You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5784)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
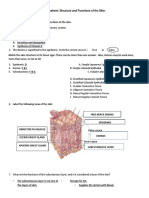
- Integumentary System Worksheet 1Document3 pagesIntegumentary System Worksheet 1Bea Lou SabadoNo ratings yet
- Article: A Ges An e EnviDocument5 pagesArticle: A Ges An e EnviKh SajolNo ratings yet
- Source of Infections 1. Endogenous SourceDocument6 pagesSource of Infections 1. Endogenous SourceDeepu VijayaBhanuNo ratings yet
- Examen Tema 1 y 2 InglesDocument7 pagesExamen Tema 1 y 2 InglesmamenNo ratings yet
- Soal Uts IngDocument6 pagesSoal Uts Ingmiftah nurohmanNo ratings yet
- Premature Cervical DilationDocument22 pagesPremature Cervical DilationJanelle Lois EscolanoNo ratings yet
- Animal Farm VocabularyDocument5 pagesAnimal Farm VocabularyMicaela DavisNo ratings yet
- Patho Physiology Ovarian CystDocument3 pagesPatho Physiology Ovarian CystSig Deliso100% (1)
- Paediatric Life SupportDocument88 pagesPaediatric Life SupportmaharaniNo ratings yet
- Latihan Soal Bahasa Inggris Kelas 1Document5 pagesLatihan Soal Bahasa Inggris Kelas 1oktavia kiranaNo ratings yet
- Protein WoolDocument11 pagesProtein WoolAnkita PrakashNo ratings yet
- Embriologia Del CerdoDocument11 pagesEmbriologia Del CerdoTadase Hotori0% (1)
- Hyper Tension and Diabetes Mellitus: Gurushankar Govindarajan James R Sowers Craig S StumpDocument7 pagesHyper Tension and Diabetes Mellitus: Gurushankar Govindarajan James R Sowers Craig S StumpAdina StoianNo ratings yet
- Wild Places Spark Wild BehaviorsDocument6 pagesWild Places Spark Wild BehaviorsqihongNo ratings yet
- Final I. Competency-checklist-VenipunctureDocument2 pagesFinal I. Competency-checklist-Venipuncturejohn100% (3)
- Intensive Writing AssessmentDocument4 pagesIntensive Writing Assessmentapi-331504072No ratings yet
- Nature Walk Assignment 3 Humans and Nature: Submitted By: Abinaya Thiruselvam BFT/18/487Document8 pagesNature Walk Assignment 3 Humans and Nature: Submitted By: Abinaya Thiruselvam BFT/18/487abiNo ratings yet
- Amazing Kid 1Document8 pagesAmazing Kid 1Harry MartinezNo ratings yet
- Anatomy I MCQDocument10 pagesAnatomy I MCQARIJITBANIK36100% (3)
- Nri1201 Hiv PosterDocument1 pageNri1201 Hiv Posteranirbanmanna88320100% (1)
- Graceful hips and deep bosomDocument41 pagesGraceful hips and deep bosomrkamin5No ratings yet
- Indian Writing in EnglishDocument4 pagesIndian Writing in EnglishAnonymous Udy9PDNo ratings yet
- Crochet Now March 2019Document128 pagesCrochet Now March 2019Chris Chrys100% (23)
- Beyond The Dragon of Icespire Peak - Part 1 - Storm Lord's Wrath - GM BinderDocument62 pagesBeyond The Dragon of Icespire Peak - Part 1 - Storm Lord's Wrath - GM BinderSébastien MIKA100% (4)
- IELTS Speaking - Useful Idioms and ProverbsDocument6 pagesIELTS Speaking - Useful Idioms and ProverbsPratik Paudel100% (2)
- Evolution and Natural SelectionDocument7 pagesEvolution and Natural SelectionMichelleneChenTadleNo ratings yet
- Animal EyesDocument291 pagesAnimal EyesjesusNo ratings yet
- Oral ContraceptivesDocument25 pagesOral ContraceptivesAdhitya Yudha MaulanaNo ratings yet
- The FOOD CHAIN by Moraa Siika XXXDocument5 pagesThe FOOD CHAIN by Moraa Siika XXXakamy5kNo ratings yet
- Understanding Culture, Society and Politics: First Quarter Module 4 - Week 4Document12 pagesUnderstanding Culture, Society and Politics: First Quarter Module 4 - Week 4Rose DiNo ratings yet