Professional Documents
Culture Documents
Lectura 61
Uploaded by
Antonio AngelitoOriginal Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Lectura 61
Uploaded by
Antonio AngelitoCopyright:
Available Formats
36
Reflejos medulares
Los reflejos son muy adaptables y controlan el movimiento de una forma dotada de finalidad Los reflejos medulares producen patrones coordinados de contraccin muscular Los reflejos cutneos producen movimientos complejos que desempean funciones protectoras y posturales El reflejo miottico acta contrarrestando el alargamiento de un msculo Las redes neuronales de la mdula espinal contribuyen a la integracin de las respuestas reflejas dotadas de finalidad El reflejo miottico implica una va monosinptica Las interneuronas inhibidoras coordinan los msculos que rodean una articulacin La divergencia en las vas reflejas amplifica las entradas sensitivas y coordina las contracciones musculares La convergencia de las aferencias (entradas) sobre las interneuronas aumenta la flexibilidad de las respuestas reflejas Las rdenes motoras generadas centralmente pueden alterar la transmisin en las vas reflejas medulares Los mecanismos tnicos y dinmicos regulan la intensidad de los reflejos medulares Las neuronas motoras y proporcionan un mecanismo de ajuste de la sensibilidad de los husos musculares Los reflejos propioceptivos desempean un papel importante en la regulacin de los movimientos voluntarios y automticos Los reflejos que afectan a los msculos de las extremidades estn mediados por vas medulares y supramedulares Los reflejos miotticos refuerzan las rdenes centrales de movimiento La lesin del sistema nervioso central produce alteraciones caractersticas en los reflejos y en el tono muscular La interrupcin de las vas descendentes a la mdula espinal a menudo produce espasricidad
La seccin de la mdula espinal en los humanos provoca un perodo de shock medular seguido de hiperreflexia Resumen
URANTE LOS MOVIMIENTOS NORMALES, el sistema nervioso central emplea informacin de un vasto conjunto de receptores sensitivos con el fin de asegurar la generacin de un patrn correcto de actividad muscular. La informacin sensitiva de los msculos, las articulaciones y la piel, por ejemplo, es esencial para regular el movimiento. Sin estas aferencias somatosensi-tivas, los grandes movimientos tienden a ser imprecisos, mientras que las tareas que requieren una coordinacin fina de la manos, como abrocharse los botones, son imposibles (vase Captulo 33). Charles Sherrington figura entre los pioneros en reconocer la importancia de la informacin sensitiva para regular los movimientos. En una monografa de gran influencia publicada en 1906, propona que los reflejos simples, movimientos estereotipados desencadenados por la activacin de los receptores situados en la piel o los msculos, son las unidades bsicas del movimiento. Propuso adems que se pueden producir secuencias complejas de movimientos combinando reflejos simples. Este punto de vista ha sido el principio rector de la fisiologa motora durante buena parte de este siglo. Slo hace relativamente poco se ha modificado, al reconocerse el hecho de que se pueden producir muchos movimientos coordinados en ausencia de informacin sensitiva. Por ejemplo, en diversas especies se pueden iniciar y mantener patrones locomotores en ausencia de aferencias sensitivas estructuradas (Captulo 37). Sin embargo est fuera de toda duda la nocin de que los reflejos desempean un papel importante en la estructuracin de la actividad motora. El punto de vista actual es que los reflejos se mtegran con rdenes motoras generadas centralmente para producir movimientos adaptativos.
Figura 36-1. Las respuestas reflejas con frecuencia son complejas y pueden variar segn la tarea. A. La perturbacin de un brazo causa una respuesta refleja excitadora en el msculo extensor del codo contralateral cuando se emplea la extremidad contralateral para evitar que el cuerpo se vaya hacia delante, pero el mismo estmulo provoca una respuesta inhibidora en el msculo (disminucin del EMG) cuando
la mano contraria sostiene una taza llena. (Adaptado de Marsden y cois., 1981.) B. La carga del pulgar durante una secuencia rtmica de movimientos de tocar el dedo y el pulgar provoca una respuesta refleja (zona sombreada en azul) en el msculo que mueve el dedo, as como en el msculo del pulgar cargado. El movimiento adicional del dedo asegura que el movimiento de pinza sigue siendo preciso. (Adaptado de Col y cois., 1984.)
En este captulo consideraremos los principios que subyacen a la organizacin y funcin de los reflejos, centrndonos en los reflejos medulares. Los estmulos sensitivos de los reflejos medulares proceden de receptores situados en los msculos, las articulaciones y la piel, y los circuitos nerviosos responsables de la respuesta motora estn localizados, en su totalidad, en la mdula espinal. Tradicionalmente, los reflejos se han considerado movimientos automticos, estereotipados, en respuesta a la estimulacin de los receptores perifricos. Este punto de vista proceda fundamentalmente de los primeros estudios sobre preparados reducidos con animales en los que se estudiaban los reflejos bajo un conjunto de condiciones normalizadas. Sin embargo, cuando los investigadores extendieron sus estudios para medir los reflejos en el transcurso de la conducta normal, nuestro concepto de los reflejos vari sustancialmente. Sabemos en la actualidad que en condiciones normales los reflejos se pueden modificar para adaptarlos a la tarea. Esta flexibilidad permite la incorporacin lenta de los reflejos a movimientos complejos iniciados por rdenes centrales.
Los reflejos son muy adaptables y controlan el movimiento de una forma dotada de finalidad Un buen ejemplo de la adaptabilidad de los reflejos se ve cuando se estiran los msculos de la mueca de un brazo mientras un sujeto est de rodillas o en pie. Los msculos estirados se contraen, pero los msculos de otras extremidades se contraen tambin para evitar una prdida de equilibrio. Es interesante que la respuesta refleja del extensor del codo del brazo contrario depende de la tarea que est realizando ese brazo. Si el brazo se est empleando para estabilizar el cuerpo (agarrado al borde de una mesa) se provoca una potente respuesta excitadora en los msculos extensores del codo para oponerse a la cada del cuerpo hacia delante. Si el brazo est sujetando un objeto inestable, como una taza de t, una inhibicin refleja de los extensores del codo impide los movimientos de la taza (Fig. 36-1A). Otro ejemplo de la adaptabilidad de los reflejos es el reflejo de los msculos flexores de los dedos y del pulgar
Captulo 36 / Reflejos medulares 5
71
en respuesta al estiramiento de los msculos del pulgar. Si nos oponemos a la flexin del pulgar mientras el sujeto est intentando tocar rtmicamente la punta del pulgar con la punta del dedo, se produce una respuesta de la-tencia corta tanto en los msculos flexores de los dedos como en los del pulgar. El reflejo en el msculo flexor de los dedos produce un movimiento mayor de flexin del dedo para compensar la disminucin de la flexin del pulgar, asegurando as la realizacin de la tarea pretendida (Fig. 36-1B). Si el sujeto est realizando simplemente movimientos rtmicos del pulgar, se produce una respuesta refleja slo en el msculo flexor del pulgar. Un tercer ejemplo de la naturaleza adaptable de los reflejos implica a un reflejo condicionado de flexin-retirada. La retirada en flexin puede asociarse con un tono auditivo por tcnicas clsicas de condicionamiento (Captulo 62). Se pide a los sujetos que coloquen el dedo ndice, con la superficie palmar hacia abajo, sobre un electrodo. Despus se administra una corriente elctrica suave unida al tono. Como cabe esperar, despus de slo unos cuantos de estos pares de estmulos el tono auditivo desencadenar el reflejo de retirada. Qu es exactamente lo que se ha condicionado? Se trata de la contraccin de un grupo fijo de msculos o un acto de conducta que retira el dedo del estmulo nocivo? Esa cuestin se puede resolver pidiendo a los sujetos que den la vuelta a sus manos despus de que el condicionamiento sea completo, de forma que sea ahora la superficie dorsal del dedo la que est en contacto con el electrodo. La mayora de los sujetos retirar los dedos del electrodo cuando suene el tono, incluso aunque esto signifique la contraccin de los msculos opuestos. Por lo tanto, la respuesta refleja condicionada al tono no es slo la contraccin de un conjunto estereotipado de msculos, sino una conducta apropiada. Estos ejemplos ilustran tres principios importantes. Primero, la transmisin en las vas reflejas se establece de acuerdo con la tarea motora. El estado de las vas reflejas para cualquier tarea se denomina conjunto funcional. Se desconoce en gran parte la forma exacta en que se establece el conjunto funcional para la mayora de las tareas motoras (el descubrimiento de los mecanismos subyacentes constituye uno de los mayores retos y de los aspectos ms excitantes de la investigacin actual sobre los sistemas motores). Segundo, la aferencia sensitiva de una fuente localizada generalmente produce respuestas reflejas en muchos msculos, algunos de los cuales pueden estar distantes del estmulo. Estas mltiples respuestas son coordinadas para lograr el objetivo pretendido. Tercero, los centros suprame-dulares desempean un papel importante para regular y adaptar los reflejos medulares, incluso hasta el punto de invertir movimientos cuando es apropiado. Con el fin de comprender la base nerviosa de los reflejos y cmo se modifican para una tarea determinada, hemos de tener primero un conocimiento meticuloso de cmo estn organizadas las vas reflejas en la mdula
espinal. La mdula espinal es un lugar importante de integracin de los reflejos con rdenes centrales, y muchas caractersticas cualitativas de los reflejos se mantienen despus de la seccin medular completa.
Los reflejos medulares producen patrones coordinados de contraccin muscular
Los reflejos cutneos producen movimientos complejos que desempean funciones protectoras y posturales Un ejemplo familiar de reflejo medular es el reflejo de flexin-retirada, en el que aleja rpidamente una extremidad de un estmulo doloroso, habitualmente por la contraccin simultnea de todos los msculos flexores de la extremidad. Sabemos que es un reflejo medular porque persiste despus de la seccin medular completa, una situacin que aisla los circuitos medulares del encfalo. La flexin-retirada es un reflejo de proteccin en el que un estmulo aislado provoca que los msculos se contraigan de forma coordinada en muchas articulaciones. A travs de vas polisinpticas divergentes la seal sensitiva excita las neuronas motoras que inervan los msculos flexores de la extremidad estimulada a la vez que inhibe las neuronas motoras que inervan los extensores de la extremidad (Fig. 36-2A). La excitacin de un grupo de msculos y la inhibicin de sus antagonistas es lo que Sherrington denomin por primera vez inervacin recproca. (Los msculos antagonistas son los que actan en direccin opuesta a un determinado msculo; por ejemplo, los extensores de la rodilla son los antagonistas de los flexores de la rodilla.) La inervacin recproca es un principio esencial de la organizacin motora y se tratar ms adelante en este captulo. Junto con la flexin de la extremidad estimulada, el reflejo puede producir un efecto contrario en la extremidad contralateral, es decir, una excitacin de las neuronas motoras extensoras y una inhibicin de las flexoras. Este reflejo de extensin cruzada sirve para facilitar el apoyo postural durante el alejamiento de un pie de un estmulo doloroso. La contraccin de los msculos extensores de la pierna contraria contrarresta el aumento de la carga causado por la elevacin de la extremidad estimulada. As, la flexin-retirada es un acto motor completo, aunque simple. Aunque los reflejos de flexin son relativamente estereotipados en su forma, tanto la extensin espacial como la fuerza de la contraccin muscular dependen de la intensidad del estmulo (Captulo 33). Tocar una estufa que est poco caliente puede producir una retirada moderadamente rpida slo de la mueca y el codo, mientras que si est muy caliente conduce invariablemente a una contraccin enrgica de todas las articulaciones, provocando el rpido alejamiento de toda la extremidad. La duracin del reflejo suele aumentar con la intensidad del estmulo, y las contracciones producidas en un reflejo de
Captulo 36 / Reflejos medulares 717
flexin siempre duran ms que el estmulo. Por tanto, los reflejos no son simplemente repeticiones de un patrn estereotipado de movimiento; son modulados por propiedades del estmulo. Dada la similitud del reflejo de flexin-retirada con el dar pasos, en el pasado se pensaba que la marcha podra ser generada simplemente como una serie de reflejos de flexin. En la actualidad sabemos que un componente importante del sistema de control nervioso de la marcha es un conjunto de circuitos medulares intrnsecos que no requieren estmulos sensitivos para producir el patrn bsico de la marcha (Vase Captulo 37). Sin embargo, en los mamferos los circuitos medulares intrnsecos que controlan la marcha comparten muchas interneuronas que participan en los reflejos de flexin. El reflejo miottico acta contrarrestando el alargamiento de un msculo Quiz el reflejo medular ms importante, y con seguridad el ms estudiado, sea el reflejo miottico, una contraccin muscular que se produce cuando un msculo se alarga. Inicialmente se pensaba que los reflejos miotti-cos eran ma propiedad intrnseca de los msculos. Sin embargo, Sherrington demostr en el cambio de siglo que se podan abolir seccionando tanto las races dorsales como las ventrales, estableciendo, por tanto, que requeran una aferencia sensitiva desde el msculo a la mdula espinal y una va de regreso a los msculos. Ahora sabemos que el receptor que detecta la variacin de longitud es el huso muscular (vase Recuadro 36-1) y que los axones aferentes establecen conexiones excitadoras directas con las neuronas motoras (Fig. 36-2B). En su investigacin sobre los reflejos, Sherrington desarroll un valioso modelo experimental para investigar los circuitos medulares. Hizo sus experimentos con gatos cuyos troncos enceflicos haban sido seccionados a nivel mesenceflico, entre los tubrculos cuadrigminos superiores e inferiores. Esto se denomina preparacin descerebra-da. El efecto de este procedimiento es desconectar el resto del encfalo de la mdula espinal, bloqueando as las sensaciones de dolor, adems de interrumpir la regulacin normal de los reflejos por centros cerebrales superiores. Cuando Sherrington trataba de flexionar pasivamente la pata trasera en extensin rgida de un gato descerebrado, not un aumento de la contraccin de los msculos que estaban siendo estirados (Fig. 36-B2). A esto lo denomin reflejo miottico. Sherrington descubri tambin que el estiramiento de un msculo provocaba que los msculos antagonistas se relajaran. Concluy que el estmulo de estiramiento causaba la excitacin de ciertas neuronas motoras y la inhibicin de otras (inervacin recproca). Los animales descerebrados tienen reflejos medulares estereotipados y habitualmente exaltados, lo que facilita el examen de los factores que controlan su expresin, tambin muestran un aumento considerable del tono
muscular extensor, a veces suficiente como para mantener en pie al animal. El tono muscular est aumentado porque, en ausencia de control por centros cerebrales superiores, las vas descendentes del tronco enceflico facilitan poderosamente los circuitos neuronales que participan en los reflejos miotticos de los msculos extensores. En animales normales los reflejos medulares son ms dbiles y considerablemente ms variados en cuanto a su intensidad que en los descerebrados, porque existe un equilibrio entre la facilitacin y la inhibicin. Las vas descendentes de la corteza cerebral y otros centros superiores del cerebro regulan de forma continua la fuerza de los reflejos miotticos. Las redes neuronales de la mdula espinal contribuyen a la integracin de las respuestas reflejas dotadas de finalidad El reflejo miottico implica una va monosinaptica El circuito nervioso responsable del reflejo miottico fue una de las primeras vas reflejas en ser estudiada con detalle (vase Captulo 4). La fase fisiolgica de este reflejo se aclar midiendo la latencia de la respuesta provocada en las races ventrales cuando se estimulaban elctricamente las races dorsales. Cuando se activan selectivamente las grandes fibras aferentes la de las terminaciones primarias del huso muscular, la latencia del reflejo a travs de la mdula espinal es inferior a 1 ms. (La clasificacin de las fibras sensitivas del msculo se muestra en el Recuadro 36-2.) Como el retraso que produce una sola sinapsis es de 0.5 a 0.9 ms, se puede inferir que las fibras la establecen conexiones directas con las neuronas motoras a, creando una va monosinptica (Fig. 36-4). Las fibras la de un msculo no slo excitan las neuronas motoras que inervan ese msculo (conexiones homnimas) sino tambin las que inervan msculos con una accin mecnica similar (conexiones heternimas). El patrn de conexiones entre las fibras la y las neuronas motoras se puede demostrar directamente mediante tcnicas de registro intracelular. El registro intracelular ha mostrado tambin que las neuronas motoras a que inervan los msculos antagonistas reciben inhibicin de las fibras la a travs de una clase especfica de interneuronas inhibidoras, las interneuronas inhibidoras la. Esta va inhibidora bisinptica es la base de la inervacin recproca; cuando un msculo se estira, los antagonistas se relajan. Las interneuronas inhibidoras coordinan los msculos que rodean una articulacin La inervacin recproca no slo es til en los reflejos miotticos, sino tambin en los movimientos voluntarios. La relajacin del msculo antagonista durante los movimientos facilita la velocidad y la eficiencia porque los
718
Parte VI / Movimiento
Recuadro 36-1.
Husos musculares
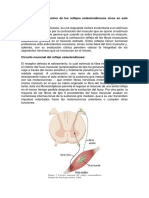
rodo en cuyo transcurso est cambiando la longitud, y una fa esttica o de estado estacionario, cuando el msculo se ha esta bilizado en una nueva longitud. Las especializaciones estructi rales en el seno de cada componente de los husos muscularp permiten que las aferencias del huso enven por separado sea les de cada fase. Existen dos tipos de fibras musculares intrafusales: las fibras nucleares de bolsa y las fibras nucleares en cadena. Un huso muscular tpico posee 2 6 3 fibras de bolsa y un nmero variable de fibras en cadena, aproximadamente 5. Adems, existen dos tipos de terminaciones nerviosas sensitivas: una terminacin primaria nica y un nmero variable de terminaciones secundarias (hasta 8). La terminacin primaria (fibra la) forma una espiral en torno a la regin central de todas las fibras musculares intrafusales (Fig. 36-3B). Las terminaciones secundarias (fibras del grupo II) estn localizadas en la vecindad de las regiones centrales de las fibras estticas de bolsa y de cadena. Las neuronas motoras y se pueden dividir tambin en dos clases, dinmicas y estticas. Las motoneuronas y dinmicas inervan las fibras de bolsa dinmicas, mientras que las neuronas motoras y inervan las fibras de bolsa y de cadena estticas. Esta dualidad estructural se refleja en una dualidad funcional. La descarga de estado estacionario, o tnica, de las terminaciones sensitivas primarias y secundarias enva seales sobre la longitud del msculo en estado estacionario. Las terminaciones primarias son adems muy sensibles a la velocidad de estiramiento, lo que les permite suministrar informacin respecto a la velocidad de los movimientos. Como son muy sensibles a pequeas variaciones, las terminaciones primarias proporcionan una informacin rpida sobre variaciones inesperadas de la longitud, que resultan tiles para generar reacciones rpidas de correccin. El aumento de la actividad de las neuronas motoras y dinmicas aumenta la sensibilidad dinmica de las terminaciones primarias pero carece de influencia en las terminaciones secundarias. El aumento de actividad de las motoneuronas y estticas aumenta el nivel tnico de actividad tanto de las terminaciones primarias como secundarias, disminuye la sensibilidad dinmica de las terminaciones primarias, y puede evitar la supresin de la actividad cuando el msculo se libera del estiramiento (Fig. 36-3C). Por tanto, el sistema nervioso central puede ajusfar la sensibilidad dinmica y esttica de las fibras sensitivas de los husos musculares de forma independiente.
Los husos musculares son pequeos receptores sensitivos en-capsulados, en forma de huso, localizados en la parte carnosa del msculo. Su principal fruicin es enviar seales sobre las variaciones en la longitud del msculo en cuyo interior se alojan. Las variaciones en la longitud de los msculos estn estrechamente asociadas con los cambios en los ngulos de las articulaciones que atraviesan. Por ello, los husos musculares pueden ser empleados por el sistema nervioso central para detectar las posiciones relativas de los diferentes segmentos corporales. Cada huso posee tres componentes principales: 1) un grupo de fibras musculares especializadas intrafusales cuyas regiones centrales no son contrctiles; 2) terminaciones sensitivas miel-nicas de gran dimetro, que se originan a partir de las regiones centrales de las fibras intrafusales; y 3) terminaciones motoras mielnicas de pequeo dimetro que inervan las regiones polares contrctiles de las fibras intrafusales (Fig. 36-3A). Cuando las fibras intrafusales se estiran, lo que a menudo se denomina cargar el huso, tambin se distienden las terminaciones sensitivas, que aumentan su ritmo de activacin. Como los husos musculares estn dispuestos en paralelo con el vientre principal del msculo, las fibras musculares varan de longitud cuando lo hace el conjunto del msculo. As, al estirarse el msculo, aumenta la actividad de las terminaciones sensitivas de los husos musculares. Cuando se acorta el msculo, el huso se descarga y la actividad disminuye. La inervacin motora de las fibras musculares intrafusales procede de las neuronas motoras de pequeo dimetro, denominadas neuronas motoras y, para diferenciarlas de las moto-neuronas a, de gran dimetro, que inervan las fibras musculares extrafusales. La contraccin de las fibras musculares intrafusales no contribuye a la fuerza de la contraccin muscular. La activacin de las motoneuronas y ms bien causa un acortamiento de las regiones polares de las fibras intrafusales. Esto a su vez distiende la regin central no contrctil, tirando de sus dos extremos, lo que induce un aumento del ritmo de activacin de las terminaciones sensitivas o una mayor probabilidad de que la distensin del msculo haga que la terminacin sensitiva descargue. As, las neuronas motoras y proporcionan un mecanismo para ajusfar la sensibilidad de los husos musculares. La estructura y el comportamiento funcional de los husos musculares es considerablemente ms compleja de lo que implica esta descripcin simple. Cuando un msculo se estira, existen dos fases de la variacin de longitud: una fase dinmica, el pe-
720
Parte VI / Movimiento
Recuadro 36-2.
Activacin selectiva de las fibras
sensitivas del msculo
las respuestas centrales provocadas por diferentes clases de fibr sensitivas se pueden evaluar graduando la fuerza del estrruil' elctrico; y ciertas clases de receptores se pueden activar de form aislada (lo que resulta imposible en condiciones naturales). La fuerza de los estmulos elctricos necesaria para activa una fibra sensitiva se mide en relacin con la fuerza necesaria para activar las fibras aferentes mayores, porque las fibras aferentes de mayor tamao tienen el umbral de activacin elctrica ms bajo. Estas fibras del grupo I se suelen activar en un nivel de una o dos veces el umbral de las aferentes de mayor tamao (teniendo las fibras la en promedio un umbral algo ms bajo que las fibras Ib). La mayora de las fibras del grupo II se activan en un nivel de 2-5 veces el umbral, mientras que las fibras pequeas del grupo III y IV requieren potencias de estimulacin del orden de 10 a 50 veces el umbral de activacin.
Las fibras sensitivas se clasifican por su dimetro. Los axones con dimetros grandes conducen ms rpidamente los potenciales de accin. Como cada clase de receptor origina fibras aferentes con dimetros de una gama restringida, este mtodo de clasificacin diferencia hasta cierto punto las fibras que proceden de diferentes grupos de receptores sensitivos. Los principales grupos de fibras sensitivas del msculo figuran en el Cuadro 36-1 (vase en el Captulo 24, la clasificacin de las fibras sensitivas de la piel y las articulaciones). La organizacin de las vas reflejas en la mdula espinal se ha determinado fundamentalmente estimulando elctricamente las fibras sensitivas y registrando las respuestas provocadas en las distintas clases de neuronas de la mdula espinal. Este mtodo de activacin posee tres ventajas sobre la estimulacin natural. Permite establecer con precisin la cronologa de la entrada aferente;
msculos que realizan el movimiento primario no trabajan contra la contraccin de los msculos que se oponen a ellos. Las interneuronas inhibidoras la que participan en el reflejo miottico se emplean tambin para coordinar la contraccin muscular durante los movimientos voluntarios. Las interneuronas reciben aferencias de fibras colaterales de los axones que descienden procedentes de las neuronas de la corteza motora, estableciendo conexiones excitadoras directas con las neuronas motoras medulares (Fig. 36-5A). Esta organizacin caracterstica simplifica el control de los movimientos voluntarios, porque los centros superiores no tienen que enviar rdenes independientes a los msculos antagonistas. La inervacin recproca de los msculos de acciones opuestas no slo es la nica forma til de coordinacin. A veces resulta ventajoso contraer al mismo tiempo el msculo que realiza el movimiento primario y el antagonista. Esta contraccin simultnea posee el efecto de fijar la articulacin,
y es ms til cuando resultan cruciales la precisin y la estabilizacin articular. Un ejemplo de este fenmeno es la contraccin simultnea de los msculos flexores y extensores del codo inmediatamente antes de coger una pelota (vase el Captulo 33). Las interneuronas inhibidoras la reciben tanto seales excitadoras como inhibidoras de todas las vas descendentes importantes (Fig. 36-5A). Al variar el equilibrio de las entradas excitadoras e inhibidoras sobre estas interneuronas, los centros supramedulares pueden reducir la inhibicin recproca y permitir la contraccin simultnea, controlando de este modo la relativa fijacin articular necesaria para cumplir los requisitos del acto motor. La actividad de las neuronas motoras medulares tambin est regulada por otra clase importante de interneuronas inhibidoras, las clulas de Renshaw (Fig. 36-5B). Las clulas de Renshaw son excitadas por colaterales de los axones de las neuronas motoras y establecen conexiones sinpticas inhibidoras con varias poblaciones de neuro-
Captulo 36 / Reflejos medulares
721
Captulo 36 / Reflejos medulares
723
724
Parte VI / Movimiento
Captulo 36 / Reflejos medulares 725
las neuronas motoras y es mantener la tensin en el huso muscular durante la contraccin activa, asegurando as la respuesta a diferentes longitudes. Cuando las neuronas motoras a son estimuladas selectivamente en condiciones experimentales, la activacin de la fibra sensitiva Hel huso muestra una pausa caracterstica durante la contraccin., porque el msculo se est acortando y por lo tanto est descargando (aflojando) el huso. Sin embargo, si se activan al mismo tiempo las neuronas motoras y, la pausa se rellena porque la contraccin de las fibras in-trafusales mantiene cargada, o en tensin, la regin central (Fig. 36-9). Por lo tanto, una funcin esencial de la inervacin de las fibras intrafusales por las neuronas motoras y es evitar que la fibra sensitiva del huso se inactive cuando el msculo se acorta como resultado de la contraccin activa, permitindole as transmitir seales sobre los cambios de longitud a lo largo de todo el espectro de longitudes musculares. Este mecanismo mantiene el ritmo de activacin del huso en un nivel ptimo para transmitir cambios de longitud, sea cual sea la longitud real del msculo. En muchos movimientos voluntarios las neuronas motoras a se activan normalmente ms o menos en paralelo con las neuronas y, un patrn denominado coactivacin a-y. Esto conlleva el mantenimiento automtico de la sensibilidad. Adems de los axones de las neuronas motoras y, las neuronas motoras a tambin inervan las fibras intrafusales. stas se denominan eferentes esqueletofusimotoras, o i. Se ha detectado una cantidad importante, aunque todava no cuantificada, de la inervacin esqueletofusi-motora en los husos musculares de los gatos y los seres humanos. Estas eferencias proporcionan el equivalente a la coactivacin oc-y; cuando se activan las neuronas esqueletofusimotoras, se compensa, por lo menos en parte, la descarga del uso por la contraccin de las fibras extrafusales, al cargarse a causa de la contraccin intra-fusal. Sin embargo, la existencia de un sistema esqueletofusi-motor, con su vinculacin forzada entre la contraccin extrafusal y la intrafusal, sirve para resaltar la importancia del sistema fusimotor independiente constituido por las neuronas motoras y. Aparentemente, en los mamferos se ha desarrollado un mecanismo que permite desacoplar el control de los husos musculares del de los msculos en los que estn situados. En principio, este desacoplamiento debera permitir una mayor flexibilidad para controlar la eferencia del huso en diferentes tipos de tareas motoras. En apoyo de esta conclusin se encuentran los registros en aferentes fusales primarias durante diversos movimientos naturales en los gatos. La cantidad y el tipo de 'envidad de las neuronas motoras y (esttica o dinmica), se preprograma a un nivel bastante estable, pero va-ria de acuerdo con la tarea o el contexto especfico. En general, tanto las neuronas y estticas como dinmicas
Figura 36-8. La fuerza de un reflejo medular puede ser modulada por variaciones en la transmisin en la va refleja. A. Una va refleja se puede modificar en tres sitios: 1) en las neuronas motoras ex, 2) en las interneuronas en vas polisinpticas, y 3) en los terminales de los axones aferentes. La liberacin de transmisor de las fibras aferentes primarias est regulada por la inhibicin presinptica (vase Captulo 13). B. Un aumento de la aferencia excitadora tnica mantiene la des polarizacin en la neurona (sombreado) y permite que una afe rencia por lo dems ineficaz inicie potenciales de accin en la neurona (Vu = voltaje umbral; Vm = potencial de membrana).
estn preprogramadas a niveles ms elevados a medida que aumenta la velocidad y dificultad del movimiento. Las situaciones impredecibles, como cuando un gato es agarrado o manipulado, inducen un aumento llamativo de la actividad y dinmica, que se refleja en un aumento de la respuesta del huso cuando los msculos se estiran. Cuando un animal est realizando una tarea difcil, como caminar sobre una barra fina, existen niveles elevados de actividad y tanto esttica como dinmica (Fig. 36-10). As, el sistema nervioso emplea el sistema fusimotor ajusfando el nivel de activacin y el equilibrio entre la activacin de las neuronas motoras y estticas y dinmicas para conseguir una sintonizacin fina de los husos, de forma que el conjunto de la aferencia de los husos musculares proporcione la informacin ms adaptada a la tarea. En los seres humanos todava no se han determinado las condiciones de las tareas en las cuales tiene lugar una activacin independiente de las neuronas motoras a y y.
Captulo 36 / Reflejos medulares
727
728
Parte VI / Movimiento
Captulo 36 / Reflejos medulares
729
730
Parte VI / Movimiento
estiramiento no ejercen una influencia suficiente en las neuronas motoras como para vencer grandes cargas inesperadas. Esto resulta inmediatamente evidente si consideramos lo que ocurre cuando tratamos de levantar una maleta pesada que creamos que estaba vaca. Tenemos que parar un momento y realizar un nuevo movimiento con una activacin muscular mucho mayor. Por lo tanto, las vas del reflejo miottico proporcionan un mecanismo de compensacin de pequeas variaciones en la carga e irregularidades intrnsecas de la contraccin muscular. Esta accin est mediada tanto por las vas monosinpticas como por las vas de arco largo, y la contribucin relativa de cada va depende del msculo y de la tarea. La lesin del sistema nervioso central produce alteraciones caractersticas en los reflejos y en el tono muscular Se pueden provocar reflejos miotticos en muchos msculos de todo el cuerpo, y se emplean de forma sistemtica en la exploracin clnica de pacientes con trastornos neurolgicos. La forma tpica de obtenerlos es golpeando bruscamente el tendn de un msculo con un martillo de reflejos. Por ello reciben el nombre de reflejos osteotendinosos, aunque el receptor que se estimula, el huso muscular, est situado en el vientre muscular y no en el tendn. Slo participan en el reflejo osteotendinoso las fibras sensitivas primarias, porque se estimulan de forma selectiva por el estiramiento rpido producido al golpear el tendn. Un anlogo elctrico al reflejo osteotendinoso es el reflejo de Hoffmann (Recuadro 36-4). La medicin de las alteraciones de la fuerza del reflejo miottico puede ayudar a establecer el diagnstico de ciertos trastornos y a localizar la lesin o la enfermedad en el sistema nervioso central. La ausencia o debilidad de algunos reflejos miotticos a menudo indica un trastorno de uno o ms componentes del arco reflejo: los axones sensitivos o motores, los cuerpos celulares de las neuronas, o el propio msculo. Sin embargo, como la excitabilidad de las neuronas motoras depende de las influencias descendentes excitadoras e inhibidoras, las lesiones del sistema nervioso central pueden tener como consecuencia reflejos hiperactivos o hipoactivos. La interrupcin de las vas descendentes a la mdula espinal a menudo produce espasticidad El tono muscular, la fuerza con la que un msculo se resiste a ser alargado, depende de la elasticidad intrnseca, o rigidez, de los msculos. Como el msculo posee elementos elsticos en serie y en paralelo que se oponen al alargamiento, se comporta como un resorte. Sin embargo, adems de la rigidez intrnseca, existe una contribucin nerviosa al tono muscular; el bucle de retroaccin del reflejo miottico tambin se opone al alargamiento
del nvsculo. Los circuitos nerviosos responsables de 10 reflejos miotticos proporcionan a los centros superiore del sistema nervioso un mecanismo para ajustar el tono muscular en diferentes circunstancias. Los trastornos del tono muscular a menudo se asocian a lesiones del sistema motor, especialmente las que interfieren en las vas descendentes, porque la fuerza de los reflejos miotticos est controlada por los centros cerebrales superiores. Pueden implicar tanto aumentos anormales del tono (hipertona) como disminuciones {hipotmv'a) La forma ms frecuente de hipertona es la espasticidad que se caracteriza por reflejos osteotendinosos hiperactivos y un aumento de la resistencia al estiramiento muscular rpido. El estiramiento lento de un msculo puede encontrar poca resistencia en un paciente con espasticidad. A medida que se incrementa la velocidad de estiramiento, la resistencia aumenta progresivamente. La espasticidad es, por tanto, un fenmeno fsico. Slo se produce una contraccin refleja activa durante un estiramiento rpido; cuando el msculo se mantiene en una posicin alargada, la contraccin refleja cede. Sin embargo, en algunos pacientes la hipertona tiene tambin un componente tnico, es decir, la contraccin refleja contina incluso despus de que haya cesado el estiramiento del msculo. Todava no est clara la fisiopatologa de la espasticidad. Se crea desde hace mucho tiempo que el aumento de la ganancia de los reflejos miotticos en la espasticidad obedeca a una hiperactividad de las neuronas motoras y. Sin embargo, experimentos recientes han puesto en duda esta explicacin. Aunque puede haber hiperactividad y en algunos casos, probablemente un factor ms importante son las alteraciones en la actividad de fondo de las neuronas motoras a y las interneuronas. Sea cual sea el mecanismo preciso que produce la espasticidad, el efecto es una potente facilitacin de la transmisin de la va refleja monosinptica desde las fibras sensitivas la a las neuronas motoras oc. De hecho, sta es la base del tratamiento. Un procedimiento frecuente hoy da es imitar la inhibicin presinptica en las terminales de las fibras la. Esto se realiza mediante la administracin intra-tecal del frmaco baclofeno a la mdula espinal. Este frmaco es un agonista de los receptores del cido y-aminobu-trico (GABA)B; la unin del GABA a estos receptores disminuye la entrada de calcio en las terminales presinpti-cas y por tanto reduce la cantidad de transmisor liberado. La seccin completa de la mdula espinal en los seres humanos provoca un perodo de shock medular seguido de hiperreflexia La lesin de la mdula espinal puede provocar grandes variaciones en la intensidad de los reflejos medulares. Cada ao, unas 10 000 personas sufren lesiones medulares en las que se produce una seccin medular completa-Ms de la mitad de estas lesiones producen incapacidad permanente, que incluye trastornos de las funciones mo-
Captulo 36 / Reflejos medulares
731
You might also like
- Tests ProyectivosDocument187 pagesTests ProyectivosJorge Quito75% (4)
- 10.guía AnsiedadDocument43 pages10.guía AnsiedadPedro Camacho100% (1)
- Reflejos OsteotendinososDocument30 pagesReflejos OsteotendinososFabi.129100% (2)
- Perfil Del Wais IVDocument12 pagesPerfil Del Wais IVJoneyris ParraNo ratings yet
- Reflejos .Fisiologia y Exploracion.Document35 pagesReflejos .Fisiologia y Exploracion.Erika Fernanda YazoNo ratings yet
- Hoja de RegistroDocument6 pagesHoja de RegistroFormación Continua Poliestudios0% (1)
- 2578-Texto Del Artículo-25896-1-10-20210217Document13 pages2578-Texto Del Artículo-25896-1-10-20210217Antonio AngelitoNo ratings yet
- Redessocialesparaempresas PDFDocument341 pagesRedessocialesparaempresas PDFsimonepcNo ratings yet
- El Futuro de La Administracion Parte IDocument36 pagesEl Futuro de La Administracion Parte IangleogNo ratings yet
- Cuestionario Multidimensional de CelosDocument2 pagesCuestionario Multidimensional de Celossapito677100% (1)
- Antcedentes Generales, Semana 3Document3 pagesAntcedentes Generales, Semana 3Antonio AngelitoNo ratings yet
- Recetario ChocolateDocument28 pagesRecetario ChocolatenagbostNo ratings yet
- La Gestión de La Innovación en La Empresa: Evolución de Su Campo de EstudioDocument12 pagesLa Gestión de La Innovación en La Empresa: Evolución de Su Campo de Estudiodanilo enrique torres pimientoNo ratings yet
- La Gestión de La Innovación en La Empresa: Evolución de Su Campo de EstudioDocument12 pagesLa Gestión de La Innovación en La Empresa: Evolución de Su Campo de Estudiodanilo enrique torres pimientoNo ratings yet
- Material de Estudio, Semana 1 PDFDocument38 pagesMaterial de Estudio, Semana 1 PDFAntonio AngelitoNo ratings yet
- Ideas Fuerza, Semana 5Document1 pageIdeas Fuerza, Semana 5Antonio AngelitoNo ratings yet
- Cocina SaludableDocument148 pagesCocina SaludableIsra HernandezNo ratings yet
- Marketing DigitalDocument20 pagesMarketing DigitalAntonio AngelitoNo ratings yet
- Administración de RRHH y Productividad en Pymes Tesis 2009Document0 pagesAdministración de RRHH y Productividad en Pymes Tesis 2009Antonio AngelitoNo ratings yet
- 12.7 Trabajo Psicologia Del Aprendizaje MMDocument9 pages12.7 Trabajo Psicologia Del Aprendizaje MMAntonio Angelito100% (1)
- Receta EmpolvadosDocument5 pagesReceta EmpolvadosAntonio AngelitoNo ratings yet
- Libro de RecetasDocument68 pagesLibro de Recetas3elizaNo ratings yet
- Condicionamiento Clásico y Condicionamiento InstrumentalDocument31 pagesCondicionamiento Clásico y Condicionamiento InstrumentalElard SánchezNo ratings yet
- Trabajo Psicobiologia S1Document6 pagesTrabajo Psicobiologia S1Antonio AngelitoNo ratings yet
- Trabajo Neurociencias S6Document11 pagesTrabajo Neurociencias S6Antonio AngelitoNo ratings yet
- Esquema de InvestigacionDocument1 pageEsquema de InvestigacionAntonio AngelitoNo ratings yet
- Características de los principales neurotransmisores: funciones y observacionesDocument3 pagesCaracterísticas de los principales neurotransmisores: funciones y observacionesAntonio Angelito67% (3)
- Herramientas de Comunicación en El Aprendizaje de AdultosDocument1 pageHerramientas de Comunicación en El Aprendizaje de AdultosAntonio AngelitoNo ratings yet
- Autoconcepto y comunicaciónDocument62 pagesAutoconcepto y comunicaciónrolotico100% (1)
- Medula Espinal y ReflejosDocument28 pagesMedula Espinal y ReflejosMayrim OchoaNo ratings yet
- Explique El Motivo de Los Reflejos Osteotendinosos Vivos en Este PacienteDocument6 pagesExplique El Motivo de Los Reflejos Osteotendinosos Vivos en Este PacienteGretel Romero SalinasNo ratings yet
- MP Neurorehabilitación 2022Document48 pagesMP Neurorehabilitación 2022Sobera BegoNo ratings yet
- Sistema Nervioso IiDocument92 pagesSistema Nervioso IiMayra RetamozoNo ratings yet
- Reflejos y diagnósticoDocument3 pagesReflejos y diagnósticoAndreaQuezadaCarrionNo ratings yet
- Reflejos Osteotendinosos - Hernandez ZamornaoDocument12 pagesReflejos Osteotendinosos - Hernandez ZamornaoANGEL DANIEL GONZALEZ MELGOZANo ratings yet
- Reflejos OsteotendinososDocument10 pagesReflejos OsteotendinososRivero Negrete NatalyNo ratings yet
- Exposición TRMDocument60 pagesExposición TRMLina MarcelaNo ratings yet
- Reflejos osteotendinosos y cutáneosDocument4 pagesReflejos osteotendinosos y cutáneosYhan D. KoronelNo ratings yet
- Reflejo Miotatico 2020 PDFDocument5 pagesReflejo Miotatico 2020 PDFliz0% (1)
- Reflejos OsteotendinososDocument7 pagesReflejos OsteotendinososJesús Choque ZuritaNo ratings yet
- Ortega. Informe NeurologiaDocument10 pagesOrtega. Informe NeurologiaJoshua Camilo Ortega DurangoNo ratings yet
- Exploración de reflejos osteotendinososDocument25 pagesExploración de reflejos osteotendinososJENNIFERNo ratings yet
- Reflejos - Andrea ParadaDocument6 pagesReflejos - Andrea ParadaAndrea ParadaNo ratings yet
- Dermatomas - Miotomas 1Document79 pagesDermatomas - Miotomas 1nidiaNo ratings yet
- Práctica 3Document14 pagesPráctica 3adriana toainga vallinNo ratings yet
- Guía de Laboratorio. Reflejos FinalDocument6 pagesGuía de Laboratorio. Reflejos FinalLaura Fonnegra MendezNo ratings yet
- 13 Neurologia y Neurocirugia by MedikandoDocument165 pages13 Neurologia y Neurocirugia by MedikandoMateo CortesNo ratings yet
- REFLEJOS - ResumenDocument6 pagesREFLEJOS - ResumenShonnys Pantoja FranNo ratings yet
- Grupo #4 - Motilidad y SensibilidadDocument19 pagesGrupo #4 - Motilidad y SensibilidadDiana MoranNo ratings yet
- Neuroanatomia AtmDocument49 pagesNeuroanatomia AtmPe T. ErNo ratings yet
- Capítulo 43Document9 pagesCapítulo 43naxittaNo ratings yet
- Informe Fisiología Sensorial IIDocument27 pagesInforme Fisiología Sensorial IILILIAN YAMILE LINARES OROZCONo ratings yet
- Semiologia de Los ReflejosDocument19 pagesSemiologia de Los Reflejoslesther cifuentesNo ratings yet
- 02funcion NeuromuscularDocument28 pages02funcion NeuromuscularUlises AlonzoNo ratings yet
- Reflejo Miotactico (De Distension)Document10 pagesReflejo Miotactico (De Distension)Drables MejiaNo ratings yet
- Panteologias de Los ReflejosDocument2 pagesPanteologias de Los ReflejosYuu BalsaNo ratings yet
- Reflejo MiotaticoDocument2 pagesReflejo MiotaticoNaye PerdomoNo ratings yet