You might also like
- Perkembangan Embrio AmfibiDocument22 pagesPerkembangan Embrio Amfibiratna100% (1)
- Laporan Praktikum Perkembangan Embrio AyamDocument13 pagesLaporan Praktikum Perkembangan Embrio AyamMuhammadRifqiAbdurrahmanNo ratings yet
- SEDIAAN AWETANDocument15 pagesSEDIAAN AWETANSintaNo ratings yet
- STRUKTUR AKAR MONOKOTIL DAN DIKOTILDocument3 pagesSTRUKTUR AKAR MONOKOTIL DAN DIKOTILRudy JazzerNo ratings yet
- Makalah Asal Usul Kehidupan. v1.2Document13 pagesMakalah Asal Usul Kehidupan. v1.2Frangklin BarapaNo ratings yet
- Laporan Embrio AyamDocument7 pagesLaporan Embrio AyamYudhisNo ratings yet
- Perkembangan DaunDocument9 pagesPerkembangan DaunnurulNo ratings yet
- Terjemahan Teratogenik Modern Toxycology Ed 4Document11 pagesTerjemahan Teratogenik Modern Toxycology Ed 4asmha997No ratings yet
- PALYNOLOGYDocument43 pagesPALYNOLOGYJuniSianiparNo ratings yet
- Struktur Anatomi Sistem Pernapasan Pada VertebratalDocument24 pagesStruktur Anatomi Sistem Pernapasan Pada VertebratalvanisNo ratings yet
- Viranika - K4319086 - C - Whole Mount HewanDocument16 pagesViranika - K4319086 - C - Whole Mount HewanViranika Devi Talita MahardaniNo ratings yet
- Makalah Embriologi Hewan Pembelahan CleaDocument20 pagesMakalah Embriologi Hewan Pembelahan CleaMartinus BuddhyNo ratings yet
- REPRODUKSIDocument7 pagesREPRODUKSIGusti Bagus Wibawa PutraNo ratings yet
- Laporan Embrio KatakDocument7 pagesLaporan Embrio KatakYudhis0% (1)
- CLEAVAGE DAN PEMBELAHAN SEL PADA ZIGOTDocument30 pagesCLEAVAGE DAN PEMBELAHAN SEL PADA ZIGOTJulista NadotNo ratings yet
- GAMETDocument17 pagesGAMETadnan_unm3177100% (2)
- LAPORAN GENETIKA PoliploidiDocument10 pagesLAPORAN GENETIKA PoliploidiRizki PrasNo ratings yet
- Gastrulasi dan Pembentukan Lapisan GerminalDocument10 pagesGastrulasi dan Pembentukan Lapisan GerminalTasafima TesariNo ratings yet
- Laporan Resmi Genetika Alel GandaDocument14 pagesLaporan Resmi Genetika Alel GandaJessicavonRichthofenNo ratings yet
- SPH Prk. 2Document25 pagesSPH Prk. 2Waode Narvia0% (1)
- Tugas 5 Ovum TypeDocument2 pagesTugas 5 Ovum Typesakinatuz zakiahNo ratings yet
- Genetika Ternak DrosophilaDocument9 pagesGenetika Ternak DrosophilaEnikaaNo ratings yet
- Plasmid Dan EpisomDocument6 pagesPlasmid Dan EpisomDevintaNo ratings yet
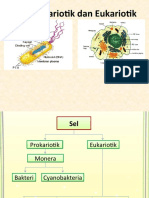
- Pertemuan 3. Sel Prokariotik Dan Sel EukariotikDocument63 pagesPertemuan 3. Sel Prokariotik Dan Sel EukariotikHilda SabrinaNo ratings yet
- Transposable Element GeneticDocument23 pagesTransposable Element GeneticOki Osaka100% (1)
- Pengembangan Embrio AyamDocument9 pagesPengembangan Embrio AyamDiah Nanda Utari100% (1)
- SPT - Sistem Simpodian Dan Anatomi NodalDocument12 pagesSPT - Sistem Simpodian Dan Anatomi NodalAhmad RusdyNo ratings yet
- TERATOGEN DAN MEDIKASIDocument40 pagesTERATOGEN DAN MEDIKASIfujimeister100% (1)
- Proyek Gelap Terang Kel.2 Rizky Pipit Off.B2Document31 pagesProyek Gelap Terang Kel.2 Rizky Pipit Off.B2rizkyalfarizyNo ratings yet
- BIOTEKNOLOGI EMBRIOLOGIDocument21 pagesBIOTEKNOLOGI EMBRIOLOGIEka Dian SofianaNo ratings yet
- Revisi Mendel 2Document34 pagesRevisi Mendel 2Naimmatus SholikhahNo ratings yet
- Tugas Online Mata Kuliah Struktur Dan Perkembangan Hewan IIDocument2 pagesTugas Online Mata Kuliah Struktur Dan Perkembangan Hewan IIgraciaNo ratings yet
- Evolusi MolekulerDocument15 pagesEvolusi Molekulerkoko murdiantoNo ratings yet
- Generasi Sporofit Dan Sporogenesis GymnospermaeDocument3 pagesGenerasi Sporofit Dan Sporogenesis GymnospermaeNaya'bee Pengend Dimengerty100% (1)
- Kelainan Embrio Ok 2Document33 pagesKelainan Embrio Ok 2yuliadewisaputri0% (1)
- Embrio KatakDocument14 pagesEmbrio KatakFadila TurahmahNo ratings yet
- AmfibiDocument10 pagesAmfibiAlbert Sanada YapriNo ratings yet
- Plasentasi BungaDocument7 pagesPlasentasi BungaNeni Inayah100% (2)
- ORGAN REPRODUKSIDocument6 pagesORGAN REPRODUKSIwahyuddinNo ratings yet
- Metamorfosis KatakDocument10 pagesMetamorfosis Katakrahma adilahNo ratings yet
- Makalah SPH2 Regenerasi Kel.5Document14 pagesMakalah SPH2 Regenerasi Kel.5AnasNo ratings yet
- MonohibridDocument7 pagesMonohibridBerty Veibrita SinagaNo ratings yet
- Nakalah Nematoda Kelompok 1 KuDocument15 pagesNakalah Nematoda Kelompok 1 KuWaOde Jumriani SittiEkaNo ratings yet
- Pembuatan Preparat Wholemount Embrio Ayam KampungDocument16 pagesPembuatan Preparat Wholemount Embrio Ayam KampungicsniNo ratings yet
- KECENDERUNGAN KAWIN DROSOPHILADocument38 pagesKECENDERUNGAN KAWIN DROSOPHILADiah Ajeng MustikariniNo ratings yet
- Malformasi Kongenital 2Document12 pagesMalformasi Kongenital 2AJihan Anggia JauzaNo ratings yet
- Ayam PDFDocument18 pagesAyam PDFShabrina LieNo ratings yet
- PERKEMBANGAN EMBRIO AYAM WHOLEMOUNTDocument20 pagesPERKEMBANGAN EMBRIO AYAM WHOLEMOUNTMega FrezaaNo ratings yet
- Makalah Ekologi (Faktor Biotik)Document14 pagesMakalah Ekologi (Faktor Biotik)Ghio NajoanNo ratings yet
- Data Pengamatan Embrio KatakDocument7 pagesData Pengamatan Embrio KataklusyNo ratings yet
- Preparat PollenDocument5 pagesPreparat PollenardihaniNo ratings yet
- Proposal Mita FENOMENA PAUTAN KELAMIN PERSILANGAN Drosophila Melanogaster STRAIN NDocument11 pagesProposal Mita FENOMENA PAUTAN KELAMIN PERSILANGAN Drosophila Melanogaster STRAIN NLyana BerlyNo ratings yet
- Aprina Dilla - Tugas PteridophytaDocument12 pagesAprina Dilla - Tugas Pteridophytaaprina dillaNo ratings yet
- Magnoliopsida Prak 2Document18 pagesMagnoliopsida Prak 2Aiiu MilawatiNo ratings yet
- Dwi Laporan SPH Embrio AyamDocument12 pagesDwi Laporan SPH Embrio AyamDwi Narlinda100% (1)
- Resume EkstrakromosomalDocument9 pagesResume EkstrakromosomalAstrinoPurmannaNo ratings yet
- Perkembangan Embrio AvesDocument20 pagesPerkembangan Embrio AvesriscaNo ratings yet
- EMBRIOGENESIS AVESDocument22 pagesEMBRIOGENESIS AVESPatimmNo ratings yet
- PERKEMBANGAN EMBRIO AVESDocument19 pagesPERKEMBANGAN EMBRIO AVESChairil AkmalNo ratings yet
- Perkembangan Embrio AvesDocument22 pagesPerkembangan Embrio AvesTania Rizki AmeliaNo ratings yet
- STRATEGIC CONTENTSDocument41 pagesSTRATEGIC CONTENTSMifroatin ShawolocketElforeverNo ratings yet
- RPP KD 2Document126 pagesRPP KD 2Mifroatin ShawolocketElforeverNo ratings yet
- Kisi-Kisi PH 2 Bahasa JawaDocument1 pageKisi-Kisi PH 2 Bahasa JawaMifroatin ShawolocketElforeverNo ratings yet
- Nilai MutlakDocument5 pagesNilai MutlakMifroatin ShawolocketElforeverNo ratings yet
- Resume 2 FixDocument11 pagesResume 2 FixMifroatin ShawolocketElforeverNo ratings yet
- LKS SISTEM PERKEMBANGBIAAKANDocument2 pagesLKS SISTEM PERKEMBANGBIAAKANMifroatin ShawolocketElforeverNo ratings yet
- Pembelahan SelDocument21 pagesPembelahan SelMifroatin ShawolocketElforeverNo ratings yet
- KORESPONDENSIDocument194 pagesKORESPONDENSIMifroatin ShawolocketElforeverNo ratings yet
- Nilai MutlakDocument5 pagesNilai MutlakMifroatin ShawolocketElforeverNo ratings yet
- RPP Dan SoalDocument11 pagesRPP Dan SoalMifroatin ShawolocketElforeverNo ratings yet
- AttachmentDocument3 pagesAttachmentMifroatin ShawolocketElforeverNo ratings yet
- Ekspresi Gen Dan PerkembangannyaDocument7 pagesEkspresi Gen Dan PerkembangannyaAfif Iffy DeraNo ratings yet
- LKS SISTEM PERKEMBANGBIAAKANDocument2 pagesLKS SISTEM PERKEMBANGBIAAKANMifroatin ShawolocketElforeverNo ratings yet
- Pengadaan Peralatan Laboratorium IpaDocument1 pagePengadaan Peralatan Laboratorium IpaMifroatin ShawolocketElforeverNo ratings yet
- LKS SISTEM PERKEMBANGBIAAKANDocument2 pagesLKS SISTEM PERKEMBANGBIAAKANMifroatin ShawolocketElforeverNo ratings yet
- Mind MapDocument1 pageMind MapMifroatin ShawolocketElforeverNo ratings yet
- Makalah 1Document8 pagesMakalah 1Mifroatin ShawolocketElforeverNo ratings yet
- Kementerian LingkunganDocument2 pagesKementerian LingkunganMifroatin ShawolocketElforeverNo ratings yet
- AttachmentDocument3 pagesAttachmentMifroatin ShawolocketElforeverNo ratings yet
- Contoh AnalisisDocument5 pagesContoh AnalisisMifroatin ShawolocketElforeverNo ratings yet
- PertanyaanDocument1 pagePertanyaanMifroatin ShawolocketElforeverNo ratings yet
- Contoh Undangan TahlilDocument1 pageContoh Undangan TahlilToyaNo ratings yet
- Analisis Atikel PenilaianDocument5 pagesAnalisis Atikel PenilaianMifroatin ShawolocketElforeverNo ratings yet
- Foto ZulaDocument5 pagesFoto ZulaMifroatin ShawolocketElforeverNo ratings yet
- ManfaatDocument2 pagesManfaatMifroatin ShawolocketElforeverNo ratings yet
- LksDocument1 pageLksMifroatin ShawolocketElforeverNo ratings yet
- Analisis Pembahasan 2Document1 pageAnalisis Pembahasan 2Mifroatin ShawolocketElforeverNo ratings yet
- IsiDocument9 pagesIsiMifroatin ShawolocketElforeverNo ratings yet
- Mind Map CoverDocument1 pageMind Map CoverMifroatin ShawolocketElforeverNo ratings yet
- Repost Silabus IPA Revisi 2016Document39 pagesRepost Silabus IPA Revisi 2016Mifroatin ShawolocketElforeverNo ratings yet