You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- File Synchronization With Vector Time Pairs: Computer Science and Artificial Intelligence Laboratory Technical ReportDocument15 pagesFile Synchronization With Vector Time Pairs: Computer Science and Artificial Intelligence Laboratory Technical Reportsatch76No ratings yet
- Bela Bartok S Concerto For OrchestraDocument43 pagesBela Bartok S Concerto For Orchestrasatch76No ratings yet
- Cycles and Coltrane Changes Giant StepsDocument12 pagesCycles and Coltrane Changes Giant Stepsprocopio99No ratings yet
- Rootless ExercisesDocument1 pageRootless Exercisessatch76No ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Heat Stress Fact SheetDocument3 pagesHeat Stress Fact SheetjacquestsNo ratings yet
- Lca Mangatarem1 PDFDocument58 pagesLca Mangatarem1 PDFBea 'Jane0% (1)
- Spheciospongia Vesparium, Commonly Known As The LoggerheadDocument2 pagesSpheciospongia Vesparium, Commonly Known As The Loggerheadandrew_yeap_2No ratings yet
- Chapter 8 Physical Development BerkDocument14 pagesChapter 8 Physical Development BerkMARIAH HOLLETTNo ratings yet
- Acute Effects of Ayahuasca in A Juvenile Non-Human Primate Model of DepressionDocument9 pagesAcute Effects of Ayahuasca in A Juvenile Non-Human Primate Model of DepressionGonzaloBimonteNo ratings yet
- A Brief History of HemophiliaDocument127 pagesA Brief History of HemophiliaApurba SahaNo ratings yet
- Physiology - Last Minute Revision Points: DR - SreetejaDocument25 pagesPhysiology - Last Minute Revision Points: DR - SreetejaBanu KubendiranNo ratings yet
- Non-Critically Ill Hyperglycemia ManagementDocument39 pagesNon-Critically Ill Hyperglycemia ManagementHerry KongkoNo ratings yet
- Pharmacogenetics Drug Drug Interactions1Document16 pagesPharmacogenetics Drug Drug Interactions1Marfu'ah Mar'ahNo ratings yet
- TissuesDocument23 pagesTissuesWubalem DestaNo ratings yet
- PD22 Hap1 L03Document33 pagesPD22 Hap1 L03Ka Yan LAUNo ratings yet
- 1 - CPomar - Pork Carcass Evaluation and Grading-2Document50 pages1 - CPomar - Pork Carcass Evaluation and Grading-2Jacob AguilarNo ratings yet
- Composite Materials For Biomedical Applications: A ReviewDocument17 pagesComposite Materials For Biomedical Applications: A ReviewDuc Anh NguyenNo ratings yet
- DocumentDocument7 pagesDocumentDébàshis Dash50% (2)
- Comprehension QuestionDocument2 pagesComprehension QuestionAhmad Safuan Bin Mohd Sukri -No ratings yet
- 1 2 Receptor Tyrosine Kinases 2021Document44 pages1 2 Receptor Tyrosine Kinases 2021Bilakovics NoemiNo ratings yet
- Fphar 12 741623Document17 pagesFphar 12 741623Hendri AldratNo ratings yet
- Karakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiDocument7 pagesKarakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiAlexanderNo ratings yet
- Morphological Characterization of Pichia Guilliermondii and Saccharomyces Cerevisiae Yeast and Their Effects On Adherence of IntesDocument7 pagesMorphological Characterization of Pichia Guilliermondii and Saccharomyces Cerevisiae Yeast and Their Effects On Adherence of IntesAnonymous Y83Rx25No ratings yet
- Geriatric PanchakarmaDocument24 pagesGeriatric Panchakarmaksr prasadNo ratings yet
- To Study The Quantity Case in Present in Different Sample of MilkDocument7 pagesTo Study The Quantity Case in Present in Different Sample of Milkbelly4u100% (2)
- Review ImportanteDocument23 pagesReview Importantejano castroNo ratings yet
- Rmsa NotificationDocument17 pagesRmsa NotificationSrinivasu RongaliNo ratings yet
- Myofascial Meridians PDFDocument9 pagesMyofascial Meridians PDFPedja Ley100% (3)
- EnvironmentDocument629 pagesEnvironmentSanchay SaxenaNo ratings yet
- FCPS SCERETS Rabia Ali Final - UPDATED Version 2014Document9 pagesFCPS SCERETS Rabia Ali Final - UPDATED Version 2014Akas Rehman100% (4)
- Chapter 15 - TeratogenesisDocument29 pagesChapter 15 - TeratogenesisMaula AhmadNo ratings yet
- 3. The structure and function of large biological moleculesDocument62 pages3. The structure and function of large biological moleculesOussema Ben KasdallahNo ratings yet
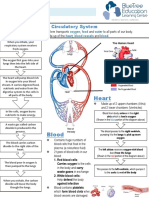
- Circulatory System Notes SLDocument1 pageCirculatory System Notes SLRathna PalaniappanNo ratings yet
- Establishment of Cacao Nursery and ManagementDocument121 pagesEstablishment of Cacao Nursery and ManagementPlantacion de Sikwate100% (29)