You might also like
- Healthcare Impact of Covid-19 Epidemic in IndiaDocument2 pagesHealthcare Impact of Covid-19 Epidemic in IndiaAnupam GuptaNo ratings yet
- Covid - 19Document1 pageCovid - 19Anupam GuptaNo ratings yet
- I MCSE-011: Time: 3 Hours Maximum Marks: 100Document2 pagesI MCSE-011: Time: 3 Hours Maximum Marks: 100Anupam GuptaNo ratings yet
- Mcse 004Document5 pagesMcse 004Saroj SinghNo ratings yet
- Instructions-Activity 6 Individual Work FINALDocument2 pagesInstructions-Activity 6 Individual Work FINALAnupam GuptaNo ratings yet
- Basic ComputerDocument59 pagesBasic ComputerAnupam GuptaNo ratings yet
- Script Unit 3: What Will You Learn in Unit 3?Document4 pagesScript Unit 3: What Will You Learn in Unit 3?Anupam GuptaNo ratings yet
- First Love PDFDocument189 pagesFirst Love PDFnityaNo ratings yet
- MCSE-004, Dec17Document158 pagesMCSE-004, Dec17Anupam GuptaNo ratings yet
- Mcse 004Document5 pagesMcse 004Saroj SinghNo ratings yet
- Importance of Assessment in Education SystemDocument7 pagesImportance of Assessment in Education SystemAnupam GuptaNo ratings yet
- Importance of Assessment in Education SystemDocument7 pagesImportance of Assessment in Education SystemAnupam GuptaNo ratings yet
- Pooja CurrentDocument1 pagePooja CurrentAnupam GuptaNo ratings yet
- PISA for Development Assessment GuideDocument15 pagesPISA for Development Assessment GuideAnupam GuptaNo ratings yet
- Ip26 Uis Cost of Measuring Learning 2a.Document15 pagesIp26 Uis Cost of Measuring Learning 2a.Anupam GuptaNo ratings yet
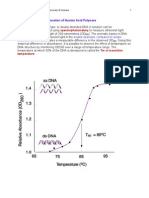
- Factors Affecting DNA Denaturation and RenaturationDocument4 pagesFactors Affecting DNA Denaturation and RenaturationVenkateswarlu Yadavalli100% (2)
- Glyc o Conjugate SDocument3 pagesGlyc o Conjugate SAnupam GuptaNo ratings yet
- Protochordates 141124083819 Conversion Gate02 2Document21 pagesProtochordates 141124083819 Conversion Gate02 2Anupam GuptaNo ratings yet
- State Writeup NEUPA - 9!11!2016Document20 pagesState Writeup NEUPA - 9!11!2016Anupam GuptaNo ratings yet
- Objective Type QuestionsDocument11 pagesObjective Type QuestionsAnupam GuptaNo ratings yet
- My First Teaching ExperienceDocument10 pagesMy First Teaching ExperienceAnupam GuptaNo ratings yet
- Students' Use of Facebook For Peer-To-Peer LearningDocument17 pagesStudents' Use of Facebook For Peer-To-Peer LearningAnupam GuptaNo ratings yet
- Slide Outline CDocument1 pageSlide Outline CAnupam GuptaNo ratings yet
- Mantra4success motto nurturing students quality education skills dreamsDocument2 pagesMantra4success motto nurturing students quality education skills dreamsAnupam GuptaNo ratings yet
- HCLDocument2 pagesHCLAnupam GuptaNo ratings yet
- Hackinf PDFDocument4 pagesHackinf PDFAnupam GuptaNo ratings yet
- National Disaster Management Authority (NDMA) : Government of IndiaDocument1 pageNational Disaster Management Authority (NDMA) : Government of IndiaAnupam GuptaNo ratings yet
- Bioinformatics: Mark Gerstein, Yale University Bioinfo - Mbb.yale - Edu/mbb452aDocument21 pagesBioinformatics: Mark Gerstein, Yale University Bioinfo - Mbb.yale - Edu/mbb452azuhalNo ratings yet
- HCL Candidate Training AgreementDocument14 pagesHCL Candidate Training AgreementAnupam GuptaNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5782)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- GKB Data Analytics Mandate Part 2Document9 pagesGKB Data Analytics Mandate Part 2Vilma TejadaNo ratings yet
- TLS landing system enables accessible airportsDocument12 pagesTLS landing system enables accessible airportsHasan MalikNo ratings yet
- Artifact c2 - Group Counseling ProposalDocument21 pagesArtifact c2 - Group Counseling Proposalapi-310924257No ratings yet
- Reliability Centered Maintenance ExplainationDocument6 pagesReliability Centered Maintenance Explainationambuenaflor100% (1)
- ISO 15184 Paint Hardness TesterDocument6 pagesISO 15184 Paint Hardness TesterAvinash SilimkarNo ratings yet
- ServiceNow Certified System Administrator CSA Practice Test Set 2Document76 pagesServiceNow Certified System Administrator CSA Practice Test Set 2Apoorv DiwanNo ratings yet
- CargadoresDocument3 pagesCargadoresAmber SmithNo ratings yet
- Different Types of AstrologyDocument16 pagesDifferent Types of AstrologyastrologerrajeevjiNo ratings yet
- Oracle NetSuite To Oracle PBCS IntegrationDocument11 pagesOracle NetSuite To Oracle PBCS IntegrationAmit SharmaNo ratings yet
- Field Trip 2 - Coreslab Structures Inc Essay - FinalDocument11 pagesField Trip 2 - Coreslab Structures Inc Essay - Finalmmg009No ratings yet
- Revised New PTLLS Assignment 1 Levels 3 and 4 Nov 2011Document3 pagesRevised New PTLLS Assignment 1 Levels 3 and 4 Nov 2011dave_perry189218No ratings yet
- 4aa0 7171enwDocument22 pages4aa0 7171enwOng Ka HuiNo ratings yet
- Law Movie Analysis Erin BrockovichDocument3 pagesLaw Movie Analysis Erin Brockovichshaju0% (3)
- BLACK BOOK-PDF GautamiDocument63 pagesBLACK BOOK-PDF GautamiHemant DasNo ratings yet
- FT 180aDocument60 pagesFT 180aRobert/YG2AKR75% (4)
- MASAR - Volume 02Document214 pagesMASAR - Volume 02Khaled MamdouhNo ratings yet
- Python Programming For Beginners 2 Books in 1 B0B7QPFY8KDocument243 pagesPython Programming For Beginners 2 Books in 1 B0B7QPFY8KVaradaraju ThirunavukkarasanNo ratings yet
- Pe 4.1 - SwimmingDocument21 pagesPe 4.1 - SwimmingJacinthe Angelou D. PeñalosaNo ratings yet
- Zinkgruvan Suecia Polimetálico TechRep - 2017Document178 pagesZinkgruvan Suecia Polimetálico TechRep - 2017manudemNo ratings yet
- Form Initial AssessmentDocument2 pagesForm Initial Assessmentari andiNo ratings yet
- Caterpillar Oil Filter Pehj0068-02Document2 pagesCaterpillar Oil Filter Pehj0068-02Parinpa Ketar100% (1)
- Labprograms 1A2A3ADocument11 pagesLabprograms 1A2A3ARajiv SethNo ratings yet
- Case Study Dorothea Orem (Francar Jade For Sbmission) - TFNDocument2 pagesCase Study Dorothea Orem (Francar Jade For Sbmission) - TFNFrancar Jade De VeraNo ratings yet
- Relevant CostingDocument5 pagesRelevant CostingRechelleNo ratings yet
- MoRTH CompleteDocument725 pagesMoRTH CompleteSekharmantri Sitapati85% (71)
- LG Beko 28cz10rx Service ManualDocument32 pagesLG Beko 28cz10rx Service ManualAlan ScarfidiNo ratings yet
- ViewPoint - Global Shopping Centre DevelopmentDocument9 pagesViewPoint - Global Shopping Centre DevelopmentAiohue FalconiNo ratings yet
- The Development Cultural Studies in BritishDocument6 pagesThe Development Cultural Studies in BritishIeie HollyNo ratings yet
- Shaft Alignment Tools TMEA Series: SKF Product Data SheetDocument3 pagesShaft Alignment Tools TMEA Series: SKF Product Data SheetGts ImportadoraNo ratings yet
- Elements of PoetryDocument50 pagesElements of PoetryBeing LoisNo ratings yet