You might also like
- Inspiration From Nature: Insights From Crustacean Chemical Sensors Can Lead To Successful Design of Artificial Chemical SensorsDocument21 pagesInspiration From Nature: Insights From Crustacean Chemical Sensors Can Lead To Successful Design of Artificial Chemical SensorsAnisahNo ratings yet
- Social Predation: How Group Living Benefits Predators and PreyFrom EverandSocial Predation: How Group Living Benefits Predators and PreyNo ratings yet
- Taylor Et Al 2012Document6 pagesTaylor Et Al 2012drinkto remenberNo ratings yet
- Chemical Communication: The Language Of PheromonesFrom EverandChemical Communication: The Language Of PheromonesRating: 3.5 out of 5 stars3.5/5 (5)
- 2013 CellcalciumimagingDocument397 pages2013 CellcalciumimagingThanh ThuanNo ratings yet
- Research ProposalDocument15 pagesResearch Proposalapi-358035887No ratings yet
- Oestrus Pheromones in Farm Animals With Special Reference To CowsDocument12 pagesOestrus Pheromones in Farm Animals With Special Reference To Cowssumantvyas100% (1)
- Using Insect Sniffing Devices For Detection: Glen C. Rains, Jeffery K. Tomberlin and Don KulasiriDocument7 pagesUsing Insect Sniffing Devices For Detection: Glen C. Rains, Jeffery K. Tomberlin and Don KulasirinemanjafilipovicNo ratings yet
- Use of Animals in Experimental Research: An Ethical Dilemma?Document4 pagesUse of Animals in Experimental Research: An Ethical Dilemma?Sergiu MindruNo ratings yet
- 683 FullDocument10 pages683 FullDwina RahmawatiNo ratings yet
- Final Bio PaperDocument3 pagesFinal Bio Paperapi-240980698No ratings yet
- Älfactory Control of Qnsect Behavior: Deurobiological Explorationofthe - Moth's Nervous SystemDocument25 pagesÄlfactory Control of Qnsect Behavior: Deurobiological Explorationofthe - Moth's Nervous SystemHelena QuintNo ratings yet
- Musa Lawan WidaDocument6 pagesMusa Lawan WidaSouleyman AhmadNo ratings yet
- Rat-Park-acceptedman Bruce K. AlexanderDocument9 pagesRat-Park-acceptedman Bruce K. AlexanderLevente BalázsNo ratings yet
- Use of Animals in Experimental Research: An Ethical Dilemma?Document4 pagesUse of Animals in Experimental Research: An Ethical Dilemma?Mikela St. JohnNo ratings yet
- Behavioral and Physiological Effects of Social Isolation On MiceDocument7 pagesBehavioral and Physiological Effects of Social Isolation On Miceapi-281130314No ratings yet
- Carlos-Identificación de Feromonas Sexuales Volátiles Putativas en Perras DomésticasDocument6 pagesCarlos-Identificación de Feromonas Sexuales Volátiles Putativas en Perras DomésticasCARLOS PANCHENo ratings yet
- EthologyDocument6 pagesEthologyDheeraj K VeeranagoudarNo ratings yet
- RSPB 2010 2189 FullDocument7 pagesRSPB 2010 2189 FullLaney SommerNo ratings yet
- tmp3656 TMPDocument14 pagestmp3656 TMPFrontiersNo ratings yet
- Quignon Et Al Genome Biology 2005Document9 pagesQuignon Et Al Genome Biology 2005jean-francois.polletlive.beNo ratings yet
- Cheluvappa - Et - Al-2017 Ethics of Animal Research in Human Disease RemediationDocument14 pagesCheluvappa - Et - Al-2017 Ethics of Animal Research in Human Disease RemediationraphaelpdaNo ratings yet
- Descriptive ToxicologyDocument40 pagesDescriptive ToxicologyAhmed AbouhashemNo ratings yet
- J Exp Biol 2014 Marshall 160 1Document3 pagesJ Exp Biol 2014 Marshall 160 1GinaMariePasquiniNo ratings yet
- Aenigmamus (Rodentia: Diatomyidae) С Эволюционной Точки ЗренияDocument16 pagesAenigmamus (Rodentia: Diatomyidae) С Эволюционной Точки ЗренияFildzah AliyahNo ratings yet
- AquiloniGherardi 2010 649 249-254Document6 pagesAquiloniGherardi 2010 649 249-254Rimka BakNo ratings yet
- The Physiology of Taste in Fish Potential Implications For Feeding Stimulation and Gut Chemical SensingDocument18 pagesThe Physiology of Taste in Fish Potential Implications For Feeding Stimulation and Gut Chemical SensingDewi NurhayatiNo ratings yet
- Edexcel GCE Biology Unit 2 Exam Revision NotesDocument18 pagesEdexcel GCE Biology Unit 2 Exam Revision NotescolachauNo ratings yet
- Lucifer YellowDocument11 pagesLucifer YellowtvvsagarNo ratings yet
- Neureither Et Al 2017 (Mouse Odor Detection)Document13 pagesNeureither Et Al 2017 (Mouse Odor Detection)jean-francois.polletlive.beNo ratings yet
- Scents and Sense-Ability K9 ArticleDocument5 pagesScents and Sense-Ability K9 ArticleAlejandro MerelesNo ratings yet
- Animals in The Neighborhood Project OriginalDocument5 pagesAnimals in The Neighborhood Project OriginalFabrizio Alfredo Chitay MedinaNo ratings yet
- Show Me Your Fucking Girlbutt (Asshole)Document9 pagesShow Me Your Fucking Girlbutt (Asshole)Restroom ChemistryNo ratings yet
- PropoDocument12 pagesPropoapi-295869438No ratings yet
- Topic 4 - Biodiversity and Natural Resources PDFDocument11 pagesTopic 4 - Biodiversity and Natural Resources PDFLamia IslamNo ratings yet
- Pheromones Dogs PDFDocument25 pagesPheromones Dogs PDFRayman RushNo ratings yet
- Occasion-Setting Mechanism Color Modulates Olfactory Learning in Honeybees by AnDocument13 pagesOccasion-Setting Mechanism Color Modulates Olfactory Learning in Honeybees by AnFrontiersNo ratings yet
- Guinea Pig's Project Proposal - LineDocument15 pagesGuinea Pig's Project Proposal - LineThunyapornNo ratings yet
- The Environmental Predation Risk (Tree Sparrow)Document6 pagesThe Environmental Predation Risk (Tree Sparrow)Sophany PhaukNo ratings yet
- The Effect of Food and Music To Spatial Memory of Hamsters in T-MazeDocument10 pagesThe Effect of Food and Music To Spatial Memory of Hamsters in T-MazeMoreno MJNo ratings yet
- Lynch Et Al 2011Document16 pagesLynch Et Al 2011Diana MonroyNo ratings yet
- Aq27 Mechanism of OlfactionDocument5 pagesAq27 Mechanism of OlfactionHomeground entertainmentNo ratings yet
- Olfactory Senses in Different AnimalsDocument15 pagesOlfactory Senses in Different AnimalsNatalia avila AperadorNo ratings yet
- Psychological Reporfs, 1996,: in Al.Document10 pagesPsychological Reporfs, 1996,: in Al.dsafasdfsdafaNo ratings yet
- Animal BehaviorDocument4 pagesAnimal BehaviorifraNo ratings yet
- How Does The Zoo Environment Affect The Behaviour of Captive PrimatesDocument23 pagesHow Does The Zoo Environment Affect The Behaviour of Captive PrimatesMiguel LessaNo ratings yet
- Kraus Epley2015Document10 pagesKraus Epley2015aulia rahmahNo ratings yet
- La Evolución de La Mirmecofagia y Sus Correlatos en Ranas VenenosasDocument27 pagesLa Evolución de La Mirmecofagia y Sus Correlatos en Ranas VenenosasBrandon HerreraNo ratings yet
- Network Analysis of Gut Microbiota Literature: An Overview of The Research Landscape in Non-Human Animal StudiesDocument8 pagesNetwork Analysis of Gut Microbiota Literature: An Overview of The Research Landscape in Non-Human Animal Studiesvicrisyihab24No ratings yet
- Forensic Science International: Xiaoshan Liu, Yanwei Shi, Haiyang Wang, Runjie ZhangDocument5 pagesForensic Science International: Xiaoshan Liu, Yanwei Shi, Haiyang Wang, Runjie ZhangAndi SriNo ratings yet
- M - 30 Biochemical Taxonomy, Its Approaches, Advantages and LimitationDocument5 pagesM - 30 Biochemical Taxonomy, Its Approaches, Advantages and LimitationDr. Tapan Kr. Dutta100% (1)
- S.A.Raja Pharmacy College: Vadakkangulam 627 116Document35 pagesS.A.Raja Pharmacy College: Vadakkangulam 627 116bhanupratap singhNo ratings yet
- Combined AssignmentDocument21 pagesCombined AssignmentRohailNo ratings yet
- Volume 42 1 97 3simpanzeDocument9 pagesVolume 42 1 97 3simpanzeAnindyaMustikaNo ratings yet
- Volume 42 1 97 5Document8 pagesVolume 42 1 97 5AnindyaMustikaNo ratings yet
- Of Mice and EthicsDocument4 pagesOf Mice and EthicsGabriel IonescuNo ratings yet
- Activity 1 3Document4 pagesActivity 1 3Nicole Shane PampangaNo ratings yet
- Resumo Ouro Preto 2011 2Document10 pagesResumo Ouro Preto 2011 2Carlos Massuretti de JesusNo ratings yet
- 16783Document51 pages16783uddinnadeemNo ratings yet
- Latched, Flip-Flops, and TimersDocument36 pagesLatched, Flip-Flops, and TimersMuhammad Umair AslamNo ratings yet
- JIS G 3141: Cold-Reduced Carbon Steel Sheet and StripDocument6 pagesJIS G 3141: Cold-Reduced Carbon Steel Sheet and StripHari0% (2)
- Student Exploration: Inclined Plane - Simple MachineDocument9 pagesStudent Exploration: Inclined Plane - Simple MachineLuka MkrtichyanNo ratings yet
- UT Dallas Syllabus For Govt4396.002.08s Taught by Gregory Thielemann (Gregt)Document2 pagesUT Dallas Syllabus For Govt4396.002.08s Taught by Gregory Thielemann (Gregt)UT Dallas Provost's Technology GroupNo ratings yet
- Neonatal Mortality - A Community ApproachDocument13 pagesNeonatal Mortality - A Community ApproachJalam Singh RathoreNo ratings yet
- postedcontentadminuploadsFAQs20for20Organization PDFDocument10 pagespostedcontentadminuploadsFAQs20for20Organization PDFMohd Adil AliNo ratings yet
- Cooperative Learning: Complied By: ANGELICA T. ORDINEZADocument16 pagesCooperative Learning: Complied By: ANGELICA T. ORDINEZAAlexis Kaye GullaNo ratings yet
- National Anthems of Selected Countries: Country: United States of America Country: CanadaDocument6 pagesNational Anthems of Selected Countries: Country: United States of America Country: CanadaHappyNo ratings yet
- Brand Strategy - in B2BDocument6 pagesBrand Strategy - in B2BKrishan SahuNo ratings yet
- Delonghi Esam Series Service Info ItalyDocument10 pagesDelonghi Esam Series Service Info ItalyBrko BrkoskiNo ratings yet
- AMULDocument11 pagesAMULkeshav956No ratings yet
- Enochian Dragon Ritual PDFDocument4 pagesEnochian Dragon Ritual PDFDenis NantelNo ratings yet
- Cable To Metal Surface, Cathodic - CAHAAW3Document2 pagesCable To Metal Surface, Cathodic - CAHAAW3lhanx2No ratings yet
- The RBG Blueprint For Black Power Study Cell GuidebookDocument8 pagesThe RBG Blueprint For Black Power Study Cell GuidebookAra SparkmanNo ratings yet
- Modlist - Modlist 1.4Document145 pagesModlist - Modlist 1.4Tattorin vemariaNo ratings yet
- Sociology As A Form of Consciousness - 20231206 - 013840 - 0000Document4 pagesSociology As A Form of Consciousness - 20231206 - 013840 - 0000Gargi sharmaNo ratings yet
- Core ValuesDocument1 pageCore ValuesIan Abel AntiverosNo ratings yet
- Where We Are in Place and Time "We Are Part of The Universe and Feel Compelled To Explore It."Document1 pageWhere We Are in Place and Time "We Are Part of The Universe and Feel Compelled To Explore It."Safia-umm Suhaim- FareedNo ratings yet
- LM2576/LM2576HV Series Simple Switcher 3A Step-Down Voltage RegulatorDocument21 pagesLM2576/LM2576HV Series Simple Switcher 3A Step-Down Voltage RegulatorcgmannerheimNo ratings yet
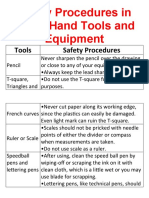
- Safety Procedures in Using Hand Tools and EquipmentDocument12 pagesSafety Procedures in Using Hand Tools and EquipmentJan IcejimenezNo ratings yet
- Comparison Between CompetitorsDocument2 pagesComparison Between Competitorsritesh singhNo ratings yet
- Hướng Dẫn Chấm: Ngày thi: 27 tháng 7 năm 2019 Thời gian làm bài: 180 phút (không kể thời gian giao đề) HDC gồm có 4 trangDocument4 pagesHướng Dẫn Chấm: Ngày thi: 27 tháng 7 năm 2019 Thời gian làm bài: 180 phút (không kể thời gian giao đề) HDC gồm có 4 trangHưng Quân VõNo ratings yet
- Pg2022 ResultDocument86 pagesPg2022 ResultkapilNo ratings yet
- ECE Companies ListDocument9 pagesECE Companies ListPolaiah Geriki100% (1)
- Development Developmental Biology EmbryologyDocument6 pagesDevelopment Developmental Biology EmbryologyBiju ThomasNo ratings yet
- Kazon Greater Predator MothershipDocument1 pageKazon Greater Predator MothershipknavealphaNo ratings yet
- Sample Resume For Supply Chain Logistics PersonDocument2 pagesSample Resume For Supply Chain Logistics PersonAmmar AbbasNo ratings yet
- Algoritm BackTracking EnglezaDocument6 pagesAlgoritm BackTracking Englezaionutz_67No ratings yet
- Nanolithography - Processing Methods PDFDocument10 pagesNanolithography - Processing Methods PDFNilesh BhardwajNo ratings yet
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (393)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (5)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (517)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (812)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseFrom EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseRating: 4.5 out of 5 stars4.5/5 (52)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (411)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (216)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (110)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (595)
- Minds Make Societies: How Cognition Explains the World Humans CreateFrom EverandMinds Make Societies: How Cognition Explains the World Humans CreateRating: 4.5 out of 5 stars4.5/5 (23)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)