You might also like
- Resistance Exercise in Men Receiving Androgen DeprivationDocument7 pagesResistance Exercise in Men Receiving Androgen DeprivationMatheus BarbosaNo ratings yet
- The Effects of An IndividualizedDocument7 pagesThe Effects of An IndividualizedMatheus BarbosaNo ratings yet
- The Benefits of Exercise Training For Quality of Life in HIVDocument13 pagesThe Benefits of Exercise Training For Quality of Life in HIVMatheus BarbosaNo ratings yet
- Percentis Bmi 2-20 CDC FemDocument1 pagePercentis Bmi 2-20 CDC FemMatheus BarbosaNo ratings yet
- Progressive Resistance Training in Breast Cancer - A SystematicDocument18 pagesProgressive Resistance Training in Breast Cancer - A SystematicMatheus BarbosaNo ratings yet
- Implications of Chronic Heart Failure On Peripheral Vasculature and Skeletal Muscle Before and After Exercise TrainingDocument17 pagesImplications of Chronic Heart Failure On Peripheral Vasculature and Skeletal Muscle Before and After Exercise TrainingMatheus BarbosaNo ratings yet
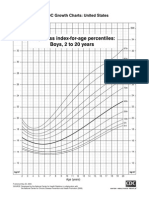
- Bmi For Age, Boys 2-20 YoDocument1 pageBmi For Age, Boys 2-20 YoLeny Karlina HakimNo ratings yet
- Resistance Exercise and SupraphysiologicDocument10 pagesResistance Exercise and SupraphysiologicMatheus BarbosaNo ratings yet
- Immune Response To A 30-Minute WalkDocument7 pagesImmune Response To A 30-Minute WalkMatheus BarbosaNo ratings yet
- Physical Activity and Hormonal Regulation of Appetite Sex Differences and Weight ControlDocument6 pagesPhysical Activity and Hormonal Regulation of Appetite Sex Differences and Weight ControlMatheus BarbosaNo ratings yet
- Hodgkin's Lymphorna in An Elite Endurance AthleteDocument5 pagesHodgkin's Lymphorna in An Elite Endurance AthleteMatheus BarbosaNo ratings yet
- Effects of Detraining On The Functional CapacityDocument8 pagesEffects of Detraining On The Functional CapacityMatheus BarbosaNo ratings yet
- Exercise Training in HIV-1-InfectedDocument8 pagesExercise Training in HIV-1-InfectedMatheus BarbosaNo ratings yet
- Lymphocyte and Cytokines After Short Periods of ExerciseDocument5 pagesLymphocyte and Cytokines After Short Periods of ExerciseMatheus BarbosaNo ratings yet
- Kinetics of Lactate Metabolism After Submaximal ErgometricDocument6 pagesKinetics of Lactate Metabolism After Submaximal ErgometricMatheus BarbosaNo ratings yet
- Evidence For Prescribing Exercise As Therapy in Chronic DiseaseDocument61 pagesEvidence For Prescribing Exercise As Therapy in Chronic DiseaseMatheus BarbosaNo ratings yet
- Exercise and The Nitric Oxide Vasodilator SystemDocument23 pagesExercise and The Nitric Oxide Vasodilator SystemMatheus BarbosaNo ratings yet
- Effects of Exercise Training On Inflammatory Markers in Patients With Heart FailureDocument11 pagesEffects of Exercise Training On Inflammatory Markers in Patients With Heart FailureMatheus BarbosaNo ratings yet
- Effects of Pharmacological Doses of NandroloneDocument10 pagesEffects of Pharmacological Doses of NandroloneMatheus BarbosaNo ratings yet
- Effects of Testosterone and Progressive Resistance Training inDocument8 pagesEffects of Testosterone and Progressive Resistance Training inMatheus BarbosaNo ratings yet
- Dynamics of Fat Cell Turnover in HumansDocument5 pagesDynamics of Fat Cell Turnover in HumansMatheus BarbosaNo ratings yet
- Effects of An Intrahospital Exercise Program Intervention For Children With LeukemiaDocument10 pagesEffects of An Intrahospital Exercise Program Intervention For Children With LeukemiaMatheus BarbosaNo ratings yet
- Effects of High-Intensity Endurance and Resistance Exercise On HIV Metabolic AbnormalitiesDocument10 pagesEffects of High-Intensity Endurance and Resistance Exercise On HIV Metabolic AbnormalitiesMatheus BarbosaNo ratings yet
- Effects of Exercise Training and Metformin On Body.13Document9 pagesEffects of Exercise Training and Metformin On Body.13Matheus BarbosaNo ratings yet
- Effects of A Supervised Home-Based Aerobic andDocument7 pagesEffects of A Supervised Home-Based Aerobic andMatheus BarbosaNo ratings yet
- Deleterious Effects of Short-Term, High-IntensityDocument6 pagesDeleterious Effects of Short-Term, High-IntensityMatheus BarbosaNo ratings yet
- Cortisol - Essential Adaptation Hormone in ExerciseDocument4 pagesCortisol - Essential Adaptation Hormone in ExerciseMatheus BarbosaNo ratings yet
- Beta-Blockers, Exercise, and The Immune System in Men With Coronary Artery DiseaseDocument6 pagesBeta-Blockers, Exercise, and The Immune System in Men With Coronary Artery DiseaseMatheus BarbosaNo ratings yet
- Effectiveness of Aerobic Exercise in AdultsDocument9 pagesEffectiveness of Aerobic Exercise in AdultsMatheus BarbosaNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Digit Symbol Substitution TestDocument9 pagesDigit Symbol Substitution TestSuresh Lukose100% (1)
- Plagiarism - Paraphrase and Summary HandoutDocument10 pagesPlagiarism - Paraphrase and Summary HandoutTika VirginiyaNo ratings yet
- SELF-EFFICACY THEORY SOURCESDocument14 pagesSELF-EFFICACY THEORY SOURCESagungNo ratings yet
- How Much Does A Kidney Transplant Cost?Document1 pageHow Much Does A Kidney Transplant Cost?Ivan RodriguezNo ratings yet
- Full Report Wastewater Paling Latest BiaDocument44 pagesFull Report Wastewater Paling Latest BiaSyazwi HakimiNo ratings yet
- Study - Social and Economic Effects of COVID-19Document151 pagesStudy - Social and Economic Effects of COVID-19мајаNo ratings yet
- Justice in Oral Health CareDocument371 pagesJustice in Oral Health CareSara Maria Menck SangiorgioNo ratings yet
- Assessment of Clients' Satisfaction With Health Service Deliveries at Jimma University Specialized HospitalDocument10 pagesAssessment of Clients' Satisfaction With Health Service Deliveries at Jimma University Specialized HospitalUrfan MajidNo ratings yet
- Gram Negative CocciDocument14 pagesGram Negative CocciRachel Marie M. GaniaNo ratings yet
- Fire Resistance of Rolling Stock Elements - 45545-3 Standard and Special Case of TunnelsDocument34 pagesFire Resistance of Rolling Stock Elements - 45545-3 Standard and Special Case of TunnelspnagarajjNo ratings yet
- Hornby Zeller Associates - Executive Resumes - Dennis E. Zeller, Helaine Hornby, Karen Hallenbeck...Document112 pagesHornby Zeller Associates - Executive Resumes - Dennis E. Zeller, Helaine Hornby, Karen Hallenbeck...Rick ThomaNo ratings yet
- Philippine HMO not liable for DSTDocument2 pagesPhilippine HMO not liable for DSTLara YuloNo ratings yet
- ESSAY 9 (When Grandiosity Speaks, Medical Students Are Supposed To Listen..)Document5 pagesESSAY 9 (When Grandiosity Speaks, Medical Students Are Supposed To Listen..)Kay BorzsonyNo ratings yet
- Imago Couples Therapy: Instructor's ManualDocument72 pagesImago Couples Therapy: Instructor's ManualSam EmekaNo ratings yet
- Nursing TheoriesDocument4 pagesNursing TheoriesSamantha Joy VidalNo ratings yet
- Ritin Kumar Srivastava Block No. 31/4, Kabeer Nagar Colony: TH THDocument3 pagesRitin Kumar Srivastava Block No. 31/4, Kabeer Nagar Colony: TH THswapnilkhadke89No ratings yet
- Visitor T30 User's Manual - Rev07 PDFDocument37 pagesVisitor T30 User's Manual - Rev07 PDFDoaà SsalamNo ratings yet
- Bacabac Vs NYK-FIL Ship Management, G.R. No. 228550, July 28, 2021Document3 pagesBacabac Vs NYK-FIL Ship Management, G.R. No. 228550, July 28, 2021Gi NoNo ratings yet
- Caffeine AP Bio InfographicDocument1 pageCaffeine AP Bio Infographic23sofia06No ratings yet
- Acrysil LTDDocument64 pagesAcrysil LTDreciprocationsNo ratings yet
- PETTERUTI, WALSH & VELÁZQUEZ The Costs of Confinement Why Good Juvenile Justice Policies Make Good Fiscal SenseDocument25 pagesPETTERUTI, WALSH & VELÁZQUEZ The Costs of Confinement Why Good Juvenile Justice Policies Make Good Fiscal SenseFrancisco EstradaNo ratings yet
- Analysis of Classification Techniques For Medical Data: April 2018Document6 pagesAnalysis of Classification Techniques For Medical Data: April 2018Ahmed MostafaNo ratings yet
- Mental Health and WellbeingDocument5 pagesMental Health and WellbeingNicole Ofili100% (1)
- Retinal Drawing A Lost Art of MedicineDocument3 pagesRetinal Drawing A Lost Art of MedicinekavyaNo ratings yet
- Bulliying 1Document34 pagesBulliying 1ahmed abybakarNo ratings yet
- Lambert - Portfolio Self-Assessment Flower Diagram 1Document1 pageLambert - Portfolio Self-Assessment Flower Diagram 1api-585093321No ratings yet
- User Manual For The SpirometerDocument4 pagesUser Manual For The SpirometerAlex WilliamsNo ratings yet
- Unofficial Developer Guide To HL7v3 BasicsDocument24 pagesUnofficial Developer Guide To HL7v3 BasicsShamilNo ratings yet
- Different Intervention Techniques For PTSDDocument2 pagesDifferent Intervention Techniques For PTSDnicolaNo ratings yet
- Long Case DysphagiaDocument3 pagesLong Case DysphagiaNadia SalwaniNo ratings yet