female worms migrate along subcutaneous tissues to reach the skin below the knee, forming a painful ulcerating blister.[3] They can also emerge from other parts of the body like the head, torso, upper extremities, buttocks, and genitalia. Life cycle Humans become infected by drinking unfiltered water containing copepods (small crus taceans) that have been infected with D. medinensis larvae. After ingestion, the copepods die and release the stage 3 larvae, which then penetrate the host's stomach, intestinal wall, and enter into the abdominal cavity and retroperitoneal space. After maturing, adult male worms die while the females migrate in the subcutaneous tissues towards the surface of the skin. After about a year of infection, the female worm forms a blister on the skin, generally on the distal lower extremity (foot), which breaks open. The patient then seeks to relieve the local discomfort by placing their foot in water, but when the lesion comes into contact with water, the female worm emerges and releases her stage 1 larvae. The larvae are then ingested by a copepod, and after two weeks (and two molts) the stage 3 larvae becomes infectious. Ingestion of the copepods is the last stage that completes the cycle. Filaria, is a parasitic filarial nematode (roundworm) spread by a mosquito vector. It is one of the three parasites that cause lymphatic filariasis, an infection of the lymphatic system by filarial worms. Life cycle W. bancrofti carry out their life cycle in two hosts. Human beings serve as the definitive host and mosquitoes as their intermediate hosts. The adult parasites reside in the lymphatics of the human host. They are viviparous. The first stage larvae are known as microfilariae. The microfilariae are present in the circulation. The microfilariae migrate between the deep and the peripheral circulation. W. bancrofti is a periodic strain that exhibits nocturnal periodicity. During the day they are present in the deep veins and during the night they migrate to the peripheral circulation. Next, the microfilariae are transferred into a vector; the most common vectors are the mosquito species: Culex, Anopheles, Mansonia, and Aedes. Inside the mosquito vector, also known as the intermediate host, the microfilariae mature into motile larvae called juveniles. When the mosquito vector has its next blood meal, W. bancrofti is egested via the mosquitos proboscis into the blood stream of the new human host. The larvae move through the Lymphatic system to regional lymph nodes, predominantly in the legs and genital area. The larvae develop into adult worms over the course of a year and reach sexual maturity in the afferent lymphatic vessels. After mating, the adult female worm can produce thousands of microfilariae that migrate into the bloodstream. A mosquito vector can bite the infected human host, ingest the microfilariae, and thus repeat the life cycle of W. bancrofti.
Notes made by kozi1989 email:kozi1989@yahoo.gr
194
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Loa loa is the filarial nematode (roundworm) species that causes Loa loa filariasis. It is commonly known as the "eye worm". Its geographic distribution includes Africa, Sudan and India. L. loa is one of four parasitic filarial nematodes that cause subcutaneous filariasis in humans. The three other filarial nematodes are Mansonella streptocerca, Onchocerca volvulus (causes river blindness), and Dracunculus medinensis (guinea worm). Maturing larvae and adults of the "eye worm" occupy the subcutaneous layer of the skin the fat layer of humans, causing disease. The young larvae develop in horseflies of the genus Chrysops (deer flies, yellow flies), including the species C. dimidiata and C. silacea, which infect humans by biting them. Loa loa worms have a simple body including a head, body, and tail. Males range from 20mm to 34mm long and 350m to 430m wide. Females range from 20mm to 70mm l ong and are about 425m wide. Life cycle Three species involved in the life cycle include the parasite Loa loa, the fly vector, and the human host: 1) 2) 3) 4) A vector fly bites an infected human host and ingests microfilariase. Microfilariae move to the thoracic muscles of the host. Microfilariae develop into first stage larvae, then third stage larvae. Third stage larvae (infective) travel to the proboscis of fly.
An infected vector fly bites an uninfected human host and the third stage larvae penetrates the skin and enters human subcutaneous tissue. Larvae mature into adults, who produce microfilariae that have been found in spinal fluid, urine, peripheral blood, and lungs.
87. Phylum Annelida. Lumbricus terrestris. Hirudo medicinalis.
Lumbricus terrestris is a large reddish worm native to Europe, but now also widely distributed elsewhere around the world (along with several other lumbricids), due to human introductions. In some areas where it has been introduced, some people consider it to be a serious pest species, since it is out-competing locally native worms. Through much of Europe it is the largest naturally occurring species of earthworm, typically reaching 20 - 25 cm in length when extended (though in parts of southern Europe there are native species which are much larger). It has an unusual habit of copulating on the surface at night, which makes it more visible than most other earthworms. Medicinal leeches are any of several species of leeches, but most commonly Hirudo medicinalis, the European medicinal leech. General morphology follows that of most other leeches. Fully mature adults can be
Notes made by kozi1989 email:kozi1989@yahoo.gr
195
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
up to 20 cm in length and are green, brown or greenish brown with a darker tone on the dorsal side and a lighter ventral side, the dorsal side also has a thin red stripe. These organisms have two suckers, one at each end, called the anterior and posterior sucker. The posterior is mainly used for leverage while the anterior sucker, consisting of the jaw and teeth, is where the feeding takes place. Medicinal leeches have three jaws (tripartite) that look like little saws, and on them are about 100 sharp teeth used to incise the host. The incision leaves a mark which is an inverted Y inside of a circle. After piercing the skin and injecting anti-coagulants (Hirudin) and anaesthetics they suck out blood. Large adults can consume up to ten times their body weight in a single meal, with 5-15 ml being the average volume taken.[2] These leeches can live for up to a year between feeding. Medicinal leeches are hermaphrodites which reproduce by sexual mating, laying eggs in clutches of up to 50 near (but not under) water, and in shaded, humid places.
88. Phylum Arthropoda general characterization. Class Crustacea general
characterization. Class Arachnoidea(Arachnida, arachnids). Order Scorpiones(scorpions) and order Aranei (spiders) An arthropod is an invertebrate animal having an exoskeleton (external skeleton), a segmented body, and jointed appendages. Arthropods are members of the phylum Arthropoda (from Greek rthron, "joint", and pods "foot", which together mean "jointed feet"), and include the insects, arachnids, crustaceans, and others. Arthropods are characterized by their jointed limbs and cuticles, which are mainly made of -chitin; the cuticles of crustaceans are also biomineralized with calcium carbonate. The rigid cuticle inhibits growth, so arthropods replace it periodically by molting. The arthropod body plan consists of repeated segments, each with a pair of appendages. It is so versatile that they have been compared to Swiss Army knives, and it has enabled them to become the most species-rich members of all ecological guilds in most environments. They have over a million described species, making up more than 80% of all described living animal species, and are one of only two animal groups that are very successful in dry environments the other being the amniotes. They range in size from microscopic plankton up to forms a few meters long. Arthropods' primary internal cavity is a hemocoel, which accommodates their internal organs and through which their blood circulates; they have open circulatory systems. Like their exteriors, the internal organs of arthropods are generally built of repeated segments. Their nervous system is "ladder-like", with paired ventral nerve cords running through all segments and forming paired ganglia in each segment. Their heads are formed by fusion of varying numbers of segments, and their brains are formed by fusion of the ganglia of these segments and encircle the esophagus. The respiratory and excretory systems of arthropods vary, depending as much on their environment as on the subphylum to which they belong. Their vision relies on various combinations of compound eyes and pigment-pit ocelli: in most species the ocelli can only detect the direction from which light is coming, and the
Notes made by kozi1989 email:kozi1989@yahoo.gr
196
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
compound eyes are the main source of information, but the main eyes of spiders are ocelli that can form images and, in a few cases, can swivel to track prey. Arthropods also have a wide range of chemical and mechanical sensors, mostly based on modifications of the many setae (bristles) that project through their cuticles. Arthropods' methods of reproduction and development are diverse; all terrestrial species use internal fertilization, but this is often by indirect transfer of the sperm via an appendage or the ground, rather than by direct injection. Aquatic species use either internal or external fertilization. Almost all arthropods lay eggs, but scorpions give birth to live young after the eggs have hatched inside the mother. Arthropod hatchlings vary from miniature adults to grubs and caterpillars that lack jointed limbs and eventually undergo a total metamorphosis to produce the adult form. The level of maternal care for hatchlings varies from nonexistent to the prolonged care provided by scorpions. Crustaceans (Crustacea) form a very large group of arthropods, usually treated as a subphylum, which includes such familiar animals as crabs, lobsters, crayfish, shrimp, krill and barnacles. The 50,000 described species range in size from Stygotantulus stocki at 0.1 mm (0.004 in), to the Japanese spider crab with a leg span of up to 12.5 ft (3.8 m) and a mass of 44 lb (20 kg). Like other arthropods, crustaceans have an exoskeleton, which they moult to grow. They are distinguished from other groups of arthropods, such as insects, myriapods and chelicerates by the possession of biramous (two-parted) limbs, and by the nauplius form of the larvae. Most crustaceans are free-living aquatic animals, but some are terrestrial (e.g. woodlice), some are parasitic (e.g. fish lice, tongue worms) and some are sessile (e.g. barnacles). The group has an extensive fossil record, reaching back to the Cambrian, and includes living fossils such as Triops cancriformis, which has existed apparently unchanged since the Triassic period. More than 10 million tons of crustaceans are produced by fishery or farming for human consumption, the majority of it being shrimps and prawns. Krill and copepods are not as widely fished, but may be the animals with the greatest biomass on the planet, and form a vital part of the food chain. The scientific study of crustaceans is known as carcinology (alternatively, malacostracology, crustaceology or crustalogy), and a scientist who works in carcinology is a carcinologist. Arachnids are a class (Arachnida) of joint-legged invertebrate animals in the subphylum Chelicerata. All arachnids have eight legs, although in some species the front pair may convert to a sensory function. The term is derived from the Greek word (archn), meaning "spider".lmost all extant arachnids are terrestrial. However, some inhabit freshwater environments and, with the exception of the pelagic zone, marine environments as well. They comprise over 100,000 named species, including spiders, scorpions, harvestmen, ticks, mites and Solifugae.[3] Basic characteristics of arachnids include four pairs of legs (1) and a body divided into two segments: the cephalothorax (2) and the abdomen (3). Almost all adult arachnids have eight legs, and arachnids may be easily distinguished
Notes made by kozi1989 email:kozi1989@yahoo.gr
197
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
from insects by this fact, since insects have six legs. However, arachnids also have two further pairs of appendages that have become adapted for feeding, defense, and sensory perception. The first pair, the chelicerae, serve in feeding and defense. The next pair of appendages, the pedipalps have been adapted for feeding, locomotion, and/or reproductive functions. In Solifugae, the palps are quite leg-like, so that these animals appear to have ten legs. The larvae of mites and Ricinulei have only six legs; the fourth pair appears when they moult into nymphs. However, there are also adult mites with six, or even four legs. Arachnids are further distinguished from insects by the fact they have no antennae or wings. Their body is organized into two tagma called the prosoma, or cephalothorax, and the opisthosoma, or abdomen. The cephalothorax is derived from the fusion of the cephalon (head) and the thorax, and is usually covered by a single, unsegmented carapace. The abdomen is segmented in the more primitive forms, but varying degrees of fusion between the segments occur in many groups. It is typically divided into a preabdomen and postabdomen, although this is only clearly visible in scorpions, and in some orders, such as the Acari, the abdominal sections are completely fused. Like all arthropods, arachnids have an exoskeleton, and they also have an internal structure of cartilage-like tissue called the endosternite, to which certain muscle groups are attached. The endosternite is even calcified in some Opiliones. Scorpions are predatory arthropod animals of the order Scorpiones within the class Arachnida. They have eight legs and are easily recognized by the pair of grasping claws and the narrow, segmented tail, often carried in a characteristic forward curve over the back, ending with a venomous stinger. Scorpions are characterised by a metasoma (tail) comprising six segments, the last containing the scorpion's anus and bearing the telson (the sting). The telson, in turn, consists of the vesicle, which holds a pair of venom glands and the hypodermic aculeus, the venominjecting barb. The abdomen's front half, the mesosoma, is made up of six segments. The first segment contains the sexual organs as well as a pair of vestigial and modified appendages forming a structure called the genital operculum. The second segment bears a pair of featherlike sensory organs known as the pectines; the final four segments each contain a pair of book lungs. The mesosoma is armored with chitinous plates, known as tergites on the upper surface and sternites on the lower surface. The cuticle of scorpions is covered with hairs in some places that act like balance organs. An outer layer that makes them fluorescent green under ultraviolet light is called the hyaline layer. Newly molted scorpions do not glow until after their cuticle has hardened. The fluorescent hyaline layer can be intact in fossil rocks that are hundreds of millions of years old. Scorpions are opportunistic predators of small arthropods and insects . They use their chela (pincers) to catch the prey initially. Depending on the toxicity of their venom and size of their claws, they will then either crush the prey or inject it with neurotoxic venom. The neurotoxins consist of a variety of small proteins as well as sodium and potassium cations, which serve to interfere with neurotransmission in the victim. Scorpions use their
Notes made by kozi1989 email:kozi1989@yahoo.gr
198
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
venom to kill or paralyze their prey so that it can be eaten; in general it is fast acting, allowing for effective prey capture. Scorpion venoms are optimised for action upon other arthropods and therefore most scorpions are relatively harmless to humans; stings produce only local effects (such as pain, numbness or swelling). A few scorpion species, however, mostly in the family Buthidae, can be dangerous to humans. The scorpion that is responsible for the most human deaths is the Androctonus australis, or fat-tailed scorpion of North Africa. The toxicity of A. australis's venom is roughly half that of the deathstalker (Leiurus quinquestriatus), but since A. australis injects quite a bit more venom into its prey, it is the most deadly to humans. Human deaths normally occur in the young, elderly, or infirm; scorpions are generally unable to deliver enough venom to kill healthy adults. Some people, however may be allergic to the venom of some species, in which case the scorpion's sting can more likely kill. A primary symptom of a scorpion sting is numbing at the injection site, sometimes lasting for several days. It has been found that scorpions have two types of venom: a translucent, weaker venom designed to stun only, and an opaque, more potent venom designed to kill heavier threats. Unlike the majority of Arachnida species, scorpions are viviparous. The young are born one by one, and the brood is carried about on its mother's back until the young have undergone at least one moult.[24] The young generally resemble their parents, requiring between five and seven moults to reach maturity. Scorpions have quite variable lifespans and the lifespan of most species is not known. The age range appears to be approximately 425 years (25 years being the maximum reported life span in the giant desert hairy scorpion, Hadrurus arizonensis). They are nocturnal and fossorial, finding shelter during the day in the relative cool of underground holes or undersides of rocks and coming out at night to hunt and feed. Scorpions prefer to live in areas where the temperature is 2037 C (6899 F), but may survive in the temperature range of 1445 C (57113 F). Spiders (order Araneae) are air-breathing arthropods that have eight legs, and chelicerae with fangs that inject venom. They are the largest order of arachnids and rank seventh in total species diversity among all other groups of organisms.[1] Spiders are found worldwide on every continent except for Antarctica, and have become established in nearly every habitat with the exception of air and sea colonization. As of 2008, approximately 40,000 spider species, and 109 families have been recorded by taxonomists;[2] however, there has been confusion within the scientific community as to how all these families should be classified, as evidenced by the over 20 different classifications that have been proposed since 1900.[3] Anatomically, spiders differ from other arthropods in that the usual body segments are fused into two tagmata, the cephalothorax and abdomen, and joined by a small, cylindrical pedicel. Unlike insects, spiders do not have antennae. In all except the most primitive group, the Mesothelae, spiders have the most centralized nervous systems of all arthropods, as all their ganglia are fused into one mass in the cephalothorax. Unlike most arthropods, spiders have no extensor muscles in their limbs and instead extend them by hydraulic pressure.
Notes made by kozi1989 email:kozi1989@yahoo.gr
199
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Their abdomens bear appendages that have been modified into spinnerets that extrude silk from up to six types of silk glands within their abdomen. Spider webs vary widely in size, shape and the amount of sticky thread used. It now appears that the spiral orb web may be one of the earliest forms, and spiders that produce tangled cobwebs are more abundant and diverse than orb-web spiders. Spider-like arachnids with silk-producing spigots appear in the Devonian period about 386 million years ago, but these animals apparently lacked spinnerets. True spiders have been found in Carboniferous rocks from 318 to 299 million years ago, and are very similar to the most primitive surviving order, the Mesothelae. The main groups of modern spiders, Mygalomorphae
89. Order Acari (ticks and mites). The role of ticks as vectors of transmissive
diseases Acari (or Acarina) are a taxon of arachnids that contains mites and ticks. Mites are arachnids and, as such, should have a segmented body with the segments organised into two tagmata: a prosoma (cephalothorax) and an opisthosoma (abdomen). However, only the faintest traces of primary segmentation remain in mites; the prosoma and opisthosoma are insensibly fused, and a region of flexible cuticle (the cirumcapitular furrow) separates the chelicerae and pedipalps from the rest of the body. This anterior body region is called the capitulum or gnathosoma and, according to some workers, is also found in Ricinulei. The remainder of the body is called the idiosoma and is unique to mites. Most adult mites have four pairs of legs, like other arachnids, but some have fewer. For example, gall mites like Phyllocoptes variabilis (family Eriophyidae) have a worm-like body with only two pairs of legs; some parasitic mites have only one or three pairs of legs in the adult stage. Larval and prelarval stages have a maximum of three pairs of legs; adult mites with only three pairs of legs may be called 'larviform'. The mouth parts of mites may be adapted for biting, stinging, sawing or sucking. They breathe through tracheae, stigmata (small openings of the skin), intestines and the skin itself. Species hunting for other mites have very acute senses, but many mites are eyeless. The central eyes of arachnids are always missing, or they are fused into a single eye. Thus, any eye number from none to five may occur. Ticks are excellent vectors for disease transmission.[1] More than 800 species of these obligate blood-sucking creatures inhabit the planet. They are second only to mosquitoes as vectors of human disease, both infectious and toxic. From the perspective of disease transmission to humans, the essential characteristic of ticks is their need to ingest a blood meal to transform to their next stage of development. Not picky in their eating habits, they take their requisite blood meal from all classes of vertebrates (eg, mammals, reptiles, birds), with the exception of fish. Ticks feed by perching in low vegetation and waiting (questing) for a susceptible host on which they can attach and feed. Once on a host, the tick attaches its hypostome, a central piercing element with hooks, into the host's skin. Some ticks secrete a cementing material to
Notes made by kozi1989 email:kozi1989@yahoo.gr
200
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
fasten themselves to the host. In addition, Ixodes ticks secrete anticoagulant, immunosuppressive, and anti-inflammatory substances into the area of the tick bite. These substances presumably help the tick to obtain a blood meal without the host's noticing. These same substances also help any freeloading pathogens to establish a foothold in the host. Ticks can carry and transmit a remarkable array of pathogens, such as bacteria, spirochetes, rickettsiae, protozoa, viruses, nematodes, and toxins. A single tick bite can transmit multiple pathogens, a phenomenon that has led to atypical presentations of some classic tick-borne diseases. In the United States, ticks are the most common vectors of vector-borne diseases.
90. Class insecta (inspects). The role of insects as vectors of transmissive diseases
Insects (from Latin insectum, a calque of Greek *ntomon+, "cut into sections") are a class of living creatures within the arthropods that have a chitinous exoskeleton, a three-part body (head, thorax, and abdomen), three pairs of jointed legs, compound eyes, and two antennae. They are among the most diverse groups of animals on the planet and include more than a million described species and represent more than half of all known living organisms.[2][3] The number of extant species is estimated at between six and ten million,[2][4][5] and potentially represent over 90% of the differing metazoan life forms on Earth.[6] Insects may be found in nearly all environments, although only a small number of species occur in the oceans, a habitat dominated by another arthropod group, the crustaceans. The life cycles of insects vary but most hatch from eggs. Insect growth is constrained by the inelastic exoskeleton and development involves a series of molts. The immature stages can differ from the adults in structure, habit and habitat and can include a passive pupal stage in those groups that undergo complete metamorphosis. Insects that undergo incomplete metamorphosis lack a pupal stage and adults develop through a series of nymphal stages.[7] The higher level relationship of the hexapoda is unclear. Fossilized insects of enormous size have been found from the Paleozoic Era, including giant dragonflies with wingspans of 55 to 70 cm (2228 in). The most diverse insect groups appear to have coevolved with flowering plants. Insects typically move about by walking, flying or occasionally swimming. As it allows for rapid yet stable movement, many insects adopt a tripedal gait in which they walk with their legs touching the ground in alternating triangles. Insects are the only invertebrates to have evolved flight. Many insects spend at least part of their life underwater, with larval adaptations that include gills and some adult insects are aquatic and have adaptations for swimming. Some species, like water striders, are capable of walking on the surface of water. Insects are mostly solitary, but some insects, such as certain bees, ants, and termites are social and live in large, well-organized colonies. Some insects, like earwigs, show maternal care, guarding their eggs and young. Insects can communicate with each other in a variety of ways. Male moths can sense the pheromones of female moths over distances of many
Notes made by kozi1989 email:kozi1989@yahoo.gr
201
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
kilometers. Other species communicate with sounds: crickets stridulate, or rub their wings together, to attract a mate and repel other males. Lampyridae in the beetle order Coleoptera communicate with light. Humans regard certain insects as pests and attempt to control them using insecticides and a host of other techniques. Some insects damage crops by feeding on sap, leaves or fruits, a few bite humans and livestock, alive and dead, to feed on blood and some are capable of transmitting diseases to humans, pets and livestock. Many other insects are considered ecologically beneficial and a few provide direct economic benefit. Silkworms and bees have been domesticated by humans for the production of silk and honey, respectively. Vectors are organisms that can introduce a pathogen such as a bacterium or virus into a plant to cause aninfection. Insects, mites, and nematode vectors focusthe movement of plant pathogens among immobileplants. Many insects or other arthropods may containplant pathogens but cannot transmit these to plants and thus are not vectors. Some of our most important plant diseases require mobile vectors. Almost all plant viruses and all wall-free, plant pathogenic bacteria known as mollicutes have recognized or suspected vectors. Arthropods form a major group of disease vectors with mosquitoes, flies, sand flies , lice, fleas, ticks and mites transmitting a huge number of diseases. Many such vectors are haematophagous, which feed on blood at some or all stages of their lives. When the insects blood feed, the parasite enters the blood stream of the host. This can happen in different ways. The Anopheles mosquito, a vector for Malaria, Filariasis and various arthropodborne-viruses (arboviruses), inserts its delicate mouthpart under the skin and feeds on its host's blood. The parasites the mosquito carries are usually located in its salivary glands (used by mosquitoes to anaesthetise the host). Therefore, the parasites are transmitted directly into the host's blood stream. Pool feeders such as the sand fly and black fly, vectors for Leishmaniasis and Onchocerciasis respectively, will chew a well in the host's skin, forming a small pool of blood from which they feed. Leishmania parasites then infect the host through the saliva of the sand fly. Onchocerca force their own way out of the insect's head into the pool of blood. Triatomine bugs are responsible for the transmission of a trypanosome , Trypanosoma cruzi, which causes Chagas' Disease. The Triatomine bugs defecate during feeding and the excrement contains the parasites which are accidentally smeared into the open wound by the host responding to pain and irritation from the bite.
91. Class Insecta (insects). Pediculus hominis (Pediculus humanus). Phthirius pubis.
Cimex lectularius. Pulex irritans Sucking lice (Anoplura, formerly known as Siphunculata) have around 500 species and represent the smaller of the two traditional suborders of lice. As opposed to the paraphyletic chewing lice, which are now divided among three suborders, the sucking lice are monophyletic.
Notes made by kozi1989 email:kozi1989@yahoo.gr
202
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The Anoplura are all blood-feeding ectoparasites of mammals. They can cause localised skin irritations and are vectors of several blood-borne diseases. Children appear particularly susceptible to attracting lice, possibly due to their fine hair. At least three species of Anoplura are parasites of humans; the human condition of being infested with sucking lice is called pediculosis. Pediculus humanus is divided into two subspecies, Pediculus humanus humanus, or the body louse, sometimes nicknamed "the seam squirrel" for its habit of laying of eggs in the seams of clothing, and Pediculus humanus capitis, or the head louse. Phthirus pubis (the crab louse) is the cause of the condition known as crabs. The body louse (Pediculus humanus humanus, sometimes called Pediculus humanus corporis)[1] is a louse which infests humans. The condition of being infested with head lice, body lice, or pubic lice is known as pediculosis. Pediculus humanus humanus (the body louse) is indistinguishable in appearance from Pediculus humanus capitis (the head louse) and under laboratory conditions they will interbreed. In their natural state, however, the two subspecies do not interbreed and occupy different habitats. In particular, body lice have evolved to attach their eggs to clothes, whereas head lice attach their eggs to the base of hairs. Body lice frequently lay their eggs on or near the seams of clothing. Body lice must feed on blood and usually only move to the skin to feed. Body lice exist worldwide and infest people of all races. Body lice infestations can spread rapidly under crowded living conditions where hygiene is poor (homeless, refugees, victims of war or natural disasters). In the United States, body lice infestations are rare, typically found mainly in homeless transient populations who do not have access to bathing and regular changes of clean clothes. Infestation is unlikely to persist on anyone who bathes regularly and who has at least weekly access to freshly laundered clothing and bedding. Body lice are spread through prolonged direct physical contact with a person who has body lice or through contact with articles such as clothing, beds, bed linens, or towels that have been in contact with an infested person. Body lice are a nuisance in themselves and cause intense itching. They are however, also vectors (transmitters) of other diseases. Body lice can spread epidemic typhus, trench fever, and louse-borne relapsing fever. Although louse-borne (epidemic) typhus is no longer widespread, outbreaks of this disease still occur during times of war, civil unrest, natural or man-made disasters, and in prisons where people live together in unsanitary conditions. Louse-borne typhus still exists in places where climate, chronic poverty, and social customs or war and social upheaval prevent regular changes and laundering of clothing. Life stages Body lice have three forms: the egg (also called a nit), the nymph, and the adult. 1) Nits are lice eggs. They are generally easy to see in the seams of an infested persons clothing, particularly around the waistline and under armpits. Body lice
Notes made by kozi1989 email:kozi1989@yahoo.gr
203
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
nits occasionally also may be attached to body hair. They are oval and usually yellow to white in color. Body lice nits may take 12 weeks to hatch. 2) A nymph is an immature louse that hatches from the nit (egg). A nymph looks like an adult body louse, but is smaller. Nymphs mature into adults about 9 12 days after hatching. To live, the nymph must feed on blood. 3) The adult body louse is about the size of a sesame seed , has 6 legs, and is tan to greyish-white. Females lay eggs. To live, lice must feed on blood. If a louse is separated from its person, it dies at room temperature. Crab lice (also known as "crabs", Phthirus pubis, Pthirus pubis, and "pubic lice"[1]) are parasitic insects notorious for infesting human genitals. The species may also live on other areas with hair, including the eyelashes. They feed exclusively on blood. Humans are the only known hosts of this parasite, although a closely related species, Pthirus gorillae, infects gorilla populations.[2] Infestation with pubic lice is called phthiriasis pubis, while infestation of eyelashes with pubic lice is called phthiriasis palpebrarum.[3] Pubic lice usually infect a new host only by close contact between individuals, usually through sexual intercourse. Parent-to-child infestations are more likely to occur through routes of shared towels, clothing, beds or closets. Adults are more frequently infested than children. As with most sexually transmitted pathogens, they can only survive a short time away from the warmth and humidity of the human body. Pubic lice are primarily spread through sweat, body contact or sexual contact. Therefore, all partners with whom the patient has had sexual contact within the previous 30 days should be evaluated and treated, and sexual contact should be avoided until all partners have successfully completed treatment and are thought to be cured. Because of the strong association between the presence of pubic lice and classic sexually transmitted infections (STIs), patients diagnosed with pubic lice should undergo evaluation for other STIs. Cimicidae or bed bugs (sometimes bedbugs), are small parasitic insects. The most common type is Cimex lectularius.[2] The term usually refers to species that prefer to feed on human blood. All insects in this family live by feeding exclusively on the blood of warmblooded animals. Adult bed bugs are reddish-brown, flattened, oval and wingless. Bed bugs have microscopic hairs that give them a banded appearance. Adults grow to 45 mm in length and 1.53 mm wide. Newly hatched nymphs are translucent, lighter in color and become browner as they moult and reach maturity. Bed bugs may be mistaken for other insects such as booklice and carpet beetles, or vice-versa. Bed bugs use pheromones and kairomones to communicate regarding nesting locations, feeding and reproduction. The life span of bed bugs varies by species and is also dependent on feeding. Bed bugs can survive a wide range of temperatures and atmospheric compositions. Below 16.1 C (61.0 F), adults enter semi-hibernation and can survive longer.[10] Bed bugs
Notes made by kozi1989 email:kozi1989@yahoo.gr
204
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
can survive for at least five days at 10 C (14.0 F) but will die after 15 minutes of exposure to 32 C (26 F).*11+ They show high desiccation tolerance, surviving low humidity and a 3540 C range even with loss of one-third of body weight; earlier life stages are more susceptible to drying out than later ones.[12] The thermal death point for C. lectularius is high: 45 C (113 F), and all stages of life are killed by 7 minutes of exposure to 46 C (115 F).*11+ Bed bugs apparently cannot survive high concentrations of carbon dioxide for very long; exposure to nearly pure nitrogen atmospheres, however, appears to have relatively little effect even after 72 hours. Bed bugs are obligatory hematophagous (bloodsucking) insects. Most species feed on humans only when other prey are unavailable.[14][15][16] Bed bugs are attracted to their hosts primarily by carbon dioxide, secondarily by warmth, and also by certain chemicals.[17]
A bed bug pierces the skin of its host with two hollow feeding tubes shaped like tongues. With one tube it injects its saliva, which contains anticoagulants and anesthetics, while with the other it withdraws the blood of its host. After feeding for about five minutes, the bug returns to its hiding place.[4] It takes between five to ten minutes for a bed bug to become completely engorged with blood. Although bed bugs can live for a year without feeding,[19] they normally try to feed every five to ten days. In cold weather, bed bugs can live for about a year; at temperatures more conducive to activity and feeding, about 5 months. The human flea, Pulex irritans, is a cosmopolitan flea species that has, in spite of the common name, a wide host spectrum. It is one of six species in the genus Pulex; the other five are all confined to the Nearctic and Neotropical regions.[1] The species is thought to have originated in South America, where its original host may have been the guinea pig or peccary.[2] This species bites many species of mammals and birds, including domesticated ones. It has been found on dogs and wild canids, monkeys in captivity, opossums, domestic cats, wild felids in captivity, chickens, black rats and Norwegian rats, wild rodents, pigs, free-tailed bats, and other species. It can also be an intermediate host for the cestode, Dipylidium caninum.
92. Genus Culex and genus Anopheles-morphological differences. Phlebotomus
papatasii Culex is a genus of mosquito, and is important in that several species serve as vectors of important diseases, such as West Nile virus, filariasis, Japanese encephalitis, St. Louis encephalitis and avian malaria. The adult mosquito can measure from 410 millimetres (0.160.39 in), and morphologically has the three body parts common to insects: head, thorax, and abdomen. As a fly, it has one pair of wings.
Notes made by kozi1989 email:kozi1989@yahoo.gr
205
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Life cycle The developmental cycle takes two weeks and is by complete metamorphosis. Eggs are laid singularly or in batches, depending on the species. Eggs will only hatch in the presence of water. During the larval stage the mosquito lives in water and feeds on organic matter and plants, then develops into a pupa. The pupa is comma-shaped and also lives in water. It does not feed and becomes an adult after one or two days. Anopheles, pronounced /nfliz/,*1+ is a genus of mosquito. There are approximately 460 recognized species: while over 100 can transmit human malaria, only 30 40 commonly transmit parasites of the genus Plasmodium, which cause malaria in humans in endemic areas. Anopheles gambiae is one of the best known, because of its predominant role in the transmission of the most dangerous malaria parasite species Plasmodium falciparum. The name comes from the Greek , an, meaning not, and , pheles, meaning profit, and translates to useless.[1]Some species of Anopheles also can serve as the vectors for canine heartworm Dirofilaria immitis, the Filariidae Wuchereria bancrofti and Brugia malayi, and viruses such as one that causes O'nyong'nyong fever. There is an association of brain tumor incidence and malaria, suggesting that the Anopheles might transmit a virus or other agent that could cause a brain tumor. Mosquitoes in other genera (Aedes, Culex) can also serve as vectors of disease agents. Like all mosquitoes, anophelines go through four stages in their life cycle: egg, larva, pupa, and imago. The first three stages are aquatic and last 514 days, depending on the species and the ambient temperature. The adult stage is when the female Anopheles mosquito acts as malaria vector. The adult females can live up to a month (or more in captivity) but most probably do not live more than 12 weeks in nature. Eggs Adult females lay 50200 eggs per oviposition. The eggs are quite small (~0.5 0.2 mm). Eggs are laid singly and directly on water. They are unique in that they have floats on either side. Eggs are not resistant to drying and hatch within 2 3 days, although hatching may take up to 23 weeks in colder climates. Larvae Mosquito larvae have a well-developed head with mouth brushes used for feeding, a large thorax and a nine segmented abdomen. They have no legs. In contrast to other mosquitoes, Anopheles larvae lack a respiratory siphon and for this reason position themselves so that their body is parallel to the surface of the water.Larvae breathe through spiracles located on the 8th abdominal segment and therefore must come to the surface frequently. The larvae spend most of their time feeding on algae, bacteria, and other microorganisms in the surface microlayer. They dive below the surface only when disturbed.
Notes made by kozi1989 email:kozi1989@yahoo.gr
206
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Larvae swim either by jerky movements of the entire body or through propulsion with the mouth brushes. Larvae develop through 4 stages, or instars, after which they metamorphose into pupae. At the end of each instar, the larvae molt, shedding their exoskeleton, or skin, to allow for further growth. 1st stage larvae are ~1 mm in length; 4th stage larvae are normally 58 mm in length. The process from egg laying to emergence of the adult is temperature dependent, with a minimum time of 7 days. The larvae occur in a wide range of habitats but most species prefer clean, unpolluted water. Larvae of Anopheles mosquitoes have been found in fresh- or salt-water marshes, mangrove swamps, rice fields, grassy ditches, the edges of streams and rivers, and small, temporary rain pools. Many species prefer habitats with vegetation. Others prefer habitats that have none. Some breed in open, sun-lit pools while others are found only in shaded breeding sites in forests. A few species breed in tree holes or the leaf axils of some plants. Pupae The pupa is comma-shaped when viewed from the side. The head and thorax are merged into a cephalothorax with the abdomen curving around underneath. As with the larvae, pupae must come to the surface frequently to breathe, which they do through a pair of respiratory trumpets on the cephalothorax. After a few days as a pupa, the dorsal surface of the cephalothorax splits and the adult mosquito emerges. Adults The duration from egg to adult varies considerably among species and is strongly influenced by ambient temperature. Mosquitoes can develop from egg to adult in as little as 5 days but usually take 1014 days in tropical conditions. Like all mosquitoes, adult Anopheles have slender bodies with 3 sections: head, thorax and abdomen. The head is specialized for acquiring sensory information and for feeding. The head contains the eyes and a pair of long, many-segmented antennae. The antennae are important for detecting host odors as well as odors of breeding sites where females lay eggs. The head also has an elongated, forward-projecting proboscis used for feeding, and two sensory palps. The thorax is specialized for locomotion. Three pairs of legs and a pair of wings are attached to the thorax. The abdomen is specialized for food digestion and egg development. This segmented body part expands considerably when a female takes a blood meal. The blood is digested over time serving as a source of protein for the production of eggs, which gradually fill the abdomen. Anopheles mosquitoes can be distinguished from other mosquitoes by the palps,
Notes made by kozi1989 email:kozi1989@yahoo.gr
207
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
which are as long as the proboscis, and by the presence of discrete blocks of black and white scales on the wings. Adult Anopheles can also be identified by their typical resting position: males and females rest with their abdomens sticking up in the air rather than parallel to the surface on which they are resting. Human malaria is transmitted only by females of anopheles Phlebotomus is a genus of "sand fly" in the Dipteran family Psychodidae. In the past, they have sometimes been considered to belong in a separate family, Phlebotomidae, but this alternative classification has not gained wide acceptance. Adults are small sized about 1.53 mm, yellowish in colour with conspicuous black eyes, hairy body, wings and legs. The oval lanceolate wings are carried erect on the humped thorax. Males possess long prominent genital terminalia known as claspers. Females have a pair of anal recti. Life cycle Females lay eggs in batches in cracks and crevices which are dark and rich in organic matter. They are small, elliptical and brownish in colour. Hatching occurs into the larval stage. Larvae are small whitish in colour with a black head capsule. Characteristic of this species is a pair of long, dark caudal bristles which arise from the posterior end. It is the cause of pappataci fever. Its isacute, infectious, febrile disease caused by a phlebovirus (family Bunyaviridae) and producing temporary incapacitation. It is transmitted to humans by the bloodsucking female sand fly (notably Phlebotomus papatasii, P. perniciosus, and P. perfiliewsi) and is prevalent in the moist subtropical countries. Comparative anatomy of vertebrates
93. Comparative anatomy of skin. Epidermis and glands. Dermis
The corium, or dermis, contains down growths from the epidermis, such as hair follicles and glands. It presents a superficial papillary layer of loose collagenous and elastic fibers, together with fibroblasts, mast cells, and macrophages. Elevations (papillae) project toward the epidermis. The thicker, deep reticular layer of the dermis consists of dense, coarse bundles of collagenous fibers. Some of the fibers enter the subcutaneous tissue, where they form bundles between lobules of fat. Smooth muscle is found in some regions (areola and nipple, scrotum and penis, and perineum). In some areas, muscle fibers of skeletal type (e.g., platysma) may be inserted into the skin. In tattooing, foreign particles, such as carbon, are introduced into the dermis. The skin lies on the subcutaneous tissue ("superficial fascia"), a layer of fatty areolar tissue that overlies the more densely fibrous fascia. It should be remembered that fat is liquid, or nearly so, at body temperature. The subcutaneous tissue serves as a depot for fat storage and aids in preventing loss of heat. When a pinch of skin is picked up, subcutaneous tissue is included. A hypodermic injection is one given into the subcutaneous
Notes made by kozi1989 email:kozi1989@yahoo.gr
208
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
tissue. The cells covering the embryo after neurulation form the presumptive epidermis (Dermis). Originally, this tissue is one cell layer thick, but in most vertebrates it shortly becomes a two-layered structure. The outer layer gives rise to the periderm, a temporary covering that is shed once the inner layer differentiates to form a true epidermis. The inner layer, called the basal layer (or stratum germinativum), is a germinal epithelium that gives rise to all the cells of the epidermis. The basal layer divides to give rise to another, outer population of cells that constitutes the spinous layer. These two epidermal layers together are referred to as the Malpighian layer. The cells of the Malpighian layer divide to produce the granular layer of the epidermis, so called because its cells are characterized by granules of the protein keratin. Unlike the cells remaining in the Malpighian layer, the cells of the granular layer do not divide, but begin to differentiate into epidermal skin cells, the keratinocytes. The keratin granules become more prominent as the keratinocytes of the granular layer age and migrate outward to form the cornified layer (stratum corneum) These cells become flattened sacs of keratin protein, and their nuclei are pushed to one edge of the cell.. The depth of the cornified layer varies from site to site, but it is usually 10 to 30 cells thick. Shortly after birth, the outer cells of the cornified layer are shed and are replaced by new cells coming up from the granular layer. Throughout life, the dead keratinized cells of the cornified layer are shed (humans lose about 1.5 grams of these cells each day*) and are replaced by new cells, the source of which is the mitotic cells of the Malpighian layer. Pigment cells (melanocytes) from the neural crest also reside in the Malpighian layer, where they transfer their pigment sacs (melanosomes) to the developing keratinocytes. The epidermal stem cells of the Malpighian layer are bound to the basal lamina by their integrin proteins. However, as these cells become committed to differentiate, they down-regulate their integrins and eventually lose them as they migrate into the spinous layer (Jones and Watt 1993). The skin is covered by a film of emulsified material produced by glands and by cornification. The epidermis is an avascular layer of stratified squamous epithelium that is thickest on the palms and soles. The epidermis, where it is thick, presents five layers, as listed in table 4-1. In the outer layers, which may conveniently be grouped as the horny zone, the cells become converted into soft-keratin flakes that are worn away from the surface continuously. The stratum corneum is a tough, resilient, semitransparent cellular membrane that acts as a barrier to water transfer. Under normal conditions, mitotic figures are practically confined to the deepest layer, the stratum basale, which is, therefore, the normal germinative layer of the epidermis. The various layers show the stages through which the basal cells pass before their keratinization and shedding. The cells of the epidermis are replaced approximately once per month. Keratin is a protein that is present throughout the epidermis, perhaps in a modified form. It is readily hydrated-hence the swelling of skin on immersion in water-and dryness of the skin is due chiefly to a lack of
Notes made by kozi1989 email:kozi1989@yahoo.gr
209
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
water. Several growth factors stimulate the development of the epidermis. One of these is transforming growth factor. TGF is made by the basal cells and stimulates their own division. When a growth factor is made by the same cell that receives it, that factor is called an autocrine growth factor. Such factors have to be carefully regulated because if their levels are elevated, more cells are rapidly produced. In adult skin, a cell born in the Malpighian layer takes roughly 8 weeks to reach the cornified layer, and remains there for about 2 weeks. In individuals with psoriasis, a disease characterized by the exfoliation of enormous amounts of epidermal cells, a cell's time in the cornified layer is only 2 days (Weinstein and van Scott 1965; Halprin 1972). This condition has been linked to the overexpression of TGF(which occurs secondarily to an inflammation) (Elder et al. 1989). Another growth factor needed for epidermal production is keratinocyte growth factor (KGF; also known as fibroblast growth factor 7), a paracrine factor that is produced by the fibroblasts of the underlying (mesodermally derived) dermis. KGF is received by the basal cells of the epidermis and is thought to regulate their proliferation. The basal cells are "forced" into the epidermal pathway of differentiation. The alternative pathway for basal cells leads to the generation of hair follicles. The epidermis and dermis also interact at specific sites to create the sweat glands and the cutaneous appendages: hairs, scales, or feathers (depending on the species). In mammals, the first indication that a hair follicle primordium, or hair germ, will form at a particular place is an aggregation of cells in the basal layer of the epidermis. This aggregation is directed by the underlying dermal fibroblast cells and occurs at different times and different places in the embryo. It is probable that the dermal signals cause the stabilization of -catenin in the ectoderm (Gat et al. 1998). The basal cells elongate, divide, and sink into the dermis. The dermal fibroblasts respond to this ingression of epidermal cells by forming a small node (the dermal papilla) beneath the hair germ. The dermal papilla then pushes up on the basal stem cells and stimulates them to divide more rapidly. The basal cells respond by producing postmitotic cells that will differentiate into the keratinized hair shaft (see Hardy 1992; Miller et al. 1993). Melanoblasts, which were present among the epidermal cells as they ingressed, differentiate into melanocytes and transfer their pigment to the shaft (Figure 12.34). It is obvious that cutaneous appendages such as hair, feathers, or scales do not grow randomly over the body. Rather, there are spaces between them, and these spaces (for instance, on the scalp) are very similar from region to region (Widelitz and Chuong 1998). Recent research suggests that a reaction-diffusion process may be responsible for this pattern (see Chapter 1). The activator is Sonic hedgehog, a paracrine factor that acts locally without much diffusion (Nohno et al. 1995). The inhibitor is believed to be BMP4 or BMP2, both of which are paracrine factors with a greater range of diffusion (Jung et al. 1998; Noramly and Morgan 1998). BMPs may prevent the dermal fibroblasts from aggregating, while Sonic hedgehog may support the formation and retention of the dermal papilla
Notes made by kozi1989 email:kozi1989@yahoo.gr
210
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
94. Horny, structures formed by the epidermis. Bony scales and plates
Hairs Hairs (or pili; pilus in the singular) are characteristic of mammals. The functions of hair include protection, regulation of body temperature, and facilitation of evaporation of perspiration; hairs also act as sense organs. Hairs develop in the fetus as epidermal downgrowths that invade the underlying dermis. Each downgrowth terminates in an expanded end that becomes invaginated by a mesodermal papilla. The central cells of the downgrowth become keratinized to form a hair, which then grows outward to reach the surface. The hairs first developed constitute the lanugo, or down, which is shed shortly before birth. The fine hairs that develop later constitute the vellus. Although hairs on many portions of the human body are inconspicuous, their actual number per unit area is large. In a few places (such as the palms and the soles and the dorsal aspect of the distal phalanges) the skin is glabrous, that is, devoid of hair. The shaft of a hair consists of a cuticle and a cortex of hard-keratin surrounding, in many hairs, a soft-keratin medulla (fig. 4-2). Pigmented hairs contain melanin in the cortex and medulla, but pigment is absent from the surrounding sheaths. The color of hair depends mainly on the shade and the amount of pigment in the cortex and, to a lesser extent, on air spaces in the hair. In white hairs pig ment is absent from the cortex, and the contained air is responsible for the whiteness; "gray hair" is generally a mixture of white and colored hairs. The root of a hair is situated in an epidermal tube known as the hair follicle, sunken into either the dermis or the subcutaneous tissue. The follicle is dilated at its base to form the bulb (matrix). In the obtuse angle between the root of a hair and the surface of the skin, a bundle of smooth muscle fibers, known as an arrector pili muscle, is usually found. It extends from the deep part of the hair follicle to the papillary layer of the dermis. On contraction it makes the hair erect. The arrectores pilorum are innervated by sympathetic fibers and contract in response to emotion or cold. This results in an unevenness of the surface called "goose pimples" or "goose skin." Nails The nails (or ungues; unguis in the singular) are hardenings of the horny zone of the epidermis. They overlie the dorsal aspect of the distal phalanges (fig. 4-3). They protect the sensitive tips of the digits and, in the fingers, serve in scratching. Nails develop in the fetus as epidermal thickenings that undercut the skin to form folds from which the horny substance of the nail grows distally. The horny zone of the nail is composed of hard-keratin and has a distal, exposed part, or body, and a proximal, hidden portion, or root. The root is covered by a distalward prolongation of the stratum corneum of the skin. This narrow fold is composed of soft-keratin and is termed the eponychium. Distal to the eponychium is the "half-moon," or lunula, a part of the horny zone that is opaque to the underlying capillaries.
Notes made by kozi1989 email:kozi1989@yahoo.gr
211
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Deep to the distal or free border of the nail, the horny zone of the fingertip is thickened and is frequently termed the hyponychium. The horny zone of the nail is attached to the underlying nail bed. The matrix, or proximal part of the bed, produces hard-keratin. Further distally, however, the bed may also generate nail substance. Moreover, the most superficiallayer of the nail may be produced by the epithelium immediately dorsal to the root and proximal to the eponychium. The growth of the nail is affected by nutrition, hormones, and disease. Nail growth involves considerable protein synthesis, as a result of which nonspecific changes occur in the nails in response to various local and systemic disturbances. White spots indicate incomplete keratinization.
95. Origins and evolution of vertebral column
In human anatomy, the vertebral column (Latin Columna vertebralis) (backbone or spine) is a column usually consisting of 24 articulating vertebrae ,[1] and 9 fused vertebrae in the sacrum and the coccyx. It is situated in the dorsal aspect of the torso, separated by intervertebral discs. It houses and protects the spinal cord in its spinal canal. The striking segmented pattern of the human spine is established during embryogenesis when the precursor of the vertebrae, the somites, are rhythmically added to the forming posterior part of the embryo. In humans, somite formation begins around the third week post-fertilization and continues until a total of around 52 somites are formed. The somites are epithelial spheres that contain the precursors of the vertebrae, the ribs, the skeletal muscles of the body wall and limbs, and the dermis of the back. The periodicity of somite distribution and production is thought to be imposed by a molecular oscillator or clock acting in cells of the presomitic mesoderm (PSM). Somites form soon after the beginning of gastrulation, on both sides of the neural tube from a tissue called the presomitic mesoderm (PSM). The PSM is part of the paraxial mesoderm and is generated caudally by gastrulation when cells ingress through the primitive streak, and later, through the tail bud. Soon after their formation, somites become subdivided into the dermomyotome dorsally, which gives rise to the muscles and dermis, and the sclerotome ventrally, which will form the spine components. Sclerotomes become subvidided into an anterior and a posterior compartment. This subdivision plays a key role in the definitive patterning of vertebrae that form when the posterior part of one somite fuses to the anterior part of the consecutive somite during a process termed resegmentation. Disruption of the somitogenesis process in humans results in diseases such as congenital scoliosis. So far, the human homologues of three genes associated to the mouse segmentation clock (MESP2, DLL3 and LFNG) have been shown to be mutated in human patients with human congenital scoliosis suggesting that the mechanisms involved in vertebral segmentation are conserved across vertebrates. In humans the first four somites are incorporated in the basioccipital bone of the skull and the next 33 somites will form the vertebrae. The remaining posterior somites degenerate. During the fourth week of embryonic development, the sclerotomes shift their position to surround the spinal cord and the notochord. The sclerotome is made of mesoderm and originates from the ventromedial part of the somites.
Notes made by kozi1989 email:kozi1989@yahoo.gr
212
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
This column of tissue has a segmented appearance, with alternating areas of dense and less dense areas. The notochord, which constitutes the earliest structure that stiffens the embryo, appeared in animals before the true vertebral column evolved. A vertebra includes a centrum and a neural arch surrounding the spinal cord. Lower chordates and fishes Possession of the notochord is what distinguishes members of the most-advanced phylum, Chordata. In the sea squirts (Urochordata), the notochord is present in the tail region of the larva but disappears after the animal transforms into the adult.
96. Origins and evolution of skull. Cartilaginous cerebral skull. Bony cerebral skull
The skull, the skeleton of the head, is the most complex bony structure in the body because it: Encloses the brain, which is irregular in shape; Houses the organs of special senses for seeing, hearing, tasting, and smelling; and Surrounds the openings in to the digestive and respiratory tracts. In the anatomical position, the skull is oriented so that the inferior margin of the orbit (eye socket) and the superior margin of the external acoustic meatus (auditory canal) are horizontal. This is called the orbitomenial plane (Frankfort plane). The term cranium (L. skull) is sometimes used when referring to the skull without the mandible (lower jaw), but the cranium is often used when referring to the part of the skull containing the brain. The superior part is the box-like structure called the calvaria (cranial vault, brain case); the remainder of the cranium, including the maxilla (upper jaw), orbits (eyeball sockets) and nasal cavities, forms the facial skeleton. The term skullcap (calotte) refers to the superior part of the calvaria, which is removed during autopsies and dissections. The inferior aspect of the cranium is called the cranial base.
97. Origins and evolution of skull. Cartilaginous visceral skull. Bony visceral skull
The skeleton of vertebrates is formed not only the bones: it includes the cartilage and connective tissue, and sometimes it consists of various skin formation. In vertebrates is assumed to specify the axial skeleton (skull, chord, spine, ribs) and the skeleton of the limbs, including their belt (shoulder and pelvic) and free sections. The skull (cranium) - the skeleton of the head of vertebrates. There are brain-skull (cranium celebrale, s. neurocranium) and visceral (cranium viscerale, s. splanchnocranium). Visceral elements of the skull - derivatives of cartilaginous gill arches that have
Notes made by kozi1989 email:kozi1989@yahoo.gr
213
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
arisen in the walls of the pharynx in the development of vertebrate gills. The fish the first two arcs have changed and turned into the mandibular and hypoglossal apparatus. In typical cases, they have saved an additional 5 gill arches, but in some genera the number fell. In primitive modern sharks semizhabernika (Heptanchus) behind the jaw and hyoid arches gill arches seven. In bony fishes jaw cartilages are faced with numerous investing bone, recent form and gill covers that protect the delicate gill lobes. In the course of the evolution of vertebrate jaws of the original cartilage were reduced steadily until it disappeared completely. If the crocodiles of the initial balance of cartilage in the mandible veneer 5 pairs covering the bones, the mammals of them is only one - a tooth, which forms the skeleton of the full lower jaw. The skull of ancient amphibians contain heavy Cover the plate and was similar in this respect, a typical skull Crossopterygii. In modern amphibians as overhead, and replacing the bone greatly reduced. In the skull of frogs and salamanders are fewer than in other vertebrates with bony skeleton, and in the latter group, many elements remain cartilaginous. Turtles and crocodiles are numerous bones of the skull and tightly adherent to each other. In lizards and snakes are relatively small, with exterior elements are separated by wide intervals, like frogs or toads. Snakes right and left branches of the lower jaw very loosely connected to each other and with the skull elastic ligaments, allowing reptiles to swallow relatively large prey. In birds, the skull bones are thin, but very solid, with adults they have grown together so completely that a few stitches disappeared. The orbital cavity is very high, relative to the huge roof brain box is formed by thin investing bone, lungs jaws are covered with horny covers. In mammals, the skull is heavy and includes powerful jaws with teeth. The remains of cartilaginous jaws moved into the middle ear, and formed his bones
98. Comparative anatomy of skull
The skull is a bony structure in the head of many animals that supports the structures of the face and forms a cavity for the brain. The skull is composed of two parts: the cranium and the mandible. A skull without a mandible is only a cranium. Animals that have skulls are called craniates. The skull is a part of the skeleton. Functions of the skull include protection of the brain, fixing the distance between the eyes to allow stereoscopic vision, and fixing the position of the ears to help the brain use auditory cues to judge direction and distance of sounds. In some animals, the skull also has a defensive function (e.g. horned ungulates); the frontal bone is where horns are mounted. The English word "Skull" is probably derived from Old Norse "skalli" meaning bald, while the word cranium comes from the Greek root (kranion). Human skull In humans, the adult skull is normally made up of 22 bones. Except for the mandible, all of the bones of the skull are joined together by sutures, rigid articulations permitting very little movement. Eight bones including one frontal, two parietals, one occipital bone, one
Notes made by kozi1989 email:kozi1989@yahoo.gr
214
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
sphenoid, two temporals and one ethmoid form the neurocranium (braincase), a protective vault surrounding the brain. Fourteen bones form the splanchnocranium, the bones supporting the face. Encased within the temporal bones are the six ear ossicles of the middle ears, though these are not part of the skull. The hyoid bone, supporting the tongue, is usually not considered as part of the skull either, as it does not articulate with any other bones. The skull is a protector of the brain. Tetrapod skulls The skull of the earliest tetrapods closely resembles that of their ancestors amongst the lobe-finned fishes. The skull roof is formed of a series of plate-like bones, including the maxilla, frontals, parietals, and lacrimals, among others. It is overlaying the endocranium, corresponding to the cartilaginous skull in sharks and rays. The various separate bones that compose the temporal bone of humans are also part of the skull roof series. A further plate composed of four pairs of bones forms the roof of the mouth; these include the vomer and palatine bones. The base of the cranium is formed from a ring of bones surrounding the foramen magnum and a median bone lying further forward; these are homologous with the occipital bone and parts of the sphenoid in mammals. Finally, the lower jaw is composed of multiple bones, only the most anterior of which (the dentary) is homologous with the mammalian mandible. In living tetrapods, a great many of the original bones have either disappeared, or fused into one another in various arrangements. In mammals and birds, in particular, there have been modifications of the skull to allow for the expansion of the brain. The fusion between the various bones is especially notable in birds, in which the individual structures may be difficult to identify . Living amphibians typically have greatly reduced skulls, with many of the bones either absent or wholly or partly replaced by cartilage.[1] Temporal fenestrae This Massospondylus skull shows the two temporal fenestrae typical of diapsids. The temporal fenestrae are anatomical features of the skulls of several types of amniotes, characterised by bilaterally symmetrical holes (fenestrae) in the temporal bone. Depending on the lineage of a given animal, two, one, or no pairs of temporal fenestrae may be present, above or below the postorbital and squamosal bones. The upper temporal fenestrae are also known as the supratemporal fenestrae, and the lower temporal fenestrae are also known as the infratemporal fenestrae. The presence and morphology of the temporal fenestra are critical for taxonomic classification of the synapsids, of which mammals are part. Physiological speculation associates it with a rise in metabolic rates and an increase in jaw musculature. The earlier amniotes of the Carboniferous did not have temporal fenestrae but two more advanced lines did: the Synapsids (mammal-like reptiles) and the Diapsids (most reptiles and later birds). As time progressed, diapsids' and synapsids' temporal fenestrae became more modified and larger to make stronger bites and more jaw
Notes made by kozi1989 email:kozi1989@yahoo.gr
215
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
muscles. Dinosaurs, which are sauropsids, have large advanced openings and their descendants, the birds, have temporal fenestrae which have been modified. Mammals, which are synapsids, possess no fenestral openings in the skull, as the trait has been modified. They do, though, still have the temporal orbit (which resembles an opening) and the temporal muscles. It is a hole in the head and is situated to the rear of the orbit behind the eye. Classification Chimpanzee skull There are four types of amniote skull, classified by the number and location of their fenestra. These are: Anapsida - no openings Synapsida - one low opening (beneath the postorbital and squamosal bones) Euryapsida - one high opening (above the postorbital and squamosal bones); euryapsids actually evolved from a diapsid configuration, losing their lower temporal fenestra. Diapsida - two openings
99. Origins and evolution of the spinal cord
The spinal cord is a long, thin, tubular bundle of nervous tissue and support cells that extends from the brain (the medulla oblongata specifically). The brain and spinal cord together make up the central nervous system. The spinal cord begins at the Occipital bone and extends down to the space between the first and second lumbar vertebrae; it does not extend the entire length of the vertebral column. It is around 45 cm (18 in) in men and around 43 cm (17 in) long in women. Also, the spinal cord has a varying width, ranging from 1/2 inch thick in the cervical and lumbar regions to 1/4 inch thick in the thoracic area. The enclosing bony vertebral column protects the relatively shorter spinal cord. The spinal cord functions primarily in the transmission of neural signals between the brain and the rest of the body but also contains neural circuits that can independently control numerous reflexes and central pattern generators. The spinal cord has three major functions: A. Serve as a conduit for motor information, which travels down the spinal cord. B. Serve as a conduit for sensory information, which travels up the spinal cord. C. Serve as a center for coordinating certain reflexes. The spinal cord is made from part of the neural tube during development. As the neural tube begins to develop, the notochord begins to secrete a factor known as Sonic hedgehog or SHH. As a result, the floor plate then also begins to secrete SHH, and this will induce the basal plate to develop motor neurons. Meanwhile, the overlying ectoderm secretes bone morphogenetic protein (BMP). This induces the roof plate to begin to secrete BMP, which will induce the alar plate to develop sensory neurons. The alar plate and the basal plate are separated by the sulcus limitans.
Notes made by kozi1989 email:kozi1989@yahoo.gr
216
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Additionally, the floor plate also secretes netrins. The netrins act as chemoattractants to decussation of pain and temperature sensory neurons in the alar plate across the anterior white commissure, where they then ascend towards the thalamus. Lastly, it is important to note that the past studies of Viktor Hamburger and Rita LeviMontalcini in the chick embryo have been further proven by more recent studies which demonstrated that the elimination of neuronal cells by programmed cell death (PCD) is necessary for the correct assembly of the nervous system. Overall, spontaneous embryonic activity has been shown to play a role in neuron and muscle development but is probably not involved in the initial formation of connections between spinal neurons. Secondary neurulation involves the making of a medullary cord and its subsequent hollowing into a neural tube. Knowledge of the mechanisms of secondary neurulation may be important in medicine, given the prevalence of human posterior spinal cord malformations. In frogs and chicks, secondary neurulation is usually seen in the neural tube of the lumbar (abdominal) and tail vertebrae. In both cases, it can be seen as a continuation of gastrulation. In the frog, instead of involuting into the embryo, the cells of the dorsal blastopore lip keep growing ventrally. 100. Origins and evolution of brain Telencephalon, diencephalon, mesencephalon, metencephalon, myelencephalon The differentiation of the neural tube into the various regions of the central nervous system occurs simultaneously in three different ways. On the gross anatomical level, the neural tube and its lumen bulge and constrict to form the chambers of the brain and the spinal cord. At the tissue level, the cell populations within the wall of the neural tube rearrange themselves to form the different functional regions of the brain and the spinal cord. Finally, on the cellular level, the neuroepithelial cells themselves differentiate into the numerous types of nerve cells (neurons) and supportive cells (glia) present in the body. The early development of most vertebrate brains is similar. The early mammalian neural tube is a straight structure. However, even before the posterior portion of the tube has formed, the most anterior portion of the tube is undergoing drastic changes. In this region, the neural tube balloons into three primary vesicles.
Notes made by kozi1989 email:kozi1989@yahoo.gr
217
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The prosencephalon becomes subdivided into the anterior telencephalon and the more caudal diencephalon. The telencephalon will eventually form the cerebral hemispheres, and the diencephalon will form the thalamic and hypothalamic brain regions that receive neural input from the retina. Indeed, the retina itself is a derivative of the diencephalon. The mesencephalon does not become subdivided, and its lumen eventually becomes the cerebral aqueduct. The rhombencephalon becomes subdivided into a posterior myelencephalon and a more anterior metencephalon. The myelencephalon eventually becomes the medulla oblongata, whose neurons generate the nerves that regulate respiratory, gastrointestinal, and cardiovascular movements. The metencephalon gives rise to the cerebellum, the part of the brain responsible for coordinating movements, posture, and balance. The rhombencephalon develops a segmental pattern that specifies the places where certain nerves originate. Periodic swellings called rhombomeres divide the rhombencephalon into smaller compartments. These rhombomeres represent separate developmental "territories" in that the cells within each rhombomere can mix freely within it, but not with cells from adjacent rhombomeres. Moreover, each rhombomere has a different developmental fate. Each rhombomere will form ganglia clusters of neuronal cell bodies whose axons form a nerve. 101. Skin sense organs of vertebrates. Evolution of the olfactory organ The olfactory system is the sensory system used for olfaction, or the sense of smell. Most mammals and reptiles have two distinct parts to their olfactory system: a main olfactory system and an accessory olfactory system. The main olfactory system detects volatile, airborne substances, while the accessory olfactory system senses fluid-phase stimuli. Behavioral evidence indicates that most often, the stimuli detected by the accessory olfactory system are awareness. The olfactory system is often spoken of along with the gustatory system as the
Notes made by kozi1989 email:kozi1989@yahoo.gr
218
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
chemosensory senses because both transduce chemical signals into perception. The mechanism of the olfactory system can be divided into a peripheral one, sensing an external stimulus and encoding it as an electric signal in neurons, and a central one, where all signals are integrated and processed in the central nervous system. Peripheral 1: Olfactory bulb 2: Mitral cells 3: Bone 4: Nasal Epithelium 5: Glomerulus 6: Olfactory receptor cells Amygdala location in each hemisphere of the human brain In mammals, the main olfactory system detects odorants that are inhaled through the nose, where they contact the main olfactory epithelium, which contains various olfactory receptors. These olfactory receptors are membrane proteins of bipolar olfactory receptor neurons in the olfactory epithelium. Rather than binding specific ligands like most receptors, olfactory receptors display affinity for a range of odor molecules. Olfactory neurons transduce receptor activation into electrical signals in neurons. The signals travel along the olfactory nerve, which belongs to the peripheral nervous system. This nerve terminates in the olfactory bulb, which belongs to the central nervous system. The complex set of olfactory receptors on different olfactory neurons can distinguish a new odor from the background environmental odors and determine the concentration of the odor. The general features of the olfactory system are remarkably consistent across vertebrates. A phylogenetic analysis of central olfactory projections indicates that at least three distinct olfactory subsystems may be broadly present in vertebrates and that a fourth, the accessory olfactory or vomeronasal system, arose in tetrapods. The origin and function of the vomeronasal system have been the subject of much controversy, but some conclusions can be drawn. The vomeronasal system did not arise as an adaptation to terrestrial life, as indicated by the presence of a vomeronasal system in modern aquatic amphibians and the increasing paleontological evidence that the last common ancestor of amphibians and amniotes was aquatic. The vomeronasal system is involved in both foraging and reproductive behaviors in reptiles and has been shown to be involved in some pheromonally mediated behaviors in mammals. However, among mammals, some pheromonal responses are not mediated by the vomeronasal system, and the possible involvement of the vomeronasal system in other type of behaviors has not yet been investigated. Thus, the relative functions of the olfactory and vomeronasal systems of tetrapods remain unclear. Other hypotheses that features of the olfactory system are specialized for aquatic chemoreception or for pheromone detection are similarly insupportable. For example, the suggestion that members of the olfactory receptor family can be separated into two groups that function for transduction of air-borne or water-borne odorants is contradicted by the presence of both groups in aquatic amphibians and by a phylogenetic analysis of the sequences for these genes. Interestingly, the putative odorant
Notes made by kozi1989 email:kozi1989@yahoo.gr
219
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
receptors from the vomeronasal epithelium share little sequence similarity with those from the olfactory epithelium, indicating that these receptors may have been independently coopted from the larger family of seven transmembrane domain receptors for use in odor transduction. A phylogenetic analysis of the distribution of olfactory receptor cell types indicates that microvillar olfactory receptor cells are widespread among vertebrates and are not restricted to aquatic animals or to the vomeronasal epithelium of tetrapods. Previous suggestions that all microvillar receptor cells are specialized for the detection of pheromones are not tenable. Attempts to recognize features of the olfactory system that are common to all vertebrates and might be specialized for the detection of pheromones vs. more general odorants, or for the detection of water-borne vs. air-borne odorants, are not supported by current evidence. 102. Comparative anatomy of ear as organ of balance and bearing The ear is the anatomical organ that detects sound. It not only acts as a receiver for sound, but also plays a major role in the sense of balance and body position. The ear is part of the auditory system. The word "ear" may be used correctly to describe the entire organ or just the visible portion. In most mammals, the visible ear is a flap of tissue that is also called the pinna and is the first of many steps in hearing. In people, the pinna is often called the auricle. Vertebrates have a pair of ears, placed symmetrically on opposite sides of the head. This arrangement aids in the ability to localize sound sources. The shape of outer ear of mammals varies widely across species. However the inner workings of mammalian ears (including humans') are very similar. Outer ear (pinna, ear canal, surface of ear drum) The outer ear is the most external portion of the ear. The outer ear includes the pinna (also called auricle), the ear canal, and the very most superficial layer of the ear drum (also called the tympanic membrane). In humans, and almost all vertebrates, the only visible portion of the ear is the outer ear. The word "ear" may properly refer to the pinna (the flesh covered cartilage appendage on either side of the head). This portion of the ear is very vital for hearing. The outer ear does help get sound (and imposes filtering), but the ear canal is very important. Unless the canal is open, hearing will be dampened. Ear wax (cerumen) is produced by glands in the skin of the outer portion of the ear canal. This outer ear canal skin is applied to cartilage; the thinner skin of the deep canal lies on the bone of the skull. Only the thicker cerumen-producing ear canal skin has hairs. The outer ear ends at the most superficial layer of the tympanic membrane. The tympanic membrane is commonly called the ear drum. The pinna helps direct sound through the ear canal to the tympanic membrane (eardrum). The framework of the auricle consists of a single piece of yellow fibrocartilage with a complicated relief on the anterior, concave side and a fairly smooth configuration on the
Notes made by kozi1989 email:kozi1989@yahoo.gr
220
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
posterior, convex side. The Darwinian tubercle, which is present in some people, lies in the descending part of the helix and corresponds to the true ear tip of the long-eared mammals. The lobule merely contains subcutaneous tissue.[2] In some animals with mobile pinnae (like the horse), each pinna can be aimed independently to better receive the sound. For these animals, the pinnae help localize the direction of the sound source. Human beings localize sound within the central nervous system, by comparing arrival-time differences and loudness from each ear, in brain circuits that are connected to both ears. This process is commonly referred to as EPS, or Echo Positioning System. Human outer ear and culture The auricles also have an effect on facial appearance. In Western societies, protruding ears (present in about 5% of ethnic Europeans) have been considered unattractive, particularly if asymmetric. The first surgery to reduce the projection of prominent ears was published in the medical literature in 1881. The ears have also been ornamented with jewelry for thousands of years, traditionally by piercing of the earlobe. In some cultures, ornaments are placed to stretch and enlarge the earlobes to make them very large. Tearing of the earlobe from the weight of heavy earrings, or from traumatic pull of an earring (for example by snagging on a sweater being removed), is fairly common.[3] The repair of such a tear is usually not difficult. A cosmetic surgical procedure to reduce the size or change the shape of the ear is called an otoplasty. In the rare cases when no pinna is formed (atresia), or is extremely small (microtia) reconstruction of the auricle is possible. Most often, a cartilage graft from another part of the body (generally, rib cartilage) is used to form the matrix of the ear, and skin grafts or rotation flaps are used to provide the covering skin. Recently ears have been grown on a rat's back and attached to human heads after. However, when babies are born without an auricle on one or both sides, or when the auricle is very tiny, the ear canal is ordinarily either small or absent, and the middle ear often has deformities. The initial medical intervention is aimed at assessing the baby's hearing and the condition of the ear canal, as well as the middle and inner ear. Depending on the results of tests, reconstruction of the outer ear is done in stages, with planning for any possible repairs of the rest of the ear.[4][5][6] Middle ear The middle ear, an air-filled cavity behind the ear drum (tympanic membrane), includes the three ear bones or ossicles: the malleus (or hammer), incus (or anvil), and stapes (or stirrup). The opening of the Eustachian tube is also within the middle ear. The malleus has a long process (the manubrium, or handle) that is attached to the mobile portion of the eardrum. The incus is the bridge between the malleus and stapes. The stapes is the smallest named bone in the human body. The three bones are arranged so that movement of the tympanic membrane causes movement of the malleus, which causes movement of the incus, which causes movement of the stapes. When the stapes footplate pushes on the oval window, it causes movement of fluid within the cochlea (a portion of the
Notes made by kozi1989 email:kozi1989@yahoo.gr
221
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
inner ear). In humans and other land animals the middle ear (like the ear canal) is normally filled with air. Unlike the open ear canal, however, the air of the middle ear is not in direct contact with the atmosphere outside the body. The Eustachian tube connects from the chamber of the middle ear to the back of the nasopharynx. The middle ear is very much like a specialized paranasal sinus, called the tympanic cavity; it, like the paranasal sinuses, is a hollow mucosalined cavity in the skull that is ventilated through the nose. The mastoid portion of the human temporal bone, which can be felt as a bump in the skull behind the pinna, also contains air, which is ventilated through the middle ear. Middle Ear Components of the middle ear Normally, the Eustachian tube is collapsed, but it gapes open both with swallowing and with positive pressure. When taking off in an airplane, the surrounding air pressure goes from higher (on the ground) to lower (in the sky). The air in the middle ear expands as the plane gains altitude, and pushes its way into the back of the nose and mouth. On the way down, the volume of air in the middle ear shrinks, and a slight vacuum is produced. Active opening of the Eustachian tube is required to equalize the pressure between the middle ear and the surrounding atmosphere as the plane descends. The diver also experiences this change in pressure, but with greater rates of pressure change; active opening of the Eustachian tube is required more frequently as the diver goes deeper into higher pressure. The arrangement of the tympanic membrane and ossicles works to efficiently couple the sound from the opening of the ear canal to the cochlea. There are several simple mechanisms that combine to increase the sound pressure. The first is the "hydraulic principle". The surface area of the tympanic membrane is many times that of the stapes footplate. Sound energy strikes the tympanic membrane and is concentrated to the smaller footplate. A second mechanism is the "lever principle". The dimensions of the articulating ear ossicles lead to an increase in the force applied to the stapes footplate compared with that applied to the malleus. A third mechanism channels the sound pressure to one end of the cochlea, and protects the other end from being struck by sound waves. In humans, this is called "round window protection", and will be more fully discussed in the next section. Abnormalities such as impacted ear wax (occlusion of the external ear canal), fixed or missing ossicles, or holes in the tympanic membrane generally produce conductive hearing loss. Conductive hearing loss may also result from middle ear inflammation causing fluid build-up in the normally air-filled space. Tympanoplasty is the general name of the operation to repair the middle ear's tympanic membrane and ossicles. Grafts from muscle fascia are ordinarily used to rebuild an intact ear drum. Sometimes artificial ear bones are placed to substitute for damaged ones, or a disrupted ossicular chain is rebuilt in order to conduct sound effectively. Inner Ear
Notes made by kozi1989 email:kozi1989@yahoo.gr
222
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Components of the inner ear The inner ear includes both the organ of hearing (the cochlea) and a sense organ that is attuned to the effects of both gravity and motion (labyrinth or vestibular apparatus). The balance portion of the inner ear consists of three semicircular canals and the vestibule. The inner ear is encased in the hardest bone of the body. Within this ivory hard bone, there are fluid-filled hollows. Within the cochlea are three fluid filled spaces: the scala tympani, the scala vestibuli and the scala media. The eighth cranial nerve comes from the brain stem to enter the inner ear. When sound strikes the ear drum, the movement is transferred to the footplate of the stapes, which presses it into one of its fluid-filled ducts through the oval window of cochlea . The fluid inside this duct is moved, flowing against the receptor cells of the Organ of Corti, which fire. These stimulate the spiral ganglion, which sends information through the auditory portion of the eighth cranial nerve to the brain. Hair cells are also the receptor cells involved in balance, although the hair cells of the auditory and vestibular systems of the ear are not identical. Vestibular hair cells are stimulated by movement of fluid in the semicircular canals and the utricle and saccule. Firing of vestibular hair cells stimulates the Vestibular portion of the eighth cranial nerve. 103. Evolution of the organ of vision - Structure of the mammalian eyeball. Embryonic development and accessory organs of the eye. Peculiarities of the eye in different classes of vertebrates. An individual gains knowledge of its environment through its sensory organs. The major sensory organs of the head develop from the interactions of the neural tube with a series of epidermal thickenings called the cranial ectodermal placodes. The most anterior of these are the two olfactory placodes that form the ganglia for the olfactory nerves, which are responsible for the sense of smell. The auditory placodes similarly invaginate to form the inner ear labyrinth, whose neurons form the acoustic ganglion, which enables us to hear. In this section, we will focus on the eye. The dynamics of optic development At gastrulation, the involuting endoderm and mesoderm interact with the adjacent prospective head ectoderm to give the head ectoderm a lens-forming bias (Saha et al. 1989). But not all parts of the head ectoderm eventually form lenses, and the lens must have a precise spatial relationship with the retina. The activation of the head ectoderm's latent lens-forming ability and the positioning of the lens in relation to the retina is accomplished by the optic vesicle. It extends from the diencephalon, and when it meets the head ectoderm, it induces the formation of a lens placode, which then invaginates to form the lens. The optic vesicle becomes the two-walled optic cup, whose two layers differentiate in different directions. The cells of the outer layer produce melanin pigment (being one of the few tissues other than the neural crest cells that can form this pigment) and ultimately become the pigmented retina. The cells of the inner layer proliferate rapidly and generate a
Notes made by kozi1989 email:kozi1989@yahoo.gr
223
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
variety of glia, ganglion cells, interneurons, and light-sensitive photoreceptor neurons. Collectively, these cells constitute the neural retina. The retinal ganglion cells are neurons whose axons send electrical impulses to the brain. Their axons meet at the base of the eye and travel down the optic stalk. This stalk is then called the optic nerve. Neural retina differentiation Like the cerebral and cerebellar cortices, the neural retina develops into a layered array of different neuronal types (Figure 12.29). These layers include the light- and colorsensitive photoreceptor (rod and cone) cells, the cell bodies of the ganglion cells, and the bipolar interneurons that transmit electrical stimuli from the rods and cones to the ganglion cells. In addition, there are numerous Mller glial cells, which maintain the integrity of the retina, as well as amacrine neurons (which lack large axons) and horizontal neurons that transmit electrical impulses in the plane of the retina. In the early stages of retinal development, cell division in a germinal layer and the migration and differential death of the resulting cells form the striated, laminar pattern of the neural retina. The formation of this highly structured tissue is one of the most intensely studied problems of developmental neurobiology. It has been shown (Turner and Cepko 1987) that a single neuroblast precursor cell from the retinal germinal layer can give rise to at least three types of neurons or to two types of neurons and a glial cell. This analysis was performed using an ingenious technique to label the cells generated by one particular neuroblast precursor cell. Newborn rats (whose retinas are still developing) were injected in the back of their eyes with a virus that can integrate into their DNA. This virus contained a galactosidase gene (not present in rat retina) that would be expressed only in the infected cells. A month after the rats' eyes were infected, the retinas were removed and stained for the presence of -galactosidase. Only the progeny of the infected cells should have stained blue. Figure 12.30 shows one of the strips of cells derived from an infected precursor cell. The stain can be seen in five rods, a bipolar neuron, During its continued development into a lens, the lens placode rounds up and contacts the new overlying ectoderm. The lens vesicle then induces the ectoderm to form the transparent cornea. Here, physical parameters play an important role in the development of the eye. Intraocular fluid pressure is necessary for the correct curvature of the cornea so that light can be focused on the retina. The importance of this pressure can be demonstrated experimentally: the cornea will not develop its characteristic curve when a small glass tube is inserted through the wall of a developing chick eye to drain away the intraocular fluid (Coulombre 1956, 1965). Intraocular pressure is sustained by a ring of scleral bones (probably derived from the neural crest), which acts as an inelastic restraint. The differentiation of the lens tissue into a transparent membrane capable of directing light onto the retina involves changes in cell structure and shape as well as the synthesis of transparent, lens-specific proteins called crystallins (Figure 12.31). The cells at the inner portion of the lens vesicle elongate and, under the influence of the neural retina,
Notes made by kozi1989 email:kozi1989@yahoo.gr
224
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
become the lens fibers (Piatigorsky 1981). As these fibers continue to grow, they synthesize crystallins, which eventually fill up the cell and cause the extrusion of the nucleus. The crystallin-synthesizing fibers continue to grow and eventually fill the space between the two layers of the lens vesicle. The anterior cells of the lens vesicle constitute a germinal epithelium, which keeps dividing. These dividing cells move toward the equator of the vesicle, and as they pass through the equatorial region, they, too, begin to elongate (Figure 12.31D). Thus, the lens contains three regions: an anterior zone of dividing epithelial cells, an equatorial zone of cellular elongation, and a posterior and central zone of crystallincontaining fiber cells. This arrangement persists throughout the lifetime of the animal as fibers are continuously being laid down. In the adult chicken, the process of differentiation from an epithelial cell to a lens fiber takes 2 years (Papaconstantinou 1967). Directly in front of the lens is a pigmented and muscular tissue called the iris. The iris muscles control the size of the pupil (and give an individual his or her characteristic eye color). Unlike the other muscles of the body (which are derived from the mesoderm), part of the iris is derived from the ectodermal layer. Specifically, this region of the iris develops from a portion of the optic cup that is continuous with the neural retina, but does not make photoreceptors. The evolution of the eye has been a subject of significant study, as a distinctive example of a homologous organ present in a wide variety of taxa. Certain components of the eye, such as the visual pigments, appear to have a common ancestry that is, they evolved once, before the animals radiated. However, complex, image-forming eyes evolved some 50 to 100 times[1] using many of the same proteins and genetic toolkits in their construction. Complex eyes appear to have first evolved within a few million years, in the rapid burst of evolution known as the Cambrian explosion. There is no evidence of eyes before the Cambrian, but a wide range of diversity is evident in the Middle Cambrian Burgess shale. Eyes show a wide range of adaptations to meet the requirements of the organisms which bear them. Eyes may vary in their acuity, the range of wavelengths they can detect, their sensitivity in low light levels, their ability to detect motion or resolve objects, and whether they can discriminate colours. 104. Comparative anatomy of digestive system. Oral cavity. Teeth - evolutionary changes Digestive tract - tube from mouth to vent or anus that functions in: Ingestion Digestion Absorption Egestion
Notes made by kozi1989 email:kozi1989@yahoo.gr
225
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Major subdivisions include the oral cavity, pharynx, esophagus, stomach, small & large intestines, and cloaca. Accessory organs include the tongue, teeth, oral glands, pancreas, liver, & gall bladder. Differences in the anatomy of vertebrate digestive tracts is often correlated with the nature & abundance of food:
1) readily absorbed (e.g., hummingbirds) vs. requiring extensive enzymatic activity (e.g., carnivores) 2) constant food supply (e.g., herbivores) vs. scattered supply (e.g., carnivores)
Mouth & oral cavity. The oral cavity begins at the mouth & ends at the pharynx. Fish have a very short oral cavity, while tetrapods typically have longer oral cavities. The mammalian mouth is specialized to serve as a suckling and masticatory organ (with muscular cheeks).
1) Palate = roof of the oral cavity 2) primary palate - internal nares lead into the oral cavity anteriorly 3) secondary palate - nasal passages are located above the secondary palate and open at the end of the oral cavity In amphioxus the digestive tract consists of only three components: the oral cavity, the pharynx, and a tubular postpharyngeal gut without subdivisions. The same condition holds in the most primitive living vertebrates, the cyclostomes (lampreys and hagfishes). In higher vertebrates, however, the postpharyngeal gut is almost always subdivided into a series of regions that are both anatomically and functionally distinct. The most common is the esophagusstomachsmall intestinelarge intestinerectum (or cloaca) sequence. Teeth are derivations of dermal armor. 1) Placoid scales - show gradual transition to teeth at the edge of the jaw 2) Composition of teeth - primarily dentin surrounded by enamel 3) Vary among vertebrates in number, distribution in the oral cavity, degree of permanence, mode of attachment, & shape
Toothless vertebrates are found in every class of vertebrates and include agnathans, sturgeons, some toads, turtles, birds, & baleen whales. A right whale swims at or near the surface of the water with its mouth open. Water and food enter through a gap in the front baleen plates, and food is caught in the matted baleen fringes inside. Toothed vertebrates
Notes made by kozi1989 email:kozi1989@yahoo.gr
226
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
1) Fish - teeth are numerous & widely distributed in the oral cavity & pharynx 2) Early tetrapods - teeth widely distributed on the palate; most amphibians & some reptiles still have teeth on the vomer, palatine, & pterygoid bones 3) Crocodilians, toothed birds, & mammals - teeth are limited to the jaws Teeth have tended toward reduced numbers & distribution most vertebrates (through reptiles) have succession of teeth most vertebrates (except mammals) replace teeth in waves (back to front; every other tooth) mammals generally develop 2 sets of teeth: milk (deciduous) teeth & permanent teeth
Morphological variation in teeth: Vertebrates other than mammals - all teeth are shaped alike (homodont dentition) Mammals - teeth exhibit morphological variation: incisors, canines, premolars, & molars (heterodont dentition) Incisors = cutting Canines = piercing & tearing Premolars & molars = macerating On jaws normally Cheeks in mammals form pocket Acrodont teeth- fish and snakes Bicuspid- amphibians Tricuspid- lizards Pleurodont teeth- snakes Thecodont teeth- crocodilians
1) 2) 3) 4) 5) 6) 7)
The intestine is located between the stomach & the cloaca or anus & is an important site for digestion & absorption. Vertebrate intestines are differentiated to varying degrees into small & large intestines. Fishes - relatively straight & short intestine in cartilaginous fishes & in primitive bony fishes (lungfish & sturgeon). However, the intestine of cartilaginous fishes has a spiral valve. Amphibians - intestines differentiated into coiled small intestine and short, straight
Notes made by kozi1989 email:kozi1989@yahoo.gr
227
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
large intestine Reptiles & Birds - coiled small intestines & a relatively short large intestine (that empties into the cloaca) Mammals - small intestine long & coiled and differentiated into duodenum, jejunum, & ileum. The large intestine is often relatively long (but not as long as the small intestine). A cecum is often present at the junction of the small & large intestines in herbivores. Cloaca 1) chamber at end of digestive tract that receives the intestine, & urinary & genital ducts, & opens to the exterior via the vent 2) shallow or non-existent in lampreys, ray-finned fishes, & mammals (except monotremes) 3) if no cloaca is present, the intestine opens directly to the exterior via anus 105. Comparative anatomy of tongue and oral glands. Stomach - evolutionary changes The first appearance of a salivary apparatus has been observed by Owen, in a genus of Entozoa found in the stomach of the tiger, and named by him Gnathostoma. It consists of four elongated straight blind tubes, each about two lines in length, placed at equal distances around the commencement of the alimentary canal, having their small extremities directed forward, and opening into the mouth. Among the Echinodermata the salivary organs in Holothuria regalis are represented by elongated cwcal processes, surrounding the cesophagus, and continued into the branched tentacles around the mouth. They exude a viscid secretion, which assists in entangling the objects which constitute the food of the animal, lubricating them, and adapting them for deglutition. In Myriapoda the salivary glands consist of small transparent vesicles, constituting in Julus terrestris, for example, a clavate mass, the small extremity of which terminates in a twisted excretory canal opening into the pharynx. They are large and very vascular in the Scolopendridm. In the Insecta the salivary glands evacuate themselves either into the mouth, or the com mencement of the intestine in front of the stomach. A. Salivary vessels opening into the mouth, generally beneath the tongue, and more seldom at the base of the mandibles. In Cirrhopoda the salivary glands are two in number and of considerable size, opening into the commencement of a short cesopha gus. Among Pteropoda they are found in Clio as two long and slender glands placed at the sides of the cesophagus, and pouring their secretion into the mouth. They " present in the Gasteropoda different forms and degrees of development bearing the ordinary relations to the construction of the mouth and the nature of the food. In the Calyptrwa they are represented by two simple elongated se creting tubes. In the whelk tbey present a conglomerate structure, and are situate at each side of the cesophagus at the base of the
Notes made by kozi1989 email:kozi1989@yahoo.gr
228
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
proboscis, along which they transmit their slender ducts to terminate on each side the anterior spines of the tongue." (Owen.) In the snail they are flattened, elongated, and irregular in form, and conglomerate in struc ture, diminishing in breadth as they proceed upwards to the pharynx, where their ducts terminate. In the Vaginulus an additional slender tube whicb lies first on the stomach, passes through the nervous collar to join the duct by which the saliva is discharged. The salivary glands are present in all the Cephalopoda, with the exception of Loligo psis. In the Onychoteuthis two glands are situated at the root of the tongue. They are in general, however, four in number, two at the root of the tongue, which give off distinct ducts which terminate at the commencement of the cesophagus ; the other pair, generally longer than the superior, is lodged in the vis ceral sac, on each side of the upper part of the crop. The ducts of the last form a single tube which opens in the neighbourhood of the spiny portion of the tongue. The salivary glands are absent in Pisces. Among reptiles in the Chelonian, Saurian, and Batrachian orders, the substance of the tongue seems principally made up of a glan dular mass formed by a multitude of little tubes united at their bases, but becoming se parate towards the surface of the tongue. In the Ophidian reptiles two glandular organs placed immediately beneath the skin of the gums surround the margins of the upper and lower jaw, and pour an abundant salivary secretion into the mouth. (Rymer Jones.) In many genera the salivary apparatus is de ficient. The poison glands of serpents can hardly be reckoned among the salivary organs, being destined for a special secretion, and forming the analogues to similar glandg in the Arachnida. In Ave: the salivary glands present con siderable variation in their number, position, and degree of development. In the crow the only indication of a salivary apparatus is a series of simple cone-shaped follicles, placed along the sides of the oral cavity, upon the rnucous membrane of which they open by distinct orifices. In general, however, there are four pairs, two sublingual on each side beneath the tongue, two maxillary, divided each into an anterior and posterior, and opening by special ducts in front of the tongue, and a gland which can be compared to the parotid. These are generally all present in the Rapaces, Pas seres, and Gallimm ; and appear to be absent in Sula, Carbo, and Phmnicopterus, and but slightly developed in the Grallce and Palmi pedes generally. In the goose they occupy the entire space included between the rami of the lower jaw, being closely united in the median line, and opening into the mouth on each side of this by a series of orifices. In the watercoot and Hirundo esculenta, the parotid is highly developed, in the latter the secretion serving for the preparation of its edible nests. In the woodpecker the glan dular mass is of extraordinary size, extending from the angle to the symphysis of the jaw on each side, and opening by the confluence of the two ducts into a single orifice at the apex of the mouth. In Illanznzalia the salivary glands present considerable variation. In the Monotremata they are partially deficient : in the Echidna there appears to be no parotid ; the submax Diary, on the other hand, is highly developed, extending from the meatus auditorius along the neck, and upon the anterior part of the thorax. Its ducts terminate by numerous orifices on the membranous floor of the mouth, and pour out a secretion for the lubrification of its
Notes made by kozi1989 email:kozi1989@yahoo.gr
229
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
long and slender tongue. In the Cetacea the salivary.glands are absent. In the Dugong, however, one of the herbivo rous Cetacea, the parotids are highly deve loped. In the Ruminantia the three pairs are highly developed, particularly the parotid ; and in addition to these there is a group, apparently continuous with the molar, which mounts up along the superior maxillary bone, beneath the zygonia, to the globe of the eye, as observed in the ox, the sheep, and the horse. The excretory ducts pierce the mu cous membrane near the posterior margin of the superior alveolar ridge. In the armadillo, among the Edentata, the submaxillary gland has appended a reservoir or bladder, receiving the saliva by small ducts, which open into it posteriorly in a valvular manner. A single duct comes off from its anterior part, and terminates just behind the symphysis of the lower jaw. The saliva is very tenacious, the serous part being pro bably absorbed during its detention in the reservoir, and is expelled at the extremity of the mouth, in order to lubricate the tongue, which is by this means rendered subservient, as in the anteater, to the catching of insects. In the latter animal the salivary secretion takes place from two glands, situated, accord ing to Cuvier, the one in contact below with the upper edge of the masseter, and filling up a great part of the temporal, zygomatic, and orbital foss ; the excretory duct opening into the mouth behind the superior maxilla: the other, probably furnishing the viscid secre tion that coats the tongue in front of the tendon of the masseter, behind the angle of the lips, and then running along the edge of the lower lip as far as the middle. Its canal opens externally at the commissure of the lips. In the Carnivora the variations of the sali vary glands are but slight. The submaxillary in them, as in the Rodentia and Ruminantia, are large. The sublingual gland is absent in the cat. Differences in the anatomy of vertebrate digestive tracts is often correlated with the nature & abundance of food:
1) readily absorbed (e.g., hummingbirds) vs. requiring extensive enzymatic activity (e.g., carnivores) 2) constant food supply (e.g., herbivores) vs. scattered supply (e.g., carnivores)
The embryonic digestive tract of vertebrates consists of 3 regions:
1) Midgut - contains yolk or attached yolk sac 2) Foregut - oral cavity, pharynx, esophagus, stomach, & small intestine 3) Hindgut - large intestine & cloaca Tongue
Notes made by kozi1989 email:kozi1989@yahoo.gr
230
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
1) Gnathostome fish & primitive amphibians - tongue is a simple crescent-shaped elevation in the floor of the oral cavity caused by the underlying hyoid skeleton & is called the primary tongue 2) Most amphibians - primary tongue (or hypobranchial eminence) + glandular field (or tuberculum impar) ('stuffed' with hypobranchial musculature) 3) Reptiles & mammals - primary tongue + glandular field (or tuberculum impar) + lateral lingual swellings (more hypobranchial muscle) 4) Birds - lateral lingual swellings are suppressed & intrinsic muscle is usually lacking
Tongue mobility: Turtles, crocodilians, some birds, & whales - tongue is largely immobilized in the floor of the oral cavity & cannot be extended Snakes, insectivorous lizards & amphibians, & some birds - tongue sometimes long and may move in and out of the oral cavity Mammals - tongue is attached to the floor of the oral cavity (via the frenulum) but can still be extended out of the oral cavity Using a keen sense of smell, anteaters are able to effectively track down ant nests on the forest floor. Once a nest is found, the mammal usually rips it open with its sharp foreclaws to expose its delectable contents. The anteater then proceeds to catch and eat the ants by repetitively flicking its long sticky tongue in and out of the nest. The giant anteater's unique tongue can measure as long as two feet (60 cm). Functions of vertebrate tongues:
1) 2) 3) 4) 5) 6) 7)
Capturing & gathering food (see woodpecker tongue below) Taste Manipulate fluids & solids in oral cavity Swallowing Thermoregulation Grooming Human speech
Oral glands - secrete a variety of substances including: Lubrication and binding: the mucus in saliva is extremely effective in binding masticated food into a slippery bolus that (usually) slides easily through the esophagus without inflicting damage to the mucosa. Saliva also coats the oral cavity and esophagus, and food basically never directly touches the epithelial cells of those tissues. Solubilizes dry food: in order to be tasted (by taste buds), the molecules in food must be solubilized.
Notes made by kozi1989 email:kozi1989@yahoo.gr
231
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Oral hygiene: The oral cavity is almost constantly flushed with saliva, which floats away food debris and keeps the mouth relatively clean. Saliva also contains lysozyme, an enzyme that lyses many bacteria and prevents overgrowth of oral microbial populations. Initiates starch digestion: in most species, amylase is present in saliva and begins to digest dietary starch into maltose. Amylase does not occur in the saliva of carnivores. Provides alkaline buffering and fluid: this is of great importance in ruminants, which have non-secretory forestomachs. Evaporative cooling: clearly of importance in dogs, which have very poorly developed sweat glands - look at a dog panting after a long run and this function will be clear. 1) poison (lizards, snakes, and mammals) 2) anticoagulant (vampire bats; video) Stomach = muscular chamber(s) at end of esophagus serves as storage & macerating site for ingested solids & secretes digestive enzymes Vertebrate stomachs: 1) Cyclostomes - weakly developed; similar to esophagus 2) Fish, amphibians, & reptiles - increasing specialization (more differentiated from the esophagus) 3) Birds - proventriculus (glandular stomach) and ventriculus (muscular stomach, or gizzard) 4) Mammals - well-developed stomach; ruminants have multichambered stomachs: Reticulum and rumen are often discussed together since each compartment is separated by a low partition. Eighty percent of the capacity of the stomach is related to the reticulo-rumen. The contents of the reticulum and rumen intermix freely. The rumen is the main fermentation vat where billions of microorganisms attack and break down the relatively indigestible feed components of the ruminant's diet. Omasum After fermentation in the reticulum and rumen, food passes to the omasum. The omasum acts as a filter pump to sort liquid and fine food particles. Coarse fibre particles are not allowed to enter the omasum. Also, the omasum may be the site for absorption of water, minerals and nitrogen. Abomasum The abomasum is the true stomach and the only site on the digestive tract that produces gastric juices (HCl and the enzymes, pepsin and rennin). Ingesta only remains here for 1 to 2 hours
Notes made by kozi1989 email:kozi1989@yahoo.gr
232
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
106. Comparative anatomy of circulatory system. Development of heart and aortic arches The circulatory system is one of the great achievements of the lateral plate mesoderm. Consisting of a heart, blood cells, and an intricate system of blood vessels, the circulatory system provides nourishment to the developing vertebrate embryo. The circulatory system is the first functional unit in the developing embryo, and the heart is the first functional organ. The vertebrate heart arises from two regions of splanchnic mesoderm one on each side of the body that interact with adjacent tissue to become specified for heart development. In amniote vertebrates, the embryo is a flattened disc, and the lateral plate mesoderm does not completely encircle the yolk sac. The presumptive heart cells originate in the early primitive streak, just posterior to Hensen's node and extending about halfway down its length. These cells migrate through the streak and form two groups of mesodermal cells lateral to (and at the same level as) Hensen's node. These groups of cells are called the cardiogenic mesoderm. The cells forming the atrial and ventricular musculature, the cushion cells of the valves, the Purkinje conducting fibers, and the endothelial lining of the heart are all generated from these two clusters. When the chick embryo is only 18 20 hours old, the presumptive heart cells move anteriorly between the ectoderm and endoderm toward the middle of the embryo, remaining in close contact with the endodermal surface. When these cells reach the lateral walls of the anterior gut tube, migration ceases. The directionality for this migration appears to be provided by the foregut endoderm. If the cardiac region endoderm is rotated with respect to the rest of the embryo, migration of the cardiogenic mesoderm cells is reversed. It is thought that the endodermal componentresponsible for this movement is an anterior-toposterior concentration gradient of fibronectin. Antibodies against fibronectin stop the migration, while antibodies against other extracellular matrix components do not. The endoderm and primitive streak also specify the some of the cardiogenic cells to become heart muscles. Cerberus and an unknown factor, possibly BMP2 in the anterior endoderm, induce the synthesis of the Nkx2-5 transcription factor in the migrating mesodermal cells that will become the heart. Nkx2-5 is a critical protein in instructing the mesoderm to become heart tissue, and it activates the synthesis of other transcription factors (especially members of the GATA and MEF2 families). Working together, these transcription factors activate the expression of genes encoding cardiac muscle-specific proteins (such as cardiac actin, atrial naturetic factor, and the alpha myosin heavy chains). Specification of the heart cells occurs gradually, with the ventricular cells becoming specified prior to the atrial cells. Cell differentiation occurs independently in the two heart-forming primordia that are migrating toward each other. As they migrate, the cells begin to express N-cadherin on their apices and join into an epithelium. A small population of these cells then downregulates N-
Notes made by kozi1989 email:kozi1989@yahoo.gr
233
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
cadherin and delaminates from the epithelium to form the endocardium. The myocardium will form the heart muscles that will pump for the lifetime of the organism. The endocardial cells produce many of the heart valves, secrete the proteins that regulate myocardial growth, and regulate the placement of nervous tissue in the heart. As neurulation proceeds, the foregut is formed by the inward folding of the splanchnic mesoderm . This movement brings the two cardiac tubes together, eventually uniting the myocardium into a single tube. The bilateral origin of the heart can be demonstrated by surgically preventing the merger of the lateral plate mesoderm. Thisresults
in a condition called cardia bifida, in which a separate heart forms on each side of the body. The two endocardia lie within this common tube for a short while, but these will also fuse. At this time, the originally paired coelomic chambers unite to form the body cavity in which the heart resides. This fusion occurs at about 29 hours in chick development and at 3 weeks in human gestation. The unfused posterior portions of the endocardium become the openings of the vitelline veins into the heart. These veins will carry nutrients from the yolk sac into the sinus venosus. The blood then passes through a valvelike flap into the atrial region of the heart. Contractions of the truncus arteriosus speed the blood into the aorta. Pulsations of the heart begin while the paired primordia are still fusing. The pacemaker of this contraction is the sinus venosus. Contractions begin here, and a wave of muscle contraction is propagated up the tubular heart. In this way, the heart can pump blood even before its intricate system of valves has been completed. Heart muscle cells have their own inherent ability to contract, and isolated heart cells from 7-day rat or chick embryos will continue to beat in petri dishes. In the embryo, these contractions become regulated by electrical stimuli from the medulla oblongata via the vagus nerve, and by 4 days, the electrocardiogram of a chick embryo approximates that of an adult. In 3-day chick embryos and 5week human embryos, the heart is a two -chambered tube, with one atrium and one ventricle. In the chick embryo, the unaided eye can see the remarkable cycle of blood entering the lower chamber and being pumped out through the aorta. The looping of the heart converts the original anterior-posterior polarity of the heart tube into the right-left polarity seen in the adult. Thus, the portion of the heart tube destined to become the right ventricle lies anterior to the portion that will become the left ventricle. Within the heart primordium,
Notes made by kozi1989 email:kozi1989@yahoo.gr
234
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
Nkx2 regulates the Hand1 and Hand2 transcription factors. Although the Hand proteins appear to be synthesized throughout the early heart tube, Hand1 becomes restricted to the future left ventricle, and Hand2 to the right, as looping commences. Without these proteins, looping fails to occur normally and the ventricles fail to form properly. The Pitx-2 transcription factor, activated solely in the left side of the lateral plate mesoderm, is also critical for proper heart looping, and it may regulate the expression of proteins such as the extracellular matrix protein flectin to regulate the physical tension of the heart tissues on the different sides. Transcription factors Nkx2-5 and MEF2C also activate the Xin gene, whose protein product, Xin (Chinese for "heart"), may mediate the cytoskeletal changes essential for heart looping.
Notes made by kozi1989 email:kozi1989@yahoo.gr
235
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The separation of atrium from ventricle is specified by the several transcription factors that become restricted to either the anterior or the posterior portion of the heart tube. The partitioning of this tube into a distinctive atrium and ventricle is accomplished when cells from the myocardium produce a factor (probably transforming growth factor 3) that causes cells from the adjacent endocardium to detach and enter the hyaluronate-rich "cardiac jelly" between the two layers.
In humans, these cells cause the formation of an endocardial cushion that divides the tube into right and left atrioventricular channels (Figure 15.8). Meanwhile, the primitive atrium is partitioned by two septa that grow ventrally toward the endocardial cushion. The septa, however, have holes in them, so blood can still cross from one side into the other. This crossing of blood is needed for the survival of the fetus before circulation to functional lungs has been established. Upon the first breath, however, these holes close, and the left
Notes made by kozi1989 email:kozi1989@yahoo.gr
236
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
and
right circulatory loops become established (see Sidelights & Speculations). With the formation of the septa (which usually occurs in the seventh week of human development), the heart is a four-chambered structure with the pulmonary artery connected to the right ventricle and the aorta connected to the left. The human embryonic circulatory system is a modification of that used in other amniotes, such as birds and reptiles. Blood pumped through the dorsal aorta passes over the aortic arches and down into the embryo. Some of this blood leaves the embryo through the vitelline arteries and enters the yolk sac. Nutrients and oxygen are absorbed from the yolk, and the blood returns through the vitelline veins back into the heart through the sinus venosus. The first constraint on vascular development is physiological. Unlike new machines, which do not need to function until they have left the assembly line, new organisms have to function even as they develop. The embryonic cells must obtain nourishment before there is an intestine, use oxygen before there are lungs, and excrete wastes before there are kidneys. All these functions are mediated through the embryonic circulatory system. Therefore, the circulatory physiology of the developing embryo must differ from that of the adult organism. Food is absorbed not through the intestine, but from either the yolk or the placenta, and respiration is conducted not through the gills or lungs, but through the chorionic or allantoic membranes. The major embryonic blood vessels must be constructed to serve these extraembryonic structures. The second constraint is evolutionary. The mammalian embryo extends blood vessels to the yolk sac even though there is no yolk inside. Moreover, the blood leaving the heart via the truncus arteriosus passes through vessels that loop over the foregut to reach the dorsal aorta. Six pairs of aortic arches loop over the pharynx. In primitive fishes, these arches persist and enable the gills to oxygenate the blood. In the adult bird or mammal, in which
Notes made by kozi1989 email:kozi1989@yahoo.gr
237
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
lungs oxygenate the blood, such a system makes little sense, but all six pairs of aortic arches are formed in mammalian and avian embryos before the system eventually becomes simplified into a single aortic arch. Thus, even though our physiology does not require such a structure, our embryonic condition reflects our evolutionary history. The third set of constraints is physical. According to the laws of fluid movement, the most effective transport of fluids is performed by large tubes. As the radius of a blood vessel gets smaller, resistance to flow increases as r-4 (Poiseuille's law). A blood vessel that is half as wide as another has a resistance to flow 16 times greater.
However, diffusion of nutrients can take place only when blood flows slowly and has access to cell membranes. So here is a paradox: the constraints of diffusion mandate that vessels be small, while the laws of hydraulics mandate that vessels be large. Living organisms have solved this paradox by evolving circulatory systems with a hierarchy of vessel sizes. This hierarchy is formed very early in development (and is already well established in the 3-day chick embryo). In dogs, blood in the large vessels (aorta and vena cava) flows over 100 times faster than it does in the capillaries. With a system of large vessels specialized for transport and small vessels specialized for diffusion (where the blood spends most of its time), nutrients and oxygen can reach the individual cells of the growing organism. 107. Comparative anatomy of urinary system pronephros, mesonephros, metanephros
Notes made by kozi1989 email:kozi1989@yahoo.gr
238
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The intermediate mesoderm generates the urogenital system the kidneys, the gonads, and their respective duct systems. While this statement may smack of hyperbole, the kidney is an incredibly intricate organ. Its functional unit, the nephron, contains over 10,000 cells and at least 12 different cell types, with each cell type located in a particular place in relation to the others along the length of the nephron. The development of the mammalian kidney progresses through three major stages. The first two stages are transient; only the third and last persists as a functional kidney. Early in development (day 22 in humans; day 8 in mice), the pronephric duct arises in the intermediate mesoderm just ventral to the anterior somites. The cells of this duct migrate caudally, and the anterior region of the duct induces the adjacent mesenchyme to form the tubules of the initial kidney, the pronephros. While the pronephric tubules form functioning kidneys in fish and in amphibian larvae, they are not
thought to be active in mammalian amniotes. In mammals, the pronephric tubules and the anterior portion of the pronephric duct degenerate, but the more caudal portions of the pronephric duct persist and serve as the central component of the excretory system throughout its development. This remaining duct is often referred to as the nephric or Wolffian duct. As the pronephric tubules degenerate, the middle portion of the nephric duct induces a new set of kidney tubules in the adjacent mesenchyme. This set of tubules constitutes the mesonephros, or mesonephric kidney. In some mammalian species, the mesonephros functions briefly in urine filtration, but in humans and rodents, it does not function as a working kidney. In humans, about 30 mesonephric tubules form, beginning around day 25. As more tubules are induced caudally, the anterior mesonephric tubules begin to regress through apoptosis (although in mice, the anterior tubules remain while the posterior ones regress:. During its brief existence,
Notes made by kozi1989 email:kozi1989@yahoo.gr
239
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
however, the mesonephros provides important developmental functions. First, it is the source of hematopoietic stem cells necessary for blood cell development. Second, in male mammals, some of the mesonephric tubules persist to become the sperm-carrying tubes (the vas deferens and efferent ducts) of the testes. The permanent kidney of amniotes, the metanephros, is generated by some of the same components as the earlier, transient kidney types. It is thought to originate through a complex set of interactions between epithelial and mesenchymal components of the intermediate mesoderm. In the first steps, the metanephrogenic mesenchyme forms in posteriorly located regions of the intermediate mesoderm, and it induces the formation of a branch from each of the paired nephric ducts. These epithelial branches are called the ureteric buds. These buds eventually separate from the nephric duct to become the ureters that take the urine to the bladder. When the ureteric buds emerge from the nephric duct, they enter the metanephrogenic mesenchyme. The ureteric buds induce this mesenchymal tissue to condense around them and differentiate into the nephrons of the mammalian kidney. As this mesenchyme. 108. Origins and evolution of gonads and genital tracts. This environmental view of sex determination remained the only major scientific theory until the rediscovery of Mendel's work in 1900 and the rediscovery of the sex chromosome by McClung in 1902. It was not until 1905, however, that the correlation (in insects) of the female sex with XX sex chromosomes and the male sex with XY or XO chromosomes was established. This finding suggested strongly that a specific nuclear component was responsible for directing the development of the sexual phenotype. Thus, evidence accumulated that sex determination occurs by nuclear inheritance rather than by environmental happenstance. Primary sex determination is the determination of the gonads . In mammals, primary sex determination is strictly chromosomal and is not usually influenced by the environment. In most cases, the female is XX and the male is XY. Every individual must have at least one X chromosome. Since the female is XX, each of her eggs has a single X chromosome. The male, being XY, can generate two types of sperm: half bear the X chromosome, half the Y. If the egg receives another X chromosome from the sperm, the resulting individual is XX, forms ovaries, and is female; if the egg receives a Y chromosome from the sperm, the individual is XY, forms testes, and is male. The Y chromosome carries a gene that encodes a testis-determining factor. This factor organizes the gonad into a testis rather than an ovary. Unlike the situation in Drosophila (discussed below), the mammalian Y chromosome is a crucial factor for determining sex in mammals. A person with five X chromosomes and one Y chromosome (XXXXXY) would be male. Furthermore, an individual with only a single X chromosome and no second X or Y (i.e., XO) develops as a female and begins making ovaries, although the ovarian follicles cannot be maintained. For a complete ovary, a second X chromosome is needed. In mammalian primary sex determination, there is
Notes made by kozi1989 email:kozi1989@yahoo.gr
240
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
no "default state." The formation of ovaries and testes are both active, gene-directed processes. Moreover, as we shall see, both diverge from a common precursor, the bipotential gonad. Secondary sex determination affects the bodily phenotype outside the gonads. A male mammal has a penis, seminal vesicles, and prostate gland . A female mammal has a vagina, cervix, uterus, oviducts, and mammary glands. In many species, each sex has a sexspecific size, vocal cartilage, and musculature. These secondary sex characteristics are usually determined by hormones secreted from the gonads. However, in the absence of gonads, the female phenotype is generated. The gonad is the organ that makes gametes. The gonads in males are the testes and the gonads in females are the ovaries. The product, gametes, are haploid germ cells. For example, spermatozoon and egg cells are gametes. Although scientifically, the term "gonad" can refer to either male gonads (testicles) or female gonads (ovaries), the vernacular, or slang, use of "gonads" (or "nads") usually only refers to the testicles. The gonads embody a unique embryological situation. All other organ rudiments can normally differentiate into only one type of organ. A lung rudiment can become only a lung, and a liver rudiment can develop only into a liver. The gonadal rudiment, however, has two normal options. When it differentiates, it can develop into either an ovary or a testis. The path of differentiation taken by this rudiment determines the future sexual development of the organism. But, before this decision is made, the mammalian gonad first develops through a bipotential (indifferent) stage, during which time it has neither female nor male characteristics. In humans, the gonadal rudiments appear in the intermediate mesoderm during week 4 and remains sexually indifferent until week 7. The gonadal rudiments are paired regions of the intermediate mesoderm; they form adjacent to the developing kidneys. The ventral portions of the gonadal rudiments are composed of the genital ridge epithelium. During the indifferent stage, the genital ridge epithelium proliferates into the loose connective mesenchymal tissue above it. These epithelial layers form the sex cords. The germ cells migrate into the gonad during week 6, and are surrounded by the sex cords. In both XY and XX gonads, the sex cords remain connected to the surface epithelium. If the fetus is XY, the sex cords continue to proliferate through the eighth week, extending deeply into the connective tissue. These cords fuse, forming a network of internal (medullary) sex cords and, at its most distal end, the thinner rete testis. Eventually, the sex cords now called testis cords lose contact with the surface epithelium and become separated from it by a thick extracellular matrix, the tunica albuginea. Thus, the germ cells are found in the cords within the testes. During fetal life and childhood, the testis cords remain solid. At puberty, however, the cords will hollow out to form the seminiferous tubules, and the germ cells will begin to differentiate into sperm.
Notes made by kozi1989 email:kozi1989@yahoo.gr
241
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
The cells of the seminiferous tubule are called Sertoli cells. The Sertoli cells of the testis cords nurture the sperm and secrete anti-Mllerian duct hormone. The sperm are transported from the inside of the testis through the rete testis, which joins the efferent ducts.
Notes made by kozi1989 email:kozi1989@yahoo.gr
242
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
These efferent tubules are the remnants of the mesonephric kidney, and they link the testis to the Wolffian duct, which used to be the collecting tube of the mesonephric kidney. In males, the Wolffian duct differentiates to become the epididymis (adjacent to the testis) and the vas deferens, the tube through which the sperm pass into the urethra and out of the body. Meanwhile, during fetal development, the interstitial mesenchyme cells of
the testes differentiate into Leydig cells, which make testosterone. In females, the germ cells will reside near the outer surface of the gonad. Unlike the sex cords in males, which continue their proliferation, the initial sex cords of XX gonads degenerate. However, the epithelium soon produces a new set of sex cords, which do not penetrate deeply into the mesenchyme, but stay near the outer surface (cortex) of the
Notes made by kozi1989 email:kozi1989@yahoo.gr
243
Notes made by (Dimitrios the greek)kozi1989 for medicine email:kozi1989@yahoo.gr 2010/2011
Year:
organ. Thus, they are called cortical sex cords. These cords are split into clusters, with each cluster surrounding a germ cell. The germ cells will become the ova, and the surrounding cortical sex cords will differentiate into the granulosa cells. The mesenchyme cells of the ovary differentiate into the thecal cells. Together, the thecal and granulosa cells will form the follicles that envelop the germ cells and secrete steroid hormones. Each follicle will contain a single germ cell. In females, the Mllerian duct remains intact, and it differentiates into the oviducts, uterus, cervix, and upper vagina. The Wolffian duct, deprived of testosterone, degenerates. Both sexes have initially two pairs of genital ducts: the mesonephric (Wolffian) ducts and the paramesonephric (Mllerian) ducts. The paramesonephric (Mllerian) duct arises as a longitudinal invagination of the coelomic epithelium on the anterolateral surface of the the urogenital ridge and empties into the urogenital sinus. It can be divided into three parts: a) the cranial part which opens into coelomic cavity, b) the horizontal part which crosses the mesonephric duct, and c) the caudal part which fuses with the same duct from the other side. The cranial parts develop into the uterine tubes while the caudal fused parts into the uterus, cervix and one third of the vagina. The surrounding layer of the mesenchyme forms the myometrium and perimetrium. When the paramesonephric ducts reach the urogenital sinus, two evaginations (sinovaginal bulbs) proliferate and form a solid vaginal plate. By the 5th month, this plate is completely canalized. Thus, the vagina has a dual origin: upper one third derives from the paramesonephric duct and the lower two thirds from the urogenital sinus. The paramesonephric duct in the male degenerates, except for a small portion known as the appendix testis and utriculus prostaticus. The mesonephric (Wolffian) duct arises by the canalization of a solid mass of cells in the urogenital ridge and empties into the urogenital sinus laterally to the paramesonephric duct orificies. It gives rise to the ductus epididymis, seminal vesicle, ductus deferens and ejaculatory duct. The mesonephric duct in female regresses, but may persist as the appendix vesiculosa, epoophoron or paroophoron and Gartner's duct. In both sexes, a branch of mesonephric duct forms the ureteric bud.
Notes made by kozi1989 email:kozi1989@yahoo.gr
244
You might also like
- What is DNA and its vital role in the human bodyDocument1 pageWhat is DNA and its vital role in the human bodyayeshaNo ratings yet
- Genes - and - GenomesDocument6 pagesGenes - and - GenomesEdwinNo ratings yet
- The Role of Nucleic Acids in HeredityDocument4 pagesThe Role of Nucleic Acids in HeredityRaluca GladcencoNo ratings yet
- BIO206 NotesDocument7 pagesBIO206 NotesKaraLVNo ratings yet
- All About DnaDocument65 pagesAll About DnaPeopwdNo ratings yet
- BCH 323 NotesDocument41 pagesBCH 323 NotesvictorNo ratings yet
- The Physical Structure of The Human Genome: Nuclear DNADocument14 pagesThe Physical Structure of The Human Genome: Nuclear DNAyusufibanuNo ratings yet
- From Wikipedia, The Free Encyclopedia: Jump To Navigation Jump To Search Introduction To Genetics DNA (Disambiguation)Document3 pagesFrom Wikipedia, The Free Encyclopedia: Jump To Navigation Jump To Search Introduction To Genetics DNA (Disambiguation)Lali HajzeriNo ratings yet
- Molecular Basis of InheritanceDocument54 pagesMolecular Basis of InheritanceLNo ratings yet
- Animal ReproductionDocument34 pagesAnimal ReproductionJenmae Dacullo MansiguinNo ratings yet
- Name - Ozeye Eka-WPS OfficeDocument8 pagesName - Ozeye Eka-WPS OfficeEMMANUEL SALVATIONNo ratings yet
- Introduction To Molecular BiologyDocument39 pagesIntroduction To Molecular BiologyAbbinaya100% (9)
- ReproductionDocument2 pagesReproductionPetrus GapNo ratings yet
- Lapuz - GeneticsDocument3 pagesLapuz - GeneticsEllyza LapuzNo ratings yet
- Deoxyribonucleic Acid (DNA) : DNA: Structure, Function and DiscoveryDocument18 pagesDeoxyribonucleic Acid (DNA) : DNA: Structure, Function and DiscoveryHajar Zarguan (Full stack developer)No ratings yet
- Reality of LifeDocument2 pagesReality of LifeDjela117No ratings yet
- Genetics Ug IaDocument63 pagesGenetics Ug Iamuhammadnajaatu98No ratings yet
- Lecture III - Semi-FinalsDocument16 pagesLecture III - Semi-FinalsENGETS IT7No ratings yet
- Nucleic AcidDocument3 pagesNucleic AcidSamraNo ratings yet
- Unit-I: Characteristics & Structure of GenesDocument64 pagesUnit-I: Characteristics & Structure of GenesMonika JosephNo ratings yet
- Cell StructureDocument8 pagesCell StructurePallabi ChatterjeeNo ratings yet
- Genetic MaterialDocument27 pagesGenetic Material82g7pxg2tsNo ratings yet
- Dogma Genetic and Molecular GeneticDocument11 pagesDogma Genetic and Molecular GeneticMarina Ivana MumpelNo ratings yet
- LECTURE NO 5 - 10 BiologyDocument10 pagesLECTURE NO 5 - 10 BiologyRebell AeonNo ratings yet
- By DR Ananya MandalDocument4 pagesBy DR Ananya MandalhasimNo ratings yet
- Introduction To DNA Fingerprinting (Definition, Importance of DNA Fingerprinting in Forensic Science)Document20 pagesIntroduction To DNA Fingerprinting (Definition, Importance of DNA Fingerprinting in Forensic Science)DeMetrius Sam DanielNo ratings yet
- GeneticsDocument4 pagesGeneticsvolcanowizardNo ratings yet
- Genetic CodeDocument3 pagesGenetic CodeAngelica PagaduanNo ratings yet
- Genetics: The Genetic MaterialDocument25 pagesGenetics: The Genetic Materialعمار مصعب عادلNo ratings yet
- The Structure of DNADocument3 pagesThe Structure of DNAkalaysiahart01No ratings yet
- Chapter 3 - Genetics (Fundamentals of Anatomy and Physiology For Nursing and Healthcare Students)Document87 pagesChapter 3 - Genetics (Fundamentals of Anatomy and Physiology For Nursing and Healthcare Students)Mary Grace MartinNo ratings yet
- M1 Possible QDocument10 pagesM1 Possible Qtaif_alam9758No ratings yet
- Lesson Discussion: Self-Learning Module (SLM) General Biology 2Document7 pagesLesson Discussion: Self-Learning Module (SLM) General Biology 2almafebe caselNo ratings yet
- Assignment 1Document3 pagesAssignment 1NADASHREE BOSE 2248138No ratings yet
- GeneticsDocument25 pagesGeneticsLouel Mattew LagascaNo ratings yet
- Science 10 (Reporting)Document76 pagesScience 10 (Reporting)anneNo ratings yet
- Research About DNA Group 5Document5 pagesResearch About DNA Group 5Kaye Selene Raphaelle SyNo ratings yet
- DNA (Deoxyribonucleic Acid)Document5 pagesDNA (Deoxyribonucleic Acid)Sagar MohanNo ratings yet
- Saq Biology For Aliha UniversityDocument7 pagesSaq Biology For Aliha UniversityEDEN GARDENNo ratings yet
- BiologyDocument24 pagesBiologyTom KiloitNo ratings yet
- Dna and RnaDocument11 pagesDna and Rnaabida.syeda.718No ratings yet
- What is DNADocument2 pagesWhat is DNAEly SibayanNo ratings yet
- Molecular Genetics and DNA StructureDocument14 pagesMolecular Genetics and DNA StructureYitbarek TesfayeNo ratings yet
- HeredityDocument24 pagesHeredityvivas.kznne.9No ratings yet
- Your Genes Are Part of What Makes You The Person You AreDocument4 pagesYour Genes Are Part of What Makes You The Person You ArePrincess Lourdes CarinNo ratings yet
- What Is DNA and RNADocument2 pagesWhat Is DNA and RNASabel RegalaNo ratings yet
- What Is A GenomeDocument22 pagesWhat Is A GenomeNewtonNo ratings yet
- The Importance of DNA and RNADocument13 pagesThe Importance of DNA and RNAJamieNo ratings yet
- DuplicateDocument170 pagesDuplicatecharnellelamulaNo ratings yet
- 2ndqbiotechweek5 7Document3 pages2ndqbiotechweek5 7Evefel Ruth Yparraguirre SanchezNo ratings yet
- Bicol University Nucleic Acids LectureDocument27 pagesBicol University Nucleic Acids LectureKim GoNo ratings yet
- DNArep GNT2 00431Document3 pagesDNArep GNT2 00431Abraham Flores RendónNo ratings yet
- 1 Genetics For PublicationDocument13 pages1 Genetics For PublicationSucheta BansalNo ratings yet
- Genetics: Structure of The Dna MoleculeDocument8 pagesGenetics: Structure of The Dna MoleculeLP JimeNo ratings yet
- DNA ReplicationDocument55 pagesDNA ReplicationShahil AlamNo ratings yet
- Structure and Function of DNA Presentation in Colorful Retro Illustrative StyleDocument51 pagesStructure and Function of DNA Presentation in Colorful Retro Illustrative Stylekimmymantos022No ratings yet
- ReviewerDocument75 pagesReviewerKurt SecretarioNo ratings yet
- Define!!!!Document6 pagesDefine!!!!Lenka AnastasovskiNo ratings yet
- Anatomy & Physiology of The PharynxDocument14 pagesAnatomy & Physiology of The Pharynxİsmail AtamNo ratings yet
- Amyotrophic Lateral SclerosisDocument14 pagesAmyotrophic Lateral Sclerosisİsmail AtamNo ratings yet
- Bell PalsyDocument4 pagesBell Palsyİsmail AtamNo ratings yet
- Ερωτήσεις κυταρρολογίαςDocument99 pagesΕρωτήσεις κυταρρολογίαςİsmail AtamNo ratings yet
- Genetik 1 - 40Document61 pagesGenetik 1 - 40İsmail AtamNo ratings yet
- Acid Base SmallDocument3 pagesAcid Base Smallİsmail AtamNo ratings yet
- BiologyDocument244 pagesBiologyİsmail AtamNo ratings yet
- Neuro ExamDocument5 pagesNeuro Examİsmail AtamNo ratings yet
- 4Document3 pages4İsmail AtamNo ratings yet
- Everything You Need to Know About Obstructive Sleep ApneaDocument64 pagesEverything You Need to Know About Obstructive Sleep Apneaİsmail AtamNo ratings yet
- Medical Parasitology PDFDocument150 pagesMedical Parasitology PDFYajee Macapallag100% (1)
- Medical Parasitology PDFDocument150 pagesMedical Parasitology PDFYajee Macapallag100% (1)
- EarthquakesDocument8 pagesEarthquakesİsmail AtamNo ratings yet
- Medical Parasitology PDFDocument150 pagesMedical Parasitology PDFYajee Macapallag100% (1)
- Ερωτήσεις κυταρρολογίαςDocument99 pagesΕρωτήσεις κυταρρολογίαςİsmail AtamNo ratings yet
- Physiologyv PT IDocument279 pagesPhysiologyv PT Iİsmail AtamNo ratings yet
- A Guide To Wastewater MicrobiologyDocument14 pagesA Guide To Wastewater Microbiologyabduyassin8014No ratings yet
- Unit I Introduction To The World of BiologyDocument25 pagesUnit I Introduction To The World of BiologyJericho De VeraNo ratings yet
- Ix Biology AdamjeeDocument7 pagesIx Biology AdamjeeSaif ObaidNo ratings yet
- Genetic Cheat SheetDocument11 pagesGenetic Cheat SheetLeah MaloneyNo ratings yet
- Step Forward in Plant Taxonomy Book (Final)Document113 pagesStep Forward in Plant Taxonomy Book (Final)DoctorQariNo ratings yet
- Structure Functions of The Eukaryotic Cell MembraneDocument39 pagesStructure Functions of The Eukaryotic Cell MembraneHeswer RajNo ratings yet
- Cells, Tissues and OrgansDocument6 pagesCells, Tissues and OrgansatheelNo ratings yet
- CSEC Biology Revision Guide AnswersDocument29 pagesCSEC Biology Revision Guide AnswersCharlobabooram50% (2)
- Differences Between Prokaryotic and Eukaryotic Cells (38 charactersDocument4 pagesDifferences Between Prokaryotic and Eukaryotic Cells (38 charactersمصطفى رسول هاديNo ratings yet
- The Story of Life: Chapters 7-12 (Evolution) : Thorough With Your Answers! 12points!Document3 pagesThe Story of Life: Chapters 7-12 (Evolution) : Thorough With Your Answers! 12points!Majeda Abdul-JabbaarNo ratings yet
- BIO2107 - Tutorial 1 Discussion PointsDocument5 pagesBIO2107 - Tutorial 1 Discussion PointsAjay Sookraj RamgolamNo ratings yet
- MPN Untuk AlgaDocument8 pagesMPN Untuk AlgaIyak LectureNo ratings yet
- Microbial TaxonomyDocument20 pagesMicrobial TaxonomySebna NoushadNo ratings yet
- Biology Applied Physics BiochemistryDocument6 pagesBiology Applied Physics BiochemistrychukwukerechimezirimNo ratings yet
- Activity 2. Plant Cell Vs Animal CellDocument5 pagesActivity 2. Plant Cell Vs Animal CellMary Joyce BonayonNo ratings yet
- Cells - Molecules and MechanismsDocument277 pagesCells - Molecules and MechanismsscottswallowsNo ratings yet
- Plant and Animal Cell ComparisonDocument27 pagesPlant and Animal Cell ComparisonReinalyn MendozaNo ratings yet
- Cell Parts and Their FunctionsDocument21 pagesCell Parts and Their FunctionsJeff Poi50% (2)
- Test Bank For Biology 5th Edition Robert Brooker Eric Widmaier Linda Graham Peter StilingDocument29 pagesTest Bank For Biology 5th Edition Robert Brooker Eric Widmaier Linda Graham Peter StilingJanice Linder100% (31)
- 4.microbial TaxonomyDocument26 pages4.microbial TaxonomyshibanginiNo ratings yet
- Biology (Topic 1) PowerpointDocument23 pagesBiology (Topic 1) Powerpoint18hollickmNo ratings yet
- Long Quiz 1 BiologyDocument2 pagesLong Quiz 1 BiologyNonita Caracas CubiloNo ratings yet
- Module-2 (Without Edits)Document27 pagesModule-2 (Without Edits)Jennie SoloNo ratings yet
- Activity 1Document4 pagesActivity 1sharksiedNo ratings yet
- Unit 2-Classification System in BiologyDocument38 pagesUnit 2-Classification System in BiologyCAD JunctionNo ratings yet
- Test Bank For Karps Cell and Molecular Biology 9th Edition Gerald Karp Janet Iwasa Wallace Marshall 2Document39 pagesTest Bank For Karps Cell and Molecular Biology 9th Edition Gerald Karp Janet Iwasa Wallace Marshall 2Valerie Gilliam100% (38)
- 2006 - Forterre - The Origin of Viruses and Their Possible Roles in Major Evolutionary Transitions PDFDocument12 pages2006 - Forterre - The Origin of Viruses and Their Possible Roles in Major Evolutionary Transitions PDFJames Roi BerroyaNo ratings yet
- Microbiology BiostatisticsDocument89 pagesMicrobiology BiostatisticsNiyati Karetha100% (1)
- Chapter 1 (1) .PPTX Micro-ParaDocument36 pagesChapter 1 (1) .PPTX Micro-ParaLorelie AsisNo ratings yet
- Lesson 8 Kingdom ProtistaDocument8 pagesLesson 8 Kingdom ProtistaKrish D.No ratings yet