You might also like
- A Survey of The Butterfly Fauna of Jatun Sacha, Ecuador (Lepidoptera: Hesperioidea and Papilionoidea)Document19 pagesA Survey of The Butterfly Fauna of Jatun Sacha, Ecuador (Lepidoptera: Hesperioidea and Papilionoidea)Kellyta RodriguezNo ratings yet
- Moncada 1999 ChelConservBiolDocument7 pagesMoncada 1999 ChelConservBiolThe Science Exchange International Sea Turtle InternshipsNo ratings yet
- Additional Anuran and Saurian Records For Phuket, South ThailandDocument5 pagesAdditional Anuran and Saurian Records For Phuket, South ThailandTeravut SuwansawaipholNo ratings yet
- Baumgartner & Greenberg 1985Document23 pagesBaumgartner & Greenberg 1985DIEGO ANDRES CADENA DURANNo ratings yet
- Geluso and Harner 2013 - Nerodia Clarkii Clarkii PredationDocument26 pagesGeluso and Harner 2013 - Nerodia Clarkii Clarkii PredationArlene NavaNo ratings yet
- Origin and Population Growth of The Brown Tree Snake, On Guam!Document12 pagesOrigin and Population Growth of The Brown Tree Snake, On Guam!No NameNo ratings yet
- The White-Chinned Petrel (Procellaria Aequinoctialis) On South Georgia: Population Size, Distribution and Global SignificanceDocument7 pagesThe White-Chinned Petrel (Procellaria Aequinoctialis) On South Georgia: Population Size, Distribution and Global Significance24emmaNo ratings yet
- Vitt & Vangilder (1983) - Ecology of Snake Community in The Northeastern BrazilDocument24 pagesVitt & Vangilder (1983) - Ecology of Snake Community in The Northeastern BrazilEd MyersNo ratings yet
- Brightsmith Parrot Annual Patterns in Tambopata 2006Document18 pagesBrightsmith Parrot Annual Patterns in Tambopata 2006Jeff CremerNo ratings yet
- Ballenas Jorobadas PerúDocument8 pagesBallenas Jorobadas PerúDiana A. VizcarraNo ratings yet
- The Diet of Eurasian Griffons (Gyps Fulvus) in Crete: ArticleDocument6 pagesThe Diet of Eurasian Griffons (Gyps Fulvus) in Crete: ArticlepulpopulpoNo ratings yet
- American Society of Ichthyologists and Herpetologists (ASIH), Allen Press CopeiaDocument5 pagesAmerican Society of Ichthyologists and Herpetologists (ASIH), Allen Press CopeiaDébora Samira G. NegrãoNo ratings yet
- Blake FruitsDietsNeotropical 1992Document12 pagesBlake FruitsDietsNeotropical 1992sebastian.cardona.corredorNo ratings yet
- Costa P. Riet-Sapriza F.G. Jorge G.TDocument26 pagesCosta P. Riet-Sapriza F.G. Jorge G.TKarumbéNo ratings yet
- 13 Esponjas Porifera Parque Nacional San Esteban 2010Document14 pages13 Esponjas Porifera Parque Nacional San Esteban 2010carmenNo ratings yet
- Aquaculture of Ecteinascidia Turbinata HerdmanDocument6 pagesAquaculture of Ecteinascidia Turbinata HerdmanjlcarballoNo ratings yet
- Acuña 2013 First Record of Three Shark SpeciesDocument6 pagesAcuña 2013 First Record of Three Shark SpeciesHarry JonitzNo ratings yet
- Ecology of WinteringDocument12 pagesEcology of Winteringbiologia007No ratings yet
- Métodos de Enseñanza CaninaDocument12 pagesMétodos de Enseñanza CaninaXavier LagunasNo ratings yet
- Comparative Breeding Behavior of Ammospiza caudacuta and A. maritimaFrom EverandComparative Breeding Behavior of Ammospiza caudacuta and A. maritimaNo ratings yet
- Altitudinal Distribution of Skinks Along Cantubias Ridge of Mt. Pangasugan, Baybay, LeyteDocument20 pagesAltitudinal Distribution of Skinks Along Cantubias Ridge of Mt. Pangasugan, Baybay, LeyteLitlen DaparNo ratings yet
- 115732-Article Text-321978-1-10-20150415Document11 pages115732-Article Text-321978-1-10-20150415SimamkeleNo ratings yet
- Species Richness and Conservation StatusDocument11 pagesSpecies Richness and Conservation StatusMiguel N. AnayaNo ratings yet
- Penguins!!Document10 pagesPenguins!!Travis SiroisNo ratings yet
- CheckList Article 53857 en 1Document4 pagesCheckList Article 53857 en 1Jonathan DigmaNo ratings yet
- Check List: Bivalves and Gastropods of Tubbataha Reefs Natural Park, PhilippinesDocument12 pagesCheck List: Bivalves and Gastropods of Tubbataha Reefs Natural Park, PhilippinesMarc AlexandreNo ratings yet
- Rodriguez and Eberhard 2006Document12 pagesRodriguez and Eberhard 2006marianomxNo ratings yet
- Momotus AequatorialisDocument4 pagesMomotus AequatorialisLUIS ALBERTO HIGUERANo ratings yet
- Jellyfish Aggregations and Leatherback Turtle Foraging Patterns in A Temperate Coastal EnvironmentDocument6 pagesJellyfish Aggregations and Leatherback Turtle Foraging Patterns in A Temperate Coastal EnvironmentBengt HörbergNo ratings yet
- 3 Garcia Et Al 22ADocument25 pages3 Garcia Et Al 22AJN GPNo ratings yet
- 27 Segura EpocaDocument12 pages27 Segura EpocaVanessa Usbeck LópezNo ratings yet
- Dickinson and Fa 2000Document11 pagesDickinson and Fa 2000Ze WlyNo ratings yet
- ARC 8 1 (Special Section) 25-32 E79 Low ResDocument8 pagesARC 8 1 (Special Section) 25-32 E79 Low ResAmelia She M UNo ratings yet
- A Snake Community at Espigao Do Oeste RoDocument12 pagesA Snake Community at Espigao Do Oeste RoPaulaSuuhNo ratings yet
- Fuentes Et Al. 2011 - Rhizostoma Pulmo Life CycleDocument21 pagesFuentes Et Al. 2011 - Rhizostoma Pulmo Life CycleAmrutha AravindNo ratings yet
- Comparative Breeding Behavior of Ammospiza caudacuta and A. maritimaFrom EverandComparative Breeding Behavior of Ammospiza caudacuta and A. maritimaNo ratings yet
- Burrow Structure and Microhabitat Characteristics of Nesiergus Insulanus (Araneae: Theraphosidae) From Fre Gate Island, SeychellesDocument6 pagesBurrow Structure and Microhabitat Characteristics of Nesiergus Insulanus (Araneae: Theraphosidae) From Fre Gate Island, SeychellesAneesh MathewNo ratings yet
- Conservation Biology of The Yellow AnacoDocument24 pagesConservation Biology of The Yellow AnacoGustavo TulaNo ratings yet
- Revista Mexicana de Biodiversidad 78: 303 - 310, 2007Document8 pagesRevista Mexicana de Biodiversidad 78: 303 - 310, 2007ElyRangelNo ratings yet
- Scharer (2003) A Survey of The Epibiota of Eretmochelys Imbricata (Testudines - Cheloniidae) of Mona Island, Puerto RicoDocument5 pagesScharer (2003) A Survey of The Epibiota of Eretmochelys Imbricata (Testudines - Cheloniidae) of Mona Island, Puerto RicoMurat OzaydinliNo ratings yet
- Distribution and Habitat Use by Sheldgeese Populations Wintering in Buenos Aires Province, ArgentinaDocument8 pagesDistribution and Habitat Use by Sheldgeese Populations Wintering in Buenos Aires Province, ArgentinahumbertofisicoNo ratings yet
- Vogt, 1988Document12 pagesVogt, 1988ianNo ratings yet
- Effects of Environmental Variability On Trophic Interactions and Food Web Structure in A Pelagic Upwelling EcosystemDocument19 pagesEffects of Environmental Variability On Trophic Interactions and Food Web Structure in A Pelagic Upwelling EcosystemĐức HuyNo ratings yet
- Digestive Ecology of Two Omnivorous Canarian Lizard Species (Gallotia, Lacertidae)Document14 pagesDigestive Ecology of Two Omnivorous Canarian Lizard Species (Gallotia, Lacertidae)Kevin SHNo ratings yet
- Occurrence, Distribution and Composition of Marine Mammals in The Bight of Bony, NigeriaDocument10 pagesOccurrence, Distribution and Composition of Marine Mammals in The Bight of Bony, NigeriaRizky RamaNo ratings yet
- Biotropica - 2006 - Sol RzanoDocument10 pagesBiotropica - 2006 - Sol Rzano2181800646No ratings yet
- Home Range TucanDocument8 pagesHome Range TucanPepe ZFNo ratings yet
- AMRE2417Document9 pagesAMRE2417tortugamarinaNo ratings yet
- Progress in Oceanography: Joan E. Cartes, Vanesa Papiol, Beatriz GuijarroDocument18 pagesProgress in Oceanography: Joan E. Cartes, Vanesa Papiol, Beatriz GuijarroSigmundSingulNo ratings yet
- Aguiar. 2007. Bat Frugivory in A Remnant of Southeastern Brazilian Atlantic ForestDocument11 pagesAguiar. 2007. Bat Frugivory in A Remnant of Southeastern Brazilian Atlantic ForestJ E Fernando CernaNo ratings yet
- Changes in The Structure of A Shallow Sandy-Beach Community in Peru During An El Niño Event Arntz 1987Document15 pagesChanges in The Structure of A Shallow Sandy-Beach Community in Peru During An El Niño Event Arntz 1987Eduardo LopezNo ratings yet
- Estacionalidad Desplazamiento Condor (Martino, 2011)Document12 pagesEstacionalidad Desplazamiento Condor (Martino, 2011)Gabriel Eduardo Mendoza RiquelmeNo ratings yet
- 3 MeiofaunaDocument4 pages3 MeiofaunaGonzalo Oyanguren AparicioNo ratings yet
- New and Additional Records For The Ant Fauna (Hymenoptera, Formicidae) of MoroccoDocument10 pagesNew and Additional Records For The Ant Fauna (Hymenoptera, Formicidae) of Moroccodavid fernandezNo ratings yet
- Diversity of Scorpions (Chelicerata: Arachnida) in The Atlantic Forest in Pernambuco, Northeastern BrazilDocument5 pagesDiversity of Scorpions (Chelicerata: Arachnida) in The Atlantic Forest in Pernambuco, Northeastern BrazilGonzalo Ramírez GalánNo ratings yet
- Boa Constrictor, An Introduced Predator On Cozumel Island, MexicoDocument7 pagesBoa Constrictor, An Introduced Predator On Cozumel Island, MexicoCristopher GonzalezNo ratings yet
- Nesting Biology of The Yellow-Throated Euphonia: Large Clutch Size in A Neotropical FrugivoreDocument16 pagesNesting Biology of The Yellow-Throated Euphonia: Large Clutch Size in A Neotropical FrugivoreAby MoReNo ratings yet
- Misc 14 32Document42 pagesMisc 14 32Juan Alberto Colmán ParedesNo ratings yet
- Exploration: Driving Value: Malcolm Brown, EVP Exploration September 2013Document47 pagesExploration: Driving Value: Malcolm Brown, EVP Exploration September 2013joelayfNo ratings yet
- Ielts Fever Listening Practice Test 4 PDFDocument7 pagesIelts Fever Listening Practice Test 4 PDFAlina EdelNo ratings yet
- Dutra & Stranz 04Document29 pagesDutra & Stranz 04unisinos100% (2)
- 03 Haq & Schutter, 2008 - A Chronology of Paleozoic Sea-Level ChangesDocument6 pages03 Haq & Schutter, 2008 - A Chronology of Paleozoic Sea-Level ChangesAnanda LimaNo ratings yet
- Keep404 PDFDocument7 pagesKeep404 PDFbashraaNo ratings yet
- Operator Activity EXTERNALDocument1 pageOperator Activity EXTERNALdecandana100% (1)
- Prehistoric Compared To Present CompletedDocument61 pagesPrehistoric Compared To Present Completedapi-73954052No ratings yet
- Gargaf of LibyaDocument1 pageGargaf of Libyammarios66No ratings yet
- Making InferencesDocument6 pagesMaking InferencesAndreaMaeBaltazarNo ratings yet
- Chronostratigraphy and Geological TimeDocument17 pagesChronostratigraphy and Geological TimeNamwangala Rashid NatinduNo ratings yet
- EIA.. Technically Recoverable Shale Oil and Shale Gas Resources Outside The United States PDFDocument730 pagesEIA.. Technically Recoverable Shale Oil and Shale Gas Resources Outside The United States PDFKitty FitzgeraldNo ratings yet
- Week 3 - The Dinosaur LessonDocument3 pagesWeek 3 - The Dinosaur LessonEccentricEdwardsNo ratings yet
- Wood Shells - A New Art Form: Steve GarrisonDocument43 pagesWood Shells - A New Art Form: Steve GarrisonRileyNo ratings yet
- 10-2 Evidence For EvolutionDocument11 pages10-2 Evidence For Evolutionapi-265907602No ratings yet
- Carpathian MountainsDocument9 pagesCarpathian MountainsAlex KibalionNo ratings yet
- Mammalogy SyllabusDocument8 pagesMammalogy Syllabusshabi049No ratings yet
- Planation Surfaces in Northern EthiopiaDocument10 pagesPlanation Surfaces in Northern EthiopiaPaula Louíse Lima FelipeNo ratings yet
- Kami Export - Justin Noh - 2019.05 International Questions - PDF (NEW)Document55 pagesKami Export - Justin Noh - 2019.05 International Questions - PDF (NEW)thinman110707No ratings yet
- Brill: BRILL Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To BehaviourDocument47 pagesBrill: BRILL Is Collaborating With JSTOR To Digitize, Preserve and Extend Access To Behaviourapi-283358168No ratings yet
- Animal 3Document48 pagesAnimal 3Khairy IestNo ratings yet
- GastropodaDocument22 pagesGastropodaJanuarista Amartya DyastiNo ratings yet
- Deltas Reservoir System SLIDES MEDCO 2006Document43 pagesDeltas Reservoir System SLIDES MEDCO 2006Te Siem Bro LayukaNo ratings yet
- UcspDocument6 pagesUcspdianaNo ratings yet
- Phylum EchinodermataDocument32 pagesPhylum Echinodermataapi-255334265100% (1)
- 21-Day Stretching Challenge: Danceinsight Part OneDocument2 pages21-Day Stretching Challenge: Danceinsight Part OneWijdane AoufiNo ratings yet
- C1 Advanced Listening Worksheet 5Document9 pagesC1 Advanced Listening Worksheet 5LVNo ratings yet
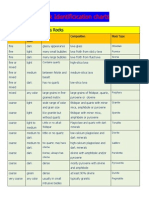
- Rock Identificication ChartsDocument3 pagesRock Identificication ChartsSan ShweNo ratings yet
- (Duellman 1990) Herpetofaunas in Neotropical Rainforests - Comparitive Composition, History, and Resource UseDocument52 pages(Duellman 1990) Herpetofaunas in Neotropical Rainforests - Comparitive Composition, History, and Resource UseDann VergaraNo ratings yet
- ReptilesDocument34 pagesReptilesZevannayasmineNo ratings yet
- New England Geological Guide 1992: January 1992Document100 pagesNew England Geological Guide 1992: January 1992JoshLeighNo ratings yet