You might also like
- Reacciones FebrilesDocument3 pagesReacciones FebrilesQUIMICO CLINICO WILLIANS SANCHEZ89% (18)
- Diferencia Entre Célula Eucariota y ProcariotaDocument11 pagesDiferencia Entre Célula Eucariota y ProcariotaLeydi Yudith Angarita Bautista89% (180)
- La PielDocument2 pagesLa PielMely FranzottiNo ratings yet
- Agua y Cuerpo HumanoDocument7 pagesAgua y Cuerpo HumanopauloarieuNo ratings yet
- Manual de Primeros Auxilios en El Laboratorio (Gutiérrez)Document19 pagesManual de Primeros Auxilios en El Laboratorio (Gutiérrez)Carlos Canel EgremiNo ratings yet
- CRIMINALISTICA-Allan Arburola ValverdeDocument80 pagesCRIMINALISTICA-Allan Arburola ValverdeHarley Ospina100% (2)
- Manual de Triage de UrgenciasDocument93 pagesManual de Triage de UrgenciashannaNo ratings yet
- Principales Síndromes Del Sistema UrinarioDocument9 pagesPrincipales Síndromes Del Sistema UrinarioOscar GutierrezNo ratings yet
- Glomerulopatías SecundariasDocument74 pagesGlomerulopatías SecundariasMonica Cochachin100% (1)
- Medida Epidemiologica Incidencia y EjerciciosDocument7 pagesMedida Epidemiologica Incidencia y EjerciciosMalejaPerdomoOspinaNo ratings yet
- Acidos - Nucleicos Estructura y FuncionDocument9 pagesAcidos - Nucleicos Estructura y FuncionArdwin Del PozoNo ratings yet
- Sistema InmunologicoDocument22 pagesSistema InmunologicoAnibal Antonio MenjivarNo ratings yet
- Tipos de ELISA, ¿Conoces Las Diferencias - Abyntek Biopharma PDFDocument1 pageTipos de ELISA, ¿Conoces Las Diferencias - Abyntek Biopharma PDFDariforiNo ratings yet
- Alteraciones Hematológicas Durante El EmbarazoDocument29 pagesAlteraciones Hematológicas Durante El EmbarazoAna Luz Muñoz CornejoNo ratings yet
- VacutainerDocument3 pagesVacutainerAhtziri MartinezNo ratings yet
- Concepto de Legislacion Internacional, Etapas Del Comercio Internacional. Power PointDocument9 pagesConcepto de Legislacion Internacional, Etapas Del Comercio Internacional. Power PointDayana VargasNo ratings yet
- BHC Valores NormalesDocument1 pageBHC Valores NormalesJeessii CiicleerNo ratings yet
- Facie de PajaritoDocument4 pagesFacie de PajaritoIvette DiazNo ratings yet
- Leishmaniasis Tegumentaria AmericanaDocument26 pagesLeishmaniasis Tegumentaria Americanadaniel c.No ratings yet
- PropanonaDocument3 pagesPropanonaAnonymous bz6kuHaJNo ratings yet
- Hipertension IntracraneanaDocument11 pagesHipertension IntracraneanaJavier Colli ReyesNo ratings yet
- Epoc DescompensadoDocument26 pagesEpoc DescompensadoAlvaroPoncedeLeón50% (2)
- Transtornos PlaquetariosDocument18 pagesTranstornos PlaquetariosEsthefany HidalgoNo ratings yet
- Etiologia de La Hipertension Arterial Sistemica PDFDocument2 pagesEtiologia de La Hipertension Arterial Sistemica PDFGaryNo ratings yet
- Urgencias CardiovascularesDocument39 pagesUrgencias CardiovascularesJuleisy Xiomi Mostajo RuizNo ratings yet
- Leucemia Mieloide Crónica (LMC) - Hematología y Oncología - Manual Merck Versión para ProfesionalesDocument6 pagesLeucemia Mieloide Crónica (LMC) - Hematología y Oncología - Manual Merck Versión para ProfesionalesAbner Hernandez LopezNo ratings yet
- Movimientos AnatomicosDocument6 pagesMovimientos AnatomicosClara Rosa Campos DuranNo ratings yet
- Los 20 Aminoacidos Mas ImportantesDocument4 pagesLos 20 Aminoacidos Mas Importantesrmtdla64No ratings yet
- Regulación de La Presión Arterial Tema 5Document9 pagesRegulación de La Presión Arterial Tema 5Richard BuendiaNo ratings yet
- Traumatismos Hepáticos PDFDocument7 pagesTraumatismos Hepáticos PDFGeorgette Minerva BocaneyNo ratings yet
- Casos Clínicos de Excitabilidad CelularDocument16 pagesCasos Clínicos de Excitabilidad CelularDamaris Castillo DuarteNo ratings yet
- Qué Pasa Con El Cuerpo Cuando Hacemos EjercicioDocument7 pagesQué Pasa Con El Cuerpo Cuando Hacemos EjercicioYesi BenitezNo ratings yet
- ElectrocardiogramaDocument20 pagesElectrocardiogramaanriodonNo ratings yet
- Adn MitocondrialDocument36 pagesAdn MitocondrialDokhur100% (1)
- LinfomasDocument30 pagesLinfomasYo MisMo Antonio Gonzalez CabralNo ratings yet
- Tema 13. Sistema HemolinfaticoDocument19 pagesTema 13. Sistema HemolinfaticoCarolina Longa100% (1)
- MCP HipertroficaDocument9 pagesMCP HipertroficaFranco PacelloNo ratings yet
- Inflamación de Amígdalas y TratamientoDocument3 pagesInflamación de Amígdalas y Tratamientoletylv2104013No ratings yet
- Libro PARASITOLOGÍADocument106 pagesLibro PARASITOLOGÍARicardo Hernandez CidesNo ratings yet
- Empiema PleuralDocument7 pagesEmpiema PleuralGaleno03No ratings yet
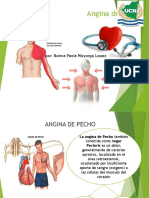
- Angina de Pecho FinalDocument20 pagesAngina de Pecho FinalDylang Yossue Diaz OrtegaNo ratings yet
- Anatomia de La Piel Cicatrizacion y HeridasDocument48 pagesAnatomia de La Piel Cicatrizacion y HeridasAdRian CMNo ratings yet
- Monografias de Enfermedad CerebrovascularDocument22 pagesMonografias de Enfermedad Cerebrovascularcalber18No ratings yet
- Fisiopatología Del Sistema Hematopoyético y HemostáticoDocument7 pagesFisiopatología Del Sistema Hematopoyético y HemostáticoDaylis YepezNo ratings yet
- Vector de VelocidadDocument13 pagesVector de Velocidadliz garciaNo ratings yet
- Caso Clínico ECVDocument16 pagesCaso Clínico ECVMarvin VivasNo ratings yet
- EPÍLEPSIADocument28 pagesEPÍLEPSIAana estefania pelayo100% (2)
- Vectores - Movimiento - DinámicaDocument15 pagesVectores - Movimiento - DinámicaAttom Nivelaciones AcadémicasNo ratings yet
- Bicarbonato en UciDocument10 pagesBicarbonato en Uciadrian mendoza croesNo ratings yet
- Mexico Vacutainer - Acerca de Los Tubos BD Vacutainer®Document13 pagesMexico Vacutainer - Acerca de Los Tubos BD Vacutainer®jorge.leo.94No ratings yet
- ¿QUÉ SON LOS MARCADORES TUMORALES ? ¿CUANTOS TIPOS HAY? - Medicina Mnemotecnias PDFDocument10 pages¿QUÉ SON LOS MARCADORES TUMORALES ? ¿CUANTOS TIPOS HAY? - Medicina Mnemotecnias PDFrafaelNo ratings yet
- Insuficiencia CardíacaDocument3 pagesInsuficiencia CardíacaThalia BenitoNo ratings yet
- Caso Clinico Cardiopatia Isquemica JCDocument16 pagesCaso Clinico Cardiopatia Isquemica JCJael NaynNo ratings yet
- Hematología CompletaDocument5 pagesHematología Completamayrel21No ratings yet
- Valoracion de Parametros Vitales Invasivos y No Invasivos 4Document22 pagesValoracion de Parametros Vitales Invasivos y No Invasivos 4Agustin Abraham Velazquez MartinezNo ratings yet
- Leucemia Linfatica AgudaDocument10 pagesLeucemia Linfatica AgudaAnonymous GNiOkl3nslNo ratings yet
- Aclaramiento o Depuración RenalDocument2 pagesAclaramiento o Depuración RenalAndrea RojasNo ratings yet
- Tema 14. Sistema Calicreína-Cininonego-Cinina MODIFICADODocument6 pagesTema 14. Sistema Calicreína-Cininonego-Cinina MODIFICADOLuis MartinezNo ratings yet
- Lineas ToracicasDocument2 pagesLineas ToracicasSllavko K. KallfaNo ratings yet
- 38 Quimio AntineoplasicaDocument11 pages38 Quimio Antineoplasicajuan laraNo ratings yet
- Movimientos AnatomicosDocument4 pagesMovimientos AnatomicosYuleixis RamírezlQa?inklpñkuNo ratings yet
- Células Madre Hematopoyéticas: Provided by Real Academia Nacional de Farmacia: Portal PublicacionesDocument23 pagesCélulas Madre Hematopoyéticas: Provided by Real Academia Nacional de Farmacia: Portal PublicacionesNicole Betancourt AlvarezNo ratings yet
- Las Células Madre HematopoyéticasDocument6 pagesLas Células Madre HematopoyéticasReyna_Maridali_9176No ratings yet
- Celulas MadreDocument9 pagesCelulas Madrearmanditocote94No ratings yet
- Control de Calidad en La Industria Farmacéutica PDFDocument102 pagesControl de Calidad en La Industria Farmacéutica PDFAbrahan HC100% (1)
- 02 Unidad 01Document29 pages02 Unidad 01Carlos Canel EgremiNo ratings yet
- 2007 Manual Metodos Inmunologicos Completo Web PDFDocument139 pages2007 Manual Metodos Inmunologicos Completo Web PDFLiliana LinoNo ratings yet
- Protocolo de Tesis Final UltimoDocument31 pagesProtocolo de Tesis Final UltimoCarlos Canel EgremiNo ratings yet
- Sopa de PanDocument2 pagesSopa de PanCarlos Canel EgremiNo ratings yet
- Atlas D - Hématologie EquateurDocument50 pagesAtlas D - Hématologie EquateurChristian Gallegos Palermo100% (3)
- Art05 Hongos PDFDocument9 pagesArt05 Hongos PDFCarlos Canel EgremiNo ratings yet
- BPLDocument34 pagesBPLctimanaNo ratings yet
- Rocamora Freud HipnosisDocument11 pagesRocamora Freud Hipnosis198809scribdNo ratings yet
- Filarias Del Hombre: I.IntroducciónDocument25 pagesFilarias Del Hombre: I.IntroducciónConan Jose Gonzalez AguilarNo ratings yet
- Charcot HisteriaDocument12 pagesCharcot HisteriaAlejandra PMNo ratings yet
- Proteina C Reactiva PDFDocument1 pageProteina C Reactiva PDFCarlos Canel EgremiNo ratings yet
- Atlas D - Hématologie EquateurDocument50 pagesAtlas D - Hématologie EquateurChristian Gallegos Palermo100% (3)
- Metabolismo EritrocitarioDocument13 pagesMetabolismo Eritrocitariomiguel angel100% (9)
- Sangre Y Hematopoyesis: Dra. Alejandra Sánchez AndradeDocument46 pagesSangre Y Hematopoyesis: Dra. Alejandra Sánchez AndradeJesús Rivera HernándezNo ratings yet
- 4 5fundatal2006 Sandra BettyDocument29 pages4 5fundatal2006 Sandra BettyCarlos Canel EgremiNo ratings yet
- ALMACENAMIENTODocument25 pagesALMACENAMIENTOCarlos Canel Egremi100% (1)
- Tapia-Gone Et Al. 2008Document4 pagesTapia-Gone Et Al. 2008Marla SaldanhaNo ratings yet
- Fisiología Leucocitaria I: Tema 27 Del Programa. Unidad Didáctica 16 Del Libro, en La Página 185Document20 pagesFisiología Leucocitaria I: Tema 27 Del Programa. Unidad Didáctica 16 Del Libro, en La Página 185Carlos Canel EgremiNo ratings yet
- Tinciones PresentacionDocument18 pagesTinciones PresentacionCarlos Canel EgremiNo ratings yet
- PRACTIKADocument4 pagesPRACTIKACarlos Canel EgremiNo ratings yet
- Raices Griegas y Latinas PDFDocument24 pagesRaices Griegas y Latinas PDFCarlos Canel Egremi67% (3)
- Ejercicios Programación Lineal y No LinealDocument7 pagesEjercicios Programación Lineal y No LinealCamilo TinocoNo ratings yet
- GUIA 3 SpvaDocument4 pagesGUIA 3 SpvaDary OrduzNo ratings yet
- Informe Psicologico GeorgeDocument5 pagesInforme Psicologico Georgemcm26No ratings yet
- PLantilla Ishikawa en Word 11Document1 pagePLantilla Ishikawa en Word 11Maxwell castillo paucarNo ratings yet
- Eqz Cognitivo MorrisonDocument105 pagesEqz Cognitivo MorrisonnatachaNo ratings yet
- Emociones NEUROCIENCIADocument29 pagesEmociones NEUROCIENCIACardenas IgnacioNo ratings yet
- Atencion Prehospitalaria en Paciente de Trauma Y EmergenciaDocument31 pagesAtencion Prehospitalaria en Paciente de Trauma Y EmergenciaXavi AzueroNo ratings yet
- Abordaje FT en Cirugía Cardiaca BYPASSDocument56 pagesAbordaje FT en Cirugía Cardiaca BYPASSTiitiimuaNo ratings yet
- Prescripcion Del Ejercicio en NiñosDocument26 pagesPrescripcion Del Ejercicio en NiñosSantiago SalamancaNo ratings yet
- Vibraciones en El TrabajoDocument44 pagesVibraciones en El TrabajoAlexandra Zapata CórdovaNo ratings yet
- Efecto de La Toxina Botul Nica Tipoa en La Funcionalidad Las Sinc - 2021 - NeurDocument6 pagesEfecto de La Toxina Botul Nica Tipoa en La Funcionalidad Las Sinc - 2021 - NeurDayan LazaroNo ratings yet
- Neurociencia de La IdentidadDocument96 pagesNeurociencia de La Identidadjuana guzmanNo ratings yet
- FOLLETODocument2 pagesFOLLETOLUZ ISMENIA GARCIA SANCHEZNo ratings yet
- SERGIO MARTINEZ ESCOBAR Adelgazar y EjercicioDocument3 pagesSERGIO MARTINEZ ESCOBAR Adelgazar y Ejerciciosergio martinezNo ratings yet
- Metodos Indirectos para Evaluar La Adherencia Al Tratamiento Una Revision SistematicaDocument29 pagesMetodos Indirectos para Evaluar La Adherencia Al Tratamiento Una Revision SistematicaEmma Sofía Peña GarcíaNo ratings yet
- Diabetes Mellitus Tipo II PDFDocument149 pagesDiabetes Mellitus Tipo II PDFWilder Bolaños Vargas50% (2)
- Instrumental QuirurgicoDocument70 pagesInstrumental Quirurgicogatrio2001No ratings yet
- Protocolo de Evaluación de DislaliasDocument14 pagesProtocolo de Evaluación de DislaliasLuis Eduardo Zárate100% (1)
- Síndrome de WilliamsDocument22 pagesSíndrome de WilliamsFrancisco Gabriel Miranda SeguraNo ratings yet
- Vivir Chevere Atencion Ambulatoria de LDocument32 pagesVivir Chevere Atencion Ambulatoria de LIv BiNo ratings yet
- El Estigma Del CáncerDocument4 pagesEl Estigma Del CáncerYuen MMNo ratings yet
- Depilacion Laser y IplDocument11 pagesDepilacion Laser y IplYenifer Nora Medina PuchuriNo ratings yet
- Sobrepeso y ObesidadDocument329 pagesSobrepeso y ObesidadIván S. Salazar100% (1)
- ELECTROLITOSDocument13 pagesELECTROLITOSAndres Ruiz100% (1)
- 01 I Informe Sis Ss Ce H 2016Document42 pages01 I Informe Sis Ss Ce H 2016Illumi ZaoldyckNo ratings yet
- Tema 1. Evolución Histórica Del Concepto de Salud y Salud PúblicaDocument25 pagesTema 1. Evolución Histórica Del Concepto de Salud y Salud PúblicaGuillermoMoralCorreaNo ratings yet
- 06 Evaluación VestibularDocument30 pages06 Evaluación VestibularMaria TeresaNo ratings yet
- La Legislación Que Regula La Profesión Farmacéutica Es Cada Vez Más InestableDocument19 pagesLa Legislación Que Regula La Profesión Farmacéutica Es Cada Vez Más InestableFernandaGuapucalNo ratings yet
- CoprofiliaDocument2 pagesCoprofiliaastrosNo ratings yet