Professional Documents
Culture Documents
Control Respiratorio
Uploaded by
Thalia LROriginal Description:
Original Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
Control Respiratorio
Uploaded by
Thalia LRCopyright:
Available Formats
CONTROL DE LA RESPIRACIN
Editores
J.M. Pino F. Garca Ro
Coordinador Dr. Francisco Garca Ro
Edita Sanitaria 2000
Imprime: Grficas ROAL S.A. I.S.B.N.: 84-611-3469-9 Depsito Legal:
Reservados todos los derechos. No est permitida la reproduccin total o parcial de este libro, ni su tratamiento informtico, ni la transmisin de ninguna forma o por cualquier medio, ya sea electrnico, mecnico, por fotocopia, por registro u otros medios, sin el permiso previo por escrito del titular del Copyrigth.
PRESENTACIN
Es posible que pocas facetas de la funcin respiratoria sean menos conocidas por la generalidad de los neumlogos que el sistema regulador de la respiracin. Frente a la necesaria y amplia formacin en mecnica ventilatoria, intercambio de gases o adaptacin al sueo y al ejercicio, el funcionamiento del sistema que vigila a los anteriores y trata de adecuarlos a las necesidades de cada momento permanece, hasta cierto punto, en el descuido. Como en cualquier rgano, tanto las funciones respiratorias como las no respiratorias de los pulmones precisan de un sistema de control que las regule. Pero esto resulta incluso ms acusado en el caso de la respiracin que en el de otras funciones corporales, puesto que depende de un mecanismo consciente y de un sistema de control automtico. En este sentido, son pocas las actividades corporales bsicas que han alcanzado tal grado de dependencia de las estructuras cerebrales superiores. Baste pensar en el trnsito intestinal, en la filtracin glomerular, en la secrecin endocrina o paracrina o en el latido cardiaco. Ninguna de estas actividades puede ser controlada de forma voluntaria ni, por supuesto, interrumpida de forma consciente. Es posible que este aspecto peculiar de la respiracin todava no haya sido reconocido en su justa importancia, pero, en cualquier caso, manifiesta una estrecha conexin entre las estructuras superiores del sistema nervioso central y el aparato respiratorio. Aun sin depender de un control voluntario, la respiracin no se limita a ser una sucesin regular de fases inspiratorias y espiratorias a lo largo del da. Muchas de las actividades cotidianas interfieren con la respiracin y precisan de estructuras reguladoras que faciliten su realizacin simultnea. Para hablar, cantar, gritar, rer, llorar, bostezar, toser, estornudar, beber o comer es necesario modificar, en mayor o menor grado, el patrn respiratorio. Desde una vertiente completamente automtica, el sistema de control de la respiracin garantiza una amplia capacidad homeosttica para esta funcin. De tal modo, se tratan de minimizar los cambios que se podran originar en los gases sanguneos durante el ejercicio, el sueo o ante determinadas enfermedades. Como es obvio, este mecanismo tiene un lmite que, cuando se sobrepasa, muchas veces se manifiesta como alguna de las formas de disnea. Resulta evidente la importancia funcional y clnica de todas las estructuras que participan en el sistema regulador de la respiracin. Desde un punto de vista meramente taxonmico, tampoco debera olvidarse que la unin a los rganos de la caja torcica de algunos rganos extratorcicos y de diversas estructuras neurales incluidas en el tronco enceflico y del mesencfalo, le confieren categora de sistema al aparato respiratorio. En cualquier caso, es preciso reconocer que el estudio del control de la respiracin plantea algunas dificultades. Sin duda, una de ellas proviene de ser un punto de contacto entre la neumologa y otras reas de la biomedicina, en particular las neurociencias. Quiz esto justifique una
mayor disociacin entre el conocimiento bsico y su aplicacin clnica que en otras facetas de la funcin respiratoria. Al igual que en muchos otros campos, en las ltimas dcadas se han producido destacados avances en la comprensin de los mecanismos moleculares que regulan la sensibilidad ante estmulos respiratorios de naturaleza qumica o fsica, en la caracterizacin de las neuronas y conexiones celulares implicadas en la generacin del ritmo respiratorio y en los mecanismos responsables del desarrollo de respuestas ventilatorias integradas. Frente a ello, disponemos de procedimientos de estudio clnico muy clsicos, con notables limitaciones en algunos casos y, a veces, con dudosa utilidad para la toma de decisiones en la prctica mdica. Por estas consideraciones, nos ha parecido que la presente coleccin de monografas Estudio de la Funcin Respiratoria debera incluir una dedicada exclusivamente al control de la respiracin. Con la misma, pretendemos proporcionar al lector una visin integrada del sistema que regula la respiracin, en la que desde el conocimiento de sus aspectos morfolgicos y celulares-moleculares se profundice en las pruebas de estudio habituales, para tratar de intuir y desarrollar su aplicacin clnica. Con esta pretensin, el libro se ha estructurado en nueve captulos. Mientras que en el primero se realiza una descripcin somera de todas las estructuras implicadas en la regulacin de la respiracin, en el segundo se profundiza en las funciones del cuerpo carotdeo y en sus mecanismos celulares y moleculares. Los dos siguientes captulos revisan los procedimientos disponibles en la actualidad para evaluar los aspectos ms destacados del control de la respiracin, esto es, el patrn respiratorio, el impulso inspiratorio central y el control qumico y mecnico de la respiracin. Para completar la descripcin de esta funcin reguladora, los captulos quinto y sexto desarrollan las adaptaciones que se producen en el mismo durante dos situaciones muy habituales en la vida cotidiana, como son el ejercicio y el sueo. Para concluir, los tres ltimos captulos tratan de proporcionar una visin del control de la respiracin desde un punto de partida clnico. Como ya se ha mencionado, la disnea, en sus diversas variantes, podra ser interpretada como una seal de alarma ante el mal funcionamiento de cualquiera de los elementos integrados en la regulacin respiratoria. Por ello, la evaluacin de sus mecanismos patognicos cobra importancia en el diagnstico y seguimiento de pacientes con enfermedades respiratorias e, incluso, con otros trastornos. Aunque el sistema regulador de la respiracin experimenta alteraciones en la mayora de las enfermedades respiratorias, los sndromes de hipo e hiperventilacin representan la expresin ms paradigmtica de su mal funcionamiento. Su valoracin, y su tratamiento, se pueden enriquecer con una aproximacin funcional a estos pacientes. Y justo es finalizar estas lneas expresando mi ms sincero reconocimiento y agradecimiento a todos los autores. Estoy convencido de que su entusiasmo, intensa dedicacin y esfuerzo desarrollado son los mejores avales para el xito de esta obra. Esperamos haber alcanzado los objetivos propuestos y nuestro mayor deseo es que resulte til para el lector.
Francisco Garca Ro
NDICE
Captulo 1.
Estructuras reguladoras de la respiracin
Francisco Garca Rio, Vanesa Lores Gutirrez, Blas Rojo Moreno-Arrones . . . . . 7
Captulo 2.
Mecanismos de quimiorrecepcin en el cuerpo carotdeo
Ana Obeso, Constancio Gonzlez, Asuncin Rocher . . . . . . . . . . . . . . . . . . . . . 29
Captulo 3.
Evaluacin del patrn respiratorio y del impulso inspiratorio central
Neus Salord, Pere Casn . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
Captulo 4.
Evaluacin del control qumico y mecnico de la respiracin
Mara Jess Rodrguez Nieto, Germn Peces-Barba Romero . . . . . . . . . . . . . . 81
Captulo 5.
Regulacin de la ventilacin durante el ejercicio
Luis Puente Maestu, Felipe Villar lvarez . . . . . . . . . . . . . . . . . . . . . . . . . . . . 99
Captulo 6.
Control de la respiracin durante el sueo
Alberto Alonso Fernndez, Mnica de La Pea Bravo, Francisco Garca Ro . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 125
Captulo 7.
Mecanismos patognicos de la disnea
Jos Mara Marn Trigo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 139
Captulo 8.
Exploracin funcional de los sndromes de hipoventilacin
Jos Miguel Rodrguez Gonzlez-Moro, Soledad Lpez Martn, Pilar de Lucas Ramos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 153
Captulo 9.
Sndromes de hiperventilacin
Mara Teresa Ramrez Prieto . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 175
CAPTULO 1
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
FRANCISCO GARCA RO, VANESA LORES GUTIRREZ, BLAS ROJO MORENOARRONES
Servicio de Neumologa. Hospital Universitario La Paz. Francisco Garca Ro Alfredo Marquere 11, izqda, 1 A 28034-Madrid Tfno: 639 911718 Fax: 91 7277096 e-mail: fgr01m@gmail.com
PALABRAS CLAVE: Centros respiratorios, quimiosensibilidad, mecanorreceptores, hipoxia, hipercapnia, patrn respiratorio.
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
8 NDICE DE CONTENIDOS: 1. Centros respiratorios A. Localizacin morfolgica 1. Bulbo raqudeo 2. Puente 3. Estructuras suprapontinas 4. Mdula espinal B. Generacin del ritmo respiratorio C. Patrn respiratorio 2. Receptores A. Quimiorreceptores B. Mecanorreceptores 1. Receptores de las vas areas superiores 2. Receptores traqueobronquiales 3. Receptores musculares 3. Efectores 1. Msculos esquelticos de la orofaringe 2. Msculos lisos traqueobronquiales 3. Msculos respiratorios 4. Bibliografa
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
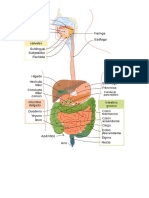
9 La necesidad de oxgeno (O2) y la eliminacin de dixido de carbono (CO2) varan mucho entre el reposo y el ejercicio extenuante. No obstante, en personas sanas, la presin arterial de oxgeno (PaO2) apenas se modifica entre ambas circunstancias. Ello es debido a la existencia de un estricto mecanismo de control, que regula la ventilacin de acuerdo a las necesidades metablicas y al gasto cardaco (1). Se ha comprobado que el incremento de cidos no voltiles en la circulacin sangunea, la vida a elevadas altitudes o ciertas enfermedades pulmonares producen una hiperventilacin refleja (2). Sin embargo, trastornos como la fiebre y enfermedades metablicas o psiquitricas se acompaan de alteraciones en la profundidad y frecuencia respiratoria que no pueden ser atribuidas exclusivamente a cambios en la composicin de la sangre arterial (1). Adems, actividades tan rutinarias como hablar, cantar, oler, aspirar, toser, tragar, tener hipo, estornudar, vomitar o aguantar la respiracin, modifican, asimismo, el patrn respiratorio (1-4). Todas estas mltiples circunstancias que afectan a la respiracin son mediadas por un intrincado mecanismo de control, constitudo por receptores centrales y perifricos y un complejo entramado de vas nerviosas y centros integradores, localizados en el cerebro, tronco cerebral y mdula espinal (3). El sistema que regula la respiracin puede ser considerado, de modo simplificado, como una organizacin formada por tres componentes (Figura 1): un "controlador", o sistema nervioso central (SNC), que integra seales propias y de las unidades sensoriales; "efectores" (pulmones, vas areas y msculos respiratorios), que ejecutan las rdenes; y "sensores", para contrastar la eficacia de la respiracin alcanzada con respecto a la propuesta (3).
Figura 1. Representacin esquemtica del sistema de control respiratorio. Tomado de Berger et al. (3).
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
10 1. CENTROS RESPIRATORIOS A. Localizacin morfolgica Las principales estructuras del sistema nervioso central relacionadas con el control de la respiracin son el bulbo raqudeo, el puente, la formacin reticular, los centros superiores y la mdula espinal. De modo muy esquemtico, se puede afirmar que ciertas neuronas localizadas en el tronco cerebral regulan la respiracin automtica, mientras que la corteza cerebral es la responsable de la respiracin voluntaria (5). Un grupo de neuronas integradoras, localizadas en la mdula espinal, procesan la informacin eferente de ambos centros respiratorios con la informacin aferente de los receptores perifricos, y envan seales a los msculos respiratorios (3). 1. Bulbo raqudeo Es el centro regulador de la respiracin espontnea en todos los mamferos estudiados hasta la fecha, incluido el hombre (6). Las neuronas respiratorias se distribuyen en dos centros: el grupo respiratorio dorsal (GRD) y el grupo respiratorio ventral (GRV) (Figura 2) (5,6).
VISIN TRANSVERSAL VISIN CORONAL
Figura 2. Representacin esquemtica de los principales grupos de neuronas respiratorias en el tronco enceflico y en la mdula espinal. La seccin transversa corresponde a la altura del obex. Abreviaturas: NA=ncleo ambiguo, nVII=ncleo facial, nXII=ncleo hipogloso, nTS=ncleo del tracto solitario, DRG= grupo respiratorio dorsal, PRG=grupo respiratorio del puente, RVLM=mdula rostral ventrolateral, pFRG=grupo respiratorio parafacial, VRG=grupo respiratorio ventral, C4=segmento correspondiente de la mdula espinal.
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
11 Grupo respiratorio dorsal Se trata de una concentracin bilateral de neuronas, predominantemente de tipo inspiratorio, localizada en una regin del bulbo raqudeo dorsal (7,8), que corresponde al ncleo del tracto solitario (nTS) (5,9,10). El 80% de estas neuronas proyectan en sentido contralateral en la mdula espinal, tanto a nivel cervical como torcico (6). Inicialmente, las neuronas inspiratorias fueron diferenciadas en dos grupos, segn que su descarga estuviera estimulada (I) o inhibida (I) por la insuflacin pulmonar (11). Una subpoblacin de neuronas I, llamadas neuronas de descarga tarda, ha sido relacionada con el final de la inspiracin, puesto que su despolarizacin se produce en ese momento (12). Un tercer tipo de neuronas inspiratorias del GRD es el integrado por las neuronas P o de despolarizacin "en rampa", que parecen intervenir en los reflejos de la va area, facilitando la fase espiratoria (8). Slo alrededor de un 5% de las neuronas del GRD tienen un patrn de descarga espiratorio (6). En conjunto, las seales aferentes, procedentes de los quimiorreceptores y receptores pulmonares, llegan al GRD a travs de los pares craneales IX y X (3). Este enva eferencias hacia motoneuronas inspiratorias, sobre todo, frnicas, y al propio GRV (3). Grupo respiratorio ventral Corresponde a una columna longitudinal bilateral localizada en la regin ventrolateral del bulbo raqudeo, que se extiende desde el nivel de la primera raz cervical hasta alcanzar la lnea media del puente (13). Esta situacin corresponde a los ncleos ambiguos, retroambiguo, paraambiguo y retrofacial, con el complejo de Btzinger (1,5,10). Posee tanto neuronas inspiratorias como espiratorias, aunque en la regin ms caudal predominan las espiratorias (6). Las neuronas inspiratorias del GRV fueron divididas en dos grupos. El primero est formado por neuronas I (que se corresponden con las neuronas I del GRD) (14). El otro grupo est constitudo por las neuronas propiobulbares, cuyo subtipo I o neuronas de descarga precoz, se relacion con el inicio de la inspiracin (14). Las neuronas espiratorias localizadas en la porcin rostral del GRV muestran patrones de despolarizacin precoz y tarda, mientras que las ms caudales nicamente presentan una descarga espiratoria tarda (15). El GRV enva diversas ramas eferentes. Del ncleo ambiguo parten las vas que se dirigen a motoneuronas inspiratorias y espiratorias de los msculos intercostales y abdominales (16). Los axones neuronales del ncleo retroambiguo se dirigen a las motoneuronas vagales de varios msculos accesorios, entre ellos el esternocleidomastoideo (1). 2. Puente La mayora de las neuronas respiratorias se localizan en la regin dorsolateral rostral del puente, configurando el grupo respiratorio pontino (PRG). Corresponde a dos ncleos ana-
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
12 tmicos: el ncleo parabraquial medial, formado por neuronas bifsicas, y el ncleo de Klliker-Fuse, integrado por neuronas inspiratorias similares a las del bulbo raqudeo (5,6,17). Se ha relacionado con la duracin de las distintas fases del ciclo respiratorio y con la frecuencia del mismo (5). Todos los centros del puente presentan ricas conexiones bilaterales con el GRV y GRD (16). En la regin ventrolateral del puente, existe un grupo de neuronas noradrenrgicas disperas denominadas A5, que proyectan a los ncleos parabraquial y de Klliker-Fuse ipsilaterales, a ambos ncleos del tracto solitario y a estructuras del tronco del encfalo relacionadas con el control cardiovascular, por lo que se supone que participan en el control cardiorrespiratorio (18). Adems, se sugiere que los grupos de neuronas pontinas modulen la respuesta a mecanorreceptores y al cuerpo carotideo (18). 3. Estructuras suprapontinas La formacin reticular se extiende desde la mdula espinal cervical superior hasta el diencfalo, en la porcin medial del tronco cerebral, y contiene neuronas del sistema activador reticular (SAR) (1,3). Dado que su actividad es inhibida por la anestesia superficial, se ha propuesto que tendra un efecto estimulante sobre el generador del ritmo respiratorio (6), y que controlara la transicin sueo-vigilia (19). En el diencfalo y el mesencfalo se han localizado neuronas con actividad respiratoria, que parecen coordinar la ventilacin con respuestas locomotoras y autnomas (6). Ya se ha mencionado que actividades como hablar, gritar, tragar o rer causan cambios marcados en la ventilacin, independientes del control automtico. As, por ejemplo, durante la fonacin disminuye la sensibilidad al CO2 y se toleran PaCO2 mayores (1). Este tipo de control voluntario y relacionado con la conducta radica en la corteza cerebral, aunque no se sabe exactamente dnde. Se ha podido comprobar que la estimulacin de algunas zonas de la corteza inhibe los movimientos respiratorios, mientras que la de otras incrementa su frecuencia (20). 4. Mdula espinal Por la sustancia blanca medular descienden los axones de los centros superiores hasta las motoneuronas respiratorias, separados los responsables del control voluntario de los encargados del control automtico (20) y los inspiratorios de los espiratorios (3,10). A cada nivel segmentario, se produce la integracin de las seales de los tractos descendentes con informacin refleja local, difiriendo posiblemente en cada grupo muscular respiratorio (1). B. Generacin del ritmo respiratorio Todos estos conocimientos, obtenidos mediante tcnicas electrofisiolgicas, biofsicas, anatmicas, farmacolgicas e histoqumicas, no han aclarado cmo se genera el ritmo respiratorio. De hecho, todava no se conoce el mecanismo que determina la alternancia inspiracin-espi-
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
13 racin en los mamferos. La mayora de las hiptesis se basan en la extrapolacin de los resultados obtenidos en animales con un sistema nervioso central ms simple (21). A partir de estudios histolgicos, Ramn y Cajal concluy, en 1909, que el ciclo respiratorio era generado en el ncleo del tracto solitario (10). Aunque esta hiptesis permaneci olvidada durante muchos aos, en las ltimas dcadas se ha confirmado la importancia del bulbo raqudeo en la determinacin del ritmo respiratorio. Von Euler (22) formul un primer modelo para explicar el funcionamiento del generador del patrn respiratorio central. Asuma que el impulso inspiratorio o actividad inspiratoria central tena una morfologa en rampa, en analoga con el registro de la despolarizacin frnica (22). La secuencia que propona consista en que la descarga de neuronas bulboespinales inspiratorias del GRD y GRV, regulada por interneuronas, originaba una despolarizacin sinptica en forma de rampa. Esta llevaba a la descarga frnica (23). Por tanto, las neuronas bulboespinales inspiratorias deberan estar agrupadas en tres clases funcionales: neuronas del grupo A, encargadas de generar la actividad inspiratoria central; del grupo C, o responsables de continuar la descarga inspiratoria; y del grupo B, a las que correspondera finalizar la actividad inspiratoria central, a partir de impulsos procedentes de la propia descarga inspiratoria y de receptores de estiramiento pulmonar (22). La enorme importancia de este modelo terico radic en que sustitua la idea de una clula "marcapaso" por la de una red neuronal inspiratoria (21). Un segundo modelo, quiz ms ajustado a la realidad, es el propuesto por Richter (24). Aportaba una interesante novedad al dividir el ciclo respiratorio en tres fases: (a) inspiracin; (b) intervalo post-inspiratorio o fase espiratoria I, en el que tanto las neuronas inspiratorias como espiratorias estn inhibidas; y (c) fase espiratoria II (24). Richter (24) consideraba que, en la generacin del ritmo respiratorio, intervienen dos clases de neuronas bulboespinales: inspiratorias (I) y espiratorias (E); y cuatro tipos de interneuronas o neuronas propiobulbares: inspiratorias precoces ("early inspiratory" o eI), inspiratorias "en rampa" ("ramp inspiratory" o Ir), inspiratorias tardas ("late inspiratory" o LI), y post-inspiratorias (pI) (Figura 3) (24). Los elementos claves en la generacin del ritmo respiratorio seran las neuronas eI y pI (21), interconectadas a travs de una compleja red sinptica. La fase inspiratoria comenzara con la descarga del sistema activador reticular (SAR) que, despus de recibir aferencias perifricas y centrales, enva un impulso estimulante a las interneuronas eI y, sobre todo, a las Ir. Estas ltimas clulas estimularan a las neuronas bulboespinales inspiratorias (I), que produciran la despolarizacin de las motoneuronas inspiratorias situadas en la mdula espinal. La fase inspiratoria terminara con la activacin de las interneuronas L-I, que no descargarn hasta que se agote la inhibicin procedente de las interneuronas eI (5,24,25). La fase post-inspiratoria o espiratoria I correspondera a la descarga de las interneuronas pI, que produce la inhibicin de las interneuronas eI e Ir, as como de las neuronas bulboespinales inspiratorias y espiratorias (21). La fase espiratoria II se iniciara al disminuir la actividad de las neuronas pI, con lo que las restantes clulas se irn liberando de su inhibicin. Esta ter-
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
14
Figura 3. Esquema de la red generadora del ritmo respiratoria porpuesto por Ritcher (24). Las flechas contnuas representan estimulacin y las discontnuas inhibicin.
cera fase del ciclo respiratorio finalizara con el restablecimiento de la actividad elctrica de las interneuronas eI, lo que permitira iniciar un nuevo ciclo (21,24). Posteriores hallazgos han confirmado que las redes neuronales que generan el ritmo respiratorio se localizan en el complejo pre-Btzinger, que est situado en la regin ventrolateral del bulbo raqudeo (26). Estas estructuras son capaces de generar no slo un tipo de actividad inspiratoria, sino hasta tres actividades diferentes, que corresponden al patrn eupneico, al suspiro y al jadeo (26). Al igual que sucede con otras redes generadoras de un patrn central, el ritmo respiratorio surge de la combinacin de las propiedades intrnsecas de la membrana neuronal, que le confieren caractersticas de marcapasos, y de una amplia red de sinapsis inhibidoras (27). Las neuronas con descarga en marcapasos se excitan por va glutamatrgica y son muy sensibles a la inhibicin sinptica. Se diferencian dos tipos de neuronas con propiedades de marcapasos. En muchas de ellas, el patrn de descargas repetidas depende de la entrada continua de sodio por canales inicos especficos, fenmeno que no puede ser bloqueado por el cadmio, por lo que se denominan neuronas marcapasos no sensibles al cadmio (27). Existe otro grupo de neuronas marcapasos, que son activadas por la entrada de calcio a travs de canales de alto voltaje activados por el calcio, y resultan sensibles al cadmio (27). Se especula que el nmero de neuronas marcapasos activas y su contribucin a la generacin del ritmo respiratorio es variable. Parece probable que durante la normoxia, ambos tipos de
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
15 neuronas marcapasos contribuyan a la generacin del ritmo respiratorio, mientras que durante la hipoxia, las neuronas sensibles al cadmio pierden gran parte de su actividad (28). Las supuestas conexiones de la red neuronal responsable de la respiracin eupneica y del jadeo se muestran en la Figura 4. En la misma, la neuronas marcapasos sensibles al cadmio (amarillo) y las neuronas marcapasos insensibles al cadmio (rojo) estn englobadas por una red de neuronas respiratorias excitatorias (E) e inhibitorias (I). Durante la hipoxia o durante el jadeo, las neuronas marcapasos sensibles al cadmio y muchas neuronas inhibitorias y excitatorias cesan su actividad, de tal modo que la actividad de descarga neuronal vendr regulada por las neuronas marcapasos no sensibles al cadmio (Figura 4) (27).
Figura 4. Modelo hipottico de la red neuronal respiratoria durante la respiracin eupneica en normoxia (A) y durante la respiracin jadeante en hipoxia (B). Abreviaturas: NM-Cads=neuronas marcapasos sensibles al cadmio, NM-Cadns=neuronas marcapasos no sensibles al cadmio, I=neuronas inspiratorias, E=neuronas espiratorias, PI=neuronas post-inspiratorias. Tomado de Ramirez y Viemari (27).
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
16 Las neuronas marcapasos dependen de la activacin endgena de receptores 5-HT2A. Esta dependencia de la modulacin serotoninrgica puede tener importantes implicaciones clnicas, puesto que su dficit en algunas regiones bulbares se relaciona con la muerte sbita del lactante (27). Englobadas en la red neuronal generadora del patrn respiratorio, las neuronas marcapasos reciben impulsos inhibitorios de neuronas excitatorias e inhibitorias. Como consecuencia de ello, se establecen las diferentes fases del ritmo respiratorio (Figura 4). Adems, la inhibicin sinptica regula la excitabilidad de conexiones glutamatrgicas y las propiedades intrnsecas de las neuronas marcapasos (29). Por tanto, la generacin del ritmo respiratorio debera ser considerada como un proceso dinmico, que depende de las propiedades intrnsecas de membrana de dos tipos de neuronas marcapasos y de una compleja red neuronal inhibitoria. En caso de no recibir aferencias procedentes del puente, la red neuronal bulbar es capaz de generar por si sla la actividad inspiratoria eupneica. C. Patrn respiratorio De forma convencional, la interpretacin del patrn respiratorio se centraba en el volumen corriente (VT) y en la frecuencia respiratoria (f). Sin embargo, resulta evidente que estos parmetros no aportan suficiente informacin. As, por ejemplo, un incremento del volumen corriente puede ser debido a un mayor tiempo inspiratorio (tI) o a un flujo inspiratorio superior. De igual modo, una mayor frecuencia respiratoria puede estar originada por un descenso de la duracin de la inspiracin, del tiempo espiratorio (tE) o de ambos (tTOT). Por ello, como se comentar en un captulo posterior, se han desarrollado expresiones ms avanzadas que permiten interpretar el patrn respiratorio en funcin de la intensidad del impulso inspiratorio central y de su duracin (30,31). La regulacin del tiempo y del volumen de la respiracin est sujeto a la interaccin entre el ritmo respiratorio generado y a las aferencias de los receptores de estiramiento (30-33). De hecho, el incremento de la ventilacin en respuesta a cualquier estmulo se produce en dos fases (33). En un primer momento, se produce a expensas del volumen corriente, hasta que aproximadamente alcanza la mitad de la capacidad vital. Despus, la ventilacin contina elevndose a costa de la frecuencia respiratoria, con muy poca o nula participacin del volumen corriente. Mientras que en la primera fase apenas se registran cambios en el tiempo inspiratorio, en la segunda se produce una finalizacin precoz de la inspiracin, atribuida al reflejo de insuflacin de Hering-Breuer, por lo que se acorta el tiempo total del ciclo respiratorio (33). Consideradas globalmente, todas las variables que integran el patrn respiratorio permiten conocer el trabajo mecnico de la respiracin, la organizacin y operacin de los centros respiratorios y el intercambio gaseoso, puesto que la ventilacin del espacio muerto depende de la relacin f VT (32,33).
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
17 2. RECEPTORES Un mecanismo que necesita autoregularse precisa sensores para verificar s se ha logrado el objetivo propuesto. Para este fin, el sistema regulador de la respiracin dispone de los quimiorreceptores perifricos y centrales y de los mecanorreceptores. A. Quimiorreceptores Desde hace varias dcadas, se ha reconocido al cuerpo carotideo como una estructura quimiosensible a nivel perifrico. En el siguiente captulo de este libro, se realiza una completa descripcin de sus caractersticas y de sus posibles mecanismos de accin. Sin embargo, el cuerpo carotdeo slo es responsable del 20-30% de la respuesta ventilatoria a la hipercapnia. El resto depende de otras estructuras localizadas en el sistema nervioso central, denominadas quimiorreceptores centrales (34). A partir de esta observacin se ha comenzado la bsqueda de reas quimiosensibles centrales, y todava existe mucha controversia sobre ellas. Se ha identificado una zona que corresponde a la superficie ventrolateral del bulbo raqudeo, medial a la salida de los pares craneales VII y X (3,35-38). El principal grupo de neuronas quimiosensibles de esta zona se ubica en el ncleo paragigantocelular lateral (39), en estrecha conexin con el grupo respiratorio dorsal (37). Al igual que las neuronas del cuerpo carotdeo, las clulas de este ncleo contienen gran variedad de agentes neuroactivos estimulantes (acetilcolina, neurotensina y sustancia P) e inhibidores (encefalinas, somatostatina y GABA) (1). Tambin se han identificado otras zonas quimiosensibles en el ncleo del tracto solitario y en el locus coeruleus (40). La estimulacin de los quimiorreceptores centrales es realizada por el incremento de hidrogeniones (H+) o de CO2 en el lquido extracelular cerebral o en el lquido cefalorraqudeo (LCR) (3). Las oscilaciones de estas sustancias en sangre tardan un tiempo en reflejarse en el LCR, puesto que deben atravesar la barrera hematoenceflica. La difusin del CO2 es rpida, mientras que la de H+ y HCO3- requiere ms tiempo. Variaciones del pH arterial alcanzan el lquido intersticial intracraneal con un retraso de treinta minutos y con una amplitud reducida; mientras que el CO2 difunde en poco ms de un minuto (1,10,41). En definitiva, la concentracin de hidrogeniones en el lquido extracelular cerebral depende de muchos factores: gases en sangre arterial y venosa, tamponamiento intracelular de H+, transporte selectivo a travs de la barrera hematoenceflica, diferencia de potencial entre sangre y LCR, y actividad metablica (41). El mecanismo de quimiorrecepcin del CO2 todava permanece sin aclarar, aunque existen evidencias de que es mediado por neurotransmisores como el cido -amino-butrico (GABA), glutamato (42), neurotensina y taquiquininas (34). El incremento de hidrogeniones en el lquido extracelular cerebral aumenta la ventilacin a travs de un mecanismo colinrgico muscarnico, en el que la acetilcolina desempea un papel fundamental (43).
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
18 Cuando la PCO2 alveolar aumenta, se desencadena un incremento compensador de la ventilacin, que requiere un tiempo para estabilizarse. La pendiente con la que se incrementa la ventilacin en relacin al aumento de la PCO2 representa la sensibilidad al CO2. Una pendiente de 24 L/min/mmHg se suele considerar normal, aunque existe una gran variabilidad intersujeto (10). Como se muestra en la Figura 5, la PO2 alveolar puede alterar la sensibilidad al anhdrido carbnico. En una situacin de hipoxia, se potencia la sensibilidad al CO2 como se desprende del aumento de la pendiente y del desplazamiento hacia la izquierda de la curva de respuesta a la PCO2 (10). Adems, debe mencionarse que cuando la PCO2 se reduce puede llegar un momento en el que no se produzca ventilacin. Este punto, que se suele calcular por extrapolacin y corresponde a la ordenada en el origen, se conoce como umbral apneico. Si la hiperventilacin reduce la PCO2 alveolar por debajo de dicho umbral, se produciria una apnea. Otro aspecto a considerar es la importancia de la PCO2 sobre el fujo sanguneo cerebral. Cuando la PaCO2 aumenta, se produce una marcada vasodilatacin cerebral, que atena el incremento cerebral de CO2 y, por tanto, puede disminuir la respuesta respiratoria a la hipercapnia.
PO2 alveolar
Ventilacin minuto (L/min)
PCO2 alveolar (mmHg)
Figura 5. Respuesta ventilatoria al incremento de la PaCO2 alveolar en tres situaciones de oxigenacin diferentes. Modificado de Nunn JF (10).
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
19 B. Mecanorreceptores Son receptores sensibles a alteraciones mecnicas de su entorno y se localizan en las vas areas superiores, traquea, pulmones y caja torcica. 1. Receptores de las vas areas superiores Se han identificado receptores en la cavidad nasal, epifaringe y laringe. La estimulacin de los receptores nasales produce estornudos, apnea y bradicardia. Sus aferencias las envan mediante el V par y el nervio olfatorio (1). La estimulacin de los receptores de la epifaringe, que envan aferencias hasta el NTS por el nervio glosofaringeo, induce a esnifar (3). Los receptores larngeos son los ms numerosos. Responden a estmulos irritantes o qumicos y producen tos, respiracin menos profunda, apnea, broncoconstriccin e hipertensin arterial (1,44). Envan aferencias por medio del nervio larngeo superior y su estimulacin antagoniza la respuesta de los quimiorreceptores centrales a la hipercapnia (45). 2. Receptores traqueobronquiales Se diferencian tres tipos: receptores de estiramiento de adaptacin lenta (SARs), receptores de estiramiento de adaptacin rpida (RARs), tambin denominados receptores irritantes, y fibras C (1,46). Los receptores de estiramiento transmiten sus aferencias por fibras vagales mielinizadas y se localizan, sobre todo, en las vas areas. Las terminaciones de las fibras C, que envan su informacin a travs de fibras vagales desmielinizadas, se encuentran principalmente en las pequeas vas areas y en el intersticio pulmonar (17). Su va aferente termina en el ncleo del tracto solitario (3,17,46). Los SARs se localizan entre las clulas de msculo liso de las vas areas y son activados por la insuflacin pulmonar. Acortan la inspiracin y estimulan la actividad espiratoria, con mayor efecto sobre el msculo geniogloso que sobre el diafragma (34). Son los responsables del reflejo de Hering-Breuer en animales, en los que la hiperinsuflacin pulmonar provoca apnea (25). Los RARs, cuyas terminales nerviosas se sitan entre las clulas epiteliales de las vas areas, son excitados por la hiperinsuflacin y por la deflacin pulmonar. Tambin responden a diversos estmulos qumicos, como sustancias nocivas en el aire inspirado, histamina o prostaglandinas (47). Su estimulacin origina tos y, cuando es poco intensa, desencadena una respiracin rpida y superficial (34,47,48). De forma ms esquemtica, se puede considerar que los RARs detectan cambios dinmicos en la mecnica pulmonar, mientras que los SARs detectan cambios estticos (49). Cuando sus terminaciones sensoriales estn sujetas a fuerzas mecnicas, se abren canales inicos activados por el estiramiento, que favorecen la entrada de calcio o sodio y promueven la despolarizacin de membrana. Una vez que el potencial generado por la sumacin de las mltiples y repetitivas despolarizaciones de las terminales sensoriales alcanza un umbral determinado, se abren canales de potasio y sodio activados por voltaje, lo que produce los potenciales de accin de los receptores (49).
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
20 Los receptores inervados por fibras C son estimulados de igual modo que los receptores irritantes, as como por sustancias endgenas como las prostaglandinas, bradiquininas y serotonina (47). Su activacin da lugar a una respiracin con mayor frecuencia y menor volumen corriente y modifica el tono del msculo liso de la va area, con distinta respuesta si el sujeto est en apnea (constriccin) o si respira (relajacin) (50). Ciertas taquiquininas como la sustancia P pueden ser los neurotransmisores utilizados por las fibras C (47). 3. Receptores musculares Las terminaciones nerviosas libres, las terminaciones nerviosas encapsuladas, los rganos tendinosos de Golgi y los husos neuromusculares, son los cuatro tipos de receptores musculares conocidos. Si bien, slo existe evidencia de los dos ltimos en los msculos respiratorios (1). Su distribucin es variable. En el diafragma predominan los rganos de Golgi (51) y en los msculos intercostales son ms numerosos los husos neuromusculares (1). Los rganos tendinosos de Golgi controlan, sobre todo, la fuerza muscular generada, y los husos neuromusculares la longitud de la fibra muscular (3). La informacin aferente desde estos receptores es procesada principalmente a nivel segmentario medular; donde interviene, junto con fibras procedentes de estructuras superiores, en la regulacin de la musculatura respiratoria mediante la activacin agonista-antagonista (1). Existen, no obstante, tractos ascendentes que se dirigen desde el diafragma a los grupos respiratorios dorsal y ventral (52) e incluso hasta la corteza cerebral (1).
3. EFECTORES Los principales efectores son los msculos respiratorios, aunque existen otros grupos musculares que tambin participan en el sistema regulador. 1. Msculos esquelticos de la orofaringe Se ha comprobado que la actividad electromiogrfica se incrementa durante la inspiracin en los msculos geniogloso, gloso, suprahioideo, infrahioideo y farngeos, y que aumenta durante la espiracin en los constrictores farngeos inferiores (53,54). 2. Msculos lisos traqueobronquiales Tambin se ha observado una actividad fsica de estas fibras musculares durante la respiracin, caracterizada por contraccin inspiratoria y relajacin espiratoria (55). 3. Msculos respiratorios El impulso neural es transmitido a los msculos respiratorios por motoneuronas y (56). La intensidad de contraccin de los msculos depender de la magnitud del estmulo que lo alcance y de las propiedades intrnsecas de la fibra muscular (3). La descripcin
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
21 de la anatoma y de las caractersticas funcionales de los msculos respiratorios (56-59) se tratan en otros captulos de esta obra. A modo de resumen, se puede considerar que el sistema regulador de la respiracin est dividido en elementos de control y elementos controlados (Figura 6). Las neuronas localizadas en el bulbo proporcionan un patrn de respiracin ideal ("valor de referencia") a las neuronas premotoras bulboespinales. Estas ltimas estimulan a los msculos respiratorios, que ventilan los pulmones y producen cambios en el pH, PaO2 y aporte tisular de oxgeno ("variables controladas"). Las variaciones en el pH y gases sanguneos actan sobre los quimiorreceptores centrales y perifricos. Desde estos receptores son enviadas seales a las neuronas bulbares. Cuando hay diferencias entre los valores de referencia y los valores reales, se modifica la actividad de las neuronas bulboespinales y se producen los cambios apropiados en la ventilacin (60). Mecanorreceptores localizados en los msculos respiratorios y en los pulmones miden las fuerzas de desplazamiento producidas por las neuronas respiratorias y, en consecuencia, modifican el patrn y nivel de su descarga (60,61).
ELEMENTOS DE CONTROL
ELEMENTOS CONTROLADOS
Figura 6. Esquema del funcionamiento del sistema regulador de la respiracin. Tomado de Coleridge y Coleridge (47).
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
22 4. BIBLIOGRAFA 1. Murray JF. Control of breathing. En: Murray JF, editor. The normal lung. The basis for diagnosis and treatment of pulmonary disease. 2 ed. Philadelphia: WB Saunders Company, 1986; p. 233-260. 2. Campbell EJM, Howell JBL. The sensation of breathlessness. Br Med Bull 1963; 19: 36-40. 3. Berger AJ, Mitchell RA, Severinghaus JW. Regulation of respiration. N Engl J Med 1977; 297: 92-97, 138-143, 194-201. 4. Cherniack NS, Altose MD. Respiratory responses to ventilatory loading. En: Hornbein TF, editor. Regulation of Breathing. II. New York: Marcel Dekker, 1981; p. 905-964. 5. Long S, Duffin J. The neuronal determinants of respiratory rhythm. Prog Neurobiol 1986; 27: 101-182. 6. Cohen MI. Central determinants of respiratory rhythm. Anun Rev Physiol 1981; 43: 91-104. 7. Lipski J, Trzebski A, Kubina L. Excitability changes of dorsal inspiratory neurons during lung inflations as studied by measurement of antidromic invasion latencies. Brain Res 1979; 161: 25-38. 8. Shannon R. Intercostal and abdominal muscle afferent influence on medullary dorsal respiratory group neurons. Respir Physiol 1980; 39: 73-94. 9. Loewy AD, Burton H. Nuclei of the solitary tract: Efferent projections to the lower brain stem and spinal cord of the cat. J Comp Neurol 1978; 181: 421-450. 10. Nunn JF. Control of breathing. En: Nunn JF, editor. Nunn's Applied Respiratory Physiology. 4 ed. Cambridge: Butterworth-Heinemann Ltd, 1993; p. 90-116. 11. Feldman JL, Cohen MI, Wolotsky P. Powerful inhibition of pontine respiratory neurons by pulmonary afferent activity. Brain Res 1976; 104: 341-346. 12. Cohen MI, Feldman JL. Models of respiratory phase-switching. Fred Proc 1977; 36: 2367-2374. 13.Taylor EK, Duffin J, Vachon BR, McCracken DH. The recruitment times and firing patterns of the medullary respiratory neurones of the cat. Respir Physiol 1978; 34: 247-266. 14. Mitchell RA. Location and function of medullary respiratory neurons. Am Rev Respir Dis 1977; 115 (Suppl): 209-216.
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
23 15. Baker JP Jr, Frazier DT, Hanley M, Zechman FW Jr. Behavior of expiratory neurons in response to mechanical and chemical loading. Respir Physiol 1979; 36: 337-351. 16. Kalia MP. Anatomical organization of central respiratory neurons. Annu Rev Physiol 1981; 43: 105-120. 17. Cherniack NS. Control of breathing in COPD. En: Cherniack NS, editor. Chronic obstructive pulmonary disease. Philadelphia: WB Saunders Co., 1991; p. 117-126. 18. Song G, Poon C-S. Functional and structural models of pontine modulation of mechanoreceptor an chemoreceptor reflexes. Respir Physiol Neurobiol 2004; 143: 281-292. 19. Orem J. Medullary respiratory neuron activity: Relationship to tonic and phasic REM sleep. J Appl Physiol 1980; 48:54-65. 20. Aminoff MJ, Sears TA. Spinal integration of segmental, cortical and breathing inputs to thoracic respiratory motoneurones. J Physiol (London) 1971; 215: 557-575. 21. Sears TA. Central rhythm generation and spinal integration. Chest 1990; 97 (Suppl.3): 45S-51S. 22. Von Euler C. On the central pattern generator for the basic breathing rhythmicity. J Appl Physiol Respir Environ Exerc Physiol 1983; 55: 1647-1659. 23. Lipski J, Duffin J. An electrophysiological investigation of propiospinal inspiratory neurons in the upper cervical cord of the cat. Exp Brain Res 1986; 61: 625-637. 24. Richter DW. Generation and maintenance of the respiratory rhythm. J Exp Biol 1982; 100: 93-97. 25. Von Euler C. Brain stem mechanisms for generation and control of breathing pattern. En: Chernicak NS, Widdicombe JG, editores. Handbook of Physiology. Bethesda: American Physiological Society, 1986; p. 1-67. 26. Smith JC, Ellenberger HH, Ballanyi K, Richter DW, Feldman JL. Pre-Btzinger complex: a brainstem region that may generate respiratory rhythm in mammals. Science 1991; 254: 726-729. 27. Ramirez JM, Viemari JC. Determinants of inspiratory activity. Respir Physiol Neurobiol 2005; 147: 145-157. 28. Rocher A, Geijo-Barrientos E, Caceres AI, Rigual R, Gonzalez C, Almaraz L. Role of voltaje-dependent calcium channels in stimulus-secretion coupling in rabbit carotid body chemoreceptor cells. J Physiol 2005;562:407-420.
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
24 29. Ramirez JM, Truba AK, Pea F. Pacemaker neurons and neuronal networks: an integrative view. Curr Opin Nerobiol 2004; 14: 665-674. 30. Milic-Emili J. Recent advantages in clinical assessment of control of breathing. Lung 1982; 160: 1-17. 31. Burki NK. Measurements of ventilatory regulation. Clin Chest Med 1989; 10: 215-226. 32. Petersen ES. The control of breathing pattern. En: Whipp BJ, editor. The control of breathing in man. Physiological Society Study Guides. Manchester: Manchester University Press 1987; p. 1-28. 33. Gardner WN. The relation between tidal volume and inspiratory and expiratory times during steady-state carbon dioxide inhalation in man. J Physiol 1977; 272: 591-611. 34. Cherniack NS. The central nervous system and respiratory muscle coordination. Chest 1990; 97 (Suppl.3): 52S-57S. 35. Allen RM. Location of neurones with cardiovascular and respiratory actions at the ventral surface of the cat's medulla. Neuroscience 1986; 18: 43-49. 36. Sato M, Severinghaus JW, Basbaum AI. Medullary CO2 chemoreceptor neuron identification by c-fos immunocytochemistry. J Appl Physiol 1992; 73: 96-100. 37. Bruce EN, Cherniack NS. Central chemoreceptors. J Appl Physiol 1987; 62: 389-402. 38. Nattie EE. Central chemosensitivity, sleep, and wakefulness. Respir Physiol 2001; 129: 257-268. 39. Mitra J, Prabhakar NR, Pantaleo T, et al. Do structures in the region of the nucleus paragigantocellularis integrate and mediate ventilatory drive inputs?. Soc Neurosci Abst 1986; 12: 304. 40. Coates EL, Li A, Nattie EE. Widespread sites of brain stem ventilatory chemoreceptors. J Appl Physiol 1993; 75: 5-14. 41. Trammeling GJ, Quanjer PhH. Regulation of breathing. En: Trammeling GJ, Quanjer PhH, editores. Contours of breathing. Leiden: Boehringer Ingelheim International GmbH., 1983; p. 195-243. 42. Kazemi H, Hoop B. Glutamic acid and -aminobutyric acid neurotransmitters in central control of breathing. J Appl Physiol 1991; 70: 1-7. 43. Burton MD, Johson DC, Kazemi H. CSF acidosis augments ventilation through cholinergic mechanisms. J Appl Physiol 1989; 66: 2565-2572.
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
25 44. Van Lunteren E, Van de Graaff WB, Parker DM, Mitra J, Haxhin MA, Strohl KP, et al. Nasal and laryngeal reflex responses to negative upper airway pressure. J Appl Physiol 1984; 56: 746-752. 45. Van Vliet BN, Uenishi M. Antagonistic interaction of laryngeal and central chemoreceptor respiratory reflexes. J Appl Physiol 1992; 72: 643-649. 46. Richardson PS, Peatfield AC. Reflexes concerned in the defense of the lungs. Bull Europ Physiopathol Respir 1981; 17: 979-1012. 47. Coleridge HM, Coleridge JCG. Reflexes evoked form the tracheobronchial tree and lungs. En: Chernicak NS, Widdicombe JG, editores. Handbook of Physiology. Bethesda: American Physiological Society, 1986; p. 395-431. 48. Whipp BJ, Wasserman K. Carotid bodies and ventilatory control dynamics in man. Fred Proc 1980; 39: 2668-2673. 49. Yu J. Airway mechanosensors. Respir Physiol Neurobiol 2005;148:217-243. 50. Haxhin MA, Deal EC, Cherniack NS. Influence of respiratory drive on airway responses to excitation of lung C fibers. J Appl Physiol 1989; 67: 203-209. 51. Frazier DT, Revelette WR. Role of phrenic nerve afferents in the control of breathing. J Appl Physiol 1991; 70: 491-496. 52. Speck DF, Revelette WR. Excitation of dorsal and ventral respiratory group neurons by phrenic nerve afferents. J Appl Physiol 1987; 62: 946-952. 53. Rothstein RJ, Narce SL, de Berry-Borowiecki B, Blanks RHI. Respiratory-related activity of upper airway muscles in anesthetized rabbit. J Appl Physiol 1983; 55: 1830-1836. 54. Orem J, Norris P, Lydic R. Laryngeal abductor activity during sleep. Chest 1978; 73 (Supp.2): 300-301. 55. Mitchell RA, Herbert DA, Baker DG. Inspiratory rhythm in airway smooth muscle tone. J Appl Physiol 1985; 58: 911-920. 56. Rochester D. Test of respiratory muscle function. Clin Chest Med 1988; 2: 249-261. 57. Herrera A, Garca-Ro F. Estudio funcional de los msculos respiratorios. Arch Bronconeumol 2000; 36: 146-158. 58. Respiratory movements and muscles. En: Shneerson J, editor. Disorders of Ventilation. Oxford: Blackwell Scientific Publications, 1988; p. 13-33.
ESTRUCTURAS REGULADORAS DE LA RESPIRACIN
26 59. Gea Guiral J. Msculos respiratorios. En: Pino Garca JM, Garca Ro F, editores. Estudio de la funcin respiratoria. Madrid, Sanitaria 2000, 2005. 60. Longobardo GS, Cherniack NS. Abnormalities in respiratory rhythm. En: Chernicak NS, Widdicombe JG, editores. Handbook of Physiology. Bethesda: American Physiological Society, 1986; p. 729-750. 61. Whipp BJ. Ventilatory control during exercise. Annu Rev Physiol 1984; 45: 393-415.
CAPTULO 2
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
ANA OBESO, CONSTANCIO GONZLEZ Y ASUNCIN ROCHER
Departamento de Bioqumica y Biologa Molecular y Fisiologa e Instituto de Biologa y Gentica Molecular. Facultad de Medicina de la Universidad de Valladolid y Consejo Superior de Investigaciones Cientficas. C/ Ramn y Cajal n 7 47005 Valladolid
Prof. Constancio Gonzlez Departamento de Bioqumica y Biologa Molecular y Fisiologa e Instituto de Biologa y Gentica Molecular. Facultad de Medicina de la Universidad de Valladolid y Consejo Superior de Investigaciones Cientficas. C/ Ramn y Cajal n 7 47005 Valladolid Telfono: +34 983 423089 Fax: +34 983 423588 e-mail: constanc@ibgm.uva.es
PALABRAS CLAVE: Cuerpo carotdeo, clulas quimiorreceptoras, hipoxia, catecolaminas, adenosina, ATP, canales de potasio.
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
30 NDICE DE CONTENIDOS: 1. Introduccin. Localizacin anatmica, estructura e inervacin de los quimiorreceptores arteriales 2. Aspectos histricos 2.1. Los primeros tiempos 2.2. El cuerpo carotdeo como receptor sensorial. El nacimiento de los quimiorreceptores arteriales 3. Funciones de los quimiorreceptores arteriales y su control 3.1. Funcin del cuerpo carotdeo en normoxia e hipoxia hipxica 3.2. Funcin del cuerpo carotdeo en las hipoxias anmica, de estasis e histotxica 3.3. Funcin del cuerpo carotdeo en hipercapnia y acidosis 3.4. Funciones del cuerpo carotdeo en el ejercicio 3.5. Otras funciones del cuerpo carotdeo 3.6. Control eferente de la funcin del cuerpo carotdeo 4. Fisiologa general del CC. Mecanismos de funcionamiento de las CQ a nivel celular y molecular 4.1. Deteccin y transduccin de los estmulos a) La hipoxia b) Estmulos cido-hipercpnicos 4.2. Neurotransmisin en el cuerpo carotdeo 5. Bibliografa
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
31 1. INTRODUCCIN Al escribir este captulo, hemos pensado que es importante presentar, aunque sea de forma breve, algunos datos histricos sobre el desarrollo de nuestros conocimientos en este campo de los quimiorreceptores arteriales, centrndonos en el cuerpo carotdeo (CC). Sin embargo, antes de nada, consideramos necesario hacer una descripcin, aunque sea sucinta, de la organizacin estructural del CC para facilitar al lector seguir el resto de la informacin. El CC de los mamferos es un rgano par, pequeo, y localizado en la bifurcacin carotdea o en una de las ramas arteriales que nacen del seno carotdeo. La localizacin precisa en el conejo se muestra en la Figura 1A. El CC est organizado en islotes celulares (Figura 1B) separados entre s por tejido conectivo y formados por dos tipos celulares, las clulas quimiorreceptores (CQ) o tipo I y las clulas sustentaculares o tipo II. Este segundo tipo celular componente del parnquima del CC no fue identificado hasta el ao 1954 por De Kock. Las primeras son ms numerosas, tienen un ncleo redondo y claro y estn localizadas hacia el centro de los islotes; las segundas rodean a las CQ y tienen un ncleo en forma discoidea con cromatina densa.
Figura 1. Esquema de la bifurcacin carotdea de conejo (A) y de un islote celular del cuerpo carotdeo (B). A: 1, cartida comn; 2 y 3, cartidas interna y externa; 4, cuerpo carotdeo; 5 y 6, ganglio petroso y nervio del seno carotdeo; 7 y 8, ganglio cervical superior y nervios ganglioglomerulares; 9, ganglio nodoso. B: 1, CQ; 2, clula sustentacular; 3, vesculas sinpticas en las CQ; 4 y 5, terminaciones y fibra sensorial; 6, capilares sanguneos. (Gonzlez, C. et al. Trends Neurosci. 15: 146-153, 1992).
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
32 El CC es un rgano muy vascularizado con un 25-30% de la superficie de una seccin histolgica ocupada por la luz de los vasos (en cerebro, la superficie de los vasos es ~5%). El CC es el rgano con mayor flujo sanguneo del organismo (1.3 l/100g/min) y tiene un consumo de O2 de 1.5 ml/100g/min, lo que implica que la diferencia arteriovenosa de O2 en el CC es mnima. Los capilares establecen una relacin estrecha con los islotes celulares con distancias mximas de capilares a tejido del orden de 20 m, habindose estimado que en condiciones de normoxia no habra ningn rea del CC con PO2 inferiores a 40 mmHg. El parnquima glmico medido como superficie inmunopositiva a tirosina hidroxilasa representa en la rata el 25% de la superficie del CC en cortes seriados del rgano, cuando ste es fijado con una presin de perfusin de 100 mmHg (Figura 2).
Figura 2. Cortes histolgicos de CC de rata, obtenidos en torno a su ecuador. A la izquierda el corte ha sido sometido a una reaccin inmunocitoqumica para tirosina hidroxilasa (TH). Las clulas positivas aparecen en verde. A la derecha, el mismo corte ha sido contrateido con cresil violeta para obtener una imagen de la estructura general del tejido. Obsrvese la gran densidad de capilares, que aparecen como espacios vacos, en el tejido del CC. Las CQ son muy ricas en catecolaminas, principalmente dopamina, y por tanto la tirosina hidroxilasa, enzima limitante en la sntesis de estas aminas bigenas, es un marcador especfico de las CQ. Aunque las terminaciones simpticas, que penetran en el CC, tambin son positivas para este enzima, son muy escasas en los islotes celulares.
El CC recibe inervacin sensorial y vegetativa. La inervacin sensorial la recibe por el nervio del seno carotdeo (NSC), rama del glosofarngeo IX par craneal. Parte de las fibras sensoriales del NSC penetran en el CC y hacen sinapsis con las CQ; desde el punto de vista funcional estas fibras son las fibras quimiorreceptoras y el otro contingente de fibras sensoriales del NSC alcanza el seno carotdeo, ya sea atravesando o rodeando el CC, y funcionalmente constituyen los barorreceptores carotdeos. En la rata, es muy comn observar cmo el NSC se bifurca en dos y cmo uno de los dos nerviecillos resultantes penetra en el CC mientras que el otro se dirige al seno carotdeo. Fibras vegetativas simpticas y parasimpticas que entran en el CC inervan fundamentalmente los vasos sanguneos. Las CQ son redondeadas y tienen un dimetro de unos 10 m. Adems de las organelas celulares generales contienen abundantes vesculas de ncleo denso descritas por Lever y Boyd
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
33 en 1957 como similares a las de las clulas adrenomedulares y que hoy sabemos contienen catecolaminas (CA; dopamina, DA y norepinefrina, NE) y ATP. Las CQ tambin poseen vesculas de ncleo claro que clsicamente se ha dicho que contendran acetilcolina (ACh). Adems, las CQ poseen pptidos opioides, pptido atrial natriurtico, galanina, etc. que estn tambin almacenados en las vesculas coexistiendo con las CA, la ACh y el ATP. La presencia o no de sustancia P en las CQ ha sido causa de intenso debate en aos recientes: la ausencia de RNAm en CC de rata sera consistente con la ausencia de sustancia P en las clulas. Las terminaciones nerviosas sensoriales que hacen sinapsis con las CQ son de forma y tamao variable y contienen vesculas de ncleo claro y de ncleo denso. Desde el punto de vista estructural las sinapsis son de dos tipos: de direccin clulas hacia las terminaciones nerviosas sensoriales, la mayora de ellas, y el resto tienen la direccin opuesta. Esta bidireccionalidad estructural de las sinapsis hace posible que neurotransmisores presentes en las terminaciones sensoriales se liberen posibilitando un control por retroalimentacin de la propia actividad de las CQ. Las fibras sensoriales que inervan a las CQ contienen DA, taquiqunina A, sustancia P, pptido relacionado con el gen de la calcitonina y neuropptido Y, con diferentes patrones de incidencia y coexistencia en distintas fibras y especies animales. Los cuerpos articos, inervados sensorialmente por el nervio artico, que es una rama del vago, estn localizados en la cara inferior del cayado artico y su organizacin estructural es comparable a la de los cuerpos carotdeos, aunque parece que su densidad de capilares y por tanto su flujo sanguneo es mucho menor. Se ha sugerido que este bajo aporte sanguneo es el responsable de que los cuerpos articos respondan a las hipoxias anmica y de estasis, o lo que es lo mismo que los CC detecten y respondan a la disminucin de la PO2 mientras que los cuerpos articos lo hagan a la disminucin del contenido arterial de O2. 2. ASPECTOS HISTRICOS 2.1. Los primeros tiempos(1) El CC fue descrito por primera vez por Taube (1743) que en su Tesis Doctoral rese por primera vez la existencia de un ganglium minutum localizado en el ngulo intercarotdeo, interconectado con el ganglio cervical superior y proveyendo de ramitas nerviosas a las arterias vecinas. Haller en 1762 y Neubauer en 1772 se refirieron al CC con los nombres de ganglium exiguum y ganglium parvum. Andersch, discpulo de Haller dio por primera vez nombre propio al CC en 1797 al referirse a l con el nombre de ganglium intercaroticum. Fue Andersch quien descubriera el ganglio petroso inferior o principal del nervio glosofarngeo. Mayer en 1833 redescubre el CC en el hombre y le da el mismo nombre y lo describe como un grano de arroz, compacto y firme al tacto, rodeado de una importante red vascular superficial y unido a la pared arterial por un ligamento. Mayer constat la constancia del gan(1)
Todas las citas dadas en esta seccin histrica han sido tomadas del libro de Adams (Adams, W.E. The comparative morphology of the carotid body and carotid sinus. Thomas, Springfield, IL. 1958). As pues, remitimos al lector a esta obra para la documentacin bibliogrfica. .
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
34 glium intercaroticum, no se trataba de un ganglio inconstante como afirmaban sus predecesores, sino de un ganglio constante, que adems de las ramas del ganglio cervical superior reciba inervacin constante a travs de una ramilla del glosofarngeo en todas las especies por l estudiadas (hombre, caballo y ternera). Esta es la primera descripcin de lo que hoy conocemos como nervio del seno carotdeo o nervio de Hering. Ntese que la denominacin del nervio del seno carotdeo puede inducir a error ya que el nervio de Hering, que debera llamarse nervio de Mayer, inerva, adems del seno carotdeo origen del componente carotdeo del reflejo barorreceptor encaminado a mantener la constancia de la presin arterial, el CC origen del reflejo quimiorreceptor encaminado a mantener la constancia de la PO2 y de la PCO2 y pH arteriales. Luscka (1862) encontraba que el ganglium intercaroticum posea una estructura glandular, y no ganglionar, por lo que le atribuy el nombre de glndula intercarotica al CC. Luscka encontraba que la relacin entre el CC y el simptico eran anlogas a las que estableca la mdula suprarrenal con el simptico abdominal. La autoridad cientfica de Luscka influy profundamente en la literatura, de tal forma que el trmino de glndula intercartica o glndula carotdea fue muy utilizado en la literatura (De Castro, 1926) y an hoy en da algn autor se refiere al CC como carotid gland. Arnold (1865) describi al CC como un conglomerado redondeado de nervios, vasos y clulas en la organizacin tpica definitoria del tejido glmico (A small body surrounded by many nerve fibers, consisting of an anastomosis between fine arterioles and veins) y en consecuencia lo bautiz como glomeruli arteriosi intercarotici. Estudios embriolgicos del propio Luscka y de Stieda (1881) atribuyen al CC un origen endodrmico, como cabe esperar para una estructura glandular, mientras que Kastschenko (1887) y Marchand (1891) suponen que tiene un origen mesododrmico para constituir un engrosamiento de la adventicia intercarotdea ricamente vascularizado. En 1882 Stilling public un artculo que teniendo la intencin de ser reconciliador supuso una gran innovacin: los vasos del CC seran de origen endodrmico, pero las clulas tpicas del rgano son independientes de los vasos en su origen embriolgico y adems se tornan marrones al tratarlas con dicromato potsico al igual que ocurre con las de la mdula adrenal. Kohn (1900) desarroll las ideas de Stilling y afirmaba que las clulas tpicas del CC tenan el mismo origen que las clulas del ganglio cervical superior. Encontr adems que haba pequeos rganos similares al CC en asociacin a vasos sanguneos en otras muchas reas y para unificar bajo el mismo nombre a todas estas estructuras cre el nombre de paraganglio, llamando al CC paraganglio intercartico y en razn de su tincin con las sales de cromo lo homolog con la mdula adrenal a la que llam paraganglio abdominal. Hoy el trmino paraganglio abarca un conjunto de rganulos o estructuras diseminadas por toda la economa corporal que contienen clulas con granos secretores y una rica inervacin. El CC es considerado como el paraganglio ms definido y a los tumores del CC se les denomina paragangliomas o chemodectomas. Por las mismas fechas Oliver y Schafer (1895) inician el estudio de la actividad secretora de la mdula adrenal y pronto se pasa a estudiar el poder vasopresor de los extractos de la mdula adrenal, y cuando se pasa a estudiar el efecto de los extractos del CC sobre la presin arterial se encuentra que produce hipotensin y enlentecimiento del pulso (Gomez, 1908), hallazgos estos ltimos ratificados por Christie en 1933. Hoy sabemos que el CC es la estructura corporal ms rica en dopamina, llegando en algunas especies a casi duplicar el contenido de los ganglios basales o del tubrculo olfactorio, por lo que la accin vasodilatadora de los extractos del CC es fcilmente explicable. Adems varios autores negaron la cromafinidad del CC, entre ellos Fernando de Castro. De otro lado, el propio Fernando
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
35 de Castro (1926) demostraba en experimentos de degeneracin walleriana que las clulas glandulares del CC no reciban inervacin de la cadena simptica como lo haca la mdula adrenal, sino, conclua en su trabajo, de un sistema autnomo bulbar que desde el tronco del encfalo enviaba sus axones por el nervio intercarotdeo (NSC) al CC. La ausencia de inervacin simptica y la cromafinidad dudosa llevaron a Watzka (1930) a crear el concepto de paraganglio parasimptico no cromafin en oposicin a los paraganglios simpticos, ejemplarizados por la medula adrenal, que son claramente cromafines. 2.2. El cuerpo carotdeo como receptor sensorial. El nacimiento de los quimiorreceptores arteriales El descubrimiento de la funcin sensorial es el fruto de unos experimentos impecablemente hechos por D. Fernando de Castro y publicados en 1928 (1). En este trabajo, adems de una descripcin bellsima de las estructuras del CC y de las terminaciones barorreceptoras del seno carotdeo y de los vasos intraglmicos, realiz experimentos de denervacin proximal (ceflico) y distal al ganglio petroso y tras verificar que la denervacin (la discontinuidad del glosofarngeo) era correcta, ya que la estimulacin del cabo distal no generaba la hipotensin y bradicardia del reflejo de Hering, estudi unos das ms tarde estructuralmente el CC encontrando que la denervacin proximal al ganglio petroso no afectaba la estructura del CC en lo esencial y no produca prdida de la inervacin de las clulas del CC, mientras que la distal eliminaba toda la inervacin de estas clulas. Veamos en las propias palabras de De Castro algunas de sus consideraciones y conclusiones (lo escrito en negrita es de los autores de esta captulo). ....En suma, el Glomus caroticum est inervado por fibras de naturaleza centrpeta, cuyo centro trfico asienta en los ganglios sensoriales del nervio glosofarngeo, y no por fibras centrfugas o por fibras secretoras como es el caso para las glndulas; esto es precisamente lo que hace suponer que el Glomus caroticum sea un rgano sensorial especial... (F. de Castro (1928). Trabajos del Laboratorio de Investigaciones Biolgicas. 25: pag.71) (1). .... Desde nuestro punto de vista, el Glomus representa un rgano con receptores sensoriales mltiples, provisto de clulas receptoras especiales semejantes a las de otros rganos sensoriales. De otro lado, no compartimos el punto de vista de Drner y Jacoborici, Mistezcu y Pop, de que se trata de un rgano destinado a recoger las variaciones de la presin sangunea. El territorio del "Sinusreflex" se encuentra en el propio seno carotdeo. Suponemos, como hiptesis verosmil, que el Glomus caroticum representa un rgano sensorial, el nico hasta el presente, encargado de recibir ciertas variaciones cualitativas de la sangre, funcin que, tal vez por accin refleja, tendra su repercusin sobre la actividad funcional de otros rganos.... (F. de Castro (1928). Trabajos del Laboratorio de Investigaciones Biolgicas. 25: pag.71) (1). ....Los elementos parenquimatosos del Glomus poseen tericamente dos polos: uno vascular y otro nervioso. Por uno de los polos, estas clulas se encuentran en relacin ntima con los capilares sinusoidales.... Por el otro polo las clulas estn inervadas.... La fibra nerviosa, por tanto, no ser estimulada directamente por la sangre, sino por el intermedio de las clulas epiteliales del rgano, las cuales, tal y como obliga a suponer su estructura, poseen una funcin secretora activa que intervendra en el fenmeno de la estimulacin centrpeta (F. de Castro (1928). Trabajos del Laboratorio de Investigaciones Biolgicas. 25: pag.71) (1).
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
36 En estos tres prrafos De Castro termina con el concepto del CC como glndula, lo define como rgano sensorial origen de reflejos, excluye el que se trate de un barorreceptor, crea el concepto de quimiorreceptor variaciones cualitativas de la sangre, adscribe a las clulas glmicas o parenquimatosas del CC (a partir de aqu clulas quimiorreceptoras, CQ, o tipo I) la funcin de sensores y define la existencia de una sinapsis qumica entre las CQ y las terminaciones sensoriales. A la fecha de hoy todas sus propuestas siguen vigentes y adems, seguimos intentando definir tanto los mecanismos por los que las CQ detectan esas modificaciones cualitativas de la sangre (la PO2, la PCO2 y la [H+]) como la identidad molecular del neurotransmisor responsable de la estimulacin centrpeta y el significado de los otros mltiples neurotransmisores que las CQ poseen. Para concluir con el nacimiento de los quimiorreceptores, y con las ideas avanzadas por De Castro, respecto al significado del CC como origen de reflejos, vamos a citar el ttulo del trabajo de Corneille Heymans que le vali el Premio Nobel en 1938: HEYMANS, C., J. J. BOUCKAERT, AND L. DAUTREBANDE. Sinus carotidien et rflexes respiratoires, II. Influences respiratoires rflexes de l'acidose, de l'alcalose, de l'anhydride carbonique, de l'ion hydrogene et de l'anoxmie: Sinus carotidiens et echanges respiratoires dans les poumons et au del des poumons. Arch. Int. Pharmacodyn. Ther. 39: 400 408, 1930) (2).
Fernando de Castro Rodrguez (1896-1967)
Corneille J. F. Heymans (1892-1968)
Los prrafos que siguen estn tomados literalmente del Discurso de G. Liljestrand en su Presentacin de Heymans como Premio Nobel en 1938(2). ...The glomus consists of a small mass of very fine intertwining vessels arising from the internal carotid and enclo(2)
Tomado de http://nobelprize.org/medicine/laureates/1938/press.html
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
37 sing various different types of cells. It has been considered by some as being a sort of endocrine gland similar to the medulla of the suprarenal glands. De Castro, however, in 1927 demonstrated that the anatomy of the glomus could in no way be compared to that of the suprarenal medulla. De Castro suggested rather that the glomus was an organ whose function was to react to variations in the composition of the blood, in other words an internal gustatory organ with special chemo-receptors. In 1931, Bouckaert, Dautrebande, and Heymans undertook to find out whether these supposed chemo-receptors were responsible for the respiratory reflexes produced by modifications in the composition of the blood. By localized destruction in the sinus area they had been able to stop reflexes initiated by pressure changes, but respiratory reflexes could still continue to occur in answer to changes in the composition of the blood. Other experiments showed that Heymans's concepts on the important role played by the glomus in the reflex control of respiration by the chemical composition of the blood were undoubtedly correct. La existencia de unos receptores en la regin cardioartica capaces de modificar la ventilacin fue puesta de manifiesto por los Heymans, padre e hijo, en 1927. En 1935 Nonidez y Muratori de forma independiente describieron el paraganglio o glomus artico a nivel histolgico, pero no fue hasta 1939 cuando Comroe los localiz funcionalmente y puso juntos en experimentos de perfusiones selectivas los hallazgos previos de los Heymans y los datos histolgicos. Es comn atribuir el crdito del descubrimientos de los cuerpos articos a Julius Comroe en su artculo The location and function of the chemoreceptors of the aorta. Am. J. Physiol. 127: 176-191, 1939 (3).
3. FUNCIONES DE LOS QUIMIORRECEPTORES ARTERIALES Y SU CONTROL 3.1. Funcin del cuerpo carotdeo en normoxia e hipoxia hipxica En este apartado resumiremos de forma muy sucinta las funciones reflejas del CC. Cuando lo consideremos oportuno haremos paralelismos entre las acciones reflejas mediadas por el CC y la actividad en el NSC o nos referiremos indistintamente a una u otra, ya que la relacin entre la salida de informacin del CC va NSC y las respuestas ventilatorias son muy similares. Comenzaremos por los efectos del reflejo quimiorreceptor sobre la ventilacin. En animales de experimentacin en estado de normoxia y eucapnia se observan descargas en fibras aisladas del NSC a frecuencia baja (~1-5 potenciales de accin/segundo), siendo posible registrar actividad en fibra nica con una PO2 de hasta 700 mmHg, lo que indica que la actividad quimiorreceptora no tiene un umbral absoluto. Al igual que pasa con la actividad elctrica, hay una depresin ventilatoria de hasta el 20-30% cuando se pasa de respirar aire a respirar 100% de O2, debido a la desactivacin del reflejo quimiorreceptor, lo que implica que el CC contribuye al mantenimiento normal de la ventilacin/minuto normal. Este efecto se observa tanto en animales como en humanos despiertos o anestesiados y se hace ms marcado si en vez de aire se esta respirando una mezcla de gases parcialmente hipxica (Figura 3).
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
38
VE ml/min
Impulsos/seg.
Segundos Figura 3. Test de Dejours. En la parte superior, se representa la actividad en el nervio del seno. Mientras el animal (gato) respira 13% de O2 la actividad es prxima a 15 impulsos/s; al respirar 100% O2 durante 5 s las descargas disminuyen hasta 2-3 impulsos/s y la actividad se recupera hacia los 40 s despus de volver a respirar 13% de O2. La parte inferior representa ventilacin a lo largo del tiempo en idnticas condiciones experimentales. Tomada de Dejours (4).
Una disminucin de la PO2 va acompaada de un aumento de la frecuencia de descarga en el NSC. Si a un animal de experimentacin le ventilamos con O2 puro y con mezclas que contengan concentraciones de O2 decrecientes y registramos actividad elctrica en fibras aisladas del NSC, nos encontramos una relacin como la que recoge la Figura 4. En el rango de PO2 de 600 mmHg a 70 mmHg la frecuencia de descarga aumenta de forma lineal con una pendiente baja, en torno a 70-75 mmHg existe un cambio brusco en la pendiente, siguiendo un aumento de la descargas de manera casi exponencial hasta 10-15 mmHg. La respuesta ventilatoria sigue el mismo patrn que las descargas en el NSC. La relacin entre PO2 y descargas en el NSC o ventilacin, est desplazada hacia arriba a medida que aumenta la PCO2 y hacia abajo en el caso contrario (Figura 4). Hay que percatarse de que este efecto de la hipoxia sobre la ventilacin es el que posibilita la adaptacin a las grandes alturas: a medida que se asciende cae la presin baromtrica y de forma paralela la PO2 en el aire que inspiramos y la PO2 en gas alveolar y en sangre arterial. Sin embargo, la hiperventilacin refleja lava, casi instantneamente, los alveolos (y la sangre) de CO2 posibilitando que la PO2 no disminuya en el gas alveolar, y
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
39
Figura 4. Lnea continua, relacin tipo entre PO2 en sangre arterial y frecuencia de actividad elctrica en fibras aisladas del NSC. Las lneas discontinuas recogen el mismo tipo de relacin a PCO2 arteriales mayores y menores de 40 mmHg (o a pH inferior o superior a 7.40). La relacin entre PaO2 y ventilacin es comparable tanto en forma como en magnitud. La ventilacin puede alcanzar valores de ms de 35 l/min desde una ventilacin basal de unos 5 l/min.
por tanto en sangre arterial, tanto como cabra esperar en razn de la cada de la PO2 en la atmsfera. El lavado alveolar de CO2 (y en sangre arterial) determinara que la relacin entre PaO2 y ventilacin se desplazara hacia abajo, pero una sensibilizacin del reflejo quimiorreceptor que se desarrolla en unas pocas horas de hipoxia sostenida, compensa y sobrepasa el efecto de disminucin de la PCO2 con el resultado neto de una hiperventilacin mayor de la que se observa en tests de hipoxia aguda. Los efectos de la hipoxia sobre el aparato cardiovascular son muy extensos. La hipoxia afecta a la frecuencia y contractibilidad cardiacas, al gasto cardiaco, presin arterial y presin venosa central. Parte de los efectos observados durante la hipoxia son mediados directamente por la hipoxia, parte son producidos directamente por el reflejo quimiorreceptor y parte son debidos a los efectos indirectos provocados por la hiperventilacin generada por el propio reflejo quimiorreceptor. En su conjunto en situacin de hipoxia nos vamos a encontrar con que: 1. La hipoxia produce vasodilatacin e hipotensin excepto en la circulacin pulmonar donde produce vasoconstriccin e hipertensin pulmonar. 2. La hipoxia modifica la contractibilidad cardiaca: aumentndola si se trata de una hipoxia moderada y disminuyndola si es hipoxia intermedia o intensa.
Actividad de los quimiorreceptores carotideos (impulsos/seg)
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
40 3. Si la combinacin de los efectos directo y reflejo de la hipoxia altera la presin arterial, el efecto barorreceptor tiende a corregir esta alteracin. 4. El estimulo hipxico produce una hiperventilacin, lo que supone cambios hemodinmicos (como es el aumento de la presin negativa intratorcica con distensin de venas y aurculas) favoreciendo el retorno venoso y aumentando la frecuencia cardiaca. Este efecto se opone a la accin directa del reflejo quimiorreceptor que tiende a producir bradicardia. 5. En la hiperventilacin, aumenta la distensin de las vas areas que produce efectos reflejos que generan taquicardia e hipotensin. 6. La actividad del reflejo quimiorreceptor provoca la secrecin de catecolaminas de la mdula adrenal, lo que modifica los efectos cardiovasculares tanto directos como indirectos de la hipoxia. 7. El efecto barorreceptor puede alterar los efectos hipxicos directos y los mediados por el reflejo quimiorreceptor sobre corazn, frecuencia y contractibilidad cardiaca. En suma, los efectos son complejos y, an cuando hay diferencias de especie, en el hombre el efecto global de la hipoxia aguda es taquicardia moderada, hipotensin moderada con aumento igualmente moderado del gasto cardiaco. La hipoxia, de forma directa y a travs del reflejo quimiorreceptor, produce una serie de respuestas endocrinas de relevancia fisiolgica. Por ejemplo, la hipoxia en distintas especies animales libera CA de la mdula adrenal (fundamentalmente adrenalina), siendo esta liberacin inhibida al seccionar bilateralmente el nervio esplcnico, lo que indica que el efecto es reflejo. El reflejo lo media el CC porque si se anestesia o se secciona el NSC se abole la respuesta; adems, la estimulacin aferente del CC libera catecolaminas adrenales (5). Tambin de forma refleja, mediada por el CC, la hipoxia aumenta la secrecin de ACTH y cortisol. La hipoxia, tambin va del reflejo quimiorreceptor, produce activacin de neuronas vasopresinrgicas de los ncleos paraventricular y supraptico del hipotlamo con liberacin de vasopresina y se produce un aumento en la secrecin de la hormona antidiurtica de unas 10 veces con estimulacin intensa del reflejo quimiorreceptor. Sin embargo, los cambios en el flujo urinario no son los esperables por el aumento de hormona antidiurtica ya que se produce un aumento del flujo urinario, tal vez mediado por las modificaciones concomitantes en los niveles plasmticos de CA y endotelina-1, que aumentan, y de aldosterona, que disminuye. Queremos considerar tambin el posible papel del reflejo quimiorreceptor en el control de la actividad de los msculos respiratorios estriados de las vas areas superiores, debido a su papel en el control de las resistencias de las vas areas y por su posible alteracin en procesos tan relevantes clnicamente como la enfermedad obstructiva del sueo. Adems, la hipoxia intermitente experimental, diseada para mimetizar el sndrome de apnea del sueo, parece producir una sensibilizacin del reflejo quimiorreceptor superior a la que ocurre durante la aclimatizacin a la hipoxia sostenida de grandes alturas (6). La hiperac-
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
41 tividad exagerada del reflejo quimiorreceptor podra representar uno de los mecanismos patognicos (va una hiperactividad simptica) de la hipertensin y otras alteraciones cardiovasculares que con frecuencia acompaan al sndrome de apnea del sueo (7). Los msculos de la laringe, faringe, lengua y del paladar blando son multifuncionales. Participan en la respiracin, en la vocalizacin, en la deglucin y en los reflejos del estornudo y de la tos (8). El patrn de actividad en los msculos estriados de las vas respiratorias superiores vara a lo largo del ciclo respiratorio. As: a) los cricoaritenoideos posteriores (separadores de las cuerdas vocales y que por tanto mantienen abierta la glotis), los genioglosos (producen un ensanchamiento de la luz farngea y de la comunicacin entre la orofaringe y la laringofaringe) y los genihioideos (producen un efecto sobre la luz farngea sinrgico con el de los genioglosos) se activan durante la inspiracin, aumentando el flujo de aire para cualquier actividad diafragmtica dada. La actividad de estos msculos comienza antes que la del diafragma y dilatan y dan consistencia a las vas areas superiores antes de que empiece el flujo areo, adems la actividad se mantiene durante toda la inspiracin con un patrn de actividad paralelo al del propio flujo areo. La hipoxia, la hipercapnia y muchos agentes farmacolgicos que estimulan los quimiorreceptores del CC activan estos msculos en grado comparable a la activacin diafragmtica y depresores del reflejo quimiorreceptor reducen la actividad de estos msculos dilatadores de las vas areas; b) por el contrario, los tiroaritenoideos (constrictores de la glotis) y los constrictores farngeos que disminuyen todos los dimetros farngeos, se activan durante la espiracin. La contraccin de estos msculos durante la espiracin frena el flujo espiratorio, hacindolo ms lento de lo predecible por la retraccin elstica pasiva del pulmn, y as mejoran la transferencia de gases a nivel alveolar. Durante el sueo, especialmente durante el sueo REM, hay una disminucin de las rdenes motoras, tanto a los msculos inspiratorios (diafragma e intercostales externos) como a los msculos de vas respiratorias altas que se activan durante la inspiracin y dilatan las vas respiratorias (9,10). El resultado de la disminucin de la fuerza de bombeo y el aumento de resistencias en vas superiores, cuando son muy marcados, generan la apnea obstructiva por cierre de las vas areas, cierre que ocurre preferentemente a nivel de la faringe (11). La apnea conduce a un aumento de la PaCO2 y a una disminucin de la PaO2 que activan al CC. El aumento de actividad en el NSC supone un aumento del input a los centros reguladores del ritmo respiratorio y a los centros que controlan el estado de vigilia, produciendo un aumento del esfuerzo respiratorio y una disminucin de las resistencias de las vas areas, asociados frecuentemente a un despertar transitorio, que por s mismo produce aumento de las rdenes motoras a los msculos respiratorios y restauracin de la ventilacin. Contribuye tambin a la restauracin del flujo areo, un reflejo originado en las propias vas areas, justo por debajo de la obstruccin, donde la presin subatmosfrica produce reflejamente aumento de la actividad de todos los msculos dilatadores de vas areas con efectos menores sobre los msculos de bombeo respiratorio (12). Restaurados los gases en sangre y recuperado el sueo se repite otro ciclo de apnea hasta completar frecuencias superiores a 30-40 apneas/hora en casos de sndromes de apnea del sueo severos. Es fcil comprender que cualquier idiosincrasia o anomala anatmica o funcional que suponga un estrechamiento de vas areas con aumento de resistencias al flujo de aire (obesidad, pues el tejido graso perifarngeo determi-
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
42 na el menor calibre farngeo; macroglosia como en el sndrome de Down o mandbula hipoplsica o retrada (retrognatismo) que limitan la comunicacin entre la orofaringe y la laringofaringe; hiperplasia del tejido adenoideo; distonas y distrofias de msculos de vas areas; etc) representa un factor de riesgo para las apneas del sueo (13,14). El reflejo quimiorreceptor aumenta la secrecin de glndulas de la submucosa traqueal. Dada la localizacin de estas glndulas, en vas de calibre grueso, no es comn que se produzca un aumento de las resistencias de las vas areas en hipoxia, exceptuando en patologas que cursen con aumento de secrecin, disminucin de la fluidez del moco o alteracin de la motilidad de los cilios del epitelio traqueal. En la enfermedad pulmonar obstructiva crnica, se dan todos los factores (hipoxia, activacin del reflejo quimiorreceptor, infeccin, aumento de secrecin, tabaquismo, disminucin del movimiento de los cilios, que en conjunto tienden a aumentar las resistencias areas (15). 3.2. Funcin del cuerpo carotdeo en las hipoxias anmica, de estasis e histotxica El CC responde de forma moderada a las hipoxias anmica y de estasis. Disminuciones del hematocrito de hasta el 50% del normal, que estimulan a los quimiorreceptores articos de manera muy intensa, apenas lo hacen a los quimiorreceptores carotdeos. Del mismo modo, una hipotensin de 80 mmHg ya activa a los quimiorreceptores articos, mientras que para estimular a los quimiorreceptores carotdeos hay que bajar la presin a 50 mmHg. Se dice que los CC son sensores rpidos a cambios de gases en sangre y responsables de la respuesta refleja ventilatoria y los cuerpos articos son sensores del flujo circulatorio y de la capacidad del transporte de O2 de sangre y responsables de las respuestas reflejas circulatorias, consistentes en taquicardia y aumento del gasto cardiaco y en menor grado de la presin arterial. La hipoxia histotxica se define como una hipoxia tisular caracterizada por la incapacidad de las clulas para utilizar el O2, lo que se traduce en una PO2 arterial y venosa esencialmente idnticas, o en cualquier caso en una diferencia arteriovenosa de O2 menor de lo normal. Cualquier veneno de los complejos enzimticos de la cadena respiratoria tiende a producir hipoxia histotxica y clsicamente los inhibidores del ltimo complejo enzimtico, tales como el cianuro y la azida, o el monxido de carbono a presiones altas, son los ms efectivos para bloquear totalmente el consumo de O2, aunque la antimicina A, inhibidor de la transferencia de electrones entre el pool de las quinonas y el complejo III es igualmente efectiva. A priori, el estudio de las hipoxias histotxicas posee muy poco inters fisiolgico, pero el hecho de que los venenos de la cadena respiratoria sean potentes estmulos de los quimiorreceptores del CC, ha hecho que se hayan utilizado mucho para explorar los reflejos (16) y los mecanismos bsicos de funcionamiento del CC (1719), por ello los describiremos brevemente. In vivo el cianuro produce un aumento de la frecuencia de descarga en el NSC y un aumento en la ventilacin de forma dosis dependiente, que remeda las producidas por la hipoxia hipxica. Tambin provoca respuestas cardiovasculares reflejas, con aumento de la frecuencia cardiaca y aumento de la presin arterial (20). Otros venenos metablicos (rotenona, azida) tambin estimulan los quimiorreceptores arteriales del CC, aumentando las descargas en el NSC y aumentando la liberacin de dopamina, lo que produce un aumento de la ventilacin (20,21). Estas observa-
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
43 ciones llevan a suponer a las mitocondrias como posibles sensores de la hipoxia, aunque hay posturas encontradas al respecto (18,19). 3.3. Funcin del cuerpo carotdeo en hipercapnia y acidosis La hipercapnia, esto es, una PCO2 por encima de 40 mmHg, produce cambios en la frecuencia de impulsos quimiosensoriales del NSC de gatos. La relacin de las frecuencias de descargas del NSC con las presiones alveolares de CO2 es lineal en el rango de PCO2 entre 20-80 mmHg, aunque a PCO2 ms extremas la relacin tiende a horizontalizarse, adquiriendo la curva una forma sigmoidea. La pendiente de las curvas aumenta a medida que disminuye la PO2 a la que se realizan los experimentos, lo que define una interaccin positiva entre el estmulo hipercpnico e hipxico tal como se pone de manifiesto en la Figura 5. La relacin entre PCO2 y ventilacin es comparable a la de las descargas, pero en la gnesis de la respuesta ventilatoria intervienen, no slo los CC, sino tambin los quimiorreceptores centrales. Haciendo estimulacin selectiva del CC (perfundiendo los CC con sangre hipercpnica y el cerebro con sangre normocpnica) o denervando los CC se puede determinar la contribucin de cada uno de los quimiorreceptores a la respuesta ventilatoria. En experimentos de este tipo realizados por Berkenbosch et al., (22) en gato, se observ que en las respuestas agudas la contribucin del CC a la hiperventilacin producida es prxima al 50%. Curiosamente, la hiperventilacin producida por la hipercapnia es menos diafragmtica que la hiperventilacin hipxica, segn se pone de manifiesto registrando actividad elctrica en el nervio frnico o actividad electromiogrfica en el diafragma a niveles de ventilacin comparables (5).
Impulsos/seg
PaCO2, mmHg Figura 5. Respuesta del NSC de gato a la PCO2 a diferentes PO2. Obsrvese la baja intensidad de la respuesta a la PCO2 en condiciones de hiperoxia. La ventilacin sigue una relacin comparable.
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
44 Un aspecto muy debatido en la literatura de la quimiorrecepcin, es la naturaleza del estmulo: es el CO2 per se el estmulo, o por el contrario, es el aumento de concentracin intracelular de H+ que el CO2 genera, el estmulo eficaz? Los datos ms recientes avalan la segunda alternativa ya que: 1) Una solucin con PCO2 alta o que contenga cidos dbiles no voltiles a pH 7.4 activa la liberacin de dopamina, las descargas en el nervio del seno y producen acidificacin intracelular; 2) El aumento de la PCO2 en el medio de perfusin de las CQ del CC, aumenta de forma brusca la concentracin intracelular de H+, y paulatinamente dicha concentracin intracelular de H+ disminuye hasta un nivel superior al normal, pero menor que el pico; las descargas en el NSC siguen un patrn idntico; 3) Si las CQ son tratadas con un inhibidor de anhidrasa carbnica antes de aumentar la presin de CO2, el proceso de acidificacin intracelular pierde el componente inicial rpido, y la liberacin de dopamina de las CQ y las descargas del NSC tambin lo pierden; 4) Si el CO2 por si mismo estimulara los quimiorreceptores, entonces los inhibidores de anhidrasa carbnica o no afectaran o aumentaran la respuesta. Adems los cidos dbiles no voltiles (propionato, butirato) no seran eficaces. Los efectos de la hipoxia y la hipercapnia sobre el aparato cardiovascular son similares. Esto no es sorprendente si tenemos en cuenta que ambos estmulos son vasodilatadores, activan el reflejo quimiorreceptor produciendo hiperventilacin y como consecuencia activan el reflejo de distensin pulmonar y sus efectos hemodinmicos. Tal vez la hipercapnia produzca mayor activacin del simptico y mayor secrecin de CA que la hipoxia, lo que contrarrestara su mayor poder vasodilatador. La inyeccin intravenosa de un cido diluido, aumenta la frecuencia de impulsos en el NSC, existiendo una relacin lineal entre la concentracin de H+ en sangre o lquido de superfusin y la frecuencia de descargas (23). La hiperventilacin que sigue a la inyeccin de cido corre paralela a las descargas, pero en el caso de las acidosis metablicas la contribucin del CC a la hiperventilacin es mayor que en el caso de las acidosis respiratorias. La mayor contribucin de los CC a la hiperventilacin provocada por la acidosis metablica (especialmente en los perodos iniciales) que por las acidosis respiratorias sera debido a la lenta difusin de los H+ al lquido cfalo-raqudeo (24). Los H+ difunden lentamente al lquido cfalo-raqudeo y la hiperventilacin promovida por los CC disminuye la PaCO2, forzando al CO2 a difundir rpidamente del lquido cfalo-raqudeo a la sangre, generndose normohidria o an alcalosis del lquido cfalo-raqudeo, lo que reduce la estimulacin de los quimiorreceptores centrales. Sin embargo, si la acidosis se prolonga, el pH del lquido cfalo-raqudeo tiende a equilibrarse con el de la sangre, con lo que se estimularn tanto los quimiorreceptores centrales como los perifricos (25). 3.4. Funciones del cuerpo carotdeo en el ejercicio Durante el ejercicio, la ventilacin sigue un curso temporal que permite dividirlo en varias fases tal como muestra la Figura 6A. En condiciones estables y para ejercicio de intensidades por debajo del umbral anaerbico, la ventilacin aumenta linealmente en proporcin directa a la intensidad del propio ejercicio, del consumo de O2 y de la produccin de CO2, pese a que la PaO2, la PaCO2 y el pHa se mantienen dentro de los rangos normales; por encima del umbral anaerbico, la ventilacin aumenta en forma desproporcionada a la intensi-
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
45 dad del ejercicio y se acompaa de una acidosis metablica, con disminucin del pHa; la acidosis metablica y la hiperventilacin adicional resultante producen hipocapnia y tendencia a aumentar la PaO2. En la Fase 0 del ejercicio, el aumento de ventilacin es instantneo y generado a nivel del sistema nervioso central. Las rdenes motoras que descienden hasta las motoneuronas de los msculos implicados en el ejercicio, envan por colaterales informacin a los centros troncoencaflicos de control respiratorio y cardiovascular, generando hiperventilacin y aumento del gasto cardiaco. Hay tambin un componente en estos ajustes iniciales que se origina en el hipotlamo, como se ha podido demostrar en distintas preparaciones de animales descerebrados, talmicos y decorticados. En la Fase 1 del ejercicio la ventilacin contina aumentando durante unos minutos a pesar de que la intensidad del ejercicio se mantenga. En esta segunda fase la hiperventilacin se genera por rdenes centrales, por aferencias de los propios msculos y por la activacin de los quimiorreceptores del CC. La Fase 2 de estado corresponde a una hiperventilacin sostenida que es proporcional al consumo de O2; los mecanismos involucrados son los mismos. En estas dos fases sabemos que el CC participa, ya que en sujetos humanos glomectomizados o en animales con los CC denervados, la pendiente de ascenso (Fase 1) y la hiperventilacin de la Fase de estado (Fase 2) son menores (ver Figura 6B). Constatada la participacin del CC en la hiperventilacin del ejercicio moderado y dado que la PaO2, la PaCO2 y el pHa se mantienen dentro de rangos normales cabe preguntarse cules son los mecanismos de activacin de los quimiorreceptores del CC? Parece ser que la hiperventilacin mediada por el CC se debera a que durante el ejercicio hay un aumento de la amplitud y frecuencia de las oscilaciones de los gases sanguneos en razn del aumento de las diferencias arteriovenosas tanto para el O2 como para el CO2. Adems, la liberacin de K+ en los msculos activos produce un aumento significativo de la potasemia, capaz de generar un aumento de la frecuencia media de descarga del NSC y de la amplitud de su oscilacin respiratoria. El aumento de la adrenalina plasmtica que produce el ejercicio tambin podra contribuir a la activacin del CC ya que el bloqueo -adrenrgico por propra-
Ventilacin/minuto (l)
Ventilacin/minuto (l)
0 0
10
20
30
40
25
50
75
Figura 6. A, Curso temporal de la ventilacin durante el ejercicio fsico (ver texto; modificada de ref. 4). B, respuesta ventilatoria al ejercicio fsico en sujetos controles (C), y en sujetos glomectomizados bilateralmente (G). Obsrvese que la respuesta ventilatoria es menor en sujetos glomectomizados que en controles a todas las intensidades de ejercicio fsico. Modificada de Honda (26).
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
46 nolol o atenolol reduce la descarga quimiosensorial del NSC. Finalmente, dada la alta sensibilidad del CC a la temperatura (27), es concebible que los aumentos de temperatura que ocurren durante el ejercicio contribuyan a aumentar la sensibilidad del CC; en el mismo sentido actuara la hipertona plasmtica en ejercicio prolongado cuando hay sudoracin profusa (ver ms adelante). En cuanto al ejercicio intenso, el esquema general sera que la acidosis metablica del ejercicio intenso junto con la hiperpotasemia creciente, el aumento de catecolaminas y el aumento de temperatura corporal, interaccionaran positivamente a nivel CC para promover la hiperventilacin aadida que se observa en situaciones de ejercicio intenso (superior al 70% del ejercicio mximo). 3.5. Otras funciones del cuerpo carotdeo En el gato se aprecia un aumento de la frecuencia de descargas quimiosensoriales del NSC si calentamos la sangre cercana a la bifurcacin carotidea y disminuye la frecuencia al enfriarse la sangre. Preparaciones in vitro, en las que se modifica la temperatura del bao manteniendo inalterables otras variables han permitido confirmar con precisin la termosensibilidad del CC. En gato anestesiado, en el que se aumenta la temperatura corporal de 35 a 40 C por calentamiento externo, se ve un aumento progresivo de la frecuencia de descarga del NSC. Este efecto es ms fcil de observarse si se secciona el NSC contralateral al del estudio, ya que de lo contrario el aumento de la ventilacin alveolar produce disminucin de la PCO2 que tiende a frenar la actividad del NSC. La termosensibilidad del CC podra participar en la hiperventilacin durante el ejercicio y cuadros febriles. Al aumentar la osmolaridad de la solucin que perfunde el CC, tanto en preparaciones in vivo como in vitro, aumenta la frecuencia de descarga del NSC. La respuesta de los CC a cambios de la osmolaridad, parecen contribuir al control de la osmolaridad de los lquidos corporales ya que al inyectar intracarotdeamente una solucin hiperosmtica en perro, se observa una disminucin del flujo urinario. Adems al perfundir con una solucin hipxica el CC o al estimular de forma elctrica el NSC se ha observado liberacin refleja de la ADH y al inyectar una solucin hipercpnica intracarotidea o una inyeccin intravenosa de NaCN, se ha observado un aumento de la frecuencia de descarga de las neuronas del ncleo supraptico del hipotlamo, siempre que los NSC estn intactos. 3.6. Control eferente de la funcin del cuerpo carotdeo Los nervios ganglio-glomerulares (ver Figura 1), que contienen fibras simpticas postganglinicas, y por tanto noradrenrgicas, terminan en la pared del seno carotdeo, la musculatura lisa de los vasos finos del CC y axones que siguen ascendiendo por el NSC; tambin hay algunas fibras simpticas postganglinicas que ascienden hasta el glosofarngeo y luego descienden por el NSC hasta el CC. La estimulacin elctrica de los nervios ganglio-glomerulares produce vasoconstriccin del CC y aumento de descargas del NSC. Una segunda va de inervacin eferente del CC estara constituida por fibras que viajan por el NSC hasta el CC y cuyo efecto sobre la actividad quimiosensorial sera inhibitorio. Experimentos recientes del laboratorio de Fidone muestran que habra dos tipos de fibras,
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
47 ambas nitrrgicas, que median la inhibicin eferente. Uno estara constituido por fibras postganglionicas parasimpticas, cuyos somas estn dispersos a lo largo del NSC (28) y que producen vasodilatacin dentro del CC, donde participaran mecanismos nitrrgicos y muscarnicos. El otro tipo de fibras liberara NO, y produciria inhibicin directa de clulas glmicas quimiorreceptoras, va mecanismos dependientes e independientes del GMP cclico. Recientemente Campanucci et al. (29) han ratificado que las neuronas localizadas en la trayectoria del NSC/glosofarngeo son nitrrgicas y colinrgicas. Adems, han demostrado que poseen corrientes leaky de K+ sensibles a hipoxia y a anestsicos voltiles e insensibles a los bloqueantes clsicos de canales de K+, esto es, son corrientes anlogas a las expresadas por las CQ (ver ms adelante). La hipoxia que activara al CC merced a la inhibicin de dichas corrientes en las CQ, tambin inhibira estas corrientes en las neuronas dispersas en el NSC activndolas e iniciando de esta forma una inhibicin de la actividad quimiorreceptora del CC va la liberacin de ACh y NO en sus terminaciones intraglmicas. Adems, en el NSC hay fibras sensoriales dopaminrgicas con sus somas en el ganglio petroso, que liberan DA al activarse. Esta DA actuara sobre receptores dopaminrgicos D2 de las clulas glmicas e inhibira su actividad quimiorreceptora. Se ha demostrado liberacin de dopamina en el CC al estimular el NSC a intensidades suficientes para reclutar las fibras C.
4. FISIOLOGA GENERAL DEL CC. MECANISMOS DE FUNCIONAMIENTO DE LAS CQ A NIVEL CELULAR Y MOLECULAR 4.1. Deteccin y transduccin de los estmulos a) La hipoxia Los mecanismos de funcionamiento del CC a nivel celular han constituido un enigma durante muchos aos, por lo que han surgido multitud de hiptesis con nombres tales como hiptesis metablica, hiptesis cida e hiptesis dopaminrgica, etc. (30). A partir de los aos ochenta tras demostrar Gonzalez and Fidone (31) y Fidone et al. (32) que las CQ liberaban DA, nuestro laboratorio comenz a considerar la transduccin sensorial como un caso especial de acoplamiento estmulo secrecin. Los estudios iniciales fueron encaminados a demostrar la universalidad de la observacin, constatando que las CQ del CC de todas las especies liberaban DA en respuesta a la hipoxia e hipercapnia en proporcin directa a la intensidad del estimulo y que esta liberacin es dependiente de Ca2+. Tambin demostramos que el alto K+ extracelular induce liberacin de dopamina dependiente de Ca2+ y que la liberacin inducida por alto K+ y la hipoxia es sensible a dihidropiridinas (antagonistas / agonistas de canales de Ca2+). Demostramos igualmente la presencia de canales de Na+ dependientes de voltaje en las CQ y que la tetrodotoxina inhiba parcialmente la liberacin de DA inducida por la hipoxia. Todos estos hechos evidenciaban que la hipoxia deba despolarizar las CQ. Tras poner a punto una preparacin de CQ en cultivo primario, iniciamos un estudio con tcnicas de patch-clamp que llev a descubrir la existencia de corrientes de K+ sensibles a la PO2, que se inhiban en hipoxia y a verificar la existencia de canales de Na+ y Ca2+ dependientes de voltaje.
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
48 Del conjunto de estas observaciones naci el modelo de transduccin del estmulo hipxico que se conoce con el nombre de modelo de membrana de la transduccin hipxica (33) que presentamos en forma de diagrama de flujo en la Figura 7 (34). En esta figura, que es autoexplicativa, se subrayan aquellas etapas de la cascada de transduccin que no estn satisfactoriamente resueltas experimentalmente, y que son las que discutiremos aqu.
Figura 7. Cascada de transduccin hipxica en CQCC. Tomada de Gonzalez et al (34).
Sensor(es) de O2. No conocemos la identidad molecular del sensor o los sensores de O2, por lo que existen varias teoras al respecto. Hasta muy recientemente haba tres teoras en boga. En 1992 nosotros propusimos que el sensor sera una hemoprotena, ya que el CO era capaz de prevenir la inhibicin de las corrientes de K+ producidas por la hipoxia, o en otras palabras, el CO era capaz de sustituir al O2 a nivel del sensor de O2. En experimentos recientes (35) hemos observado en parches de membrana aislados en configuracin inside out que la hipoxia disminua la probabilidad de apertura de canales de K+ y que el CO reverta el efecto. Dado que el efecto del CO se manifiesta en parches aislados y que el CO no es sustrato de ningn enzima conocido en clulas de animales superiores, nos reafirmamos en la conclusin previa de que el sensor sera una hemoprotena de membrana que al desaturarse de O2 cambia de conformacin y transmite alostricamente el cambio a los canales de K+ para modificar su cintica de apertura. El CO al unirse al sitio del O2 revertira la configuracin del sensor a una situacin comparable a la que tiene a PO2 normales y el canal se abrira. Desafortunadamente, la hemoprotena propuesta como sensor no ha sido identificada molecularmente. La segunda teora propona que el sensor sera una NADPH oxidasa anloga a la de las clulas fagocitarias, que reduce el O2 molecular con un solo electrn para formar O2- que se dismuta a H2O2. La propuesta est basada en la observacin de que un inhibidor de la NADPH oxidasa, el difenileneiodonio (DPI), activa las descargas del NSC y previene el efecto de la hipoxia. La propuesta era que la disminucin de la PO2 inhibira al enzima disminuyendo la produccin de O2- y de H2O2; esta disminucin de O2- y de H2O2 cambiara el potencial redox de la clula, esto es el cociente glutation reducido vs. glutation oxidado (GSH/GSSG) hacia un valor ms reducido, ms alto, y ello generara un cambio en la relacin entre grupos sulfhidrilos libres y
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
49 puentes disulfuro (Prot -SH/Prot-S-S) de los canales de K+ con la consiguiente modificacin de su cintica de apertura (36). Otro grupo de autores (37) encontraron que la hipoxia aumentaba la produccin de ROS (de O2- y de H2O2) a nivel mitocondrial y propusieron que el sensor sera la mitocondria y que la seal capaz de modificar la cintica de los canales de K+ sera el aumento de ROS, y no su disminucin como propona Acker (36). El tercer modelo propone que las mitocondrias seran los sensores de la hipoxia e iniciadores de la cascada de transduccin. Esta hiptesis es muy antigua en la literatura (38) y se conoca con el nombre de hiptesis metablica. Dado que los venenos metablicos, mitocondriales y glicolticos son fuertes activadores de la actividad quimiorreceptora, la hiptesis metablica propona que la hipoxia, al igual que los venenos metablicos, estimulara al CC a travs de una disminucin de los niveles de ATP. Ya en 1968 la hiptesis fue seriamente criticada sobre la base de que a las PO2 que el CC responde, la cadena respiratoria no estara inhibida en absoluto por dficit de O2, y entonces los niveles de ATP no disminuiran. Estos extremos fueron verificados experimentalmente por nuestro laboratorio al demostrar que los desacoplantes, por un lado, y la hipoxia, por otro, eran capaces de activar la liberacin de DA y la actividad en el NSC sin modificar los niveles de ATP. A pesar de estos hechos la literatura est salpicada de artculos en los que es recurrente la idea de las mitocondrias como sensores de la PO2, pero con puntos de vista encontrados y sin datos experimentales slidos (18,19). Recientemente Peers y colaboradores, (39) han encontrado que en normoxia la hemoxigenasa-2 genera CO que activa al maxi-K, un canal de K+ sensible a la hipoxia en las CQ de rata. En hipoxia la actividad de la enzima disminuye, debido a la disminucin de la PO2, por lo que el maxi-K tiende a inhibirse. Esta inhibicin ms la producida por la propia hipoxia conducira a la inhibicin global del maxi-K que se observa en experimentos en clula intacta o en whole-cell. Mecanismo(s) de acoplamiento. El desconocimiento de la naturaleza del sensor de O2 hace difcil buscar el mecanismo de acoplamiento entre dicho sensor y los canales de K+ sensibles a O2. En los dos modelos alternativos, en los que se atribuye un papel principal a los ROS, el mecanismo de acoplamiento es sencillo de entender: cambio en la relacin entre grupos sulfhidrilos libres y puentes disulfuro (Prot-SH/Prot-S-S) de los canales de K+ que promoveran los cambios cinticos antes descritos. Sin embargo parece muy poco probable que este mecanismo tenga un significado fisiolgico ya que el CO, que desde luego es incapaz de reducir u oxidar ninguna molcula en condiciones normales de temperatura y presin, revierte el efecto de la falta de O2 (PCO ~70 a 140 mmHg y PO2 ~30 mmHg). De otro lado, se ha demostrado que la reduccin de canales inicos y la sensibilidad al O2 son cosas distintas. As por ejemplo en clulas HEK transfectadas con canales de K+ de la familia shaker, shaker/1.2 y Kv4.2/1.2, el ditiotreitol disminuy la corriente de los tres sistemas y la hipoxia slo inhibi al ltimo de ellos (40). Adems, Riesco-Fagundo et al. (35) demostraron que el ditiotreitol aumenta la probabilidad de apertura del BK y la hipoxia lo disminuye. As pues el estado redox de los canales no parece estar involucrado en la iniciacin de la cascada de transduccin, lo que no excluye que pueda jugar un papel modulador de la misma. Ya hemos mencionado que si el sensor es realmente una hemoprotena, el mecanismo de acoplamiento sera alostrico: la reorganizacin intramolecular del sensor al desaturarse por
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
50 la disminucin del O2 producira cambios en la disposicin espacial del mismo (tal como ocurre en la hemoglobina al desaturarse) que conduciran al cierre o que dificultaran la apertura de los canales de K+ sensibles a la PO2. Si las mitocondrias (o alguno de sus componentes moleculares) fueran los sensores no sabemos qu seal produciran para modificar la cintica de los canales de K+. Sin embargo, sea cual fuere el mecanismo, parece que las subunidades reguladoras de los canales de K+ seran necesarias para el acoplamiento entre el mecanismo de deteccin o sensor(s) de O2 y la subunidad conductora de los canales de K+. En efecto, en sistemas de transfeccin las corrientes de K+ slo son sensibles a la hipoxia si se cotransfectan las subunidades con subunidades reguladoras o si la transfeccin se hace a tipos celulares que expresan dichas subunidades . En otras palabras, las subunidades interaccionaran con el presunto sensor hemoproteico de O2 o seran los componentes de los canales de K+ que reaccionan con los ROS (o el glutation) o con la seal originada en las mitocondrias. Esta misma idea es apoyada por datos recientes de Hartness et al. (41), que encuentran que la hipoxia crnica, que se acompaa de sensibilizacin de las respuestas a la hipoxia, produce un aumento de expresin de subunidades y de su colocalizacin con las subunidades de los canales de K+ sensibles a hipoxia. Sin embargo, en un estudio muy reciente (42) se ha demostrado que una variante de la subunidad conductora del maxi-K+ generada por splicing alternativo posee sensibilidad intrnseca a la hipoxia sin la necesidad de cooperacin de las subunidades . Habremos de esperar unos pocos meses (aos) a ver qu variante es la que se expresa en las CQ. Identidad molecular del canal de K+ sensible a la PO2. Este aspecto es de gran importancia para entender la transduccin hipxica. El primer canal de K+ sensible a la PO2, descrito por nosotros en clulas quimiorreceptoras de conejo, es voltaje dependiente y tiene un umbral macroscpico de activacin en torno a -40 mV (43-45), generando una corriente que se inactiva con el tiempo; dado que el potencial de membrana (Em) de las CQ oscila entre -50 y -60 mV, el dilema era evidente: si los canales de K+ sensibles a O2 estn cerrados, en condiciones de normoxia (Em = -50, -60 mV), la hipoxia no podra disminuir su probabilidad de apertura. La situacin era similar en CQ de rata en las que el primer canal sensible a la PO2 descrito fue el BK, canal de K+ dependiente de Ca2+; en reposo (en normoxia) la [Ca2+]i es bajo, en el rango nanomolar y el Em negativo por lo que el BK estara igualmente cerrado y la disminucin de la PO2 tampoco podra disminuir la Po del BK. En otras palabras, ninguno de los dos canales descritos con sensibilidad a la hipoxia pareca, en trminos macroscpicos, contribuir al mantenimiento del Em en reposo (normoxia) y por tanto no podan alterarlo. Los argumentos que se han utilizado para defender el modelo de membrana (ver Figura 7) son de doble naturaleza: o bien que la probabilidad de apertura de los canales de K+ al Em normxico fuera, aunque muy baja, distinta de cero, o bien que las clulas presentaran actividad elctrica espontnea. En ambos casos, la inhibicin de los canales de K+ por la hipoxia s que podra regular la cintica de activacin de los canales dependientes de voltaje y de los dependientes de Ca2+, y por tanto la entrada de Ca2+. Dos hallazgos recientes parecen resolver esta problemtica. El primer hallazgo se realiz en clulas quimiorreceptoras de rata. En esta especie, varios autores haban encontrado que el TEA (hasta 20 mM) y la caribdotoxina (20 nM), solos o en combinacin con hasta
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
51 5 mM de 4-aminopiridina, no modificaban en condiciones de normoxia ni el Em, ni los niveles de Ca2+ de las clulas quimiorreceptoras aisladas, ni la liberacin de DA, ni las descargas en el NSC en CC intactos; tampoco afectaban la liberacin de DA o la actividad en el NSC evocada por la hipoxia. Las conclusiones de estos estudios eran obvias: los canales de K+ dependientes de Ca2+ (BK) no tienen nada que ver con la respuesta de las clulas quimiorreceptoras a la hipoxia. Sin embargo, Buckler (46) hizo un hallazgo adicional: las clulas quimiorreceptoras de rata poseen una corriente leaky, voltaje independiente que es inhibida por la hipoxia de forma reversible. Posteriormente identific al canal que lleva dicha corriente como el TASK-1, aunque tambin podran contribuir otros canales de la misma familia. Es importante sealar que la sensibilidad a la hipoxia de estos canales leaky, evidente en registros en cell-attached, se pierde al escindir los parches de membrana y pasar a registrar en configuracin inside-out. En suma, los hallazgos de Buckler han resuelto un problema, pero han arrojado sombras sobre otros aspectos que parecan resueltos, tal como la sensibilidad intrnseca de la membrana celular a la hipoxia (ver ms arriba). El segundo hallazgo se realiz en clulas quimiorreceptoras de conejo. En clulas quimiorreceptoras de conejo con dominantes negativos para canales de la familia shaker (Kv1) o shal (Kv4) obtenidas por transferencia adenoviral de los correspondientes genes mutados, se ha encontrado en nuestro laboratorio que slo las clulas nulas para la familia Kv4 estaban despolarizadas en condiciones de normoxia y no se despolarizaban ni por la hipoxia ni por las 4-aminopiridinas, indicando que estos canales (y no los Kv1) contribuyen al mantenimiento del Em y son responsables de la despolarizacin producida por la hipoxia. En experimentos ms recientes utilizando una combinacin de tcnicas electrofisiolgicas, microfluorometra con imagen para Ca2+, RT-PCR, inmunocitoqumica e hibridacin in situ, se han confirmado las observaciones previas, al verificarse la expresin de subunidades Kv4.1 y Kv4.3 en todas las clulas quimiorreceptoras, TH-positivas. Adems, se encontr que un nmero importante de clulas, sino todas, co-expresan subunidades Kv3.4, que tambin sustentan corrientes transitorias. Datos muy recientes de nuestro laboratorio indican, que en efecto, ninguno de los inhibidores del BK (TEA e Iberiotoxina) afectan ni a la liberacin basal de DA ni a la inducida por la hipoxia en el CC intacto, ni a los niveles basales de Ca2+ ni a los aumentos de Ca2+ inducidos por la hipoxia en clulas aisladas. Pero, cmo explicar que la inhibicin farmacolgica de los canales de K+, en concreto el BK, cuyo funcin principal es repolarizar las clulas no afecta la respuesta a la hipoxia, cuando sabemos que las clulas generan potenciales de accin? La nica posibilidad que se nos ocurre es que los bloqueantes no afectan la respuesta a la hipoxia porque los canales sensibles estn total o casi totalmente inhibidos por la hipoxia a pesar de que en los registros en whole-cell las inhibiciones encontradas nunca superen el 40-50%. Estas consideraciones llevaron a buscar un estmulo que activara las clulas sin inhibir al BK. Se eligi dinitrofenol (DNP) porque no se conoce que tenga efecto sobre las corrientes de K+ dependientes de voltaje. Pues bien, el TEA y la iberiotoxina potencian la liberacin de DA y el aumento de Ca2+ intracelular producido por DNP en CC intacto. En experimentos muy recientes (Gomez-Nio, Obeso, Lopez-Lopez & Gonzalez), han confirmado la premisa de los experimentos descritos, al demostrar que en efecto el DNP no inhibe el BK en experimentos de patch-clamp en parches aislados.
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
52 En suma, pensamos que el modelo de membrana propuesto en la Figura 7 se ajusta, aunque con alguna laguna, a las observaciones experimentales macroscpicas y es capaz de acomodar un nmero muy importante de las observaciones existentes a nivel celular y molecular. b) Estmulos cido-hipercpnicos Ya hemos descrito que el estmulo eficaz para las clulas quimiorreceptoras es el aumento intracelular de la [H+]. Cmo un aumento de la [H+]i es capaz de activar las clulas? Nuestro laboratorio propuso un modelo de transduccin del estmulo cido en el CC de conejo en el que el aumento de la [H+] intracelulares activara el intercambiador Na+/H+ con el consiguiente aumento de Na+ intracelular; a su vez este aumento de Na+ intracelular activara el intercambiador Na+/Ca2+ que funcionando en modo reverso introducira Ca2+ en el interior de las clulas, promoviendo la liberacin de neurotransmisores (47). Trabajando en clulas quimiorreceptoras de CC de rata, Buckler and Vaughan-Jones (48) encuentran que los estmulos cidos, al igual que la hipoxia, despolarizan las clulas y generan potenciales de accin, que deben ser potenciales de Ca2+, porque las clulas quimiorreceptores, de rata parece que no poseen canales de Na+. La amplitud de la despolarizacin y de las rfagas de potenciales de accin, as como los aumentos de [Ca2+]i producidos por los estmulos cidos en las clulas quimiorreceptoras son comparables a las producidas por la hipoxia (49). De otro lado, el maxi-K+ es muy sensible a la acidificacin intracelular y el TASK-1 es sensible al pH extracelular. Estudios muy recientes de nuestro laboratorio (50) realizados en clulas quimiorreceptoras de conejo han confirmado que los estmulos cidos despolarizan las clulas tambin en esta especie animal, promoviendo la activacin de canales de Ca2+ voltaje dependientes y exocitosis de neurotransmisores. En suma, la transduccin del estmulo cido tiene las mismas luces y sombras que la del estmulo hipxico. 4.2. Neurotransmisin en el cuerpo carotdeo La idea de neurotransmisin entre las clulas quimiorreceptoras y las terminaciones nerviosas del NSC fue avanzada por De Castro (1) al definir el CC como un rgano sensorial. El nico neurotransmisor considerado hasta finales de 1950 fue la ACh. Fueron muchos los autores (38) que demostraron que la ACh y agentes colinestersicos eran excitatorios para la actividad en el NSC. Eyzaguirre y colaboradores en una serie de artculos compilados en Eyzaguirre y Zapata (51) proporcionaron un fuerte empuje a la llamada teora colinrgica al demostrar entre otras cosas que el CC de gato contena ACh y que sta se liberaba en respuesta a la estimulacin hipxica. Sin embargo los estudios farmacolgicos no consiguieron demostrar que la inhibicin de la transmisin colinrgica con bloqueantes nicotnicos y muscarnicos suprimiera la actividad quimiorreceptora generada por los estmulos naturales. Finalmente, en estudios muy recientes, Gauda et al. (52) han demostrado que las clulas quimiorreceptoras no expresan colinacetiltransferasa, que se localiza exclusivamente en clulas ganglionares intraglmicas, por lo que no puede ser el neurotransmisor entre CQ y terminaciones sensoriales al no encontrarse en el elemento presinptico.
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
53 En el ao 1957, Lever y Boyd (53) demostraban la presencia en las clulas quimiorreceptoras de vesculas de ncleo denso comparables a los de las clulas cromafines de la mdula adrenal, y proponan que tal vez se debiera a que las CA fueran los neurotransmisores. A este artculo pionero siguieron multitud de estudios nuestros que caracterizaron los enzimas involucrados en la sntesis de CA, el proceso de sntesis de CA en normoxia e hipoxia y la liberacin de CA en respuesta a la hipoxia (21,30). La problemtica con las CA como posibles neurotransmisores en la unin clula glmica-terminaciones sensoriales del NSC es que las CA exgenas no siempre mimetizan los efectos de los estmulos naturales: en vez de estimular como la hipoxia, con frecuencia inhiben las descargas en el NSC o la ventilacin y los bloqueantes catecolaminrgicos en vez de inhibir la actividad quimiorreceptora con frecuencia la aumentan. Adems de la presencia de serotonina en el CC, demostrada por Chiocchio et al. (54), y cuyo significado funcional es dudoso, existe evidencia bioqumica e inmunocitoqumica de la existencia de un nmero importante de neuropptidos. Aunque existen observaciones experimentales respecto a acciones concretas de uno u otro pptido, el significado funcional general de los neuropptidos en el CC, ya sea como moduladores de la funcin de las clulas glmicas o de la transmisin sinptica o como factores trficos a largo plazo, no est bien definida (55). Cabe destacar a la sustancia P por la gran atencin que ha recibido. La ausencia de mRNA que codifique para este neuropptido en las clulas quimiorreceptoras le excluye como neurotransmisor primario, pero muy bien pudiera ser que la liberacin de sustancia P de las terminaciones sensoriales, donde es muy abundante, fuera capaz de modular la actividad quimiorreceptora. Los pptidos opioides, presentes en las clulas quimiorreceptoras, son inhibitorios tanto de la actividad en el NSC como de la ventilacin (20,21), y al menos parte de sus acciones las ejercen va receptores presinpticos ya que son capaces de inhibir la respuesta secretora de las clulas quimiorreceptoras frente gran variedad de estmulos. Recientemente Nurse y cols han desarrollado una preparacin de co-cultivo de clulas quimiorreceptoras y de neuronas del ganglio petroso que forman sinapsis funcionales, de tal forma que registrando en las neuronas tras formar las sinapsis, y no antes, es posible observar actividad espontnea y un aumento de descargas en respuesta a la hipoxia que es bloqueado por una mezcla de bloqueantes nicotnicos (mecamilamina) y purinrgicos (suramina), concluyendo que la transmisin en la sinapsis clula quimiorreceptora-terminacin sensorial, al menos en el medio artificial que supone el cultivo, es una cotransmisin nicotnica y purinrgica. Los receptores purinrgicos responsables de la cotransmisin en el CC seran receptores P2X2 y P2X3 (ya sea como homo o como heterorreceptores), cuya expresin se solapa muy frecuentemente en las mismas neuronas del ganglio petroso y en las terminaciones nerviosas del NSC a nivel intraglmico. Ya hemos discutido ms arriba que las clulas quimiorreceptoras no expresan colinacetiltransferasa in vivo, por lo que el rasgo colinrgico sera una adquisicin del medio de cultivo (52). Tampoco el papel del ATP est libre de sospecha. Los trabajos clsicos de farmacologa no apoyan al ATP como neurotransmisor (56,57). Los primeros autores encuentran que mientras que el ATP y la adenosina aumentan la actividad en el NSC del gato, con curvas dosis respuestas superponibles, los anlogos no hidrolizables de ATP la disminuyen, concluyendo que la estimulacin producida por el ATP estara mediada por la adenosina liberada tras la hidrlisis del nucletido. Por su parte Spergel y Lahiri (57) encuentran que la infusin sostenida de ATP induce una acti-
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
54 vacin de las descargas en el NSC seguida por una desensibilizacin y desaparicin de la respuesta, mientras que la misma maniobra no afect a la respuesta inducida por la hipoxia: si el ATP fuera un mediador importante de la respuesta hipxica, sta debera ser atenuada por la infusin previa de ATP. Sin embargo, en estudios muy recientes, Rong et al. (58) y Spyer et al. (59) encuentran que ratones knockout para receptores purinrgicos del tipo P2X2 son incapaces de responder a la hipoxia, mientras que responden a la hipercapnia (la respuesta es mediada en ms de un 50% por los quimiorreceptores centrales); adems, registros in vitro de la actividad en el NSC demuestran que los CC son incapaces de generar actividad elctrica frente a la hipoxia en las fibras del NSC. La conclusin sera que el ATP es el neurotransmisor principal en esta sinapsis sensorial y que el resto de neurotransmisores son co-transmisores secundarios con capacidad para modular la actividad tanto a nivel presinptico como postsinptico. Sin embargo, experimentos muy recientes del laboratorio de Fidone realizados en rata (60) dan al ATP una responsabilidad de en torno al 50% de la actividad registrada en el NSC en respuesta a la hipoxia. En este sentido nuestro laboratorio ha encontrado que en la rata la adenosina sera el responsable del otro 50% de la actividad, ya que inhibidores no especficos de receptores de adenosina, como la cafena, disminuyen en ese porcentaje la actividad elctrica generada por la hipoxia. Una diseccin farmacolgica del efecto de la cafena indica que sta acta en receptores del subtipo A2A y A2B; poniendo las cosas en forma directa diramos que la adenosina liberada endgenamente es responsable del 50% de la actividad generada por la hipoxia (61). Tras esta visin sucinta vamos a referirnos a un aspecto de la neurotransmisin que consideramos de gran inters. La liberacin de DA y NE ha sido sugerida por mltiples estudios in vivo, al demostrar una disminucin de DA y/o NE en el CC asociada a un aumento del turnover de las mismas tras un episodio agudo de hipoxia. Sin embargo, han sido los laboratorios de Fidone y el nuestro propio los que han caracterizado ms en detalle, la liberacin de CA en relacin con multitud de estmulos, utilizando mtodos radioisotpicos y voltamtricos. La primera medida directa de liberacin de CA por la hipoxia fue obtenida en CC de conejo por Gonzalez y Fidone (31) y en una serie de estudios posteriores se cuantific la relacin entre la intensidad de la hipoxia, la actividad en el NSC y la cantidad de CA liberada en la misma especie, demostrando adems que la CA liberada era DA. Los estudios se hicieron extensivos a otras especies tales como el gato, la rata, la ternera y el ratn. Ha habido otros laboratorios que han comprobado igualmente la liberacin en rata, en conejo y en gato. La liberacin de DA frente a los estmulos cidos ha sido caracterizada por nosotros tambin, y determinamos la relacin entre intensidad de estmulo, actividad en el NSC y la cantidad de CA liberada. Para estmulos farmacolgicos tambin hemos establecido esa relacin entre concentracin del agente estimulante, liberacin de DA y aumento de actividad en el NSC tanto en gato como en conejo (55). Tambin hicimos la primera demostracin cuantitativa de liberacin de DA en clulas en cultivo, y posteriormente el grupo de Lopez-Barneo y de Peers lo verificaron en clula nica con tcnicas voltamtricas y ampermtricas y en clulas identificadas en una preparacin de cortes de CC. La liberacin de CA en el CC intacto o clulas quimiorreceptoras aisladas trasciende a la neurotransmisin sensu stricto, ya que es un parmetro que se utiliza como ndice de la activacin de las clulas quimiorreceptoras: no existe en nuestro conocimiento ningn estmulo para el CC que no aumente la liberacin de CA y adems existe una correlacin razonable entre intensidad
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
55 de estimulacin y aumento de liberacin de DA y de frecuencia de potenciales de accin en el NSC frente a mltiples estmulos (Figura 8).
Figura 8. Relacin entre la liberacin de DA y la actividad elctrica en el NSC evocadas por distintos estmulos. La Figura muestra slo una intensidad de estimulacin para cada estmulo, pero en los estudios originales se realizaron curvas con distintas intensidades de estimulacin en todos los casos.
Sin embargo, existen dos estudios de Donnelly (62,63) en los que muestra que frente a estimulacin repetida o en animales reserpinizados tal paralelismo se rompe: la liberacin de DA disminuye con cada nuevo estmulo pero la frecuencia de descargas en el NSC se mantiene esencialmente constante. Hay que destacar que ste es el patrn normal de conducta de los sistemas catecolaminrgicos a la estimulacin repetida y a la reserpinizacin (64-66). Finalmente, Iturriaga et al. (67), an cuando se observa el paralelismo antes mencionado, los autores dicen obtener una disociacin temporal entre liberacin de DA y actividad en el NSC, en el sentido que la ltima precedera a la primera. Con tcnicas voltamtricas idnticas Buerk et al. (68) llegaron a la conclusin opuesta. Adems la experiencia de este laboratorio (Dr. R. Rigual, comunicacin personal) demuestra que el establecimiento de la relacin temporal con tcnicas voltamtricas es imposible de hacer con fiabilidad, ya que depende de dnde, en el espesor del CC, se implante el electrodo de fibra de carbono para registrar liberacin de CA, y dnde tenga su origen la unidad quimiorreceptora del NSC que se est registrando. Ambos localizaciones son incontrolables experimentalmente. Agradecimientos Subvencionado por la DGICYT (Proyecto BFU2004-06394), por el ICiii (RED Respira, RTIC C03/011 y Proyecto PI042462) y por la JCyL Proyectos VA011C05 y VA106A05.
Veces la actividad basal en normoxia
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
56 5. BIBLIOGRAFA 1. DE CASTRO, F. (1928). Sur la structure et l'innervation du sinus carotidien de lhomme et des mammifres: Nouveaux faits sur l'innervation et la fonction du glomus caroticum. Trab. Lab. Invest. Biol. Univ. Madrid. 25: 330-380. 2. HEYMANS, C., BOUCKAERT, J.J. & DAUTREBANDE, L. (1930). Sinus carotidien et rflexes respiratoires. II. Influences respiratoires rflexes de l'acidose, de l'alcalose, de l'anhydride carbonique, de l'ion hydrogene et de l'anoxmie: Sinus carotidiens et echanges respiratoires dans les poumons et au del des poumons. Arch. Int. Pharmacodyn. Ther. 39: 400 408. 3. COMROE J. (1939) The location and function of the chemoreceptors of the aorta. Am. J. Physiol. 127: 176-191. 4. DEJOURS, P. (1968). Approaches to the study of arterial chemoreceptors. In: Arterial chemoreceptors. (Ed. Torrance, R.W.). Blackwell. Oxford. pp 41-49. 5. FITZGERALD, R.S. & SHIRAHATA, M. (1997). Systemic responses elicited by stimulating the carotid body: primary and secondary mechanisms. In: The Carotid Body Chemoreceptors. (Ed. Gonzalez, C). Springer-Verlag. Heidelberg, Germany. pp.171- 191. 6. PENG, Y.J. & PRABHAKAR, N.R. (2004). Effect of two paradigms of chronic intermittent hypoxia on carotid body sensory activity. J Appl Physiol. 96: 1236-1242. 7. PENG, Y.J., OVERHOLT, J.L., KLINE, D., KUMAR, G.K. & PRABHAKAR, N.R. (2003). Induction of sensory long-term facilitation in the carotid body by intermittent hypoxia: implications for recurrent apneas. Proc Natl Acad Sci U S A. 100:10073-8. 8. VAN LUTEREN, E., (1997). Upper airway effects on breathing. In: The Lung: Scientific Foundations. Second Edition. (Ed. Crystal, R., West, J., Weibel, E. & Barnes, P). Lippincott Raven. Philadelphia. pp: 2073-2084. 9. DEMPSEY, J., HARMS, C., MORGAN, B., BODR, S. & SKATRUD, J. (1997). Sleep effects on breathing and breathing stability. In: The Lung: Scientific Foundations. Second Edition. (Ed. Crystal, R., West, J., Weibel, E. & Barnes, P). Lippincott Raven. Philadelphia. pp.2063-2072. 10. FENIK, V., DAVIES, R.O., PACK, A.I. & KUBIN, L. (1998). Differential suppression of upper airway motor activity during carbachol-induced, REM sleep-like atonia. Am J Physiol. 275: R1013-1024. 11. MORRISON, D.L., LAUNOIS, S.H., ISONO, S., FEROAH, T.R., WHITELAW, W.A. & REMMERS, J.E. (1993). Pharyngeal narrowing and closing pressures in patients with obstructive sleep apnea. Am Rev Respir Dis. 148: 606-611.
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
57 12. HARMS, C.A., ZENG, Y.J., SMITH, CA., VIDRUK, E.H. & DEMPSEY, J.A. (1996). Negative pressure-induced deformation of the upper airway causes central apnea in awake and sleeping dogs. J Appl Physiol. 80:1528-1539. 13. CARRERA, M., BARBE, F., SAULEDA, J., TOMAS, M., GOMEZ, C. & AGUSTI, A.G. (1999). Patients with obstructive sleep apnea exhibit genioglossus dysfunction that is normalized after treatment with continuous positive airway pressure. Am J Respir Crit Care Med. 159: 1960-1966. 14. McGUIRE, M., ZHANG, Y., WHITE, D.P. & LING, L. (2002). Effect of hypoxic episode number and severity on ventilatory long-term facilitation in awake rats.J Appl Physiol. 93: 2155-2161. 15. WIDDICONBE, J.H. (2002). Regulation of the depth and composition of airway surface liquid. J Anat. 201: 313-318. 16. HEYMANS, C., BOUCKAERT, J.J. & DAUTREBANDE, L. (1931). Sinus carotidien et rflexes respiratoires.; sensibilit des sinus carotidiens aux substances chimiques. Action stimulante respiratoire rflexe du sulfure de sodium, du cyanure de potassium, de la nicotine et de la lobline. Arch Int Pharmacodyn Ther; 40: 54-91. 17. OBESO, A., ALMARAZ, L. & GONZALEZ, C. (1989). Effects of cyanide and uncouplers on chemoreceptor activity and ATP content of the cat carotid body. Brain Res. 481: 250-257. 18. ORTEGA-SAENZ, P., PARDAL, R., GARCIA-FERNANDEZ, M. & LOPEZ-BARNEO, J. (2003). Rotenone selectively occludes sensitivity to hypoxia in rat carotid body glomus cells. J Physiol. 548: 789-800. 19. WYATT, C.N. & BUCKLER, K.J. (2004). The effect of mitochondrial inhibitors on membrane currents in isolated neonatal rat carotid body type I cells. J Physiol. 556: 175-191. 20. ZAPATA, P. (1997). Chemosensory activity in the carotid nerve: Effects of pharmacological agents. In: The Carotid Body Chemoreceptors. (Ed. Gonzalez, C.). New York: Springer-Verlag; p. 119-146. 21. GONZALEZ, C., ALMARAZ, L., OBESO, A. & RIGUAL, R. (1994). Carotid body chemoreceptors: from natural stimuli to sensory discharges. Physiol. Rev. 74: 829-898. 22. BERKENBOSCH, A., VAN DISSEL, J., OLIEVIER, C.N., DE GOUDE, J. & HEERINGA, J. (1979). The contribution of the peripheral chemoreceptors to the ventilatory response to CO2 in anaesthetized cats during hyperoxia. Respir Physiol. 37: 381-90. 23. FITZGERALD, R.S. & PARKS, D.C. (1971). Effect of hypoxia on carotid chemoreceptor response to carbon dioxide in cats. Respir Physiol; 12: 218-229.
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
58 24. KAZEMI, H. (1997). Cerebrospinal fluid and the control of ventilation. In: The Lung: Scientific Foundations. (Ed. Crystal, R., West, J., Weibel, E. & Barnes, P). LippincottRaven. Philadelphia. pp.1777-1788. 25. CHERNIACK, N. & ALTOSE, M. (1997). Central chemoreceptors. In: The Lung: Scientific Foundations. (Ed. Crystal, R., West, J., Weibel, E. & Barnes, P). Lippincott Raven. Philadelphia. pp.1767-1776. 26. HONDA, Y. (1985). Role of carotid chemoreceptors in control of breathing at rest and in exercise: studies on human subjects with bilateral carotid body resection. Jpn J Physiol. 35: 535-44. 27. ZAPATA, P. (1997). Chemosensory activity in the carotid nerve: Effects of physiological variables. In: The Carotid Body Chemoreceptors. (Ed. Gonzalez, C.). New York: Springer-Verlag; p. 97-117. 28. DE CASTRO, F. (1926). Sur la structure et l'innervation de la glande intercarotidienne (glomus caroticum) de l'homme et des mammifres, et sur un nouveau systme d'innervation autonome du nerf glossopharyngien. Trab. Lab. Invest. Biol. Univ. Madrid. 24: 365-432. 29. CAMPANUCCI, V.A., FEARON, I.M. & NURSE, C.A. (2003). Sensing mechanisms in efferent neurons to the rat carotid body. Adv Exp Med Biol. 536: 179-185. 30. FIDONE, S. & GONZALEZ, C. (1986). Initiation and control of chemoreceptor activity in the carotid body. In: Fishman AP, ed. Handbook of Physiology. The Respiratoy System VI. Bethesda: Amer Physiol Soc. p. 247-312. 31. GONZALEZ, C. & FIDONE, S. (1977). Increased release of 3H-dopamine during low O2 stimulation of rabbit carotid body in vitro. Neurosci Lett. 6: 95-99. 32. FIDONE, S., GONZALEZ, C. & YOSHIZAKI, K. (1982). Effects of low oxygen on the release of dopamine from the rabbit carotid body in vitro. J Physiol Lond; 333: 93-110. 33. GONZALEZ, C., ALMARAZ, L., OBESO, A. & RIGUAL, R. (1992). Oxygen and acid chemoreception in the carotid body chemoreceptors. Trends Neurosci. 15: 146-153. 34. GONZALEZ, C., ROCHER, A. & ZAPATA, P. (2003). Arterial chemoreceptors: cellular and molecular mechanisms in the adaptative and homeostatic function of the carotid body. Rev Neurol. 36: 239-254. 35. RIESCO-FAGUNDO, A.M,, PEREZ-GARCIA, M.T., GONZALEZ, C. & LOPEZLOPEZ, J.R. (2001). O2 modulates large-conductance Ca(2+)-dependent K(+) channels of rat chemoreceptor cells by a membrane-restricted and CO-sensitive mechanism. Circ Res. 89: 430-436.
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
59 36. ACKER, H. (1994). Cellular oxygen sensors. Ann. N. Y. Acad. Sci. 718: 3-10. 37. CHANDEL, N.S. & SCHUMACKER, P.T. (2000). Cellular oxygen sensing by mitochondria: old questions, new insight. J. Appl. Physiol. 88: 1880-1889. 38. ANICHKOV, S. & BELENKII, M. (1963). Pharmacology of the carotid body chemoreceptors. Mew York. Macmillan. 39. WILLIAMS, S.E., WOOTTON, P., MASON, H.S., BOULD, J., ILES, D.E., RICCARDI, D., PEERS, C. & KEMP, P.J. (2004). Hemoxygenase-2 is an oxygen sensor for a calcium-sensitive potassium channel. Science. 306: 2093-2097. 40. PEREZ-GARCIA, M.T., LOPEZ-LOPEZ, J.R. & GONZALEZ, C. (1999). Kvbeta1.2 subunit coexpression in HEK293 cells confers O2 sensitivity to kv4.2 but not to Shaker channels. J Gen Physiol. 113: 897-907. 41. HARTNESS, M.E., BRAZIER, S.P., PEERS, C., BATESON, A.N, ASHFORD, M.L. & KEMP, P.J. (2003). Post-transcriptional control of human maxiK potassium channel activity and acute oxygen sensitivity by chronic hypoxia. J Biol Chem. 278: 51422-51432. 42. McCARTNEY CE, MCCLAFFRTY H, HUIBANT JM, ROWAN EG, SHIPSTON MJ, ROWE IC (2005). A cysteine rich motif confers hypoxia sensitivity to mammalian large conductance voltage and Ca-activated K (BK) channel alpha subunits. Proc Natl Acad Sci USA 102: 17870-17876. 43. LOPEZ-BARNEO, J., LOPEZ-LOPEZ, J.R., UREA, J. & GONZALEZ, C., (1988). Chemotransduction in the carotid body: K+ current modulated by PO2 in type I chemoreceptor cells. Science. 241: 580-582. 44. LOPEZ-LOPEZ, J.R., GONZALEZ, C., UREA, J. & LOPEZ-BARNEO, J. (1989). Low pO2 selectively inhibits K+ channel activity in chemoreceptor cells of the mammalian carotid body. J. Gen. Physiol. 93: 1001-1015. 45. LOPEZ-LOPEZ, J.R., DE LUIS, A. & GONZALEZ, C. (1993). Properties of a transient K+ current in chemoreceptor cells of rabbit carotid body. J. Physiol. 460: 15-32. 46. BUCKLER, K.J. (1997). A novel oxygen-sensitive potassium current in rat carotid body type I cells. J Physiol. 498: 649-662. 47. ROCHER, A., OBESO A., GONZALEZ, C. & HERREROS, B. (1991). Ionic mechanisms for the transduction of acidic stimuli in rabbit carotid body glomus cells. J Physiol Lond. 433: 533-548. 48. BUCKLER, K.J & VAUGHAN-JONES, R.D. (1994). Effects of hypercapnia on membrane potential and intracellular calcium in rat carotid body type I cells. J Physiol. 478: 157-171
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
60 49. BUCKLER, K.J.& VAUGHAN-JONES, R.D. (1994). Effects of hypoxia on membrane potential and intracellular calcium in rat neonatal carotid body type I cells. J Physiol. 476: 423-428. 50. ROCHER, A, GEIJO-BARRIENTOS, E., CACERES, A.I., RIGUAL, R., GONZALEZ, C. & ALMARAZ, L. (2005). Role of voltage-dependent calcium channels in stimulussecretion coupling in rabbit carotid body chemoreceptor cells. J Physiol. 562: 407-420. 51. EYZAGUIRRE, C. & ZAPATA, P. (1968). A discusin of posible transmitter or generator substances in carotid body chemoreceptors. In: Arterial Chemorreceptors. ed. (Torrance, R.). Blackwell. Oxford. pp. 213-251. 52. GAUDA, E.B, COOPER, R., JOHNSON, S.M, Mc LEMORE, G.L. & MARSHALL. (2004). C. Autonomic microganglion cells: a source of acetylcholine in the rat carotid body. J Appl Physiol. 96:384-391. 53. LEVER, J.D. & BOYD, J.D. (1957). Osmiophile granules in the glomus cells of the rabbit carotid body. Nature. 179: 1082-1083. 54. CHIOCCHIO, S., BISCARDI, A. & TRAMEZZANI, J. (1967). 5-Hydroxytryptamine in the carotid body of the cat. Science. 158: 790-791. 55. GONZALEZ, C., DINGER, B. & FIDONE, S.J. (1997). Functional significance of chemoreceptor cell neurotransmitters. In: The Carotid Body Chemocheceptors. (Ed. Gonzalez, C). Springer-Verlag. Heidelberg, Germany. pp. 47- 64. 56. McQUEEN, D.S. & RIBEIRO, J.A. (1986). Pharmacological characterization of the receptor involved in chemoexcitation induced by adenosine. Br J Pharmacol. 88: 615-620. 57. SPERGEL, D. & LAHIRI, S. (1993). Differential modulation by extracellular ATP of carotid chemosensory responses. J Appl Physiol. 74: 3052-3056. 58. RONG, W., GOURINE, A.V., COCKAYNE, D.A., XIANG, Z., FORD, A.P. & SPYER, K.M. (2003). Burnstock Pivotal role of nucleotide P2X2 receptor subunit of the ATP-gated ion channel mediating ventilatory responses to hypoxia. J Neurosci. 23: 11315-11321. 59. SPYER, K.M, DALE, N. & GOURINE, A.V. (2004). ATP is a key mediator of central and peripheral chemosensory transduction. Exp Physiol. 89: 53-59. 60. HE, L., CHEN, J., DINGER, B., STENSAAS, L. & FIDONE, S. (2006). Effect of chronic hypoxia on purinergic synaptic transmission in rat carotid body. J Appl Physiol. 100:157-62. 61. CONDE SV, OBESO A, VICARIO I, RIGUAL R, ROCHER A, (2006). Presence and functional significance of A2B adenosine receptors in rat carotid body chemoreceptor cells. J Physiol (enviado)
MECANISMOS DE QUIMIORRECEPCIN EN EL CUERPO CAROTDEO
61 62. DONNELLY, D.F. (1995). Does catecholamine secretion mediate the hypoxia-induced increase in nerve activity? Biol Signals. 4: 304-309. 63. DONNELLY, D.F. (1996). Chemoreceptor nerve excitation may not be proportional to catecholamine secretion. J Appl Physiol. 81: 657-664. 64. WAKADE, A.R. & KRUSZ, J. (1972). Effect of reserpine, phenoxybenzamine and cocaine on neuromuscular transmission in the vas deferens of the guinea pig. J Pharmacol Exp Ther. 181: 310-317. 65. FUDER, H., MUSCHOLL, E. & WEGWART, R. (1976). The effects of methacholine and calcium deprivation on the release of the false transmitter, alpha-methyladrenaline, from the isolated rabbit heart. Naunyn Schmiedebergs Arch Pharmacol. 293: 225-234. 66. ARTALEJO, A.R., GARCIA, A.G., MONTIEL, C. & SANCHEZ-GARCIA P. (1985). A dopaminergic receptor modulates catecholamine release from the cat adrenal gland. J Physiol. 362: 359-368. 67. ITURRIAGA, R., ALCAYAGA, J. & ZAPATA, P. (1996). Dissociation of hypoxia-induced chemosensory responses and catecholamine efflux in cat carotid body superfused in vitro. J Physiol. 497: 551-564 68. BUERK, D.G., LAHIRI, S., CHUGH, D. & MOKASHI, A. (1995). Electrochemical detection of rapid DA release kinetics during hypoxia in perfused-superfused cat CB. J Appl Physiol. 78: 830-837.
CAPTULO 3
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
NEUS SALORD Y PERE CASN
Departamento de Neumologa. Hospital de la Santa Creu i de Sant Pau Facultad de Medicina. UAB. Barcelona Av. Sant Antoni M Claret 167, 08025. Barcelona. Fax: 93 556 56 01 Neus Salord: msalord@santpau.es.
PALABRAS CLAVE: Control ventilatorio, Patrn ventilatorio, Impulso inspiratorio, Respuesta a la hipoxia, Respuesta a la hipercapnia, Respuesta a cargas mecnicas.
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
64 NDICE DE CONTENIDOS: 1. Introduccin 2. Patrn respiratorio 3. Mtodos de medicin 4. Impulso inspiratorio 4.1. Mediciones del impulso inspiratorio 4.2. Determinaciones ante diferentes estmulos 4.3. Respuesta ventilatoria a la hipercapnia y a la hipoxia 4.4. Respuesta ventilatoria a la carga mecnica 4.5. Prueba del tiempo de apnea o Breath holding Time 5. Conclusin 6. Bibliografa
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
65 1. INTRODUCCIN La ventilacin es el resultado de una compleja interaccin de diferentes estmulos neuronales, qumicos, musculares y mecnicos que tienen como objetivo ltimo el mantenimiento de un equilibrio gaseoso y cido-base ante las diferentes demandas metablicas y actividades de la vida diaria. Para la correcta medida e interpretacin del control de la respiracin es imprescindible recordar, como se ha desarrollado en captulos anteriores, el circuito que configura el control ventilatorio. Aunque no se puede localizar con precisin, ste se puede esquematizar en cuatro componentes (figura 1), el control voluntario, el reflejo o automtico, el qumico y el control mecnico.
Figura 1. Esquema del Sistema de Control Ventilatorio.
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
66 Control voluntario: la respiracin est influenciada por aspectos del comportamiento que interfieren en el patrn ventilatorio y que lo modifican durante el habla, las emociones, estados de vigilia-sueo y actividades diarias. Los centros reguladores estn ubicados en el crtex parietofrontal y motor as como en reas subcorticales y se proyectan mediante diferentes vas sobre los centros pontomedulares (1). El control reflejo o automtico: est configurado por diversos grupos neuronales situados en la protuberancia, en el bulbo raqudeo y en las porciones superiores de la mdula espinal. En la protuberancia se halla el ncleo parabrachialis y ncleo fusiforme de Klliker (tambin llamado clsicamente ncleo apnustico), a nivel del bulbo raqudeo estn los ncleos respiratorios ventrales y dorsales y el Ncleo del Tracto Solitario (2). Estos grupos determinan conjuntamente, a travs de neuronas exitatorias e inhibitorias, la ritmicidad del patrn respiratorio, integrando a su vez estmulos procedentes de los receptores perifrios y de los centros volicionales (3). Control qumico: los cambios producidos como consecuencia de la respiracin, reflejados en las presiones parciales de los gases y del equilibrio cido-base, se detectan en la periferia a travs del cuerpo carotdeo que responde principalmente a los cambios en pO2, pCO2 e H+ y de los cuerpos articos que responden a CO2 e H+ (4). Existen adicionalmente en los ncleos ventrolaterales medulares neuronas con capacidad de respuesta a los cambios de H+ y CO2 (5,6). Finalmente, a travs de los receptores mecnicos, formados por receptores propioceptivos y mecanoreceptores de la pared torcica y del diafragma as como los receptores de estiramiento de adaptacin lenta del parnquima pulmonar, se da respuesta a cambios fsicos provocados en diversas condiciones fisiopatolgicas, impulsos que trasmitidos a travs de la mdula espinal y del nervio vago alcanzan los centros respiratorios. Por otro lado, las agresiones agudas de la va area o del parnquima pulmonar estimulan a los receptores de estiramiento de adaptacin rpida de la va area y los receptores J del parnquima pulmonar que actan a travs del nervio vago (1). La funcin primordial de los pulmones, el intercambio de gases, es fcilmente objetivable mediante una simple determinacin de las presiones parciales de O2 y CO2 en la sangre arterial. Un aumento de la presin parcial de CO2 por encima de 45 mmHg en reposo, indica la alteracin de dicho equilibrio y se entiende como un fracaso ventilatorio. En la prctica clnica diaria es habitual interpretar los cambios en los gases sanguneos en trminos de cambios funcionales y estructurales del aparato respiratorio. Sin embargo, para el estudio de los mecanismos de produccin del fracaso ventilatorio, son necesarias diferentes pruebas que nos ayuden a entender las vas fisiopatolgicas que han conducido al paciente a la prdida de dicho equilibrio. Aunque no se conocen por completo los diferentes aspectos neuroanatmicos y neurofisiolgicos que controlan la ventilacin, a lo largo de las ltimas dcadas se han desarrollado diversos mtodos no invasivos que aplicados a la clnica, nos permiten entender y definir un amplio rango de alteraciones ventilatorias. Dichos mtodos se han basado en el estudio de la respuesta ventilatoria ante diferentes estmulos, varias mezclas gaseosas o cargas inspira-
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
67 torias, que aplicadas durante un perodo determinado de tiempo, geralmente unos pocos minutos, intentan extrapolar como se comporta el control ventilatorio en perodos prolongados. Hay que tener en cuenta este hecho, ya que la exposicin prolongada a diferentes estmulos puede dar efectos contradictorios a los producidos en perodos cortos (6). A lo largo de este captulo nos centraremos en el anlisis de las diferentes tcnicas para el estudio del control de la respiracin travs del conocimiento del patrn ventilatorio y del impulso respiratorio central, as como en su interpretacin. 2. PATRN RESPIRATORIO El ciclo respiratorio consiste en una fase de incremento del volumen pulmonar, la inspiracin y otra de descenso, la espiracin. Este hecho se puede definir por el volumen desplazado o circulante (VT) y el tiempo, que se clasifica por la duracin de la inspiracin (TI), duracin de la espiracin (TE) y la duracin total del ciclo (TTOT). Habitualmente se define el patrn ventilatorio por el volumen circulante (VT) y la frecuencia respiratoria (f).
Aunque se trata de una medicin til y sencilla, esta definicin no permite conocer los mecanismos que afectan al VT y a la f. El volumen circulante puede verse alterado por el aumento del flujo inspiratorio o por el cambio en el TI o por ambos. Variables que pueden ser controladas independientemente por diferentes vas reflejas y estmulos de los quimiorreceptores. Por ello es til desglosar el ciclo respiratorio en dos conceptos, como el resultado del impulso inspiratorio, tambin conocido en terminologia anglosajona como el driving y por el componente cronolgico o timing. De esta manera podemos desarrollar la anterior frmula de la siguiente manera:
donde el flujo inspiratorio medio (VT/ TI) es el ndice de intensidad del impulso respiratorio y TI/ TTOT representa la cronologa, definida por la fraccin inspiratoria del ciclo respiratorio (8,9,10). Aunque cada uno de estos eventos est determinado por un estmulo neuronal, el primero de ellos determina la intensidad del impulso y el segundo su cronologa. En general al exponer al aparato respiratorio a un estmulo externo, este responde modificando el flujo inspiratorio medio, sin embargo, este es el resultado de una cadena de eventos. Los centros respiratorios estimulan a los msculos respiratorios, que movilizan la caja torcica, a su vez sta a los pulmones y finalmente los pulmones movilizan el gas favoreciendo su intercambio a travs de la membrana alveolo-capilar. Por ello la medicin del patrn ventilatorio estar influenciada por mltiples factores que afectan a la mecnica pulmonar. En este sentido la medicin de la presin de oclusin de la va area en los primeros 100 milisegundos, que se
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
68 desarrollar ms adelante en este captulo, puede reflejar en este corto perodo de tiempo, el patrn que siguen los impulsos generados por el nervio frnico evitando, en parte, las interferencias de la mecnica del aparato respiratorio. 3. MTODOS DE MEDICIN La medicin del patrn ventilatorio puede realizarse directamente utilizando un saco de Douglas, a travs del clculo de la integracin de una seal de flujo, empleando un sensor a la altura de la boca o un analizador de gases. Existen diferentes tcnicas que se pueden separar entre las que utilizan boquillas para la realizacin de medidas a nivel de la va area y las tcnicas sin boquilla, presentado cada una de ellas diferentes ventajas e inconvenientes. En la figura 2 se describe un ejemplo de la tcnica llevada a cabo con boquilla. Se une la boquilla acoplada a una vlvula unidireccional, como una vlvula en Y de Collins, unida en la parte inspiratoria a un neumotacgrafo y su vez este a un transductor de presin. La seal elctrica de presin del transductor ser almacenada como flujo inspiratorio y podr amplificarse. A su vez la seal se integrar en volumen. Es importante intentar conseguir el mnimo espacio muerto para evitar as la reinhalacin de CO2 (11). Ser bsica la correcta calibracin tanto del nematocgrafo como del transductor de presin. Para la correcta realizacin de la medicin es importante que el paciente se encuentre confortable, puede estar en posicin sentado o supino, la boquilla debe encontrarse a la altura adecuada y es aconsejable un periodo de adaptacin de unos 3-5 minutos. A este sistema se pueden aadir toda una serie de determinaciones como la de la concentracin espiratoria de CO2 (End Tidal CO2)
Figura 2. Circuito para la medicin del patrn ventilatorio.
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
69 insertando un analizador de CO2 y/o O2 en la boquilla y aadiendo un saco de Douglas en la porcin espiratoria de la vlvula de Collins. Esto permitira la medicin del VO2 y VCO2 as como el cociente respiratorio (R) (12). La ventaja de la medicin con la boquilla, es la precisin de las medidas, as como la posibilidad de aadir la medida de la presin de oclusin de la va area. Sin embargo, hay que tener en cuenta que esta tcnica tiene la desventaja de alterar, por el hecho de respirar con la boquilla, el patrn ventilatorio en reposo del individuo. Se han descrito cambios del patrn ventilatorio definidos como el aumento del VT y el descenso de la frecuencia respiratoria por el uso de una simple boquilla o unas pinzas nasales (13). Por este motivo, se han desarrollado otras tcnicas mediante mascarillas faciales, que aunque tienen el inconveniente de posibles fugas, pretenden interferir en menor medida en el patrn ventilatorio espontneo del sujeto a estudio. Los efectos de la mscara sobre el patrn ventilatorio, no obstante, han sido poco estudiados (13,14). Para la medicin del patrn ventilatorio se han utilizado otras tcnicas como la magnetometra, que analiza el patrn ventilatorio a travs de los cambios del volumen torcico y la frecuencia respiratoria mediante la medicin de los movimientos de la pared torcica y abdominal (15,16). En el mismo sentido la pletismografia de induccin, utiliza transductores expandibles formados por dos coils de tefln unidos a bandas elsticas, colocados alrededor de la caja torcica y el abdomen. Se calculan los cambios del rea transversal a travs de la modificacin de la situacin que producen en los coils los movimientos torcicos y abdominales, calculando a partir de estos los cambios volumtricos (17,18,19). La utilizacin de pletismografa optoelectrnica puede ser una alternativa no invasiva para la medicin del patrn ventilatorio (20,21). Esta tcnica consiste en la colocacin de 4 cmaras alrededor del sujeto a estudio, con las que se recoge el desplazamiento de marcadores de posicin dispuestos en su caja torcica y abdomen. Las coordenadas de dichos marcadores se utilizan para extrapolar el volumen de un poliedro y a travs de una serie de frmulas geomtricas se puede calcular el volumen de cada tetraedro en que se divide el poliedro total. De la suma de los volmenes de cada unidad se obtendr el volumen total desplazado. La utilizacin de estas tcnicas sin emplear una boquilla tiene la ventaja de interferir en menor medida en el patrn ventilatorio. En su contra, la compleja calibracin en muchas ocasiones influenciada por los cambios posturales, la menor precisin y el inconveniente aadido de no poder aplicar simultniamente la medida de la presin de oclusin de la va area y una mayor dificultad para aplicar diferentes estmulos provocados por la inhalacin de diferentes mezclas gaseosas. Los valores de referencia obtenidos en el patrn ventilatorio dependern de la tcnica utilizada. 4. IMPULSO INSPIRATORIO La definicin del impulso respiratorio es un concepto artificial, creado para entender que a partir de la integracin de toda una serie de seales perifricas, de receptores qumicos y mecnicos, centrales (reflejas y voluntarias) se inicia una respuesta neuronal que tiene como objetivo
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
70 MTODOS DE MEDICIN DEL IMPULSO NEURAL
1. Electromiograma diafragmtico (EMGd): Tcnica a travs de catter esofgico. Tcnica con electrodos cutneos. 2. Presin de oclusin de la va area: Medicin de la presin de oclusin de la va area en los primeros 100 ms (P0.1) Medicin de la mxima defleccin de la onda de presin diferencial en el primer 0,2 segundo de iniciarse el esfuerzo inspiratorio (dP/tmax). 3. Presin transdiafragmtica (Pdi): Catter-baln esofgico y gstrico. Catter-baln esofgico Tabla 1. Mtodos de medicin del impulso neural.
mantener la ventilacin alveolar. La actividad del conjunto de neuronas, excitatorias e inhibitorias, que forman los centros respiratorios, no puede en la actualidad medirse directamente. Por este motivo se han creado toda una serie de pruebas indirectas para cuantificar el impulso respiratorio, que se detallan en la tabla 1. Sin embargo, hay que tener en cuenta que son tcnicas no validables en humanos, como medidas reales del impulso neural. 4.1. Mediciones del impulso inspiratorio La descarga del nervio frnico es una posible medida de dicho impulso. Su descarga moviliza al principal msculo inspiratorio, el diafragma. La medicin del electromiograma (EMG) diafragmtico nos permite valorar pues la activacin elctrica de este msculo, reflejo directo del impulso neuronal a travs del nervio frnico. Esta media requiere la colocacin de los electrodos bipolares a travs del esfago hasta alcanzar el nivel del diafragma. La seal del EMG es amplificada y requiere precisos filtros que neutralicen las mltiples interferencias como el electrocardiograma. Para la realizacin de esta tcnica se precisa una intensa colaboracin del paciente, as como la experiencia de un tcnico experto y se trata de un proceso con una complicada calibracin. Inicialmente, se realizaban medidas semicuantitativas del EMG diafragmtico (22) posteriormente Lopata et al desarrollaron las medidas del pico de actividad, que se refiere al pico medio de movimiento del EMG diafragmtico y el ndice medio de aumento de la actividad inspiratoria, que se obtiene dividiendo el pico de actividad por el tiempo inspiratorio. Este grupo describi que la curva de movimiento medio del diafragma, reflejaba el ndice medio del impulso neuronal y el reclutamiento de unidades musculares durante el esfuerzo respiratorio. Por otro lado, describieron que el pico de movimiento medio, que denotaba el pico de actividad inspiratoria alcanzada, poda ser un ndice del umbral desconector de la inspiracin. (23,24,25). Una aproximacin a esta tcnica es la a colocacin de electrodos a travs de la piel para captar el EMG diagragmtico. Es menos precisa debido a las mltiples interferencias con los
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
71 msculos intercostales, as como los cambios de postura de los electrodos con los movimientos respiratorios, lo que hace difcil su interpretacin (26). Otra aproximacin a la valoracin del impulso central es la medida de una accin que es consecuencia de la seal neuronal. En una respiracin normal la seal neuronal ser convertida finalmente en presin o flujo. De este concepto deriva la medicin de la presin de oclusin de la va area (P0.1) desarrollada durante los aos 70 (27,28,29,30,31,32). La P0.1 consiste en la medicin de la presin generada por los msculos inspiratorios contra la va area cerrada al final de la espiracin. Se mide a nivel de la boca en los primeros 100 milisegundos (ms) del inicio de la inspiracin con la va area ocluida. La P 0.1 refleja la presin inspiratoria disponible para iniciar la inspiracin y esta medicin se considera relativamente independiente de la mecnica pulmonar y del control volicional de la respiracin. Los valores de referencia de la P0.1 son 1,27 cm H2O con un coficiente de variacin de 50,6 % y la dP/dtmax s de 16,7 cm H2O por segundo, con un coeficiente de variacin de 36,2 % (28,29,30,31). La medicin de la presin de oclusin no est exenta de problemas tcnicos cuando se aplican en sujetos enfermos, ya que su valor depende del tamao diafragmtico y longitud de las fibras musculares en el momento de inicio de la inspiracin. Es el caso de la EPOC, en que los enfermos tienen un diafragma aplanado, hecho que conlleva una disminucin de la longitud del msculo, por lo que ser incapaz de producir la misma presin en respuesta al mismo estmulo. Los valores habituales en pacientes con EPOC se encuentran sobre 3 cm H2O, y en el caso de del paciente EPOC agudizado o del paciente con distrs respiratorio pueden ser mayores de 10 cm H2O. De hecho, algunos estudios han demostrado que la elevacin de la P0.1 por encima de 46 cm de H2O en el paciente con EPOC en curso de Ventilacin Mecnica (VM) se asocia al fallo de la extubacin (33). En el estudio de pacientes con enfermedades neuromusculares, la p0.1 basal suele hallarse ms elevada, aunque la curva de la respuesta de la p0.1 frente al estmulo de CO2 est disminuida, lo que no se puede traducir en una disminucin de la respuesta del centro respiratorio en los pacientes con enfermedades que cursan con debilidad muscular (34,35). El montaje necesario para la medida de la presin de oclusin es parecida al descrito anteriormente en la medida del patrn ventilatorio Hay que aadir un sistema de oclusin en la parte inspiratoria de la vlvula en Y de Collins, as como un transductor de presin que grave y amplifique la seal. La tcnica consiste en producir una serie de oclusiones inspiratorias durante la ventilacin. La vlvula de oclusin se activa justo al final de la espiracin, existen diferentes dispositivos manuales o elctricos, de manera que al inicio de la inspiracin la vlvula se encuentra ocluida. Una parte fundamental para esta medida es el hecho de que el sujeto no sea consciente del momento en el que se efecta la oclusin. Para ello el dispositivo de oclusin debe ser silencioso y su activacin debe realizarse fuera del campo de visin del sujeto, para evitar el mecanismo de aprendizaje y anticipacin del sujeto a la oclusin. Las oclusiones se realizan a intervalos determinados. Se pueden utilizar dos tipos de medidas de la presin de oclusin: La P0.1 que es la presin inspiratoria generada al final del primer 0.1 segundo desde el inicio del esfuerzo inspiratorio, que se refleja como la defleccin negativa del trazo de la onda de presin. Otra mediada es la dP/dtmax que se refiere a la mxima defleccin de la onda de presin diferencial en el primer 0,2
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
72 segundo de iniciarse el esfuerzo inspiratorio. Las dos medidas son igualmente vlidas como ndices del impulso inspiratorio neuromuscular, se describen en la figura 3 (28,30,31). En aquellos en casos que exista un bajo estmulo inspiratorio, en los que el inicio de la presin inspiratoria sera ms gradual, a veces es ms difcil extrapolar la P0.1 y, adems, en estos casos es ms fcil el clculo de la dP/dtmax (36,37). Sin embargo, actualmente el uso de mtodos de anlisis informatizados minimizan estos problemas que se hallaban en medidas ms artesanales. Estas determinaciones se repiten habitualmente unas 5-6 veces y se utiliza el valor medio.
Figura 3. Medicin de la presin de oclusin de la va area (P0.1) y la presin diferencial mxima dP/dtmax.
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
73 Otra forma ms directa pero ms invasiva, es la utilizacin de un cateter-baln situado a nivel del esfago y del estmago para la media de la presin transdiafragmtica (Pdi), que se define como la diferencia entre la presin pleural (Ppl) y la presin abdominal (Pabd) y que a la prctica se mide utilizando la diferencia entre la presin esofgica (Pes) y la gstrica (Pga) (38,39).
4.2. Determinaciones ante diferentes estmulos Con el objetivo de responder preguntas clnicas sobre el control ventilatorio en patologias concretas, durante los aos 60 y 70 se desarrollaron tcnicas que estudiaban la respuesta del aparato respiratorio a estmulos qumicos y cargas mecnicas. Cuestiones como diferenciar los pacientes con EPOC que desarrollaban hipoxia e hipercapnia frente a los que no desarrollaban hipercapnia (40,41), impulsaron el desarrollo de sistemas de anlisis ampliamente establecidos en la actualidad. En la tabla 2 se enumeran estmulos aplicados para el estudio del patrn ventilatorio y el impulso inspiratorio. ESTUDIO DEL PATRN VENTILATORIO Y DEL IMPULSO NEURAL FRENTE A ESTMULOS
1. Estmulos qumicos: mezclas gaseosas: Hipercapnia: Medicin a una concentracin determinada. Medicin a una concentracin creciente (Read 1967). Hipoxia: Medicin a una concentracin determinada. Medicin a una concentracin creciente (Rebuck and Campbell 1974). 2. Estmulos mecnicos: Respuesta cargas: Cargas elsticas Cargas resistivas Cargas umbral.
Tabla 2. Estudio del patrn ventilatorio y de la presin de oclusin de la va area frente a estmulos.
4.3. Respuesta ventilatoria a la hipercapnia y a la hipoxia Respuesta a la hipercapnia: La respuesta ventilatoria a la hipercapnia se puede medir mediante la inhalacin de una sola concentracin de CO2 o mediante su aumento progresivo. En el primer caso se utiliza un circuito parecido al descrito anteriormente en la evaluacin del patrn ventilatorio, aadiendo
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
74 un saco de Douglas con una mezcla gaseosa que contiene una concentracin determinada de CO2 (usualmente de un 5-8%) que se rellena continuametne de manera que se mantiene una concentracin constante (42). Despus de un periodo de 10-12 minutos en que se ha alcanzado la estabilidad, se realizan las mediciones de volumen/minuto y la P0.1. De esta manera se obtienen resultados de la ventilacin/minuto y de la presin de oclusin para una concentracin determinada. Este procedimiento puede repetirse con diversas concentraciones (44). Sin embargo, el mtodo de medida ms extendido para valorar la respuesta del impulso ventilatorio al CO2 es el de la reinhalacin, conocido en la terminologa anglosajona como rebreathing aunque esta tcnica fue desarrollada a lo largo de los aos 50. El mtodo ms utilizado fue descrito por Read en 1967 (43). En este sistema despus de la medicin basal del patrn ventilatorio y de la P0.1, se aade una concentracin de CO2 y O2 determinada en la parte inspiratoria del circuito a travs de una bolsa de 5-10 litros de volumen, se cierra el circuito de manera que el sujeto reinhala una mezcla gaseosa con alto contenido en oxgeno y en el que el contenido en CO2 va aumentando respiracin a respiracin hasta llegar a una PCO2 de 60 o 70 mmHg. En todo momento se registra la concentracin de CO2 en aire exhalado a nivel de la boca. Durante el proceso se mide la presin de oclusin de la va area, la ventilacin minuto y el patrn ventilatorio. Los datos se registran en volumen minuto en frente del aumento del CO2 exhalado y en trminos de presin por CO2 exhalado. Respuesta a la hipoxia: Para el estudio de la respuesta al estmulo hipxico se pueden realizar mediciones con una concentracin fija de gas pobre en oxgeno o mediante un descenso progresivo del oxgeno de la mezcla gaseosa. En el primer caso el circuito es idntico al descrito para la prueba con concentracin fija del CO2 para el estudio de la respuesta a la hipercapnia, usualmente se utilizan mezclas con una concentracin del 10% de O2 y habitualmente se alcanza la estabilidad en la muestra en un perodo inferior, de aproximadamente 2 minutos (44). Se monotoriza al paciente con un pulsioxmetro. Los resultados se registran mediante la saturacin arterial de O2 (SO2) y la fraccin inspirada de O2 frente a la presin de oclusin y al volumen/minuto. Una desventaja de este mtodo es el mantenimiento de la isocapnia, ser importante monotorizar en todo momento el CO2 exhalado y realizar ajustes en la mezcla gaseosa. En la respuesta a la hipoxia progresiva, existe una tcnica ampliamente establecida diseada por Rebuck y Campbell en 1974 (45), el circuito es idntico al descrito en la respuesta progresiva a la hipercapnia, pero en este caso se aade al circuito un absorbente de CO2 con una bomba de flujo variable, con la finalidad de manener la isocapnia, para ello se monotorizar el CO2 exhalado y se ajustar el flujo de la bomba para mantener el nivel de CO2 constante. Por otro lado la mezcla de gas inhalada ser de aire, de manera que respiracin a respiracin se conseguir un descenso progresivo de PO2 en condiciones de isocapnia. Los resultados se registraran con la SO2 y la fraccion inspirada de O2 versus la ventilacin minuto y la presin de oclusin.
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
75 4.4. Respuesta ventilatoria a la carga mecnica Con el objetivo de imitar situaciones similares a las que ocurren en la fisiopatologia de enfermedad respiratoria, se desarrollaron una serie de pruebas en que se somete al aparato respiratorio a una carga mecnica encaminada a estimular a los mecanoreceptores y se estudia su respuesta frente estmulos qumicos o se compara con un estado basal. Se puede someter al aparato respiratorio a diferentes tipos de cargas: resistivas, elsticas y de umbral. Las cargas resistivas consisten en aumentar la presin para un flujo dado, las cargas elsticas consisten en aumentar la presin para un volumen dado y pueden ser tanto inspiratorias como espiratorias. En las cargas-umbral se necesita una presin determinada para iniciar el flujo pero no para mantenerlo, imitando el fenmeno de la presin intrnseca al final de la espiracin (PEEPi) (46). Estas medidas intentan valorar la respuesta que los estmulos de los mecanoreceptores producen en los centros respiratorios. 4.5. Prueba del tiempo de apnea o Breath Holding Time Es una prueba de fcil realizacin. Se precisa tan solo una pinza nasal y un cronmetro. El sujeto realiza una espiracin profunda hasta alcanzar el volumen residual (VR), inspira hasta la capacidad pulmonar total (CPT) y mantiene la respiracin tanto tiempo como le es posible al sujeto. Si se aade el anlisis del gas espirado, se puede medir el tiempo de apnea con la presin parcial del gas (47). Se puede medir la respuesta ante diferentes mezclas gaseosas. La prueba deber repetirse hasta obtener resultados reproducibles o por la fatiga del sujeto. Esta tcnica no est exenta de mltiples interferencias a expensas de la fuerza muscular o de la geometra de la caja respiratoria (48, 49). Por otro lado, no existen unos valores de referencia establecidos, por lo que de ser utilizada debern establecerse valores propios. 5. CONCLUSIN El desarrollo de diferentes tcnicas que estudian la medida del patrn ventilatorio y del impulso inspiratorio nos acercan a una mejor comprensin de los mecanismos fisiopatolgicos que conducen al fracaso ventilatorio. La eleccin de la tcnica para su estudio depender de la cuestin planteada y de las limitaciones que presenta cada tcnica segn la patologa a estudio. Para su correcta interpretacin es necesario recordar que se tratan de medidas indirectas que son el reflejo de toda una jerarqua de estmulos reflejos, qumicos, mecnicos y voluntarios que se extienden desde el crtex cerebral a los receptores pulmonares y de la caja torcica, que se integran en una respuesta que llamamos impulso respiratorio. Queda an mucho por conocer y desarrollar de los mtodos descritos para que dichas medidas sean una herramienta til, que nos permita la toma directa de decisiones clnicas.
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
76 6. BIBLIOGRAFA 1. Euler C von. Brain stem mechanisms for generation and control of breathing pattern. In: Cherniack NS, Widdicombe JG, Handbook of Physiology; Control of Breathing . Vol 2, part 2. Bethesa ed. American Physiology society.1986; pp. 595-619. 2. Ritchter DW. Generation and maintenance of respiratory rhytm. J Exp Biol 1982; 100:93-107. 3. Euler C von. On the central pattern generador for the basic breathing rhytmicity. J Appl Physiol: Respirat Environ Exercise Physiol 1983; 55(6) 1647-1659. 4. Bisgard GE, Neubauer JA. Periferal and central effects of hypoxia. In. Dempsey JA, Pack AI, eds. Regulation of Breathig. 2on Edn. New York, Marcel Dekker 1995; pp 617-668. 5. Bruce EN, Cherniach; NS. Central chemoreceptors. J Appl Physiol 1987;62 (2):389-402. 6. Bledsoe SW, Hornbein TF. Central chemoreceptors and the regulation of their chemical environment. Hornbein, ed. Regulation of breathing. New York, Marcel Dekker, 1981: 347-406. 7. Easton PA, Slykrman LJ, Anthonisen NR. Ventilatory response to sustained hypoxia after pretreatment with aminophyline. J Appl Physiol 1988; 61:255-264. 8. Clark FJ, Von Euler C. On the regulation of depth and rate of breathing. J Physiol (London) 1972; 222:267-295. 9. Milic-Emili J, Grusnstein MM, Drive and timing components of breathing. Lung 1976;160:1-17. 10. Derenne J, Couture J, Iscoe S, Whitelaw WA, Milic-Emili J. Regulation of breathing in anesthesized human subjects. J Appl Physiol 1976; 40:804-814. 11. Maxwell DL, Cover D, Hughes JMB. Effect of respiratory apparatus on timing and depth of breathing in man. Respir Physiol 1985; 61:255-264. 12. Burki NK. Measurements of Ventilatoy Regulation. Clin in Chest Med 1989; 10 (02): 215-226. 13. Askanazi PA, Silverberg A, Foster RJ, Hyman AI, Milic-Emili J, Kinney JM l: Effects of respiratory apparatus on breathing pattern. J Appl Physiol: Respir Environ exerc Physiol 1980; 48 (4):577-580. 14. Gilbert R, Auchincloss JH, Brodsky J, Boden W: Changes in tidal volume, frequency and ventilation induced by their measurement. J Appl Physiol 33:252.
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
77 15. Konno K, Mead J. Measurements of separate volume changes of ribcage and abdomen during breathing. J Appl Physiol 1967; 22: 407-422. 16. Robertson CH, Bradley ME, Homer LD. Comparison of two and four magnetometer methods of measuring ventilation. J Appl Physiol 1980; 49:355-362. 17. Sackner JD, Nixon AJ, Davis B, Atkinson N, Sackner MA. II Effects of breathing through external deadspace on ventilacion at rest and during exercise. Am Rev Dis 1980;122: 933-940. 18. Shea SA, Walter J, Pelley K Murphy K, Guz A. The Effect of visual and auditory stimuli upon resting ventilation in man .Respir Physiol 1987 ; 68 : 345-357. 19. Tobin MJ, Chadha TS, Jenouri G, Birch SJ, Gazeroglu HB, Sackner MA. Breathing patterns, 1: Normal subjects. Chest 1983;84: 202-205. 20. Cala SJ, Kenyon CM, Ferrigo G, Carnevali P,Alivelti A, Pedotti A, Macklen PT, Rochester DI. A Chest wall and lung volume estimation by optical refelctance motion analysis. J Appl Physyol 1996; 81: 2680-2689. 21. Aliverti A, Decalla R, Pelosi P, Chiumello D, Gattinoni L, Pedotti A. Compartmental analysis of breathing in the supine and prone positions by Opto-electronic pletismography. Ann Biomed Eng 2001; 29: 60-70. 22. Agostoni E, Sant Ambrogio G, Portillo-Carrasco H. Electromyography of the diaphragm in man and transdiaphragmatic pressure.J Appl Physiol 1960; 15:1093-1097. 23. Evanich MJ, Lopata M, Lourenco RV. Analytical methods for the study of electrical activity in respiratory nerves and muscles. Chest 1976;70(S): 156-1962. 24. Lopata M, Evanich MJ, Lourenco RV. Quantification of diaphragmatic EMG response to CO2 rebreathing in humans. J Appl Physiol. Respir Environ Exerc Physiol 1977; 43: 262-270. 25. Lopata M, Myron JE, Ruy VL. Realationship between Mouth Occlusion pressure and electrical activity of the diaphragm: Effects of Flow-Resistive Loading. Am Rev Res Dis 1977;116: 449-455. 26. Lopata M, Onal E, Cromydas G. Respiratory load compensation in chronic airway obstruccion. J Appl Physiol 1985; 1947-1954 27. Fahey PJ, Hyde R W. Wont breathe vs Cant Breathe. Detection of Depressed Ventilatory Drive in Patients with Obstructive Pulmonary Disease. Chest 1983; 84 (1): 19-25. 28. Burki NK, Mitchel LK, Chaudhary BA. Zechman FW. Measurement of mouth occusion pressure as an index of respiratory centre output in man. Con Sci Mol Mel 1977;53: 117-123.
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
78 29. Burki NK. The effects of changes in functional residual capacity with posture in patients with chronics obstruction of the airways. Chest 1979;76: 629-635. 30. Mathews AW, Howel JBL. The rate of isometric inpiratory pressure development as a measure of responsivenss to carbon dioxide in man. Clin Sci Mol Med 1980; 1:4-12. 31. Whitelaw WA, Derene J, Milic-EmiliJ. Occlusion pressure as a measure of respiratoy center output in conscious man. Respir Physiol 1975; 23:181-199. 32. Whitelaw A, Derenne JP. Airway occlusion pressure. J Appl Physiol 1993; 74(4): 1475-1483. 33. Saason CSH, Te TT, Mahutte CK, Light RW. Airway occlusion pressure: an important indicator for successful weaning in patients with crhonic obstructive pulmonary disease. Am Rev Dis 1987;135:107-113. 34. Baydur A. Respiratory muscle strengh and control of ventilation in patients with neuromuscular disease. Chest 1991:99:330-338. 35. Gibson GJ, Gilmartin JJ, Veale D, Walls TJ, Serisier DE. Respiratory muscle function in neuromuscular disease. In Jones NL, Killian KJ, editors, Breathless. Hamilton, Canada: CME;1992 p.66-71. 36. Jammes Y, Auran Y, Gouvernet J. The ventilatory pattern of conscious man according to age and morphology. Bull Eur Physiopathol Respir 1979;15:527-540. 37. Javaheri S, Blum J, kazemi H. Pattern of breathing and carbon dioxide retencion in chronic obstructive pulmonary disease. An J Med 1981;71: 228-234. 38. Cherniak RM, Farhi LE, Armstrong BW, Proctor DF. A comparison of esofageal and intrapleural pressure in man. J Appl Physiol 1955;8:203-211. 39. Tzelepis GE, Nasiff L, McCool FD, Hammond J. Transmision of pressure within the abdomen. J Appl Physiol 1996;81:1111-1114. 40. Gorini M, Misuri G, Corrado A, Duranti R, Iandelli I, De la Paola E, Scano G.Breathin pattter nad carbon dioxide retention in severe obstructive pulmonary disease. Thorax 1996; 51: 677-683. 41. Gorini M, Spinelli A, Gianni R, Duranti R, Gigliotti F, Scano G. Neural respiratory dirve and neuromuscular coupling in patients with chronic obstructive pulmonary disease. Chest 1990; 98:1179-1186. 42. Gardner WN. The relation betwen tidal volume and inspiratory and expiratoy time during steady-state carbon dioxide inhalation in man. J Physiol 1977; 272:591-611
EVALUACIN DEL PATRN RESPIRATORIO Y DEL IMPULSO INSPIRATORIO CENTRAL
79 43. Read DJC. A clinical method for assessing the ventilatory response to carbon dioxide. Australas Ann Med 1967; 16:20-32. 44. Gardner WN. The pattern of breathing following step changes of alveolar partial pressures of carbon dioxide and oxygen in man. J Physiol 1980;300:55-73 45. Rebuck and Campbell EJM. A clinical method for assessing the ventilatory response ot hypoxia. Am Rev Respir Dis 1974; 109:345-350. 46. Cherniak N, Milic-Emili J. Mechanical aspects of Loaded Breathing. The Thorax. Part B. C Rousos, PT Macklan Eds. Lung Biol Health Dis. Marcel Dekker New York 1985; 29:751-786. 47. Stanley NN, Cunninghan EL, Altose MD, Kelsen SG, Ward CF, Cheniak NS. Evaluation of breath holding in hypercapnia as a simple clinicaltest of respiratory chemosensivity. Thorax 1975; 30 337-343. 48. Stanley NN, Altose MD, Kelsen SG, Ward CF, Cheniak NS. Changing effect of lung volme on respiratory drive in man. J Appl Physiol 1975; 38:768-773. 49. Whitelaw WA, Derene J, Noble S, McBride B. Similarites between behaviour of respiratory muscles in breath-holding and in elastic loading. Respir Physyol 1988; 72: 151-161.
CAPTULO 4
EVALUACIN DEL CONTROL QUMICO Y MECNICO DE LA RESPIRACIN
MARIA JESS RODRGUEZ NIETO, GERMN PECES-BARBA ROMERO
FUNDACIN JIMNEZ DIAZCAPIO AVENIDA REYES CATLICOS 2 28040 MADRID FAX: 915504954 e-mail: mjrodriguezn@fjd.es
PALABRAS CLAVE: Control de la ventilacin, quimiorreceptores centrales, quimiorreceptores perifricos, respuesta a la hipoxia, respuesta a la hipercapnia, presin de oclusin, centros respiratorios, carga mecnica, estado estable, reinhalacin, ventilacin minuto, hipoventilacin alveolar, estmulos centrales, estmulos perifricos.
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
82 NDICE DE CONTENIDOS: 1. Introduccin 2 Fundamentos de las pruebas 3 Valoracin de la respuesta 4. Respuesta de los quimiorreceptores a la hipoxia y a la hipercapnia a. Respuesta a la hipoxia i Mtodo del estado estable ii. Mtodo reinhalacin iii. Mtodo de la hiperoxia intermitente b. Respuesta a la hipercapnia i. Mtodo del estado estable ii. Mtodo reinhalacin iii. Relacin PCO2/PO,1 5. Respuesta mecnica a. Presin de oclusin (P0,1) b. Presiones transpulmonares y EMG diafragmtico c. Carga mecnica 6. Apnea voluntaria 7. Control de la respiracin en distintas enfermedades respiratorias 8. Indicaciones de realizacin de las pruebas de valoracin de los centros respiratorios 9. Bibliografa
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
83 1. INTRODUCCIN El control de la respiracin es un proceso complejo y multifactorial que pretende mantener en muy diferentes circunstancias, como son por ejemplo el ejercicio y el reposo, un rango estrecho de valores para el pH y unos valores normales de O2 en sangre que permitan el normal funcionamiento de los distintos rganos y sistemas. El fallo en el control de la ventilacin provoca niveles inadecuados de ventilacin o patrones de ventilacin anormales, que conllevan una alteracin de estos parmetros en la sangre. Las pruebas que nos permiten evaluar el control de la ventilacin requieren considerar la existencia de numerosos factores que complican su interpretacin (1). Adems de la influencia emocional y voluntaria sobre la ventilacin, sta se modifica por la estimulacin de receptores mecnicos de la pared torcica, receptores de la va area (irritacin, alargamiento) y los receptores J del parnquima pulmonar (2). Todos ellos envan impulsos al centro respiratorio medular. La respiracin se estimula por la disminucin del pH y la disminucin de la pO2 en los receptores perifricos (cuerpo carotdeo y cuerpos articos) y tambin por la disminucin del pH en el lquido cfaloraqudeo captado por los receptores centrales localizados en la superficie ventrolateral del bulbo raqudeo. Una compleja red nerviosa enva estos estmulos a los centros respiratorios, tanto medulares como supramedulares. Los medulares son los mximos responsables del ritmo ventilatorio y los supramedulares tendran una funcin probablemente reguladora de los centros medulares. El impulso nervioso central resultante debe ser transmitido a travs de los nervios intercostales y frnico hacia la musculatura respiratoria del cuello, trax y abdomen. La ventilacin pulmonar no slo se altera por la presencia de problemas en este complejo circuito, sino que la presencia de obstruccin en la va area superior o enfermedades restrictivas u obstructivas pulmonares pueden tambin modificarla. El primer paso en la clnica para evaluar el control de la respiracin es realizar una gasometra arterial. Valores de pH, pO2 y pCO2 normales excluyen, de forma grosera, anormalidades del control de la respiracin pero son compatibles con alteraciones durante el sueo o el ejercicio. Por otra parte los valores anormales no son diagnsticos de alteraciones en el control de la respiracin ya que como sabemos pueden ser debidos a otras causas (desigualdades V/Q, bajo gasto cardaco, etc). No hay una prueba sencilla que permita evaluar el control de la ventilacin. Los distintos mtodos intentan actuar sobre alguno de los puntos del circuito o arco que controla la ventilacin (figura 1), por ejemplo, estudiando la respuesta a variaciones en los niveles de O2 o de CO2 en la sangre y comparando esta respuesta a la que tienen los sujetos sanos en similares circunstancias. La amplia variabilidad tanto inter como intra individual en la respuesta observada en estas pruebas limitan su uso clnico.
2. FUNDAMENTO DE LAS PRUEBAS Las pruebas se basan en alterar un componente del impulso de la ventilacin, bien utilizando estmulos ya existentes que aumentamos o disminuimos o bien introduciendo nuevos estmulos. La respuesta consiste en un cambio en la ventilacin o en el trabajo respi-
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
84
Figura 1. Arco del sistema de control de la ventilacin.
ratorio. El cambio en la ventilacin en respuesta al estmulo puede representarse como una curva dosis-respuesta. Los estmulos. Pueden actuar directamente sobre receptores perifricos, centrales o ambos. Los receptores centrales se pueden estimular directamente al respirar una mezcla de gases que contenga CO2 y O2 (3). Tambin se estimula directamente por la hipertermia. Los receptores perifricos se estimulan con hipoxemia leve (4) o con sustancias como la progesterona. Cuando existe hipoxemia con hipercapnia se produce un estmulo tanto central como perifrico. Tipo de cambio. El estimulo que utilicemos para evaluar el control de la ventilacin puede aumentar o disminuir el impulso ventilatorio. La disminucin en el impulso ventilatorio tiene la ventaja que apenas produce aumento del trabajo del sistema respiratorio. Se puede disminuir respirando una mezcla hiperxica y tambin por hipnosis o induciendo una situacin de hipocapnia y/o alcalemia. El incremento en el impulso respiratorio se puede inducir por el ejercicio, tambin al respirar una mezcla hipxica o hipercpnica, aumentando la temperatura corporal o reduciendo el pH sanguneo. Factores de confusin. Cualquier cambio en la ventilacin alveolar de suficiente magnitud para alterar la ventilacin alveolar inevitablemente tiene efectos secundarios. Un aumento en la ventilacin que no se deba a hipercapnia conduce a una eliminacin adicional de CO2 y reduce el impulso respiratorio de la hipercapnia. Si disminuye la ventilacin el efecto ser el opuesto. Estos mecanismos de retroalimentacin se pueden interrumpir realizando las pruebas manteniendo constante una variable mientras monitorizamos el efecto de la otra. Lo ms fcil es respirar una mezcla de O2 y CO2. Se puede respirar una mezcla hiperxica que estimula el cuerpo carotdeo disminuyendo la ventilacin y el efecto observado se debe a la hipercapnia central con hiperoxia. Por otra parte se puede estudiar los efectos de distintos grados de hipoxia o hipercapnia manteniendo constante la otra variable. Esta tcnica llamada isocpnica, se controla con la concentracin del gas al final de la espiracin (end tidal o concentracin alveolar del gas) cuyo estudio nos permite interpretar los resultados.
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
85 3. VALORACIN DE LA RESPUESTA El estado de activacin de los centros respiratorios se determina mediante una serie de pruebas de laboratorio encaminadas a valorar el perfil del patrn ventilatorio y su respuesta a variaciones en los niveles de oxgeno o de carbnico en la sangre. El electromiograma (EMG) diafragmtico puede usarse para valorar el estado de los msculos y siempre que exista alteraciones de la mecnica respiratoria que puedan limitar la respuesta ventilatoria, aunque el mtodo ms til en estos casos de limitacin mecnica es el de la medicin de la presin de oclusin en la boca o P0,1. En este captulo nos vamos a centrar en las tcnicas que evalan el control qumico y mecnico de la respiracin. 4. RESPUESTA DE LOS QUIMIORRECEPTORES A LA HIPOXIA Y A LA HIPERCAPNIA La inhalacin de mezclas de gases hipxicos o hipercpnicos se ha utilizado desde hace tiempo para evaluar la sensibilidad de los quimiorreceptores. Tiene la limitacin de que evaluamos un sistema que interacta, y su interpretacin debe considerar los distintos factores que intervienen. Los distintos protocolos que se realizan durante estas tcnicas requieren que los sujetos inhalen o reinhalen a travs de una bolsa mientras medimos el volumen bien por un espirmetro o un neumotacgrafo. La tcnica de reinhalacin requiere una bolsa de aproximadamente 6-8 L de capacidad conectada por vlvulas unidireccionales que permiten la reinhalacin del gas. La tcnica del estado estable requiere una bolsa de mayor tamao (hasta 100L) o bien una de menor capacidad que vayamos rellenando durante el procedimiento. Se miden los cambios en la presin de CO2 y O2 en la boca conectando el sistema con los analizadores de estos gases. Estos cambios los relacionamos bien con la ventilacin o la presin de oclusin si hacemos esta tcnica. Para realizar esta ltima tcnica hace falta colocar una vlvula de oclusin en la va inspiratoria y medir la presin en la boca con un transductor de presin. Si se quiere mantener una situacin isocpnica se colocar en la va espiratoria un absorbente de CO2. Dado que el control de la ventilacin puede estar influenciado por la voluntad del sujeto, para obtener datos representativos debemos promediar el resultado de varias respiraciones y estandarizar en lo posible las condiciones de las pruebas (5). Los pacientes deben de estar confortablemente sentados y en reposo al menos 30 minutos antes de la prueba, la habitacin donde se realice debe estar en silencio, el sujeto no debe de comer en las 2 horas previas ni haber tomado estimulantes como cafena o alcohol. Debe tener la vejiga urinaria vaca y se le debe decir que intente no pensar en su respiracin, ni intentar controlarla. Puede ser til poner msica suave. Si durante la prueba hay que cambiar la concentracin de la mezcla de gases o se ocluye la vlvula, hay que realizarlo de una manera que el sujeto no lo prevea para que no vare voluntariamente la ventilacin. Las pruebas deben de hacerse por duplicado esperando 10 minutos de descanso entre ellas.
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
86 a. Respuesta a la hipoxia La respuesta ventilatoria a la hipoxia est mediada nicamente por receptores centrales si se mantiene una situacin isocpnica. La respuesta ventilatoria a la hipoxia se representa tradicionalmente con la relacin curvilnea ventilacin (VE) - PO2 que se vemos en la figura 2 donde tenemos la respuesta ventilatoria a la hipoxia a tres niveles constantes de PCO2. La medicin de la sensibilidad a la hipoxia tiene el inconveniente de que se ha de someter al paciente a reducciones de la PO2.
Figura 2. Relacin entre ventilacin y PO2 arterial realizada a tres niveles constantes de PaCO2: normocapnico, hipercapico e hipocapnico.
Existen 3 tcnicas que pueden usarse en la prctica: i. Mtodo del estado estable Permite la elaboracin de varias curvas de respuesta VE- PCO2 a diferentes niveles de PO2. Se requiere un mnimo de 5 minutos para alcanzar un nivel estable de PCO2 para trazar cada uno de los puntos de la curva de respuesta, aunque tambin es posible acelerar el proceso variando la PO2 mientras se mantiene estable la PCO2. Se trata de una prueba muy laboriosa e incmoda para los sujetos, principalmente cuando tienen que alcanzar niveles altos de PCO2 o bajos de PO2. ii. Mtodo de reinhalacin (hipxica-isocpnica) Mediante este mtodo el sujeto consigue alcanzar la hipoxia al consumir el O2 de la bolsa que reinhala, pero debe cuidarse de mantener un nivel estable de PCO2, para lo cual debemos
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
87 colocar un absorbente de CO2 en la va espiratoria del sistema. Afortunadamente este mtodo se puede simplificar ofreciendo el resultado mediante la relacin entre la saturacin de O2 y la ventilacin. De esta forma la saturacin de O2 puede medirse en vez de la PO2. Esta relacin es lineal y su pendiente de inclinacin depende de la sensibilidad de los quimiorreceptores (6). La dificultad de esta tcnica radica en mantener los niveles de PCO2 estables. iii. Mtodo de hiperoxia intermitente Este mtodo tiene la ventaja de evitar la exposicin de los sujetos a la hipoxia. Mediante la inhalacin de oxgeno se mide la reduccin que se va generando en el impulso ventilatorio, indicativo de la existencia de actividad de los cuerpos carotdeos. Este mtodo es mucho menos sensible que el de estado estable. b. Respuesta a la hipercapnia La tcnica que se utiliza con mayor frecuencia para estimar la respuesta ventilatoria a la hipercapnia es establecer la relacin VE - PCO2. Esta relacin es linear en sujetos normales (figura 3). La pendiente de la recta se utiliza para estimar la respuesta de los quimiorreceptores implicados, que son receptores centrales y tambin receptores perifricos si el valor de PO2 no es excesivo. Como puede verse en la figura 3, distintos valores de PO2 alteran la pendiente de la curva de respuesta de forma que la sensibilidad de los quimiorreceptores a la hipercapnia aumenta con niveles bajos de O2. Es por tanto muy importante mantener constante la PO2 durante la prueba para poder interpretar la relacin VE - PCO2.
Figura 3. caracterizacin de la respuesta ventilatoria con CO2. Vemos las pendientes de las distintas rectas con distintos niveles de PO2.
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
88 i. Mtodo del estado estable Tras elevar el valor de la PCO2 incrementando su concentracin en el gas inspirado, se mide el volumen minuto y la PCO2 alcanzados. Se necesitan unos 5 minutos de inhalacin de la mezcla de gases para considerar que se ha alcanzado el estado estable para realizar la medicin. Deben realizarse varias determinaciones a diferentes niveles de PCO2 para conseguir la curva de respuesta VE - PCO2, por lo que es una prueba prolongada e incmoda para los pacientes. ii. Mtodo de reinhalacin (hipercpnica-hiperxica) Al igual que en el anlisis de la respuesta a la hipoxia, este mtodo introducido por Read (7) simplifica la medicin y es el utilizado habitualmente en la clnica. El paciente reinhala el contenido de una bolsa con 7% de CO2, 50-90% de O2 y el resto de N2. A medida que progresa la reinhalacin, incrementa el nivel de PCO2, pero la concentracin de oxgeno se mantiene elevada, para que el resultado no se interfiera por una posible hipoxemia asociada. Habitualmente se prolonga la rinhalacion durante 4 minutos. Se registra la concentracin de CO2 en la bolsa y el nivel de ventilacin alcanzado construyendo una curva VE - PCO2 de respuesta. Las pendientes obtenidas mediante los dos mtodos, el de estado estable y el de reinhalacin son superponibles, aunque con este ltimo, la curva se desplaza levemente a la derecha (8). iii. Relacin PCO2- P0,1 Como veremos a continuacin la presin de oclusin (P0,1) es otro parmetro que nos permite evaluar el control de la ventilacin. Cambios en el valor de la PCO2 producen una alteracin en el valor de P0,1 alcanzado. Se ha sugerido que este cambio es el mejor indicador del estado de activacin de los centros respiratorios (9). 5. RESPUESTA MECNICA a. Presin de oclusin (P0,1) Los parmetros del patrn ventilatorio que se utilizan para evaluar la respuesta ventilatoria pueden verse alterados si hay algn nivel de sobrecarga mecnica a la respiracin, bien incrementos de la resistencia de la va area o bien por incremento de la resistencia elstica pulmonar. En estos casos, la limitacin mecnica del sistema provoca que la respuesta ventilatoria que debiera existir ante la activacin de los centros respiratorios est disminuida y as, el nivel de ventilacin no incrementa adecuadamente porque la sobrecarga mecnica lo impide. Estas interferencias se pueden evitar con una prueba introducida por Milic Emili et al. (10), que consiste en medir la presin en la boca durante una breve oclusin inspiratoria y considerar su valor en los primeros 100 milisegundos (P0,1) que en ese momento va a depender directamente de la activacin neuronal y est mnimamente alterada por la sobrecarga mecnica del sistema (11). Sin embargo, tambin en estos casos, el valor de la prueba puede verse influenciado por el retraso de transmisin de la presin pleural a la boca, la presencia de distorsin de la caja torcica o la activacin previa de los msculos abdominales en la espiracin (9). La prueba se realiza generalmente ocluyendo mediante un baln la va
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
89 inspiratoria mientras el sujeto est en la fase espiratoria. El baln se desinfla pasados 150 ms de iniciada la inspiracin, ya recogidos los valores de presin a los 100 ms y antes de que la oclusin apenas pueda llegar a ser perceptible por el sujeto. Tiene la ventaja de que precisa una tecnologa sencilla. La determinacin de la P0,1 puede hacerse por separado o en combinacin con la administracin de cargas mecnicas inspiratorias o espiratorias o variaciones de la PCO2 que pueden simular alguno de los aspectos presentes en las enfermedades respiratorias (12). b. Presiones transpulmonares y EMG diafragmtico El trabajo no elstico realizado por el pulmn durante la inspiracin proporciona una medida del impulso respiratorio que como la P0,1, es casi independiente de la condicin funcional del pulmn. El trabajo puede obtenerse con la medida de la presin esofgica durante la inspiracin; el trabajo inspiratorio se obtienen por el rea de la parte inspiratoria de la curva volumen-presin obtenida. La condicin de prueba invasiva por la necesidad de introducir un baln esofgico y/o gstrico, hacen que la medicin de estas presiones apenas se realicen en la prctica clnica. La realizacin del EMG diafragmtico tiene tambin importantes dificultades tcnicas, por lo que tambin se utiliza escasamente en la valoracin del estado de activacin de los centros respiratorios. c. Carga mecnica Estas tcnicas requieren tecnologa muy especializada y su utilidad en la prctica clnica est en investigacin. Se han desarrollado varios circuitos con carga durante la respiracin implicando bien exposicin a una carga simple y observar el efecto en la primera respiracin con carga en relacin a las respiraciones previas, o bien el estudio de los efectos de estimular la respiracin con hipoxemia o hipercapnia durante una carga resistiva fija (13). Los resultados pueden estar afectados por la naturaleza de la enfermedad a estudio y en este contexto la insuficiencia ventilatoria restrictiva debido a alteracin mecnica de la pared torcica o el efecto de la presencia de autoPEEP en los pacientes EPOC pueden hacer compleja la interpretacin de los resultados de la prueba (14). La disminucin de la ventilacin frente a la carga resistiva varia significativamente entre los distintos sujetos (15).
6. APNEA VOLUNTARIA Es una prueba sencilla que se realiza con una maniobra de capacidad vital seguida de una apnea que el sujeto debe mantener el mayor tiempo posible. Se recogen el tiempo de apnea y los valores de O2 y CO2 espirados. La prueba se repite hasta que se alcanza la reproductibilidad o el sujeto se fatiga. En sujetos jvenes y sanos es superior a 1 min, pero es una prueba difcil de interpretar en presencia de enfermedad, donde suele ser inferior a 20 segundos si existe disnea (9).Cuando se realiza una apnea voluntaria tras una inhalacin de aire, la PCO2 alveolar o arterial suele mantenerse constante al alcanzar el punto de ruptura, en un valor prximo a 50 mmHg, pero suele ser la hipoxia el factor desencadenante de esta ruptura. Si la apnea se precede de una inspiracin de oxgeno, la apnea se prolonga y entonces los
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
90 valores de CO2 llegan a ser mayores. Puede deducirse que los individuos con una baja respuesta al O2 o al CO2 tendrn una apnea ms prolongada. Los buceadores de lite tienen una baja respuesta a la hipercapnia pero no a la hipoxia (16). Como en otras pruebas que estudian el control de la ventilacin hay distintos factores que interfieren en su interpretacin. La fuerza de los msculos respiratorios y la geometra de la pared torcica son muy importantes para la capacidad de un sujeto de mantener la apnea. (17, 18).
7. CONTROL DE LA RESPIRACIN EN DISTINTAS ENFERMEDADES RESPIRATORIAS Enfermedad Pulmonar Obstructiva Crnica (EPOC) Es la enfermedad que cuenta con ms estudios desde esta perspectiva, intentando distinguir a los pacientes con EPOC que desarrollan hipoxia e hipercapnia de aquellos que mantienen valores de PCO2 dentro de la normalidad. La sobrecarga mecnica existente en las vas areas en la EPOC ha supuesto un inconveniente a la hora de separar las alteraciones en la regulacin de la respiracin de las propias alteraciones mecnicas de esta enfermedad. El simple incremento de la resistencia de la va area produce por s mismo una reduccin de la respuesta al CO2 (19). Si se mide la pendiente de respuesta de VE - PCO2 durante la prueba de reinhalacin, la activacin en trminos absolutos ser menor en sujetos normocpnicos con EPOC, pero si el nivel de ventilacin alcanzado durante la prueba se normaliza como porcentaje de la mxima capacidad ventilatoria, o en trminos de trabajo respiratorio generado, se obtiene que la pendiente es similar o incluso mayor en los pacientes con EPOC que en los sujetos normales sometidos a inhalacin de CO2 o a sobrecarga mecnica (20,21). Cuando la EPOC es ms grave y se presenta con hipercapnia crnica, la respuesta ya es menor (19,22). Es conocido que en presencia de hipercapnia crnica, la resistencia de la va area suele ser mayor, por lo que esta respuesta no puede ser atribuida slo a una disminucin de la activacin de los centros respiratorios. Para separar la influencia de estos dos componentes, se introdujo la tcnica del electromiograma diafragmtico en estos estudios de regulacin de la respiracin. De esta manera, se lleg a ver que los pacientes con EPOC e hipercapnia crnica tenan una actividad electromiogrfica reducida, comparada con la existente en sujetos normales o en pacientes con EPOC y normocapnia (23). Pero la dificultad de realizacin tcnica de esta prueba ha llevado a la utilizacin de la determinacin de la presin de oclusin (P0,1). Con esta prueba se vio que la activacin es mayor en pacientes normocpnicos con EPOC que en sujetos normales (21), pero que disminua claramente en presencia de hipercapnia crnica (9). El aumento de sobrecarga mecnica que se asocia a los casos de hipercapnia crnica estable no se acompaa, sin embargo, de fatiga de los msculos respiratorios. Ello se ha atribuido al patrn conformista de ventilacin adoptado, con una respiracin rpida y superficial adoptada por estos pacientes (24) y que favorece la hipercapnia por el incremento de la relacin espacio muerto/volumen corriente. En cambio, la hipercapnia inducida en sujetos normales tiene una respuesta claramente diferente, el estmulo respiratorio hace que se incremente el volumen corriente y descienda el tiempo espiratorio. El tiempo inspiratorio no aumenta, pero sube la rampa (flujo) para incrementar el
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
91 volumen inspirado. El descenso del tiempo espiratorio hace que el ciclo respiratorio se acorte (25). La hipoxia inducida en sujetos normales provoca unos cambios similares a los de la hipercapnia, excepto en que con la hipoxia tambin se acorta el tiempo inspiratorio y el ciclo respiratorio es aun ms rpido (26, 27). Dentro del campo del control de la ventilacin, en estos pacientes tambin se ha estudiado el efecto de la respiracin de altas concentraciones de O2. Hay resultados dispares que pueden estar en relacin con los distintos grados de gravedad que presentan los pacientes de los distintos estudios. Hay trabajos que apuntan a que la exposicin a concentraciones altas de O2 producen hipercapnia por aumento de las desigualdades de la ventilacin/perfusin (28), y en otros se demuestra, que algunos pacientes expuestos a altas concentraciones de O2 durante una exacerbacin desarrollan hipercapnia por una disminucin en la ventilacin, quizs debido a una disminucin de la sensibilidad de los quimiorreceptores. (29) Hipoventilacin alveolar primaria La hipoventilacin alveolar primaria o sndrome de Ondina se debe a la ausencia de respuesta de los quimiorreceptores a cambios en la PCO2 o la PO2 (30, 31). Es una enfermedad rara que se presenta principalmente en nios. Fue descrito por primera vez en 1962 (32), con la presentacin de tres pacientes sometidos a una intervencin neuroquirrgica en la regin medular alta, que presentaban periodos prolongados de apnea por alteracin del control automtico de la respiracin, pero manteniendo intacto el control voluntario. Desde entonces se han publicado 103 casos de sndrome de Ondina, un 19% de ellos desarrollado durante la edad adulta (33). La ausencia de respuesta de los quimiorreceptores se compensa durante el da por el control voluntario de la respiracin pero por la noche, cuando la influencia voluntaria desaparece, disminuye el volumen minuto cayendo la PO2 y aumentando la PCO2. Estos pacientes muestran una respuesta reducida a los estmulos de hipercapnia e hipoxia cuando estn despiertos y esto se acenta de forma dramtica durante el sueo. Respiracin peridica Existen dos tipos de respiracin peridica, la de Cheyne-Stokes y la del sndrome de apnea del sueo. La respiracin de Cheyne-Stokes puede ocurrir de manera espontnea en pacientes de varias patologas neurolgicas, metablicas y cardiacas, pero se hace ms patente durante el sueo. Supone una oscilacin en el ritmo de ventilacin, con fases de hiperpnea y fases de hipopnea o de apneas, intercaladas con periodos de respiracin normal. Las apneas son generalmente centrales, pero tambin pueden ser obstructivas, siendo frecuente que aparezcan en el mismo sujeto secuencialmente periodos de ambos tipos de apneas (35). Esta secuencia de patrones repetitivos de alteraciones del ritmo ventilatorio se debe a un anormal control de la ventilacin. La fase de hiperpnea aparece cuando los quimiorreceptores perifricos detectan la elevacin de la PCO2 y el descenso de la PO2 procedentes de la fase de apnea, el aumento de ventilacin de aqulla hace que se inviertan los valores y descienda la PCO2 a la vez que aumenta la PO2, lo que hace que vuelva a empezar el ciclo con una nueva apnea para compensar el exceso de ventilacin precedente. Quin empieza este ciclo y por qu se prolonga?. Se sabe que esta respiracin peridica aparece tambin en sujetos norma-
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
92 les en la transicin desde la vigilia al sueo. En la transicin desde la vigilia al sueo (fases I y II del sueo no REM) existen irregularidades del patrn ventilatorio, con bostezos, toses o cortos periodos de asfixia que pueden conducir a la respiracin cclica durante los primeros minutos del sueo (35), con posible aparicin de microdespertares o arousals durante las fases hiperpneicas de este periodo. En situaciones de insuficiencia cardiaca, existe un enlentecimiento del tiempo circulatorio con retardos en los tiempos de transferencia de la informacin qumica procedente de los quimiorreceptores que puede originar la respiracin cclica de Cheyne-Stokes (36). En casos de enfermedad neurolgica o metablica, tambin asociados a esta respiracin cclica, la causa podra ser un retardo en la respuesta de los centros respiratorio al estmulo procedente de los quimioreceptores. En el caso del sndrome de apnea del sueo, la respiracin peridica incorpora la obstruccin total o parcial de la va area superior que llegan a presentarse durante las fases de apnea o de hipopnea. Cuando la apnea o la hipopnea se prolongan, provocan un descenso de la pO2 y un aumento de la PCO2. Entonces, el estmulo quimiorreceptor, junto con el estmulo propioceptivo local procedente de la presin subatmosfrica intrafarngea, provocan una activacin de los centros respiratorios (37, 38), pero el flujo no se restablece hasta que esta activacin es lo suficientemente intensa como para estimular sincrnicamente y de manera suficiente a los msculos dilatadores farngeos y a los msculos respiratorios. En este momento, generalmente asociado a un microdespertar o arousal, se abre la va area colapsada y se reanuda la respiracin cerrando el ciclo que se haba iniciado con la apnea o la hipopnea. 8. INDICACIONES DE REALIZACIN DE LAS PRUEBAS DE VALORACIN DE LOS CENTROS RESPIRATORIOS En general, las pruebas de valoracin de los centros respiratorios tienen escasa aplicacin en la clnica (Tabla I). La principal indicacin de estas pruebas se hace ante la sospecha de un sndrome de hipoventilacin alveolar primario, tras confirmar la ausencia de anomalas funcionales mecnicas respiratorias, del intercambio de gases y de enfermedades neuromusEnfermedad Sndrome de hipoventilacin alveolar Indicacin Cuando las pruebas de mecnica y de intercambio de gases son normales Cuando existe hipercapnia con escasa afectacin mecnica Cuando existe escasa afectacin mecnica Para valorar el xito de la extubacin
Enfermedades neuromusculares
Sndrome de hipoventilacin-obesidad Insuficiencia respiratoria en ventilacin mecnica
Tabla 1. Principales indicaciones de realizacin de las pruebas de valoracin de los centros respiratorios.
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
93 culares. Existir una alteracin de los gases arteriales como consecuencia de la hipoventilacin debida a la reduccin del impulso respiratorio, con descensos de las respuestas a la hipoxia y a la hipercapnia. Cuando existe una enfermedad neuromuscular, la aparicin de una elevacin de la PCO2 arterial puede ser debida a la propia debilidad muscular que no es capaz de mantener una ventilacin adecuada, o a una alteracin asociada del control ventilatorio que puede provocar la hipercapnia, a pesar de conservar una adecuada capacidad ventilatoria (39). De todas formas, la interpretacin de las pruebas en presencia de neuromiopata debe considerarse siempre con cautela. Un valor reducido de P0,1 no implica necesariamente alteracin del impulso ventilatorio. Una curva plana de respuesta al CO2 puede ser atribuida a alteracin de la respuesta a nivel de quimiorreceptor o a nivel central, pero respuestas disminuidas, no completamente planas son ms difciles de interpretar. La obesidad puede presentarse con hipoventilacin alveolar, consecuencia generalmente de la sobrecarga mecnica asociada, pero tambin tiene asociada la presencia de unas respuestas reducidas a la hipoxemia y a la hipercapnia, aunque esta reduccin puede ser secundaria a la hipoventilacin en s, ms que ser una situacin iniciada por la propia obesidad (40). En algunos casos de pacientes con insuficiencia respiratoria sometidos a ventilacin mecnica y con dificultad para la extubacin, estas pruebas pueden valorar el impulso ventilatorio y predecir el xito de la desconexin del ventilador.
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
94 8. BIBLIOGRAFA 1. Euler C von. Brain stem mechanisms for generation and control of breathing pattern. In: Cherniack NS, Widdicombe JG, eds. Handbook of Physiology; Control of Breathing. Vol. 2, Part 2, Section 3. Bethesda, American Physiology Society, 1986; pp.1-67. 2. Coleridge HM, Coleridge JCG. Reflexes evoked from tracheobronchial tree and lungs. In: Cherniack NS, Widdicombe JG, eds. Handbook of Physiology; Control of Breathing. Vol. 2, Part 2, Section 3. Bethesda, American Physiology Society, 1986; pp. 395-430. 3. Bruce EN, Cherniack NS. Central chemoreceptores. J Appl Physiol 1987; 62: 389-402. 4. Easton Pa, Sllykerman LJ, Anthoninsen NR. Ventilatory response to sustained hyocia in normal aduelts. J Appl Physiol 1986; 61: 906-911. 5. Assessment of respiratory control in humans; editorial and workshop. Am Rev -Respir Dis : 177-181, 363-365, 541-544, 713, 715-716, 883-887, 1977 6. Rebuck AS, campbel EJM. A clinical method for assessing the ventilatory response to hypoxia. Am Rev Respir Dis 1974; 109: 345-50 7. Read DJC: A clinical method for assessing the ventilatory response to carbon dioxide. Australas Ann Med 16. 20-32, 1967 8. Milledge JS, Minty KB, Duncalf D. On-line assessment of ventilatory response to carbon dioxide. J Appl Physiol 1974; 37: 596-9 9. Calverley PMA. Control of breathing. En Hughes JMB, Pride NB. Lung Functions Tests. WB Saunders. London 1999. Pags. 107-120. 10. Whitelaw WA, Derenne J, Milic-Emili J: Occlusion pressure as a measure of respiratory center output in conscious man. Respir Physiol 23: 181-199, 1975 11. Whitelaw WA, Derenne JP. Airway occlusion pressure. J Appl Physiol 1993; 74:14751483. 12. Whitelaw WA, Derenne JP, Milic-Emili J. Occlusion pressure as a measure of respiratory center output in conscious man. Respir Physiol. 1975; 23: 181-99. 13. Cherniak NS, Altose MD. Respiratory response to loading. In: Hornbein TK, ed. The Regulation of Breathing Part II. New York, Marcel Dekker Inc, 1981: pp. 905-964. 14. Cherniak NS, Millic-Emili J. Mechanical aspects of loaded breathing. In: Roussos C, Macklem PT, eds. The Thorax . New York, Marcel Dekker, 1985; pp. 751-786.
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
95 15. Clague JE, Carter J, Pearson MG, Calverley PM. Effort sensation, chemoresponsiveness and breathing pattern during inspiratory resistive loading. J Appl Physiol 1992; 73:440-445. 16. Grassi B, Ferretti G, Costa M, Ferrigno M, Panzacchi A, Lundgren CE, Marconi C, Cerretelli P. Ventilatory responses to hypercapnia and hypoxia in elite breath-hold divers. Respir Physiol. 1994; 97: 323-32. 17. Stanley NN, atose MD, Kelsen SG, Ward CF, Cherniack NS. Changing effect of lung volume on respiratory drive in man. J Appl Physiol 1975; 38:68-773. 18. Whitelaw WA, Derenne J, noble S, McBride B. Similarities between behaviour of respiratory muscles in breath-holding. Respir Physiol 1988; 72. 151-161. 19. Cherniack RM, Snidal DP. The effect of obstruction to breathing on the ventilatory response to CO2. J Clin Invest. 1956; 35: 1286-90 20. Brodovsky D, McDonell JA, Cherniack RM. The respiratory response to carbon dioxide in health and in emphysema. J Clin Invest. 1960; 39: 724-9 21. Calverley PMA. Ventilatory control and dysnea. En Calverley PMA, Pride NB. Chronic Obstructive Pulmonary Disease. Chapman-Hall. London 1995. Pags: 205-242. 22. Park SS. Factors responsible for carbon dioxide retention in chronic obstructive lung disease. Am Rev Respir Dis. 1965; 92: 245-54. 23. Lourenco RV, Miranda JM. Drive and performance of the ventilatory apparatus in chronic obstructive lung disease. N Engl J Med. 1968; 279: 53-9 24. Gorini M, Spinelli A, Ginanni R, Duranti R, Gigliotti F, Scano G. Neural respiratory drive and neuromuscular coupling in patients with chronic obstructive pulmonary disease (COPD). Chest. 1990; 98: 1179-86. 25. Gautier H, Remmers JE, Bartlett D Jr. Control of the duration of expiration. Respir Physiol. 1973; 18: 205-21. 26. Remmers JE. Analysis of ventilatory response. Chest. 1976; 70 (Suppl): 134-7 27. Cherniack NS, Dempsey J, Fencl V, Fitzgerald RS, Lourenco RV, Rebuck AS, Rigg J, Severinghaus JW, Weil JW, Whitelaw WA, Zwilich CW. Methods of measurement of ventilatory responses to hypoxia and hypercapnia. Am Rev Respir Dis. 1977; 115: 177-81 28. Aubier M, Murciano d, Milic-Emili J, et al. Effects of the administration of O2 on ventilation and blood gases in patients with chronic obstructive pulmonary disease during acute respiratory failure. Am Rev Respir Dis. 1980; 122: 747-754
EVALUACION DEL CONTROL QUIMICO Y MECANICO DE LA RESPIRACION
96 29. Robinson TD, Freiberg DB, Regnis JA, Young IH. The role of hypoventilation and ventilation-perfusion redistribution in oxygen-induced hypercapnia during acute exacerbation of chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med 2000; 161: 1524-1529 30. Shea SA, Andres LE, Shannon DC, Banzett RB. Ventilatory responses to exercise in humans lacking ventilatory chemosensitivity. J Physiol (lond) 1993; 468: 623-640 31. American Thoracic Society. Idiopathic congenital central hypoventilation syndrome: diagnosis and manegement. Am. J. Respir. Crit. Care Med 1999; 16: 368-373. 32. Severinghaus, J.W. and Mittchell, R,A.: Ondines curse-failure of respiratory center automaticity while awake. Clin.Res. 1962; 10:122. 33. Debra E. Weese-Mayer, MD,Chair et al. Idiopathic Congenital Central Hipoventilation Sndrome. Am. J. Respir. Crit. Care Med. 1999; 160: 368-373. 34. Cherniak NS, Lomgobardo G. Periodic breathing during sleep. En Sleep and Breathing (2nd ed). Saunders NA y Sullivan CE Eds. Marcel Dekker, New York 1994: 157-190. 35. Ingvar DH, Bulow KB. Respiratory regulation in sleep. Ann N Y Acad Sci. 1963 Jun 24;109:870-81. 36. Cherniack NS, Longobardo GS. Cheyne-Stokes breathing. An instability in physiologic control. N Engl J Med. 1973;288:952-7 37. Gothe B, Altose MD, Goldman MD, Cherniak NS. Effect of quiet sleep on resting and CO2-stimulated breathing in humans. J Appl Physiol 1982; 50: 724-730, 38. Gleeson K, Zwillich CW, White DP. The influence of increasing ventilatory effort on arousal from sleep. Am Rev Respir Dis 1990; 142: 295-300 39. ATS/ERS stament on respiratory muscle testing. Am J Respir Crit Care Med 2002; 166: 518-624. 40. Bates DV. Altered physiologic states and associated syndromes. En Respiratory Function in disease (3rd ed). Bates DV ed. Saunders, Phyladelphia 1989: 81-105.
CAPTULO 5
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
LUIS PUENTE MAESTU Y FELIPE VILLAR LVAREZ
Hospital General Universitario Gregorio Maran Servicio de Neumologa c/ Doctor Ezquerdo 46 28007 Madrid Tfno: (+34) 91 586 8336 Fax: (+34) 91 586 8018 Email: lpuente@separ.es
PALABRAS CLAVE: Ventilacin, quimiorreceptores, ejercicio, produccin de anhdrido carbnico, equilibrio cido-base, senos carotdeos, orden motora, patrn respiratorio, lmites ventilatorios.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
100 NDICE DE CONTENIDOS: 1. Introduccin 2. Determinantes de la ventilacin durante el ejercicio 3. Patrones de Respuesta Ventilatoria al Ejercicio 3.1. Fase uno: Fase dinmica precoz 3.2. Fase dos: Fase dinmica tarda 3.3. Fase 3: Fase de estado estable 3.4. Acidosis lctica 4. Lmites ventilatorios a la capacidad de esfuerzo 4.1. Limitacin mecnica 4.2. Fatiga muscular 4.3. Demandas metablicas 5. Conclusin 6. Bibliografa
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
101 1. INTRODUCCIN El pulmn es la puerta de entrada y de salida de los gases involucrados en la respiracin celular o sea, a travs del pulmn debe pasar todo el oxgeno que necesiten las mitocondrias para mantener su metabolismo energtico y tambin se debe ventilar el organismo del anhdrido carbnico producido por estas. La actividad fsica es un poderoso estmulo metablico y para que pueda proseguir la ventilacin ha de aumentar en la proporcin necesaria para atender a las necesidades celulares, de hecho, al llevar a cabo actividades ligeras o moderadas no se producen cambios apreciables con respecto al reposo ni en las presin parcial de oxgeno ni en la de anhdrido carbnico en la sangre arterial (1) lo que implica que el intercambio de estos gases en los pulmones se ajusta exactamente a la cantidad que absorben y producen los tejidos. En las actividades ms intensas, en las que se acumula cido lctico en cantidades significativas, los pulmones eliminan anhdrido carbnico en exceso para compensar la acidosis metablica, aumentado aun ms las demandas ventilatorias. La estrecha concordancia entre la ventilacin por un lado y la demanda metablica y el equilibrio cido base por otro, proporciona un criterio para definir lo que es un respuesta adecuada y es el fundamento de este captulo.
2. DETERMINANTES DE LA VENTILACIN DURANTE EL EJERCICIO Durante la contraccin muscular la respiracin pulmonar debe aumentar en proporcin al metabolismo pare preservar el microambiente fisicoqumico de las clulas musculares en unas condiciones que permitan seguir cumpliendo sus funciones, para conseguirlo una cadena de sistemas engranados (y que empieza en lo pulmones) deben tener suficiente la capacidad para trasportar el oxgeno de la boca a la mitocondria en las cantidades necesarias (figura 1).Para comprender la magnitud de dicho transporte es conveniente empezar por considerar los requerimientos metablicos de los msculos. El msculo esqueltico se puede considerar como una mquina que convierte la energa qumica de los alimentos ingeridos en energa mecnica y trabajo. Como otras clulas el msculo no utiliza directamente la energa qumica de los sustratos usados como combustibles si no que utiliza un medio de pago energtico intracelular: La adenosina trifosfato (ATP) cuyo enlace fosfato terminal libera una gran cantidad de energa cuando se hidroliza al deshacerse los puentes actinamiosina accionando as el mecanismo contrctil. La principal fuente de produccin sostenida de ATP en las fibras musculares es el catabolismo de los carbohidratos y grasas, ya que en condiciones normales las protenas juegan un papel insignificante como sustrato durante el ejercicio. En la tabla 1 podemos observar los requerimientos estequiomtricos de utilizacin de oxgeno en la oxidacin hasta anhdrido carbnico y agua de los carbohidratos y de la grasas. La tabla 1 pone de relieve que la eficiencia relativa de la utilizacin de uno u otro sustrato depende del sistema que consideremos. Desde un punto de vista termodinmico el aprovechamiento de la energa qumica (p) es similar catabolizando grasas o carbohidratos, sin embargo, desde el punto de vista de almacenamiento, las grasas tienen una ventaja
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
102
Figura 1. Representacin esquemtica de los sistemas que se engranan para que con el consumo de oxgeno (VO2 y la produccin de anhdrido carbnico (VCO2) pulmonares se ajuste al consumo de oxgeno (QO2) y la produccin de anhdrido carbnico metablicos (QCO2 ).
~P: RQ VO2 VCO2 O 2
Carbohidratos (Glucosa)
~P: CO2
O2: ~P
CO2: Kcal /l KCal/ l ATP/ (O2) (CO2) mol Kcal/g ~P
0.17
0.17
5.28
5.28
36/37
3.81
53%
AGL 0.7 (Palmitato)
0.7
5,65
8.13
0.18
0.12
4.54
6.54
130
9.1
56%
Tabla 1. Relaciones entre energa, intercambio de gases y sustratos. RQ: cociente respiratorio celular; VO2 y VCO2 captacin de oxgeno y produccin de anhdrido carbnico en l min-1. AGL. cidos grasos libres; ~P:O2 y ~P:CO2 ATP producido por mol de oxgeno consumido y anhdrido carbnico producido respectivamente. O2:~ P y CO2:~P: Moles de oxgeno consumido o produciodos para producir un mol de ATP. Kcal/l: Kilocaloras producidas por el consumo o la producicn de un litro de oxgeno o anhdrido carbnico respectivamente. Kcal /g. Kilocaloras obtenida de un gramo de azcar o de cido palmtico. p: energa producida en ATP con respecto a la energa libre del sustrato.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
103 significativa con respecto a los hidratos de carbono, con una produccin cercana a 9 kcal/g comparada con solo 4 kcal/g de los carbohidratos. Tambin se puede ver en la tabla 1 que hay marcadas diferencias segn el sustrato en las necesidades de intercambio gaseoso, as la proporcin de ATP producida por mol de oxgeno (~P:O2) es algo mayor usando carbohidratos que cidos grasos y en consecuencia las necesidades de oxgeno para una necesidad dada de sntesis de ATP es aproximadamente un 6% menor que usando grasas, es decir, desde el punto de vista de los mecanismos de transporte de oxgeno, los hidratos de carbono son el combustible ms eficiente. En contraste a la elevada ~P:O2 de los cidos grasos, la produccin de CO2 por mol de ATP (CO2:~P) es un 30% menor que con la utilizacin de carbohidratos, y por tanto los cidos grasos son el combustible ms eficiente con respecto a la produccin de CO2. Estas diferencias en el intercambio gaseoso segn el sustrato son relevantes para los sistemas fisiolgicos encargados de transportar oxgeno y anhdrido carbnico. Por ejemplo, dado que hay una estrecha relacin entre el gasto cardiaco (Q) y el consumo de oxgeno (VO2) durante el ejercicio, el uso de carbohidratos exigir menos al sistema cardiocoruclatorio (1), por el contrario, dado que la ventilacin (VE) para un determinado esfuerzo fsico est ms relacionada con la produccin de CO2 (VCO2), el uso de una proporcin mayor de cidos grasos es menos exigente para el sistema venitalatorio durante el ejercicio (2). La contribucin proporcional de carbohidratos (c) al intercambio de energa puede calcularse en situaciones de estado-estable a partir del cociente respiratorio (RQ; p.e., la produccin de CO2 generado metablicamente, dividido por el O2 consumido): c = (RQ 0.7)/0.3 (1)
El cociente RQ se refiere por tanto a los gases consumidos y producidos por la clula y slo podemos medirlos en la boca en situaciones estables, cuando ambos coinciden. Por ello RQ se distingue del cociente VO2/VCO2 medido en la boca y que llamamos simplemente R. Es importante darse cuenta de que R puede diferir marcadamente de RQ en condiciones en las que las que el gas disuelto en el organismo (sobre todo los depsitos de CO2) estn cambiando. Este fenmeno es aparente por ejemplo al comienzo del ejercicio como se muestra en la figura 1. Podemos ver que R permanece relativamente constante durante unos 15-20 s tras comienzo del ejercicio (mas abajo se discute este intervalo que se denomina fase 1), lo que implica que mientras la sangre procedente de los msculos llegue a los pulmones, la sangre venosa que pasa por ellos tiene los mismos niveles de O2 y CO2 que en reposo. Casaburi et al. (3) han medido los contenidos venosos mixtos de O2 y CO2 directamente en arteria pulmonar y han encontrado que cambian antes de los esperado (tiempo de circulacin de la sangre desde los msculos) cuando se parte de reposo, pero no cuando se aumenta la potencia desde un ejercicio previo (4) posiblemente por los efectos del tono simptico en la aportacin al pool venoso central por los distintos territorios venosos. Tras este breve periodo inicial, en el que R permanece relativamente estable, se observa un descenso transitorio reflejo de que la respuesta de la VCO2 es apreciablemente ms lenta que la de VO2. Esta fase transitoria ha sido denominada fase 2. La diferencia en el curso temporal de las respuestas de VO2 y VCO2 se debe a que una parte del CO2 producido por
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
104 el metabolismo se acumula (disuelto) en el msculo y por tanto no todo el CO2 originado en los msculos activos pasa inmediatamente a la sangre venosa y llega al pulmn. Durante esta fase del ejercicio R es obviamente menor que RQ (no se puede determinar directamente como cambia RQ en esta fase 2). Durante el periodo de transicin ejercicio-reposo el CO2 previamente almacenado en los tejidos se descarga lo que provoca un caracterstico aumento de R por encima del RQ. El otro mecanismo importante por el que R difiere de RQ ocurre cuando se consume bicarbonato (HCO3) tamponando en acido lctico. En el ejercicio de alta intensidad se produce una acidosis lctica, desciende el bicarbonato y se libera CO2 a una tasa aproximadamente 2.5 veces mayor que cuando no hay acumulacin de cido lctico (5), es decir, la energa producida anaerbicamente produce indirectamente 2.5 veces mas CO2 que el uso de hidratos de carbono. Aproximadamente el 90% del tampn lo proporciona le bicarbonato disuelto (6)(7). El resultado es un descenso de ~ 22 mEq de bicarbonato por cada 25 mEq de aumento en la concentracin lactato. Puesto que la oxidacin de los hidratos de carbono termina en cido lctico y no en CO2 y agua (lo que hubiese producido 6 molculas de CO2 que deben ser restadas del exceso de CO2 originado al taponar el cido lctico) el aumento neto en la tasa de produccin de CO2 es : 24.6 mEq (lactato) x 0.9 22mEq (HCO3) 22mM CO2 (2) (3)
22mM CO2 anaerbico 6mM CO2 aerbico = 16mM de CO2 (produccin neta)
La consecuencia por tanto del metabolismo anaerobio es un gran aumento de la VCO2 que se ve muy claramente durante las pruebas de esfuerzo y es la base de la estimacin no invasiva del umbral de lactato (figura 3). Este exceso de CO2 debida al tamponamiento del bicarbonato es una exigencia aadida para la funcin intercambio gaseoso pulmonar impuesta por la formacin de cido lctico, pero adems los pulmones tambin intervienen en la homeostasis cido-base y para frenar la caida pH arterial (pHa), la VE tiene que incrementarse an ms, como podemos deducir de la reformulacin de la ecuacin de Herderson-Hasselbach: pHa = pK + log {(HCO3)a/(0.03 x PaCO2)} (4)
donde el pK del cido lctico tiene normalmente un valor de 6.1. Pero como PaCO2 = 863 x VCO2 (STPD)/ VA (BTPS) y VA = VE (1 VD/VT) (6) (5)
donde VA es la ventilacin alveolar y VD/VT el espacio muerto fisiolgico, entonces: pHa = pK + log {((HCO3)a/25.6) x (VE /VCO2) x (1 VD/VT)} (7)
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
105
Figura 2. Curso temporal del intercambio de gases en la boca. Obsrvese como tras un breve periodo en que permanece constante al principio del ejercicio, R disminuye por la mayor lentitud de la repuesta de la produccin de anhdrido carbnico (VCO2 ) con respecto al consumo oxgeno(VO2 ). Esto se debe a que la capacidad de almacenamiento de CO2 por le organismo es mayor que la de O2 y parte del CO2 producido por el metabolismo es almacenado. En esta fase inestable R (cociente respiratorio) no es un reflejo de RQ (cociente metablico).
Esta frmula pone de manifiesto que para detener la bajada del pH cuando la capacidad de tamponamieto del bicarbonato se agota, la VE debe aumentar ms que la VCO2 en una proporcin que depende de la eficiencia del intercambio gaseoso, definida por el trmino (1 VD/VT). En resumen hemos visto a lo largo de esta seccin que la ventilacin necesaria para que conseguir ajustar el intercambio de gases pulmonar a la respiracin celular depender del sustrato utilizado, de la capacidad del organismo para el almacenamiento CO2 a corto plazo, de la eficiencia de la ventilacin, de si se genera o no acidosis lctica y de la capacidad tampn del organismo. Adems de todo esto la ventilacin se vera influida por el set point de la presin arterial de anhdrido carbnico (PaCO2), pues para eliminar un volumen dado de CO2 por minuto se necesitar mas menos ventilacin dependiendo de la PaCO2 como podemos ver en la figura 4.
3. PATRONES DE RESPUESTA VENTILATORIA AL EJERCICIO En la seccin previa consideraremos los factores que constituyen el estmulo para la ventilacin en el ejercicio; en esta parte nos centraremos en el conocimiento actual de cmo se lleva a cabo este ajuste.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
106
Figura 3. Respuestas ventilatoria y del intercambio gaseosa a una prueba progresiva en cicloe-ergmetro. Ntese la aceleracin de la produccin de anhdrido carbnico (VCO2 ) con respecto al consumo de oxgeno (VO2 ) y al aumento de los equivalentes ventilatorios de oxgeno y anhdrido carbnico (EqO2 y EqCO2 respectivamente) que permiten estimar de forma no invasora el mural de acidosis.
Se cree que al igual que con otras respuestas fisiolgicas los determinantes de la ventilacin varan en distintos momentos a partir del comienzo del ejercicio de acuerdo con unas fases (figura 5) que conceptualmente son las siguientes: 3.1. Fase uno: Fase dinmica precoz Esta fase corresponde a los primeros 10-20 segundos del ejercicio (figura 5) y se corresponde con el tiempo transcurrido desde el momento en que se activan los msculos y la llegada a los pulmones de la sangre procedente de las extremidades. Krogh y Lindhard (8) fueron los primeros en documentar que en la VE aumentaba virtualmente en sincrona con el comienzo del ejercicio muscular. Pensaron que el mecanismo deba guardar relacin con el aumento del gasto cardaco (Q) pulmonar ya que en esta fase el VO2 tambin aumenta bruscamente antes de que pueda llegar la sangre venosa procedente de los msculos activos. Esta observacin se ha confirmado en numerosas ocasiones si bien se ha podido profundizar en algunos detalles de la respuesta ventilatoria en esta fase precoz. Se ha observado, por ejemplo, que al comienzo del ejercicio (figura 2) tanto el gasto cardaco como la VE aumentan de forma rpida y en una proporcin bastante pareja (9),(10),(11) y como consecuencia ni la presin tele-espiratoria de CO2 (PETCO2) ni R no sufren cambios apreciables. (Algunos
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
107
Figura 4. Factores que influyen en las necesidades ventilatorias para una carga metablica dada. El grfico representa las notables diferencias de ventilacin que pueden necesitarse para responder a una determinada necesidad metablica, definida por el consumo de oxgeno (VO2 ) dependiendo de del conciente respiratiorio (R), la presin arterial de CO2 (PaCO2 ) y el espacio muerto fisiolgico (VD/VT).
Ventilacin
Tiempo
Figura 5. Fases de la respuesta ventilatoria al ejercicio. En la respuesta ventilatoria, como en las otras respuestas fisiolgicas al ejercicio, se pueden considerar 3 fases, una fase inicial de respuesta dinmica precoz, que dura unos 20 s, el tiempo de circulacin de la sangre venosa desde los msculos hasta los pulmones, una fase dinmica tarda, en la que la ventilacin crece exponencialmente hasta alcanzar la respuesta mxima y un fase de estado estable en la que tras alcanzar la respuesta mxima necesaria se mantiene mientras dure el ejercicio.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
108 resultados experimentales parecen indicar que de hecho se produce una discretsima hiperventilacin(12)(13)). Por su velocidad de instauracin tradicionalmente se ha considerado esta respuesta tan rpida de la VE como de origen neurognico, bien por aferencias respiratorias procedentes de extremidades en ejercicio (14)(15)(16)(17)(18) , impulsos procendentes de la propia corteza cerebral (8) o del rea motora subtalmica (19)(20). La estimulacin de las pequeas fibras mielinizadas y no mielinizadas (grupos III y IV) de los msculos en animales produce una respuesta ventilatoria y cardiovasculares que se elimina al bloquear las fibras (21) y se ha visto que estas fibras se proyectan en el hipotlamo donde podran influir en la regin motora subtalmica y, as, en las respuestas cardiorrespiratorias (19)(20)(22). En humanos, sin embargo, no se modifica sensiblemente la respuesta ventilatoria al bloquear las aferencias de las extremidades o al estimular pasivamente los msculos en sujetos con seccin medular (23)(24). Aunque se considera que el mecanismo principal regulatorio de la hiperpnea del ejercicio es de origen central como un componente asociado de la orden motora cortical que activa los msculos perifricos, hay una serie de observaciones en esta fase difciles de explicar, como el que la hiperpnea inicial sea independiente de la magnitud del impuso central (potencia a generar por los msculos activos) (15)(25), es decir no parece que la respuesta sea directamente proporcional a la intensidad del estmulo o al nmero de neuronas motoras activadas. Adems cuando se aumenta la potencia partiendo de un ejercicio la hiperpnea precoz es mnima o inexistente a pesar de la mayor activacin central (26)(27)(28). Finalmente se ha visto que la hiperpnea precoz tambin se reduce significativamente o llega abolirse cuando la transicin reposo-ejercicio se hace en posicin supina (29)(30) y resulta sorprendente qu el mero hecho de tumbarse sea capaz de hacer desaparecer un inactivar un elemento bsico del estmulo respiratorio. Otra teora alternativa de la hiperpnea precoz es que la respuesta ventilatoria est directamente vinculada a la respuesta cardiaca (hipernea cardiodinmica) (31), mediada por estmulos originados en el ventrculo derecho (32) (33). En apoyo de esta teora estn la respiracin de Cheynes-Stokes en las que las alteraciones de la presin venosa o auricular conduce a la estimulacin ventilatoria (34)(35)(36)(37), sin embargo experimentos en humanos con marcapasos implantados a los que se les induca artificialmente aumentos del gasto cardiaco incrementando la frecuencia del marcapasos no se vio que la VE cambiase significativamente en los 20 s siguientes a cambios de frecuencia de 50 a 80 latidos min-1 a pesar de que PETCO2 subi 2.5 mmHg de media, la presin tele-espiratoria de oxgeno (PETO2) cay aproximadamente 8 mmHg y R cerca de 0,1. Dado que al aumentar la frecuencia se observaron aumentos en laVO2 y la VCO2 secundarios al aumento del gasto cardaco, pero no se vio hiperpnea es poco probable que le mecanismo caridodinmico sea fundamental (38). Adems en los transplantados pulmonares (39)(40) no se observa reduccin de la magnitud o de la rapidez de comienzo de la respuesta hiperneica al ejercicio a pesar de la denervacin cardiaca. En resumen, el ajuste tanto en el curso temporal como en la magnitud de la respuesta circulatoria y cardiaca en la fase I del ejercicio sugiere una relacin entre ambas respuestas pero ninguno de los mecanismos propuestos es capaz de explicar todas las observaciones experimentales, lo que da pie a la posibilidad de que ambos aumentos se deban a una orden central y que la similitud sea meramente accidental.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
109 3.2. Fase dos: Fase dinmica tarda En esta fase la respuesta de todas las variables de intercambio de gases se caracteriza por un crecimiento que se puede definir muy bien por una funcin exponencial (figura 5). Se observa un descenso transitorio de R (figura 1) como resultado de la cintica mas lenta de VCO2 comparada con VO2. Tradicionalmente se ha considerado que el mecanismo que regula la ventilacin en esta fase es humoral, pues la transicin empieza justo en el momento en que comienza a llegar a los pulmones la sangre procedente de los msculos activos y la elevacin de la ventilacin es lenta, sin embargo, algunos autores cuestionan que la lentitud de la respuesta necesariamente excluya un control nervioso (41). Hay bastante evidencia que confirma que la magnitud de la respuesta ventilatoria est estrechamente ligada con el intercambio pulmonar de CO2 y no con el de O2 o la produccin tisular de CO2 (6)(42)(43) (44). En estudios en los que la potencia se cambia de forma sinusoidal (oscilaciones de intensidad con un periodo fijo) (27)(45)(46)(47)(48), variando la amplitud y velocidad de los cambios se ha observado que la respuesta ventilatoria est desfasada de los cambios de potencia y de VO2, pero sigue estrechamente los cambios de VCO2. Resulta difcil explicar esta observacin si el mecanismo de control de los cambios lentos de la ventilacin fuese central (es decir ligado a la orden motora que activa los msculos que realizan el ejercicio) pues implicara que el impulso ventilatorio siguiera un curso temporal diferente que el impulso motor a las piernas a pesar de proceder del mismo origen. Otro modelo experimenta til para analizar esta fase es la respuesta ventilatoria a una potencia impulso (una potencia supra-mxima mantenida brevemente) por ejemplo 400W durante 10 s. La respuesta esperada para un sistema con dos componentes uno rpido (como el explicado ms arriba en la fase I) y otro ms lento, como se piensa que ocurre en la fase 2 del ejercicio, se muestra en la figura 6. El primer pico estara dominada por el primer componente que cuando empieza a decaer viene seguido por el segundo componente de la respuesta que aparece cuando ya no se est realizando ejercicio y por lo tanto han cesado el impuso central y las aferencias musculares relacionadas con el movimiento. Este comportamiento sugiere que no es necesario estar haciendo ejercicio para que se produzca una respuesta ventilatoria. Por tanto la respuesta a variaciones sinusoidales de la potencia y a potencias impulso demuestra que la VE est estrechamente relacionada con el intercambio pulmonar de CO2 y no con la utilizacin de O2 o la actividad muscular. Se ha especulado que los cambios en la VE podran ser los responsables de una mayor eliminacin de CO2 y no al revs (49), pero esta posibilidad es remota ya que los estudios de cambio de potencia sinusoidal demuestran que existe un desfase sistemtico entre los cambios de la VCO2 y la respuesta de VE siendo esta ltima algo mas lenta (aprox. 10%) (45) (46). La consecuencia de este desfase sera una pequea hipercapnia transitoria en la fase 2 que ha sido observada, aunque es de magnitud muy pequea (50)(51)(52) comparada con la hipoxemia transitoria que tambin se produce pues el desfase entre ventilacin y VO2 es mucho mayor ( del orden del 50-70%) (53) (54). Se sabe que el control del a ventilacin en esta fase depende en una pequea parte del control mediado por los cuerpo carotdeos. Estudios en humanos en los que la seal de los qui-
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
110
Amplitud
Tiempo (s)
Figura 6. Respuesta a un estmulo impulso de un modelo de dos componentes. Respuesta idealizada de un sistema de dos componentes a un estmulo de impuso. Obsrvese que el primer componente rpido produce una respuesta que desaparece tambin rpido antes de que haya tenido lugar el segundo componente de la respuesta que aparece (segundo pico) cuando ya ha terminado el primero y desparece mas lentamente.
miorreceptores perifricos se aumenta mediante hipoxia (55) o acidosis metablica con cloruro de amonio oral (56) demuestran que la respuesta ventilatoria es ms rpida y si por el contrario se reduce la quimiosensibilidad carotdea con hiperoxia (55) (57) o alcalosis metablica inducida por la ingesta de bicarbonato sdico (56), por la infusin intravenosa de dopamina (58) y en sujetos con reseccin quirrgica de ambos cuerpos carotdeos (59) la respuesta ventilatoria se retarda apreciablemente. Por tanto los cuerpos carotdeos parecen ser elementos importantes, aunque no los nicos como veremos mas adelante, en la modulacin de las respuesta de la VE a la VCO2. 3.3. Fase 3: Fase de estado estable La respuesta ventilatoria del estado estable es el resultado de la mxima respuesta de la suma de los mecanismos operativos en la fase 1 y 2, pero no esta claro el mecanismo por el que se consigue la estrecha regulacin de la PaCO2 en valores prximos a los de reposo*, una caracterstica que se ven en la mayora de los sujetos. Algunos autores creen que la estabilidad de la PaCO2 no es ms que un fenmeno espurio y no se debe a una autntica regulacin de la ventilacin a la produccin metablica de CO2 (60)(61). Pero la opinin mayoritaria es que hay una regulacin ligada al CO2, pero independiente de la presin arterial media de anhdrido carbnico (ya que esta no varia como se ha dicho). Los mecanismos propuestos como reguladores de la VE son: 1.- Presin de anhdrido carbnico en la sangre venosa mixta. Es dudoso que este sea un estmulo ventilatorio pues la sangre cargada de CO2 infundida en el retorno venoso no
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
111 estimula la ventilacin (62) hasta que no alcanza los quimiorreceptores arteriales (63). Adems los sujetos que han sufrido un transplante cardiopulmonar (i.e denervacin del corazn y de los pulmones) tienen ventilaciones adecuadas durante el ejercicio (40). 2.- Difusin de CO2 a travs del pulmn (desde los vasos pulmonares a los alvolos) (64). Como se dijo antes el control respiratorio no parece verse afectado paro la vagotoma ni en animales ni en humanos con transplante cardiopulmonar. 3.- Sensibilidad a las oscilaciones de CO2 en la sangre (65). Algn quimiorreceptor de los conocidos o an no identificado sera sensible la amplitud de las oscilaciones inspiratorias-espiratorias de PaCO2 y pH, aunque la media permaneciera constante (66). Se han propuesto varias caractersticas de estas oscilaciones de PCO2 como mecanismo de control de la VE que incluyen: 1.- La amplitud de la oscilacin. 2.- La velocidad de cambio de la seal de (H+). 3.- El nmero de picos por unidad de tiempo que pasa por los cuerpos carotdeos. 4.- El desfase entre la seal oscilante que llega a los cuerpos carotdeos y el ciclo respiratorio. Hay dos observaciones empricas que hay que tener en cuenta al considerar estos mecanismos. Primero, el componente de la hiperpnea durante el ejercicio en el que no se produce acidosis lctica que depende de los senos carotdeos es del 0-20% dependiendo de los sujetos y por tanto solo podran explicar parte de la respuesta ventilatoria. Segundo, en humanos la amplitud de las oscilaciones del pHa se amortigua si la frecuencia respiratoria es mayor de 20 min-1 (67) y sin embargo esta frecuencia se alcanza en muchas ocasiones sin que disminuya la ventilacin. 4- Transporte venoso de CO2: Cargando o descargando la sangre venosa de CO2 producen cambios en la ventilacin idnticos a los que produce la inhalacin de CO2 y que son de una magnitud menor a los cambios en la ventilacin que ocurren en ejercicio (68) (69)(70)(71), sin embargo la reduccin del retorno venoso (derivando la sangre a travs de un baypass veno-arterial parciales con intercambiadores gaseosos extracorpreos) si disminuye la VE en proporcin a la cantidad de CO2 intercambiada por el sistema extracorpreo (72)(73)(74) es decir la ventilacin se reduce en proporcin a la reduccin de la VCO2 pulmonar aunque el ejercicio contine y adems al cesar la derivacin la VE aumenta otra vez hasta volver a sus niveles de control. Stremel et al. (72) han observado que la VE en reposo descenda al abrir la derivacin veno-arterial a pesar de que la presin arterial de CO2 aumentase. Los receptores involucrados en el ajuste de la ventilacin a la VCO2 pulmonar secundarias a los cambios en el gasto cardiaco que pasa por los pulmones no estn claros. Se ha sugerido que puedan ser los receptores de distensin pulmonar, cuya seal es sensible al CO2 (33) (73), pero de nuevo los datos en animales
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
112 vagotimizados o en humanos con transplantares pulmonares ponen en cuestin que el estmulo proceda del mismo pulmn. 5- Potasio: El potasio se libera del interior de las clulas musculares durante su contraccin y se ha propuesto como un regulador de la ventilacin durante el ejercicio (75), basndose en dos hallazgos: 1) la correlacin observada entre la concentracin de potasio en la sangre venosa y la ventilacin y 2) la demostracin que el potasio es capaz de activar las terminaciones nerviosas tipo III y IV musculares (cuya estimulacin se ha demostrado que inducir hiperpnea) (17). Los cambios en la concentracin de potasio tambin estimulan los cuerpos carotideos (76) (77), aunque como vimos mas arriba los cuerpos carotideos juegan un papel no mayor del 20% de la regulacin ventilatoria. Podemos por tanto resumir diciendo que parece que el intercambio de CO2 en el pulmn es el determinante principal de la magnitud de la hiperpnea en ejercicio, pero influida en parte por otros factores como la PaCO2. De los mecanismos de control propuestos no hay ninguno convincente que justifique la relacin entre la produccin del CO2 y la VE. 3.4. Acidosis lctica Por encima del umbral de acidosis lctica, las respuesta de VE al ejercicio se vuelven ms complejas y deja de ser lineal. La acidosis metablica produce un incremento del impulso ventilatorio adicional para intentar conseguir la compensacin respiratoria del equilibrio cido-base reduciendo la PaCO2. Esta respuesta est sensiblemente disminuida en los sujetos a los que se les ha resecado ambos cuerpos carotdeos (59) (78) y en consecuencia el pH cae ms durante el ejercicio en ellos. La acidemia lctica no es el nico mecanismo que aumenta la VE por encima del umbral del lactato (pero si es el componente dominante). Se sabe que hay otros como por ejemplo el incremento de los niveles de catecolaminas circulantes, que slo aparece a estas potencias y que presumiblemente estimula la VE. El papel de este mecanismo no est claro pues aunque el bloqueo betaadrenrgico reduce la ventilacin notablemente en sujetos con insuficiencia cardiaca (79), no altera la respuesta de VE en sujetos normales (80).La elevacin de la temperatura corporal y de la osmolaridad sangunea secundaria a la acumulacin de cido lctico en los tejidos pueden contribuir tambin a la hiperventilacin.
4. LMITES VENTILATORIOS A LA CAPACIDAD DE ESFUERZO 4.1. Limitacin mecnica En sujetos normales existe reserva funcional ventilatoria de sobra y el ejercicio se detiene cuando todava queda entre el 25% y el 40% de la reserva funciona (83) y por tanto no parece que la ventilacin constituya un lmite en estos sujetos, sin embargo en atletas de elite bien entrenados la VE puede aproximarse al 90% y se ha sugerido que pueda llegar a ser un lmite o al menos una traba para aumentar la capacidad aerbica (81). La observacin de que
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
113 en sujetos muy entrenados se produce un aumento del gradiente alveolo-arterial sugiere que la capacidad de los pulmones para intercambiar oxgeno llega a su lmite en estas circunstancias (82) y podra explicar por qu tenemos tanta reserva funcional ventilatoria, que en realidad no sera un redundancia innecesaria, sino la consecuencia de la necesidad aumentar la superficie de difusin partiendo de la estructura de saco rodeado de capilares. El volumen corriente (VT) vara de acuerdo con el tamao corporal y tambin respiracin a respiracin. Durante el ejercicio VT aumenta de forma no lineal y aunque su lmite terico es la capacidad vital, raramente se excede el 70% de la misma (83), mximo que se alcanza aproximadamente al 70% VO2 pico. La frecuencia respiratoria normal en reposo es de 8-12 min-1. Se han podido alcanzar en maniobras forzadas frecuencia respiratoria de 300 min-1 (84), pero en ejercicio excepcionalmente se excedan 60 min-1 y normalmente no se pasa de 40 min-1 (83). Durante la respiracin existen dos elementos de resistencia, la resistencia al flujo que aumenta con la frecuencia respiratoria y la resistencia elstica que aumenta con el (VT), pero los sujetos optan por respirar generalmente con el patrn que minimaza el trabajo respiratorio (85)(86). La relacin entre el tiempo inspiratorio y el espiratorio es 0.8-1 tanto en reposo como en ejercicio. Los flujos inspiratorios y espiratorios aumenta tpicamente durante el ejercicio (figura 7). Aunque la dinmica de la respiracin es diferente durante el ejercicio que durante las maniobras respiratorias forzadas, La mejor estimacin del flujo areo mximo es la curva flujo volumen mxima en reposo (CFVM). Normalmente los flujos inspiratorios y espiratorios generados espontneamente durante el ejercicio son bastante menores que la CFVM (figura 7) en la mayora de los sujetos jvenes o no muy entrenados (87)(88) sin embargo en atletas de elite el flujo areo espiratorio generado espontneamente se
Flujo
Volumen
Flujo
Volumen
Figura 7. Flujos mximos espontneos en reposo y en ejercicio. En esta figura se ven las curvas flujovolumen espontneas generadas en reposo (lnea de puntos mas interna) y en ejercicio mximo (lnea discontinua mas externa) comparada con la curva flujo-volumen mxima (lnea continua mas externa) en un sujeto sano normal ( panel de la izquierda) y en un atleta de elite ( panel de la derecha).
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
114 aproxima al lmite cerca de la capacidad de esfuerzo mxima (aVO2 de 5-6 l min-1 y VE de 150-160 l min-1)(89)(90) y lo mismo ocurre en sujetos de mediana edad o mayores activos fsicamente (91). En sujetos normales jvenes el volumen telespiratorio normalmente baja hasta 0.5-1 l por debajo de la FRC en reposo (figura 7), y en consecuencia aumenta la capacidad inspiratoria) (92)(93). En sujetos mayores tambin disminuye el EELV a niveles moderados pero no a ejercicios intensos por la limitacin al flujo espiratorio (91). 4.2. Fatiga muscular No parece que la ventilacin est limitada por la capacidad de los msculos respiratorios salvo en atletas en los que se ha observado disminucin de la capacidad vital y de la presin transdiafragmtica tras ejercicios intensos (94) (95). 4.3. Demandas metablicas A medida que la ventilacin aumenta los msculos respiratorios necesitan producir ms energa, lo que requiere mas flujo sanguneo que niveles de ventilacin muy elevados puede distraer una proporcin significativa del flujo sanguneo de los msculos activos (96). Desde un punto de vista terico la ventilacin a partir de la cual el requerimiento de oxgeno de los msculos respiratorios se vuelve tan grande que requiere todo incremento adicional de VO2 es de 120-160 l min-1, niveles que solo alcanzan atletas entrenados (97). 5. CONCLUSIN A pesar de la gran cantidad de trabajo realizado los mecanismos de control de la ventilacin durante el ejercicio no estn totalmente aclarado. La teora mas plausible es la actividad neuronal motora que impulsa a los msculos que realizan el ejercicio produce un estmulo simultneo de los msculos respiratorios aumentado la ventilacin. Este estmulo inicial se ajusta luego por una serie de seales procedentes de los msculos perifricos, los quimiorreceptores perifricos y el propio pulmn. Muchos de estos mecanismos parecen redundantes, pero el mecanismo principal de regulacin se desconoce, aunque se sabe que es proporcional a la cantidad anhdrido carbnico que se elimina a travs de los pulmones. Cuando aparece acidosis metablica se establece una compensacin respiratoria fundamentalmente mediada por las seales procedentes del os senos carotdeos. La ventilacin no parece que normalmente sea el factor limitante del ejercicio salvo en atletas de elite y personas de edad entrenadas.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
115 6. BIBLIOGRAFA 1. Wasserman K, Hansen JE, Sue DY, Whipp BJ and Casaburi R. Principles of Exercise Testing (3rd ed.), Lippincott, Williams and Wilkins, Baltimore (1999). 2. Brown SE, Wiener S, Brown RA, Marcarelli PA, Light RW. Exercise performance following a carbohydrate load in chronic airflow obstruction. J Appl Physiol. 1985 Apr;58(4):1340-6. 3. Casaburi R, Daly J, Hansen JE, Effros RM. Abrupt changes in mixed venous blood gas composition after the onset of exercise. J Appl Physiol. 1989 Sep;67(3):1106-12. 4. Casburi, R., Cooper, C., Effros, R.M., Wasserman, K. Time course of mixed venous O2 saturation following various modes of exercise transition. 1989. Fed. Proc. 3:A849. 5. Whipp,B.J., Mahler, M. Dynamics of pulmonary gas exchange during exercise. Pulmonary Gas Exchange. 1980. New York, academic Press, pp 33-96. 6. Wasserman K, Van Kessel AL, Burton GG. Interaction of physiological mechanisms during exercise. J Appl Physiol. 1967 Jan;22(1):71-85. 7. Keull, J., Doll, E., Keppler, D. Oxidative energy supply. Energy Metabolism of Human Muscle. 1972. University Park Press, pp 52-202. 8. Krogh, A., Lindhard, J. The regulation of respiration and circulation during the initial stages of muscular work. J. Physiol.1913; 47:112-136. 9. Loeppke, J.A., Grene, E.R., Hoekenga, D.E. Caprihan, A. Luft U.C. Beat-by-beat stroke volume assessment by pulsed Doppler in upright and supine exercise. J. Appl. Physiol. 1981;50:1173-1182. 10 Miyamoto Y, Hiura T, Tamura T, Nakamura T, Higuchi J, Mikami T. Dynamics of cardiac, respiratory, and metabolic function in men in response to step work load. J Appl Physiol. 1982 May;52(5):1198-208. 11. Cummin AR, Iyawe VI, Mehta N, Saunders KB. Ventilation and cardiac output during the onset of exercise, and during voluntary hyperventilation, in humans. J Physiol. 1986 Jan;370:567-83. 12. Ward SA. The effects of sudden airway hypercapnia on the initiation of exercise hyperpnoea in man. J Physiol. 1979 Nov;296:203-14. 13. Forster HV, Pan LG, Funahashi A. Temporal pattern of arterial CO2 partial pressure during exercise in humans. J Appl Physiol. 1986 Feb;60(2):653-60.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
116 14. Kao, F.F. An experimental study of the patways involved in exercise hyperpnea employing cross-circulation techniques. The Regulation of Human Respiration. 1963. Oxford, Blackwell, pp. 461-502. 15. Dejours, P. Control of respiration in muscular exercise. Handbook of Physiology Respiration, vol I. 1964 . American Physiological Society, pp. 631-648. 16. Tibes U. Reflex inputs to the cardiovascular and respiratory centers from dynamically working canine muscles. Some evidence for involvement of group III or IV nerve fibers. Circ Res. 1977 Sep;41(3):332-41. 17. Kaufman MP, Waldrop TG, Rybicki KJ, Ordway GA, Mitchell JH. Effects of static and rhythmic twitch contractions on the discharge of group III and IV muscle afferents. Cardiovasc Res. 1984 Nov;18(11):663-8. 18. Waldrop TG, Mullins DC, Millhorn DE. Control of respiration by the hypothalamus and by feedback from contracting muscles in cats. Respir Physiol. 1986 Jun;64(3):317-28. 19. Eldridge FL, Millhorn DE, Waldrop TG. Exercise hyperpnea and locomotion: parallel activation from the hypothalamus. Science. 1981 Feb 20;211(4484):844-6. 20. Eldridge FL, Millhorn DE, Kiley JP, Waldrop TG. Stimulation by central command of locomotion, respiration and circulation during exercise. Respir Physiol. 1985 Mar;59(3):313-37. 21. Waldrop TG, Stremel RW. Muscular contraction stimulates posterior hypothalamic neurons. Am J Physiol. 1989 Feb;256(2 Pt 2):R348-56. 22. DiMarco AF, Romaniuk JR, Von Euler C, Yamamoto Y. Immediate changes in ventilation and respiratory pattern associated with onset and cessation of locomotion in the cat. J Physiol. 1983 Oct;343:1-16. 23. Adams L, Frankel H, Garlick J, Guz A, Murphy K, Semple SJ. The role of spinal cord transmission in the ventilatory response to exercise in man. J Physiol. 1984 Oct;355:85-97. 24. Brice AG, Forster HV, Pan LG, Funahashi A, Hoffman MD, Murphy CL, Lowry TF. Is the hyperpnea of muscular contractions critically dependent on spinal afferents? J Appl Physiol. 1988 Jan;64(1):226-33. 25. Jensen JI. Neural ventilatory drive during arm and leg exercise. Scand J Clin Lab Invest. 1972 Apr;29(2):177-84. 26. Linnarsson D. Dynamics of pulmonary gas exchange and heart rate changes at start and end of exercise. Acta Physiol Scand Suppl. 1974;415:1-68.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
117 27. Casaburi R, Whipp BJ, Wasserman K, Koyal SN. Ventilatory and gas exchange responses to cycling with sinusoidally varying pedal rate. J Appl Physiol. 1978 Jan;44(1):97-103. 28. Whipp BJ, Ward SA, Lamarra N, Davis JA, Wasserman K. Parameters of ventilatory and gas exchange dynamics during exercise. J Appl Physiol. 1982 Jun;52(6):1506-13. 29. Karlsson H, Lindborg B, Linnarsson D. Time courses of pulmonary gas exchange and heart rate changes in supine exercise. Acta Physiol Scand. 1975 Nov;95(3):329-40. 30. Weiler-Ravell D, Cooper DM, Whipp BJ, Wasserman K. Control of breathing at the start of exercise as influenced by posture. J Appl Physiol. 1983 Nov;55(5):1460-6. 31. Wasserman K, Whipp BJ, Castagna J. Cardiodynamic hyperpnea: hyperpnea secondary to cardiac output increase. J Appl Physiol. 1974 Apr;36(4):457-64. 32. Jones PW, Huszczuk A, Wasserman K. Cardiac output as a controller of ventilation through changes in right ventricular load. J Appl Physiol. 1982 Jul;53(1):218-24. 33. Huszczuk, A., Whipp, B.J., Oren, A., Shors, E.C., Pokorski, M., Nery, L.E., Wasserman,K. Venous return and ventilatory control. Modelling of Control of Breathing. 1983 New York, Elsevier, pp 78-85. 34. Kostreva DR, Zuperku EJ, Purtock RV, Coon RL, Kampine JP. Sympathetic afferent nerve activity of right heart origin. Am J Physiol. 1975 Oct;229(4):911-5. 35. Uchida Y. Tachypnea after stimulation of afferent cardiac sympathetic nerve fibers. Am J Physiol. 1976 Apr;230(4):1003-7. 36. Lloyd TC Jr. Control of breathing in anesthetized dogs by a left-heart baroreflex. J Appl Physiol. 1986 Dec;61(6):2095-101. 37. Lloyd TC Jr. Effect on breathing of abruptly loading and unloading the canine left heart. J Appl Physiol. 1989 May;66(5):2216-22. 38. Jones PW, French W, Weissman ML, Wasserman K. Ventilatory responses to cardiac output changes in patients with pacemakers. J Appl Physiol. 1981 Nov;51(5):1103-7. 39. Theodore J, Morris AJ, Burke CM, Glanville AR, VanKessel A, Baldwin JC, Stinson EB, Shumway NE, Robin ED. Cardiopulmonary function at maximum tolerable constant work rate exercise following human heart-lung transplantation. Chest. 1987 Sep;92(3):433-9. 40. Banner N, Guz A, Heaton R, Innes JA, Murphy K, Yacoub M. Ventilatory and circulatory responses at the onset of exercise in man following heart or heart-lung transplantation. J Physiol. 1988 May;399:437-49.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
118 41. Eldridge FL. Maintenance of respiration by central neural feedback mechanisms. Fed Proc. 1977 Sep;36(10):2400-4. 42. Jones NL. New tests to assess lung function. Exercise testing in pulmonary evaluation: rationale, methods and the normal respiratory response to exercise. N Engl J Med. 1975 Sep 11;293(11):541-4. 43. Taylor R, Jones NL. The reduction by training of CO2 output during exercise. Eur J Cardiol. 1979 Jan;9(1):53-62. 44. Wasserman DH, Whipp BJ. Coupling of ventilation to pulmonary gas exchange during nonsteady-state work in men. J Appl Physiol. 1983 Feb;54(2):587-93. 45. Casaburi R, Whipp BJ, Wasserman K, Beaver WL, Koyal SN. Ventilatory and gas exchange dynamics in response to sinusoidal work. J Appl Physiol. 1977 Feb;42(2):300-1. 46. Bakker HK, Struikenkamp RS, De Vries GA. Dynamics of ventilation, heart rate, and gas exchange: sinusoidal and impulse work loads in man. J Appl Physiol. 1980 Feb;48(2):289-301. 47. Miyamoto Y, Hiura T, Tamura T, Nakamura T, Higuchi J, Mikami T. Dynamics of cardiac, respiratory, and metabolic function in men in response to step work load. J Appl Physiol. 1982 May;52(5):1198-208. 48. Engeman, R.M., Swanson, D.G., Jones, R.H. Sinusoidal frequency allocation for estimating model parameters. I.E.E.E. Trans. (SMC) 1981;11:243.245. 49. Casaburi R, Weissman ML, Huntsman DJ, Whipp BJ, Wasserman K. Determinants of gas exchange kinetics during exercise in the dog. J Appl Physiol. 1979 Jun;46(6):1054-60. 50. Lamarra N, Ward SA, Whipp BJ. Model implications of gas exchange dynamics on blood gases in incremental exercise. J Appl Physiol. 1989 Apr;66(4):1539-46. 51. Whipp, B.J., Wasserman, K., casaburi, R., Juratsch, C., Weissman, M.L., Stremel, R.W. Ventilatory control characteristics of conditions resulting in isocapnic hyperpnea. Control of Respiration during Sleep and Anesthesia. 1978. New York, Plenum Press, pp 355-365. 52. Whipp BJ, Davis JA, Wasserman K. Ventilatory control of the 'isocapnic buffering' region in rapidly-incremental exercise. Respir Physiol. 1989 Jun;76(3):357-67. 53. Oldenburg FA, McCormack DW, Morse JL, Jones NL. A comparison of exercise responses in stairclimbing and cycling. J Appl Physiol. 1979 Mar;46(3):510-6. 54. Young IH, Woolcock AJ. Changes in arterial blood gas tensions during unsteady-state exercise. J Appl Physiol. 1978 Jan;44(1):93-6.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
119 55. Griffiths, T.G., Henson, L.C., Whipp, B.J. Influence of inspired oxygen concentration on the dynamics of the exercise hiperpnea. J Physiol 1986;380:387-407. 56. Oren A, Whipp BJ, Wasserman K. Effect of acid-base status on the kinetics of the ventilatory response to moderate exercise. J Appl Physiol. 1982 Apr;52(4):1013-7. 57. Casaburi R, Whipp BJ, Wasserman K, Stremel RW. Ventilatory control characteristics of the exercise hyperpnea as discerned from dynamic forcing techniques. Chest. 1978 Feb;73(2 Suppl):280-3. 58. Boetger CL, Ward DS. Effect of dopamine on transient ventilatory response to exercise. J Appl Physiol. 1986 Dec;61(6):2102-7. 59. Wasserman K, Whipp BJ, Koyal SN, Cleary MG. Effect of carotid body resection on ventilatory and acid-base control during exercise. J Appl Physiol. 1975 Sep;39(3):354-8. 60. Fordyce, W.E., Bennett, F.M. Some characteristics of a steady state model of exercise hyperpnea. Physilogist,. 1984;27(4):217. 61. Dempsey JA, Mitchell GS, Smith CA. Exercise and chemoreception. Am Rev Respir Dis. 1984 Feb;129(2 Pt 2):S31-4. 62. Armstrong BW, Hurt HH, Blide RW, Workman JM. The humoral regulation of breathing. Science. 1961 Jun 16;133:1897-906. 63. Sylvester JT, Whipp BJ, Wasserman K. Ventilatory control during brief infusions of CO2-laden blood in the awake dog. J Appl Physiol. 1973 Aug;35(2):178-86. 64. Filley GF, Heineken FG. A blood gas disequilibrium theory. Br J Dis Chest. 1976 Oct;70(4):223-45. 65. Waserman, K., Whipp, B.J., Casaburi, R., Beaver, W., Brown, H.V. CO2 flow to the lungs and ventilatory control. Muscular Exercise and the Lung. 1977 Madison, University of Wisconsin Pres, pp 105-135. 66. Yamamotho WS. Mathematical analysis of the time course of alveolar carbon dioxide. J Appl Physiol. 1960 Mar;15:215-9. 67. Murphy K, Stidwill RP, Cross BA, Leaver KD, Anastassiades E, Phillips M, Guz A, Semple SJ. Is hypercapnia necessary for the ventilatory response to exercise in man? Clin Sci (Lond). 1987 Dec;73(6):617-25. 68. Wasserman K, Whipp BJ, Casaburi R, Huntsman DJ, Castagna J, Lugliani R. Regulation of arterial PCO2 during intravenous CO2 loading. J Appl Physiol. 1975 Apr;38(4):651-6.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
120 69. Lewis SM. Awake baboon's ventilatory response to venous and inhaled CO2 loading. J Appl Physiol. 1975 Sep;39(3):417-22. 70. Reischl P, Gonzalez F Jr, Greco EC Jr, Fordyce WE, Grodins FS. Arterial PCO2 response to intravenous CO2 in awake dogs unencumbered by external breathing apparatus. J Appl Physiol. 1979 Nov;47(5):1099-104. 71. Shors, E.C., Huszczuk, A., Wasserman, K., Whipp, B.J. Effects os spinal-cord section and altered lung CO2 flow on the exercise hyperpnea in the dog. Modelling and Control of Breathing. 1983. New york, Elsevier, pp 274-281. 72. Stremel RW, Whipp BJ, Casaburi R, Huntsman DJ, Wasserman K. Hypopnea consequent to reduced pulmonary blood flow in the dog. J Appl Physiol. 1979 Jun;46(6):1171-7. 73. Sheldon MI, Green JF. Evidence for pulmonary CO2 chemosensitivity: effects on ventilation. J Appl Physiol. 1982 May;52(5):1192-7. 74. Huszczuk A, Whipp BJ, Oren A, Shors EC, Pokorski M, Nery LE, Wasserman K. Ventilatory responses to partial cardiopulmonary bypass at rest and exercise in dogs. J Appl Physiol. 1986 Aug;61(2):575-83. 75. Tibes U, Hemmer B, Boning D. Heart rate and ventilation in relation to venous (K+), osmolality, pH, PCO2, PO2, (orthophosphate), and (lactate) at transition from rest to exercise in athletes and non-athletes. Eur J Appl Physiol Occup Physiol. 1977 Jan 14;36(2):127-40. 76. Band DM, Linton RA, Kent R, Kurer FL. The effect of peripheral chemodenervation on the ventilatory response to potassium. Respir Physiol. 1985 May;60(2):217-25. 77. Burger RE, Estavillo JA, Kumar P, Nye PC, Paterson DJ. Effects of potassium, oxygen and carbon dioxide on the steady-state discharge of cat carotid body chemoreceptors. J Physiol. 1988 Jul;401:519-31. 78. Whipp BJ, Wasserman K. Carotid bodies and ventilatory control dynamics in man. Fed Proc. 1980 Jul;39(9):2668-73. 79. Witte KK, Thackray S, Nikitin NP, Cleland JG. The effects of long-term beta-blockade on the ventilatory responses to exercise in chronic heart failure. Eur J Heart Fail. 2005;7:612-7. 80. Dodd, S., Powers, S., OMalley, N., Brooks, E., Sommers, H. Effects of the beta-adrenergic blockade on ventilation and gas exchange during incremental exercise. Aviation Space Environ. Med 1989;59:718-722. 81. Filinsbee, L.J., Aallace, E.S., Bedi, J.F., Horvarh, S.M. Respiratory patterns and control during unrestrained human running. Modelling and Control of Breathing. 1983. New York, Elsevier, pp 205-212.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
121 82. Hammond MD, Gale GE, Kapitan KS, Ries A, Wagner PD. Pulmonary gas exchange in humans during exercise at sea level.J Appl Physiol. 1986;60:1590-8. 83. Hansen JE, Sue DY, Wasserman K. Predicted values for clinical exercise testing. Am Rev Respir Dis. 1984 Feb;129(2 Pt 2):S49-55. 84. Otis AB and Guyatt AR. The maximal frequency of breathing in man at varios tidal volumes. Respir Physiol.1968;5:118-9. 85. Milic-Emili G, Petit Jm, Deroanne R. The effects of respiratory rate on the mechanical work of breathing during muscular exercise.Arch Int Physiol Biochim. 1960;68:528-31. 86. Beck KC, Staats BA, Hyatt RE, Babb TG. Dynamics of breathing during exercise. En Exercise Pulmonary physiology and physiopathology. Whipp BJ y Wasserman K eds. Marcel Dekker 1991 New York pp 67-119. 87. Klas JV, Dempsey JA. Voluntary versus reflex regulation of maximal exercise flow: volume loops. Am Rev Respir Dis. 1989;139:150-6. 88. Grimby G, Saltin B, Wilhelmsen L. Pulmonary flow-volume and pressure-volume relationship during submaximal and maximal exercise in young well-trained men. Bull Physiopathol Respir (Nancy). 1971;7:157-72. 89. Grimby G. Peripheral limiting factors during exercise in chronic lung diseases. Bull Eur Physiopathol Respir. 1977 May-Jun;13(3):381-6. 90. Hesser CM, Linnarsson D, Fagraeus L. Pulmonary mechanisms and work of breathing at maximal ventilation and raised air pressure. J Appl Physiol. 1981 Apr;50(4):747-53. 91. Johnson BD, Reddan WG, Pegelow DF, Seow KC, Dempsey JA. Flow limitation and regulation of functional residual capacity during exercise in a physically active aging population.Am Rev Respir Dis. 1991;143:960-7. 92. Henke KG, Sharratt M, Pegelow D, Dempsey JA. Regulation of end-expiratory lung volume during exercise. J Appl Physiol 1988;64:135146 93. Younes M. Determinants of thoracic excursion during exercise. In Exercise Pulmonary Physiology and physiopathology. Whipp BJ and Wasserman K. eds. Editorial Marcel Dekker. NewYork 1991.pp 1-65. 94. Bye PT, Farkas GA, Roussos C. Respiratory factors limiting exercise. Annu Rev Physiol. 1983;45:439-51. 95. Warren GL, Cureton KJ, Sparling PB. Does lung function limit performance in a 24-hour ultramarathon? Respir Physiol. 1989 ;78:253-63.
REGULACIN DE LA VENTILACIN DURANTE EL EJERCICIO
122 96. Harms CA, Wetter TJ, McClaran SR, Pegelow DF, Nickele GA, Nelson WB, Hanson P, Dempsey JA. Effects of respiratory muscle work on cardiac output and its distribution during maximal exercise.J Appl Physiol. 1998;85:609-18. 97. Whipp BJ. The performance of the pulmonary system during exercise in athletes. En Physiological determinants of exercise tolerance in Humans. Whipp BJ y Sargenat AJ eds. Portland Press 1999.pp 135-151.
CAPTULO 6
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
ALBERTO ALONSO FERNNDEZ1 MNICA DE LA PEA BRAVO1 FRANCISCO GARCA RO2
1Servicio 2Servicio
de Neumologa. Hospital Universitario Son Dureta. Palma de Mallorca. de Neumologa. Hospital Universitario La Paz. Facultad de Medicina. Universidad Autnoma. Madrid.
Alberto Alonso Fernndez Hospital Son Dureta. Secretara Neumologa. Andrea Doria, 55. Palma de Mallorca07014 Tel: 660914808 aalonso@hsd.es
PALABRAS CLAVE: Arousals, control de la respiracin, hipercapnia sueo, hipoventilacin, hipoxia, mecanorreceptores, neuromoduladores, quimiorreceptores, reflejo de Hering-Breuer, sueo no REM, sueo REM, ventilacin, vas areas superiores.
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
126 NDICE DE CONTENIDOS: 1. Introduccin 2. Quimiorreceptores centrales 3. Quimiorreceptores perifricos 4. Receptores vagales y de las vas areas superiores 5. Control de la respiracin en nios 6. Control de la respiracin en adolescentes 7. Bibliografa
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
127 1. INTRODUCCIN A lo largo de los captulos precedentes se ha revisado como se produce el control de la respiracin y la implicacin de distintas unidades anatomo-funcionales en vigilia. La adecuada integracin de las diversas funciones del sistema nervioso central, los quimiorreceptores centrales y perifricos, as como los mecanorreceptores garantizan que se pueda llevar a cabo con xito el control de la respiracin. Durante el sueo se modifican las relaciones entre los distintos elementos implicados en el normal control de la respiracin, adems de producirse variaciones en la manera en la cual se procesa la informacin de los distintos receptores, tanto centrales como perifricos. El sueo no es un estado homogneo y bsicamente se intercalan tres o cuatro ciclos de sueo sin movimientos oculares rpidos (sueo no REM) y periodos en los que estos ltimos si estn presentes (sueo REM). Por su parte, en el sueo no REM se distingue cuatro fases, las dos ltimas de ellas conocidas como sueo de ondas lentas o sueo profundo. En el periodo REM, ocurren los sueos, y se caracteriza por una actividad metablica cerebral intensa y una activacin del sistema nervioso central. Los diferentes mecanismos implicados en el control de la respiracin varan segn el sujeto se encuentre en fase de sueo REM o no REM, adems, estos cambios evolucionan de una manera dinmica y fisiolgica a lo largo de la edad, existiendo diferencias significativas en la infancia y en los ancianos. En este captulo se revisan los mecanismos implicados en el control de la respiracin en el sueo. As mismo, se repasa la repercusin que la edad, las fases del sueo, y los neuromoduladores tienen en el mismo. 2. QUIMIORRECEPTORES CENTRALES La estimulacin de los quimiorreceptores centrales se realiza como consecuencia del incremento de hidrogeniones (H+) o de dixido de carbono (CO2) en el lquido extracelular cerebral o el lquido cefalorraqudeo. Clsicamente se crea que estaban en su mayora, ubicados en la superficie ventral de la mdula espinal. No obstante, en los ltimos aos, se han identificado otras localizaciones anatmicas de quimiorreceptores centrales que pueden ser tan importantes como el mencionado previamente, como en el ncleo fastigial, ncleo del tracto solitario, rafe caudal, locus caeruleus y ncleo ambiguo (1) (Figura 1). De todos ellos, probablemente el ncleo del tracto solitario es el principal protagonista de todas las localizaciones centrales de los quimiorreceptores, ya que es un importante centro que integra informacin de la homeostasis de funciones cardiorrespiratorias (2, 3). No slo se han identificado mltiples localizaciones de quimiorreceptores centrales, sino que adems las respuestas a los cambios en el CO2 varan segn el estado de conciencia (vigilia-sueo) y dentro del sueo, tambin se han detectado variaciones segn la fase del mismo entre las diferentes ubicaciones de los receptores (4). As, en pacientes que tienen daada la zona dorsolateral de la mdula se detectan periodos prolongados de apneas de
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
128
Figura 1. Centros respiratorios centrales.
caractersticas centrales (sin esfuerzos toracoabdominales acompaantes) durante el sueo, pero no en vigilia (5). Adems, en estos enfermos se detectaron reducciones en la respuesta ventilatoria al CO2 en vigilia, pero no a la hipoxia o al ejercicio (6). Existen datos obtenidos de experimentacin animal que han corroborado la presencia de distintos comportamientos de quimiorreceptores segn el estado vigilia-sueo y de su localizacin (2, 7). No obstante todava no est totalmente aclarado la manera exacta en la que interactan cada uno de los quimiorreceptores centrales y como se coordinan en las diferentes fases de sueo. Durante el sueo se detecta una disminucin de la ventilacin alveolar, siendo ms acusada a medida que progresa el mismo. De tal manera que la reduccin es mayor en las fases tres y cuatro de sueo no REM, y ms an en el periodo REM (8) (Figura 2A), sin cambios muy significativos en la frecuencia respiratoria (9) (Figura 2B). Esta hipoventilacin nocturna se relaciona con la pendiente de la respuesta ventilatoria a la hipercapnia, ya que se observa una disminucin de la respuesta en la fase no REM, que todava disminuye ms en REM (10, 11). Como consecuencia, se detectan incrementos de la presin de CO2 al final de las respiraciones a volumen corriente de dos a tres mm de Hg en la fase no REM, y de otros dos tres mm de Hg ms, en periodo REM. La disminucin observada en la ventilacin nocturna es desproporcionada con la reduccin que se detecta en el metabolismo basal durante el mismo tiempo. Se cree es consecuencia de una depresin en la quimiosensibilidad central, debido a la existencia de conexiones neuronales entre los sistemas responsables del control del ciclo vigilia-sueo y los centros respiratorios centrales (12). As, la activacin de sistemas gabargicos de las neuronas implicadas en el inicio del sueo en el ncleo ventrolate-
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
129
A)
B)
fR (% del valor en vigilia)
Figura 2. Representacin grfica de la influencia de las diferentes fases del sueo en la ventilacin (A) y en la frecuencia respiratoria (B). Modificado de Douglas (8, 9).
ral preptico del hipotlamo, conlleva una inhibicin de la vigilia y al mismo tiempo de la respiracin (13, 14). La reduccin de la excitacin del sistema reticular a los centros respiratorios, y el descenso de la inhibicin gabargica de grupos neuronales pontinos y medulares relacionados con la respiracin, justificara la menor sensibilidad de los quimiorreceptores centrales durante el sueo y la consecuente reduccin de la ventilacin (15). En este sentido, se ha demostrado que la infusin de anlogos de GABA a nivel sistmico o intraventricular, reducen la ventilacin minuto en ratas y sta, es adems dosis dependiente (16). De igual manera, la administracin de antagonistas de GABA en la superficie ventrolateral de la mdula de ratas provoca un incremento de la actividad del nervio frnico (17).
VE (L/min)
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
130 A pesar de que existe una disminucin de la sensibilidad de los quimiorreceptores centrales durante el sueo, la disminucin de la ventilacin no se puede explicar nicamente por este motivo, ya que tambin se eleva la resistencia de la va area superior, aumentando la carga mecnica de la respiracin, debido a una disminucin de la actividad de los msculos responsables de mantenerla permeable (18, 19), y adems se observa una disminucin de la actividad de los msculos accesorios de la respiracin que condiciona una leve hipoventilacin (20, 21). Conjuntamente, la hipocapnia tiene un potente efecto inhibitorio sobre la actividad muscular respiratoria que es mayor durante el sueo (22). No disponemos de la suficiente evidencia de que la disminucin en la sensibilidad al CO2 sea consecuencia slo de distintos comportamientos de los quimiorreceptores centrales durante el sueo. As, diferentes autores proponen que la actividad de los receptores depende del pH intracelular de dichas neuronas, ms que de los niveles plasmticos de CO2 o del pH sanguneo (23, 24). A su vez, el pH de las clulas neuronales obedece a cambios en el CO2 (que igualmente depende del balance entre metabolismo basal y flujo sanguneo local), y tambin a los diferentes transportadores de H+ de las membranas neuronales y del metabolismo intracelular, que es diferente en cada regin del tronco cerebral y adems su funcionamiento puede no seguir un patrn homogneo a lo largo del sueo (25-27). 3. QUIMIORRECEPTORES PERIFRICOS Como se ha comentado en captulos precedentes, existen quimiorreceptores perifricos localizados en la aorta y en ambas cartidas. Estas ltimas se agrupan formando el conocido como cuerpo carotdeo y debido a que su umbral de activacin est ms elevado, tienen mayor protagonismo real en el control de la respiracin. Su activacin est condicionada por los cambios en la concentracin de oxgeno arterial y son los responsables de la respuesta a la hipoxia y de un 15 a un 40% del impulso respiratorio en vigilia (28). No obstante, tambin, aunque en menor medida, estn modulados por el pH y el CO2 sanguneos (29). La respuesta a la hipoxia vara en funcin de las fases de sueo, aunque esta influencia es menor de la que se observa en relacin con la hipercapnia. De esta manera, durante la fase REM, se detecta una disminucin de la respuesta a la hipoxia, que no vara en funcin del gnero (Figura 3) (30-32). Por su parte, en el periodo no REM, se muestra una menor respuesta a la hipoxia, pero sta solo ha sido probada en varones. Se desconoce el porque de estas diferencias, no obstante hay que tener en cuenta que los varones tienen una mayor respuesta en vigilia que las mujeres y por lo tanto, un mayor margen para su posterior disminucin. No obstante, quizs estas diferencias representen comportamientos dispares del control de la respiracin durante el sueo segn el gnero y reflejen la influencia de las hormonas sexuales femeninas, lo cual, aunque no ha sido demostrado, podra justificar la diferencia en la prevalencia de ciertos trastornos respiratorios durante el sueo como el Sndrome de apneas-hipopneas (33). Recientemente se han identificado quimiorreceptores sensibles a la hipoxia en el tronco cerebral en modelos animales (34, 35), que interactan entre s y que potencialmente modulan la respuesta de los receptores perifricos (36). Sin embargo, el papel exacto que juegan
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
131
Ventilacin (L/min)
Saturacin de 02
Figura 3. Respuesta a la hipoxia en relacin con el estado de vigilia y las diferentes fases de sueo. Modificado de Douglas (9).
cada de uno de ellos, as como la influencia del sueo en estos ltimos est an por dilucidar. De tal manera, no est aclarado, si la menor respuesta a la hipoxia en el sueo se debe a un cambio en la respuesta del cuerpo carotdeo, de los receptores centrales o una combinacin de ambos factores. Adems del incremento de la ventilacin en respuesta a la hipoxia y a la hipercapnia, durante el sueo, cuando este mecanismo compensador no es suficiente, el organismo reacciona mediante la generacin de arousals o microdespertares a travs de la activacin del sistema simptico, que restauran, de forma parcial y momentnea los mecanismos de vigilia del control de la respiracin (37). Al igual que el incremento de la ventilacin como respuesta a la hipoxia, el umbral de los arousals vara en funcin del gnero (menor variacin a lo largo del sueo en las mujeres), y de las fases de sueo. Existen significativas variaciones en la presin de CO2 arterial necesaria para la aparicin de un arousal, no obstante la mayora ocurren entre 55 y 65 mm de Hg. Dicho umbral se incrementa en las fases profundas del sueo no REM, en comparacin con las fases 1 y 2 del sueo no REM y el sueo REM (10, 30). El umbral en relacin con la hipoxia es mucho ms variable, aunque se ha objetivado una respuesta con valores ms bajos de oxgeno arterial durante el sueo REM (10, 30, 37). 4. RECEPTORES VAGALES Y DE LAS VAS AREAS SUPERIORES El reflejo de Hering-Breuer se desencadena cuando se estimulan los receptores de estiramiento en el rbol bronquial, que a su vez, envan informacin a los centros respiratorios a travs del vago. Su activacin promueve la finalizacin de la fase inspiratoria y por tanto, tiene una influencia en la frecuencia respiratoria. Una mayor estimulacin de estos receptores conlleva tambin un estmulo para la finalizacin de la espiracin. Cuando la respiracin
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
132 se realiza a volumen corriente en reposo, el reflejo de Hering-Breuer tiene una influencia sutil en la modulacin de la respiracin (38, 39). En los ltimos aos, se ha demostrado que la reaccin vagal producida tras la estimulacin de los receptores de estiramiento situados en lo bronquios, tiene un mayor efecto sobre la actividad de los msculos de la va area superior y del diafragma, que sobre la modulacin de los tiempos inspiratorio y espiratorio (40-42). Durante el sueo el umbral de activacin del reflejo parece encontrarse elevado (43). La evolucin ha conseguido modular la anatoma de la va area superior de los humanos de una manera singular. As, el hioides est en una posicin sustancialmente baja y carece de capacidad de movimiento, gracias a los cual, se ha conseguido elongar la faringe y que sta, pueda estar sujeta a una mayor influencia muscular, consiguiendo as la posibilidad de emitir sonidos y por lo tanto, la comunicacin verbal. Como contrapartida, existe una faringe excesivamente larga, que demanda la existencia de una mayor actividad muscular para mantener la permeabilidad de la misma. Los msculos que principalmente estn implicados en el mantenimiento del calibre de la va area superior incluyen el geniogloso, el esternohioideo, y el tensor del paladar. La actividad del geniogloso durante fases no REM se asemeja a la que detectamos en periodos de vigilia, exceptuando una menor respuesta a la hipercapnia (44). En humanos no ha sido valorada dicha actividad en periodo REM, pero en modelos animales la respuesta de la actividad muscular del geniogloso a la hipercapnia en dicho periodo, se encuentra an ms disminuida (45). Adems de la influencia neuromecnica, la actividad de la musculatura de la va area superior tambin se halla sujeta al efecto modulador de la distorsin de la va area, el patrn del flujo respiratorio, la respiracin nasal, la temperatura del aire, y de la fase inspiratoria y espiratoria de cada uno de los ciclos respiratorios. De esta manera, cuando alguno de estos factores se encuentra comprometido (respiracin bucal, hipocapnia, cambio en la fase de sueo,..) se incrementa la posibilidad de alcanzar algn cierto grado de disfuncin de la va area superior, que favorece su capacidad para colapsarse y desarrollo ulterior de un sndrome de apneas-hipopneas del sueo (46). Existe una escasa correlacin entre la resistencia de la va area y la actividad de su musculatura (47, 48), y ello explicara que slo sean percibidos pequeos cambios en la resistencia de la va area en REM con respecto a los periodos no REM, a pesar de que si existe una disminucin clara en el tono muscular en la primera. 5. CONTROL DE LA RESPIRACIN EN NIOS La frecuencia respiratoria es muy elevada en el neonato (35-65 respiraciones por minuto), es mayor en el sueo REM que en el no REM y posteriormente se reduce la misma con el crecimiento del nio, siendo esta disminucin ms marcada en el primer ao de vida (49). Se ha comprobado que existe una relacin indirecta entre la ganancia ponderal del nio y la frecuencia respiratoria, ya que esta ltima disminuye exponencialmente al aumentar el peso.
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
133 Como consecuencia de la disminucin de la frecuencia respiratoria y el volumen corriente durante el sueo, se detecta una disminucin de la ventilacin en comparacin con la vigilia. En el sueo REM, la respiracin es irregular, con variaciones significativas en la frecuencia respiratoria y en el volumen corriente. De igual manera, no es infrecuente la aparicin de apneas centrales y obstructivas, favorecidas estas ltimas por la disminucin de la actividad de la musculatura de la va area superior, sin cambios en la contractilidad del diafragma. La respuesta ventilatoria a la hipoxia y a la hipercapnia se encuentra disminuida durante el sueo en los nios y como consecuencia se evidencian incrementos del CO2 y descensos en la saturacin de oxgeno (50). Tanto en vigilia como en sueo (sobretodo en el sueo tranquilo), existe un mayor control autonmico y una superior sensibilidad a los estmulos qumicos (51), as mismo, los quimiorreceptores perifricos tienen una respuesta independiente de las fases de sueo (52). 6. CONTROL DE LA RESPIRACIN EN ADOLESCENTES En el periodo de la adolescencia se experimentan una serie de cambios fsicos, metablicos, bioqumicos y psquicos, as como en el sueo y ritmos circadianos, que representan un periodo vital puente hacia la vida adulta. La respiracin as como el control de la misma son ya muy parecidos a los del adulto, y ya se puede detectar una frecuencia respiratoria ms elevada y variable en el periodo REM con respecto a las fases en las que no existen movimientos oculares rpidos (53).
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
134 7. BIBLIOGRAFA 1. Nattie E. Multiple sites for central chemoreception: their roles in response sensitivity and in sleep and wakefulness. Respir Physiol 2000; 122:223-235. 2. Nattie EE, Li A. CO2 dialysis in nucleus tractus solitarius region of rat increases ventilation in sleep and wakefulness. J Appl Physiol 2002; 92:2119-2130. 3. Parisian K, Wages P, Smith A, Jarosz J, Hewitt A, Leiter JC et al. Ventilatory effects of gap junction blockade in the NTS in awake rats. Respir Physiol Neurobiol 2004; 142:127-143. 4. Nattie EE. Central chemosensitivity, sleep, and wakefulness. Respir Physiol 2001; 129:257-268. 5. Manning HL, Leiter JC. Respiratory control and respiratory sensation in a patient with a ganglioglioma within the dorsocaudal brain stem. Am J Respir Crit Care Med 2000; 161:2100-2106. 6. Morrell MJ, Heywood P, Moosavi SH, Guz A, Stevens J. Unilateral focal lesions in the rostrolateral medulla influence chemosensitivity and breathing measured during wakefulness, sleep, and exercise. J Neurol Neurosurg Psychiatry 1999; 67:637-645. 7. Nattie EE, Li A. CO2 dialysis in the medullary raphe of the rat increases ventilation in sleep. J Appl Physiol 2001; 90:1247-1257. 8. Douglas NJ, White DP, Pickett CK, Weil JV, Zwillich CW. Respiration during sleep in normal man. Thorax 1982; 37:840-844. 9. Douglas NJ. Control of ventilation during sleep. Clin Chest Med 1985; 6:563-575. 10. Berthon-Jones M, Sullivan CE. Ventilation and arousal responses to hypercapnia in normal sleeping humans. J Appl Physiol 1984; 57:59-67. 11. Douglas NJ, White DP, Weil JV, Pickett CK, Zwillich CW. Hypercapnic ventilatory response in sleeping adults. Am Rev Respir Dis 1982; 126:758-762. 12. Rist KE, Daubenspeck JA, McGovern JF. Effects of non-REM sleep upon respiratory drive and the respiratory pump in humans. Respir Physiol 1986; 63:241-256. 13. Gallopin T, Fort P, Eggermann E, Cauli B, Luppi PH, Rossier J et al. Identification of sleep-promoting neurons in vitro. Nature 2000; 404:992-995. 14. Gervasoni D, Peyron C, Rampon C, Barbagli B, Chouvet G, Urbain N et al. Role and origin of the GABAergic innervation of dorsal raphe serotonergic neurons. J Neurosci 2000; 20:4217-4225.
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
135 15. Joseph V, Pequignot JM, Van Reeth O. Neurochemical perspectives on the control of breathing during sleep. Respir Physiol Neurobiol 2002; 130:253-263. 16. Hedner J, Jonason J, Lundberg D. Respiratory effects of gamma-hydroxybutyric acid in anesthetized rats. J Neural Transm 1980; 49:179-186. 17. Soto-Arape I, Burton MD, Kazemi H. Central amino acid neurotransmitters and the hypoxic ventilatory response. Am J Respir Crit Care Med 1995; 151:1113-1120. 18. Hudgel DW, Martin RJ, Johnson B, Hill P. Mechanics of the respiratory system and breathing pattern during sleep in normal humans. J Appl Physiol 1984; 56:133-137. 19. Hudgel DW, Devadatta P. Decrease in functional residual capacity during sleep in normal humans. J Appl Physiol 1984; 57:1319-1322. 20. Johnson MW, Remmers JE. Accessory muscle activity during sleep in chronic obstructive pulmonary disease. J Appl Physiol 1984; 57:1011-1017. 21. Millman RP, Knight H, Kline LR, Shore ET, Chung DC, Pack AI. Changes in compartmental ventilation in association with eye movements during REM sleep. J Appl Physiol 1988; 65:1196-1202. 22. Dempsey JA, Smith CA, Harms CA, Chow C, Saupe KW. Sleep-induced breathing instability. Sleep 1996; 19:236-247. 23. Filosa JA, Dean JB, Putnam RW. Role of intracellular and extracellular pH in the chemosensitive response of rat locus coeruleus neurones. J Physiol 2002; 541:493-509. 24. Wang W, Bradley SR, Richerson GB. Quantification of the response of rat medullary raphe neurones to independent changes in pH(o) and P(CO2). J Physiol 2002; 540:951-970. 25. Erlichman JS, Cook A, Schwab MC, Budd TW, Leiter JC. Heterogeneous patterns of pH regulation in glial cells in the dorsal and ventral medulla. Am J Physiol 2004; 286:R289R302. 26. Nottingham S, Leiter JC, Wages P, Buhay S, Erlichman JS. Developmental changes in intracellular pH regulation in medullary neurons of the rat. Am J Physiol 2001; 281:R1951. 27. Ritucci NA, Chambers-Kersh L, Dean JB, Putnam RW. Intracellular pH regulation in neurons from chemosensitive and nonchemosensitive areas of the medulla. Am J Physiol 1998; 275:R1152-R1163. 28. Pan LG, Forster HV, Martino P, Strecker PJ, Beales J, Serra A et al. Important role of carotid afferents in control of breathing. J Appl Physiol 1998; 85:1299-1306.
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
136 29. Forster HV. Plasticity in the control of breathing following sensory denervation. J Appl Physiol 2003; 94:784-794. 30. Berthon-Jones M, Sullivan CE. Ventilatory and arousal responses to hypoxia in sleeping humans. Am Rev Respir Dis 1982; 125:632-639. 31. Douglas NJ, White DP, Weil JV, Pickett CK, Martin RJ, Hudgel DW et al. Hypoxic ventilatory response decreases during sleep in normal men. Am Rev Respir Dis 1982; 125:286-289. 32. White DP, Douglas NJ, Pickett CK, Weil JV, Zwillich CW. Hypoxic ventilatory response during sleep in normal premenopausal women. Am Rev Respir Dis 1982; 126:530-533. 33. Duran J, Esnaola S, Rubio R, Iztueta A. Obstructive sleep apnea-hypopnea and related clinical features in a population-based sample of subjects aged 30 to 70 yr. Am J Respir Crit Care Med 2001; 163:685-689. 34. Golanov EV, Ruggiero DA, Reis DJ. A brainstem area mediating cerebrovascular and EEG responses to hypoxic excitation of rostral ventrolateral medulla in rat. J Physiol 2000; 529 Pt 2:413-29.:413-429. 35. Solomon IC, Edelman NH, Neubauer JA. Pre-Botzinger complex functions as a central hypoxia chemosensor for respiration in vivo. J Neurophysiol 2000; 83:2854-2868. 36. Gozal D, Gaultier C. Evolving concepts of the maturation of central pathways underlying the hypoxic ventilatory response. Am J Respir Crit Care Med 2001; 164:325-329. 37. Berry RB, Gleeson K. Respiratory arousal from sleep: mechanisms and significance. Sleep 1997; 20:654-675. 38. BuSha BF, Judd BG, Manning HL, Simon PM, Searle BC, Daubenspeck JA et al. Identification of respiratory vagal feedback in awake normal subjects using pseudorandom unloading. J Appl Physiol 2001; 90:2330-2340. 39. BuSha BF, Stella MH, Manning HL, Leiter JC. Termination of inspiration by phasedependent respiratory vagal feedback in awake normal humans. J Appl Physiol 2002; 93:903-910. 40. Bartlett D, Jr., St John WM. Influence of lung volume on phrenic, hypoglossal and mylohyoid nerve activities. Respir Physiol 1988; 73:97-109. 41. Kuna ST. Inhibition of inspiratory upper airway motoneuron activity by phasic volume feedback. J Appl Physiol 1986; 60:1373-1379.
CONTROL DE LA RESPIRACIN DURANTE EL SUEO
137 42. Wilson CR, Satoh M, Skatrud JB, Dempsey JA. Non-chemical inhibition of respiratory motor output during mechanical ventilation in sleeping humans. J Physiol 1999; 518:605-618. 43. Iber C, Simon P, Skatrud JB, Mahowald MW, Dempsey JA. The Breuer-Hering reflex in humans. Effects of pulmonary denervation and hypocapnia. Am J Respir Crit Care Med 1995; 152:217-224. 44. Pillar G, Malhotra A, Fogel RB, Beauregard J, Slamowitz DI, Shea SA et al. Upper airway muscle responsiveness to rising PCO(2) during NREM sleep. J Appl Physiol 2000; 89:1275-1282. 45. Horner RL, Liu X, Gill H, Nolan P, Liu H, Sood S. Effects of sleep-wake state on the genioglossus vs.diaphragm muscle response to CO(2) in rats. J Appl Physiol 2002; 92:878-887. 46. Krimsky WR, Leiter JC. Physiology of breathing and respiratory control during sleep. Semin Respir Crit Care Med 2005; 26:5-12. 47. Henke KG, Dempsey JA, Kowitz JM, Skatrud JB. Effects of sleep-induced increases in upper airway resistance on ventilation. J Appl Physiol 1990; 69:617-624. 48. Wiegand L, Zwillich CW, White DP. Collapsibility of the human upper airway during normal sleep. J Appl Physiol 1989; 66:1800-1808. 49. Carskadon MA, Harvey K, Dement WC, Guilleminault C, Simmons FB, Anders TF. Respiration during sleep in children. West J Med 1978; 128:477-481. 50. Duran CJ, Rubio Aramendi R. El sueo y la respiracin en las edades extremas: lo normal y lo anormal. In: Alvarez-Sala JL, Gonzalez-Mangado N, editors. Trastornos respiratorios del sueo. Madrid: Ergn; 2004. p. 189-211. 51. Waggener TB, Stark AR, Cohlan BA, Frantz ID, III. Apnea duration is related to ventilatory oscillation characteristics in newborn infants. J Appl Physiol 1984; 57:536-544. 52. Rigatto H. Control of breathing during sleep in the fetus and neonate. Principles and practice of sleep medicine in the child. Philadelphia: WB Saunders Co; 1995. p. 29-43. 53. Tabachnik E, Muller NL, Bryan AC, Levison H. Changes in ventilation and chest wall mechanics during sleep in normal adolescents. J Appl Physiol 1981; 51:557-564.
CAPTULO 7
MECANISMOS PATOGNICOS DE LA DISNEA
JOS M MARN TRIGO
Servicio de Neumologa, Hospital Universitario Miguel Servet Avda Isabel la Catlica, 1-3; Zaragoza 50009 jmmarint@unizar.es
PALABRAS CLAVE: Disnea, fisiopatologa respiratoria.
MECANISMOS PATOGNICOS DE LA DISNEA
140 NDICE DE CONTENIDOS: 1. Definicin de disnea 2. Bases fisiopatolgicas de la disnea 2.1. Origen multifactorial de la disnea 2.2. Intercambio gaseoso y disnea 2.3. Trax y disnea 2.3.1. Receptores pulmonares 2.3.2. Receptores musculares y de pared torcica 2.3.3. Sensibilidad de los centros respiratorios 3. Desacoplamiento neuromecnico 4. Disnea en diferentes entidades clnicas 4.1. Insuficiencia cardaca 4.2. Asma 4.3. EPOC 4.4. Fibrosis pulmonar 4.5. Enfermedad neuromuscular 5. Resumen 6. Bibliografa
MECANISMOS PATOGNICOS DE LA DISNEA
141 1. DEFINICIN DE DISNEA Se trata de un trmino clnico para expresar la dificultad respiratoria o la molestia que representa el esfuerzo de la respiracin. Este sntoma indica el mecanismo de alerta con el que el ser humano ha sido dotado para informarle de que el nivel de ventilacin es inadecuado o de que se est produciendo una alteracin orgnica o funcional en el aparato respiratorio. En sujetos sanos la disnea aparece con el ejercicio o la altura de forma natural. En pacientes, la disnea aparece con niveles mas reducidos de ejercicio y caractersticamente su intensidad es percibida de forma inhomogenea frente a un mismo estmulo disneizante; situacin que por otra parte sucede tambin con otras sensaciones como el dolor. Existe una definicin operativa de la disnea propuesta por la Sociedad Torcica Americana: un trmino utilizado para caracterizar la experiencia subjetiva de respiracin molesta de calidad e intensidad variables. La sensacin deriva de la interaccin de mltiples factores fisiolgicos, psicolgicos, sociales y ambientales y puede inducir respuestas secundarios fisiolgicas y de comportamiento (1). En base a esta definicin, en el presente captulo se revisa brevemente las bases fisiopatolgicas de la disnea y sus mecanismos de expresin clnica.
2. BASES FISIOPATOLGICAS DE LA DISNEA 2.1. Origen multifactorial de la disnea Numerosos factores son los que interaccionan para determinar finalmente la sensacin de disnea (figura 1). Un ejemplo clsico de este polimorfismo es el asma. Ante estmulos
Figura 1. Representacin esquemtica de las diferentes seales de aferencias y eferencias que contribuyen a la gnesis de la disnea.
MECANISMOS PATOGNICOS DE LA DISNEA
142 como por ejemplo un incremento de ejercicio en casos de asma, el haber recibido previamente un broncodilatador reduce la intensidad de la disnea y ello pese a no incrementarse los flujos areos. Por otra parte en asmticos, las crisis de broncoconstriccin naturales se asocian a una mayor disnea que las crisis de broncoconstriccin inducidas en el laboratorio mediante sustancias broncoconstrictoras. La disnea puede modularse por factores externos. Por ejemplo muchos pacientes con disnea crnica encuentran alivio cuando el aire fri choca contra la cara (2). Factores internos cognitivos, psicolgicos, y de comportamiento la afectan de forma significativa (3-6). El propio entrenamiento de la disnea ante un determinado estmulo ayuda a reducir su intensidad. 2.2. Intercambio gaseoso y disnea El control de la ventilacin y el grado de ventilacin alveolar estn estrechamente relacionados con la actividad de quimiorreceptores centrales y perifricos que sensan el nivel de PaO2 y de PaCO2 en sangre arterial. La hipercapnia e hipoxemia experimental puede inducir disnea. Incluso en sujetos cuyos msculos respiratorios se paralizan con curare o derivados, la inhalacin de aire enriquecido con CO2 induce disnea (7). El efecto de la hipoxemia es mas controvertido. En sujetos sanos, la induccin de hipoxemia produce efectos variables y la administracin de O2 suplementario a pacientes con hipoxemia basal tambin produce una respuesta no predecible (8). A nivel tisular, la hipoxia altera el ambiente metablico y produce una activacin simptica que modula la respuesta ventilatoria y sensorial durante el ejercicio (9). La idea imperante actual es que la hipoxemia en general produce hiperventilacin (e hiperinsuflacin en pacientes con EPOC) y se asocia a fatiga muscular con necesidad de mayor esfuerzo de contraccin muscular y finalmente mayor esfuerzo ventilatorio y disnea. 2.3. Trax y disnea 2.3.1. Receptores pulmonares A nivel pulmonar se han identificado tres tipos de receptores de estmulos claramente diferenciados (10). Los receptores de adaptacin lenta al estiramiento localizados en vas areas de grueso calibre y que responden al incremento de volumen pulmonar; un segundo grupo de receptores de adaptacin rpida, presentes en el epitelio de las vas areas y que responden a irritantes inhalados y finalmente los receptores justa-pulmonares (tambin llamados J o receptores de fibras C) que son fibras no mielinizadas distribuidas en el parnquima pulmonar, junto a los capilares pulmonares y en la mucosa bronquial. Estas fibras son estimuladas por multitud de factores exgenos (irritantes) y endgenos (elevacin de la presin local) y su estimulacin se sigue de apnea, boncoconstriccin e hipersecrecin mucosa (11). La estimulacin de estos receptores producen seales aferentes hacia el sistema nervioso central (SNC) conducidas por el n. vago. La vagotoma o el bloqueo del nervio y su efecto sobre la disnea ha sido estudiada extensamente. Por un lado, en respuesta a la apnea voluntaria y a cargas resistivas tanto sujetos sanos como en pacientes con enfisema no se han encontrado alteraciones (12,13); ni siquiera en el paciente que recibe un trasplante cardiopulmonar y que sufre una vagotoma existe diferencias en la sensacin de la disnea de
MECANISMOS PATOGNICOS DE LA DISNEA
143 esfuerzo frente a sujetos sanos (14). En pacientes cuadripljicos de alto nivel se produce vagotoma adems de una alteracin sensorial procedente de msculos y caja torcica. Cuando a estos pacientes se les induce boncoconstriccin por metacolina, desarrollan un grado de disnea similar al de los asmticos (15). La informacin proporcionada por estos estudios podra resumirse en que si bien los receptores pulmonares modulan el grado de disnea, stos no son imprescindibles como generadores nicos de disconfort respiratorio. 2.3.2. Receptores musculares y de pared torcica El cerebro recibe proyecciones procedentes de una gran variedad de receptores situados en los tendones, msculos y articulaciones de la caja torcica que pueden influenciar la sensacin de disnea. Estudios ya clsicos de disnea inducida por la apnea voluntaria en sujetos normales demostraron como puede aliviarse la disnea, al permitir la relajacin muscular o paralizando los msculos respiratorios (16). Paradjicamente, tambin puede inducirse disnea a pesar de paralizar los msculos durante las pruebas de estimulacin con CO2 (17). As pues los mecanoreceptores torcicos parecen jugar un papel importante pero no nico en la patognesis de la disnea. 2.3.3. Sensibilidad de los centros respiratorios Algunos autores han relacionado el grado de disnea percibida como una expresin del nivel de actividad de los centros respiratorios y del impulso ventilatorio central. Puesto que ante estmulos homogneos como la hiperinsuflacin pulmonar inducida, la respiracin a travs de resistencias o la induccin de debilidad muscular con agentes paralizantes, los sujetos sensan el esfuerzo ventilatorio en diferentes grados, se sugiere la idea de que debe existir diferencias innatas en el procesamiento central de un mismo esfuerzo ventilatorio cuya expresin sera la disnea. Nuestro grupo estudi en 26 pacientes con EPOC severa (FEV1 < 50% predicho) la respuesta del centro respiratorio a la hipercapnia inducida y el impulso ventilatorio subsiguiente (18). Encontramos 2 tipos de patrones de comportamiento: pacientes con alta sensibilidad al CO2 y baja sensibilidad al CO2 expresado como nivel de ventilacin por mmHg de CO2 inhalado (VE/PetCO2). Al realizar pruebas de esfuerzo mximas, se encontr que el grado de disnea de esfuerzo se relacionaba con el estado del impulso ventilatorio central obtenido por la inhalacin de CO2 a nivel basal (Figura 2). Es decir, la disnea no solo est relacionada con la carga ventilatoria impuesta (ejercicio en este caso), si no con el estado innato del comando central ventilatorio. De nuestros datos se sugiri que exista poca adaptabilidad del sistema nervioso central a los cambios en el rendimiento de los msculos respiratorios, al menos en pacientes con EPOC. Adems existi un cambio en el patrn ventilatorio, un aumento en el trabajo de los msculos respiratorios durante el ejercicio y un aumento en la ventilacin total que no se relacionaron de forma directa con el grado de disnea. Parece probable que un grupo de pacientes con EPOC severa, an presentando respuestas musculares similares a una carga determinada refieran diferentes grados de percepcin frente a ste trabajo. Los pacientes con altos grados de disnea pareceran los mejores candidatos a recibir intervencin mdica para modular su disnea independientemente de su grado de obstruccin. Estos datos traen de nuevo la idea de lo simplista que puede resultar tratar
MECANISMOS PATOGNICOS DE LA DISNEA
144
Delta Disnea
P0.1/PETCO2 Figura 2. Relacin entre el cambio de disnea desde el nivel de reposo hasta la percibida a mximo esfuerzo (delta dyspnea) y la respuesta central innata a la estimulacin de CO2 en reposo (tomado de cita 18).
a pacientes con EPOC basndose en el grado de alteracin de un nico parmetro fisiolgico como puede ser el FEV1.
3. DESACOPLAMIENTO NEURO-MECNICO De la informacin proporcionada mas arriba se deduce que no existe un nico grupo de receptores o sensores responsables del nivel de disnea. Tambin es evidente que es simplista pensar que exista una relacin directa entre el esfuerzo impuesto al sistema respiratorio, una respuesta ventilatoria paralela y la aparicin de disnea en un grado proporcional. Se han realizado numerosos esfuerzos para integrar ambas circunstancias (estimulacin aferente hacia el SNC y sensacin eferente desde el SNC). La teora mas aceptada es la que propone la existencia de una desproporcin en la relacin entre la longitud de una fibra muscular y la tensin que debe realizar para cumplir con su misin de contraerse y generar movimiento ventilatorio (19). En la realidad del movimiento respiratorio, la longitud se puede traducir en incremento de volumen inspiratorio y la tensin hace referencia a la presin generada por los msculos respiratorios en su contraccin. De acuerdo con esta teora la disnea aparece cuando la tensin muscular realizada es anormalmente elevada para generar un determinado incremento de volumen pulmonar. Una modificacin reciente de esta teora llamada, disociacin neuromecnica, indica que adems de los msculos respiratorios, existira una multitud de receptores a lo largo del sistema respiratorio de forma que la disnea aparece cuando no existe una relacin equilibrada entre el impulso ventilatorio central y la estimulacin del SNC por parte de aferencias desde el sistema respiratorio (20). La magnitud del impulso central es determinado por las necesidades metablicas (produccin de CO2 y consumo de O2) y en parte por el nivel de impulso central dirigido a la activacin de los msculos respiratorios.
MECANISMOS PATOGNICOS DE LA DISNEA
145 4. DISNEA EN DIFERENTES ENTIDADES CLNICAS 4.1. Insuficiencia cardaca Muchos pacientes con insuficiencia cardiaca detienen su ejercicio por disnea o molestias en piernas en un punto en que todava mantienen reserva cardiopulmonar para seguir realizando ejercicio fsico. Todo ello adems pese a que la mayora de ellos no retienen CO2 ni desaturan durante el ejercicio y por tanto los quimiorreceptores no han sido activados. La explicacin mas plausible para la disnea en estos pacientes es la presencia de una sobrecarga de msculos respiratorio debida a la isquemia local, aumento de elasticidad/congestin pulmonar y estimulacin de fibras C por aumento de la presin venosa pulmonar (11,21). La disnea descrita por estos pacientes habitualmente se refiere como sofocante, respiracin rpida, hambre de aire o respiracin demasiado trabajosa (tabla 1). 4.2. Asma En general el tipo de disnea que sufren los asmticos cuando desarrollan crisis en reposo o durante el ejercicio, es similar. En el asma inducida por el ejercicio o por un broncoconstric-
Tipo de respiracin
ICC
Asma
EPOC
Enf. Pulmonar difusa
Enf. NeuroVascular Muscular Pulmonar
X
Rpida Espiratoria Superficial Trabajosa Asfixiante hambre de aire Tensin torcica
X X X X X X X X X X X
Tabla 1. Lenguaje de la disnea en diferentes entidades clnicas.
MECANISMOS PATOGNICOS DE LA DISNEA
146 tor, la sensacin inicial es de tensin torcica y cuando progresa la obstruccin bronquial (p.e. medida por la cada del FEV1), los pacientes suelen referir respiracin trabajosa o con mucho esfuerzo. El nivel de disnea en respuesta a las crisis de broncoconstriccin inducidas tiene implicaciones clnicas. Los pacientes que sufren de ataques cuasi-fatales, tienen una percepcin disneica reducida durante las crisis (22). Estos pacientes han demostrado presentar un mayor nmero de hospitalizaciones por asma, episodios de ataques casi-fatales y muertes por asma (23). Los mecanismos que explicaran la disnea en asmticos parecen centrarse en el efecto de la estimulacin de receptores de irritacin vagal situados en las vas areas (24) y en la hiperinsuflacin dinmica que estimulara los mecanoreceptores de la caja torcica (25). 4.3. EPOC Es posiblemente la entidad clnica en la que la disnea ha sido mas ampliamente estudiada. Numerosos factores han demostrado estar implicados en la gnesis de la disnea de los pacientes con EPOC. Los mas ampliamente aceptados son: el aumento de la carga de trabajo impuesta a los msculos inspiratorios por la alteracin de su configuracin longitud-tensin secundaria al aumento de volumen torcico asociado a la hiperinsuflacin y al desarrollo de presin positiva espiratoria (26), debilidad muscular, aumento de la demanda ventilatoria debido al excesivo espacio muerto anatmico, compresin dinmica de las vas areas, factores cardiovasculares y en general, un conjunto de todos ellos. Cuando se valora el grado de severidad de la enfermedad mediante el nivel del FEV1, no se encuentra una relacin significativa entre ste y el grado de disnea de esfuerzo que refieren los pacientes en la vida diaria. Por el contrario, el grado de disnea de esfuerzo durante las pruebas de ejercicio mximo, se relaciona estrechamente con el nivel de atrapamiento areo dinmico que se desarrolla durante el ejercicio (20). El nivel de capacidad inspiratoria (CI) es el volumen espejo de la capacidad residual funcional y refleja el grado de atrapamiento areo. Nuestro grupo demostr que no solo durante el ejercicio mximo se produce hiperinsuflacin dinmica, sino que tambin durante actividades moderadas como el paseo se desarrolla una reduccin en la CI que es reflejo de atrapamiento areo e hiperinsuflacin (27). Encontramos que el grado de disnea que los pacientes con EPOC cuantifican al pasear, se relaciona de forma inversa con el nivel de CI de que disponen al final del paseo (figura 3). Esta relacin era mucho mas potente que la que exista con el nivel de FEV1 medido antes de la prueba. En sujetos sanos, la sensacin de exceso de trabajo respiratorio o disnea durante el ejercicio se describe en el momento de mximo esfuerzo. En contraposicin, los pacientes con EPOC describen la disnea a lo largo del esfuerzo y con descriptores tipo no puedo llenar mis pulmones o no puedo inspirar. Estas sensaciones probablemente tienen que ver con la hiperinsuflacin pulmonar que se desarrolla, con la produccin de presin intratorcica positiva al final de la espiracin y con el desacoplamiento de la relacin percepcin del esfuerzo respiratorio e impulso ventilatorio. 4.4. Fibrosis pulmonar En la enfermedad pulmonar difusa de localizacin intersticial existe una reduccin en la distensibilidad pulmonar y un aumento del espacio muerto. La rigidez del pulmn obliga a un
MECANISMOS PATOGNICOS DE LA DISNEA
147
Exertional dyspnea ( Borg)
IC (% TLC) Figura 3. Relacin entre el nivel de disnea al final de la prueba de marcha de 6 minutos y el cambio en la capacidad inspiratoria desde el nivel basal hasta el final de la prueba en pacientes con EPOC. (tomado de cita 27).
exceso de trabajo a los msculos respiratorios y la reduccin del espacio alveolar obliga a un incremento de la ventilacin. Ambos factores producen un aumento de la sensacin de esfuerzo ventilatorio y del impulso central ventilatorio con la consiguiente sensacin de disnea (28). Los pacientes afectos de fibrosis adoptan un patrn ventilatorio tipo taquipnea y platipnea, es decir frecuencias respiratorias rpidas y con volumen corriente reducido y describen la disnea como sofocante, hambre de aire, tensin torcica o muy trabajosa. 4.5. Enfermedad neuromuscular La alteracin primaria de la musculatura respiratoria en entidades como la miastenia gravis o la esclerosis lateral amiotrfica o distrofia muscular, induce un estado de debilidad de muscular que requiere un aumento del impulso ventilatorio central para activar la accin muscular (29). Los pacientes afectos describen su disnea como respiracin muy trabajosa o trax rgido.
5. RESUMEN La disnea es un sntoma que expresa la presencia de seales aferentes anormales hacia el sistema nervioso central procedentes del sistema respiratorio. No existe habitualmente un nico estimulante o receptor implicado en la disnea, por el contrario suelen estar implicados varios receptores (quimiorreceptores y mecanoreceptores) y diferentes aferencias centrales. Esto explica las diferentes formas individuales e interindividuales de expresar la sensacin disneica (30,31). Los pacientes suelen usar mas de un descriptor de disnea si bien algunas entida-
MECANISMOS PATOGNICOS DE LA DISNEA
148 des clnicas comparten algunas formas comunes para describir la disnea. Habitualmente los diferentes descriptores en el lenguaje de la disnea suelen representar el predominio de la actividad de determinados receptores si bien no suele existir un nico factor que explique el mecanismo fisiopatolgico de la disnea en una determinada entidad clnica.
MECANISMOS PATOGNICOS DE LA DISNEA
149 6. BIBLIOGRAFA 1. American Thoracic Society. Dyspnea, mechanisms, assessment and management: a consensus statement. Am J Respir Crit Care Med 1999; 159:321-40. 2. Schwartzstein, R. M., K. Lahive, A. Pope, S. E. Weinberger, and J. W. Weiss. Cold facial stimulation reduces breathlessness induced in normal subjects. Am. Rev. Respir. Dis. 1978; 136: 58-61. 3. Dudley, D. L., C. J. Martin, and T. H. Holmes. Dyspnea: psychological and physiologic observations. J. Psychosom. Res 1968;12: 205-214. 4. Gift, A. G., and C. A. Cahill. Psychophysiologic aspects of dyspnea in chronic obstructive pulmonary disease: a pilot study. Heart Lung 1990;19: 252-257. 5. Janson-Bjerklie, S., V. K. Carrieri, and M. Hudes. The sensations of pulmonary dyspnea. Nurs. Res 1986;35: 154-159 6. Demediuk, B. H., H. Manning, J. Lilly, V. Fencl, S. E. Weinberger, J. W. Weiss, and R. M. Schwartzstein. Dissociation between dyspnea and respiratory effort. Am. Rev. Respir. Dis. 1992;146: 1222-1225. 7. Campbell, E. J. M., S. C. Gandevia, K. J. Killian, C. K. Mahutte, and J. R. A. Rigg. Changes in the perception of inspiratory resistive loads during partial curarization. J. Physiol. 1990;309: 93-100. 8. Mak VHF, Bugler JR, Roberts CM, Spiro SG. Effect of arterial oxygen desaturation on six-minute walk distance, perceived effort and perceived breathlessness in patients with airway limitation. Thorax 1993; 48:33-48. 9. O'Donnell, D. E., D. J. Bain, and K. A. Webb. Factors contributing to relief of exertional breathlessness during hyperoxia in chronic airflow limitation. Am. J. Respir. Crit. Care Med. 1997;155: 530-535. 10. Widdicombe, J. G. Nervous receptors in the respiratory tract and lung. In T. D. Hornbein, editor. Lung Biology in Health and Disease, Vol. 1: Regulation of Breathing, 17th ed. Marcel Dekker, New York. 1981;429-472. 11. Paintal, A. S.. Sensations from J receptors. NIPS 1995;10: 238-243. 12. Guz, A.. Brain, breathing and breathlessness. Respir. Physiol. 1997;109: 197-204 13. Bradley GW, hale T, Pimble J. Effect of vagotomy in the breathing pattern and exercise ability in emphysematouse patients. Clin Sci 1982; 62:311-319.
MECANISMOS PATOGNICOS DE LA DISNEA
150 14. Kimoff, R. J., T. T. Cheong, M. G. Cosio, A. Querraty, and R. D. Levy. Pulmonary denervation: effects on dyspnea and ventilatory pattern during exercise. Am. Rev. Respir. Dis. 1990;142: 1034-1040. 15. Lougheed, M. D., K. A. Webb, and D. E. O'Donnell. Breathlessness during induced lung hyperinflation in asthma: the role of the inspiratory threshold load. Am. J. Respir. Crit. Care Med. 1995;152: 911-920 16. Campbell, E. J. M., S. C. Gandevia, K. J. Killian, C. K. Mahutte, and J. R. A. Rigg. Changes in the perception of inspiratory resistive loads during partial curarization. J. Physiol. 1990;309: 93-100 17. Banzett, R. B., R. W. Lansing, and R. Brown. `Air hunger' from increased PCO2 persists after complete neuromuscular block in humans. Respir. Physiol 1990;81: 1-17 18. Marin JM, Montes de Oca M, Rassulo J, Celli BR. Ventilatory drive at rest and perception of external dyspnea in severe COPD. Chest 1999; 115: 1293-1300. 19. Howell, J. B. L., and E. J. M. Campbell, editors. Breathlessness. Blackwell Scientific Publications, 1966; London. 20. O'Donnell, D. E., and K. A. Webb. Exertional breathlessness in patients with chronic airflow limitation. Am. Rev. Respir. Dis 1993;148: 1351-1357. 21. Mancini DM, Ferraro N, Nazzaro D, Chance B, Wilson JR. Respiratory muscle deoxygenation during exercise in patients with heart failure. J Am Coll Cardiol 1991; 18:492-498. 22. Kikuchi, Y., S. Okabe, G. Tamura, W. Hida, M. Homma, K. Shirato, and T. Takishima. Chemosensitivity and perception of dyspnea in patients with a history of near-fatal asthma. N. Engl. J. Med. 1994;330: 1329-1334. 23. Magadle R, Berar-Yanay N, Weiner P. The risk of hospitalization and near-fatal and fatal asthma in relation to perception of dyspnea. Chest 2002; 121:329-333. 24. Kikuchi, Y., S. Okabe, G. Tamura, W. Hida, M. Homma, K. Shirato, and T. Takishima. Chemosensitivity and perception of dyspnea in patients with a history of near-fatal asthma. N. Engl. J. Med. 1994;330: 1329-1334 25. Lougheed, M. D., K. A. Webb, and D. E. O'Donnell. Breathlessness during induced lung hyperinflation in asthma: the role of the inspiratory threshold load. Am. J. Respir. Crit. Care Med. 1995;152: 911-920. 26. Manning, H. L., and R. M. Schwartzstein. Pathophysiology of dyspnea. N. Engl. J. Med 1995;333: 1547-1553.
MECANISMOS PATOGNICOS DE LA DISNEA
151 27. Marin, J.M., Carrizo, S.J, Gascon, M., Sanchez, A., Gallego, B., Celli B.R. Inspiratory capacity, dynamic hyperinflation, breathlessness, and exercise performance during the 6minute-walk test in chronic obstructive pulmonary disease. Am J Respir Crit Care Med 2001; 163;1395-1399. 28. Brack T, Jubran A, Tobin M. Dyspnea and decrease variability of breathing in patients with restrictive lung disease. Am J Respir Crit Care Med 2002; 165:1260-1264. 29. Begin R, Bureau M, Lupien L, Bernier JP, Lemieux B. Pathogenesis of respiratory insuffiency in myotonic dystrophy. Am Rev Respir Dis 1982; 125:312-318. 30. Elliott, M. W., L. Adams, A. Cockcroft, K. D. McRae, K. Murphy, and A. Guz. The language of breathlessness: use of verbal descriptors by patients with cardiopulmonary disease. Am. Rev. Respir. Dis 1991;144: 826-832 31. Simon, P. M., R. M. Schwartzstein, J. W. Weiss, K. LaHive, V. Fencl, M. Teghtsoonian, and S. E. Weinberger. Distinguishable sensations of breathlessness in normal volunteers. Am. Rev. Respir. Dis 1989; 140: 1021-1027
CAPTULO 8
EXPLORACIN FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
JM. RODRGUEZ GONZLEZ-MORO, S. LPEZ MARTN, P. DE LUCAS RAMOS
Servicio de Neumologa. Seccin SVTS. H.G.U. Gregorio Maran. Madrid
Jos Miguel Rodrguez Gonzlez-Moro Servicio de Neumologa. Hospital Gregorio Maran C/ Dr. Esquerdo n 46-28007-MADRID Telfono: 91-5868328-30. Fax 91-5968018 Direccin electrnica: jrodriguezgo.hgugm@salud.madrid.org
PALABRAS CLAVE: Hipoventilacin, control de la ventilacin, funcin ventilatoria, sndrome de hipoventilacin.
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
154 NDICE DE CONTENIDOS: 1. Introduccin 2. Exploracin de la funcin ventilatoria 2.1. Estudios en vigilia 2.1.1. Conduccin neural 2.1.2. Conduccin neuromuscular a. Anlisis del patrn ventilatorio b. Presin de oclusin c. Presin inspiratoria mxima (PImax) 2.1.3. Estudios de quimiosensibilidad a. Respuesta ventilatoria al CO2 b. Respuesta ventilatoria a la hipoxia 2.2. Estudios durante el sueo 3. Sndromes clnicos de hipoventilacin 3.1. Sndrome de Hipoventilacin Alveolar Primaria 3.2. Hipoventilacin Alveolar Central 3.3. Sndrome de ObesidadHipoventilacin 3.4. Hipoventilacin en enfermedades musculares 3.5. Hipoventilacin en las alteraciones de la pared torcica 4. Evaluacin diagnstica 4.1. Estudio bsico 4.2. Estudio avanzado 4.3. Aplicacin clnica: algoritmo prctico 5. Resumen 6. Bibliografa
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
155 1. INTRODUCCIN El nivel de la pO2 alveolar est determinado por un equilibrio entre la rapidez con que la sangre retira O2 (lo que depende de las demandas metablicas de los tejidos) y la celeridad con que la ventilacin alveolar repone este O2. Por tanto, si esta ltima es demasiado baja, la pO2 alveolar desciende, y por razones similares la pCO2 asciende. Este fenmeno se denomina hipoventilacin alveolar y presenta dos caractersticas fisiolgicas fundamentales. En primer lugar, siempre provoca un incremento de la PaCO2 (arterial), lo que constituye un elemento diagnstico. La relacin entre la pCO2 y el nivel de ventilacin alveolar en el pulmn normal es muy simple: pCO2 = (VCO2/VA) x K donde VCO2 es la produccin de CO2, VA es la ventilacin alveolar y K es una constante. Por ello, si la ventilacin alveolar se reduce a la mitad, la pCO2 se duplicar. En segundo lugar, la hipoxemia puede ser corregida fcilmente aumentando la pO2 inspirada mediante la administracin de O2. Es importante destacar que la PaO2 (arterial) no puede descender a niveles muy bajos por la hipoventilacin pura. Si recordamos la ecuacin simplificada del gas alveolar para el oxgeno (PAO2= PIO2-PaCO2/R), si R es igual a 1, la pO2 alveolar desciende 1 mmHg por cada aumento de 1 mmHg de la pCO2. Esto significa que, para una hipoventilacin grave como para duplicar la PaCO2 de 40 a 80 mmHg, har descender la PaO2 de 100 a 60 mmHg, pero, an as, la saturacin de O2 ser de alrededor del 90%. Obviamente, mientras que el incremento de la PaCO2 tendr una importante traduccin clnica, la existencia de una saturacin del 90% asegura un aporte todava adecuado de O2 a los tejidos. Estas caractersticas son la base fisiolgica de los denominados cuadros de hipoventilacin alveolar crnica (1). En conjunto, constituyen un grupo de enfermedades frecuentes en la prctica neumolgica y, aunque pueden responder a diferentes mecanismos etiolgicos y patognicos, todas tienen el denominador comn de presentar una elevacin mantenida de la PaCO2 por encima de los 45 mmHg que consideramos como valor superior de la normalidad, Una gran mayora de los casos son debidos a la existencia de una obstruccin crnica al flujo areo (EPOC) en fase evolucionada, pero en otras ocasiones existen otros factores que los condicionan como son las alteraciones primarias o secundarias del control de la ventilacin, las enfermedades de la caja torcica (cifoescoliosis, espondilitis, fibrotrax, toracoplastias, etc) y los cuadros que cursan con debilidad de los msculos respiratorios. Estos procesos son los que pueden incluirse propiamente en los sndromes de hipoventilacin alveolar crnica (Tabla 1) (2-3). El objetivo general de las pruebas de funcin pulmonar consiste en la evaluacin del tipo y grado de disfuncin del aparato respiratorio. En este sentido, el papel que desempean en el estudio de los sndromes de hipoventilacin es fundamental y pueden clasificarse de manera un tanto esquemtica en dos categoras: 1) Pruebas que exploran la funcin ventilatoria y 2) Pruebas que permiten analizar la respuesta del aparato respiratorio ante diver-
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
156
Alteraciones del sistema nervi oso central Encefalitis Accidentes cerebrovasculares Traumatismos Tumores Enfermedades desmielinizantes Frmacos Txicos Idioptica Enfermedades mdula espinal y neuromusculares Poliomielitis Enfermedad de motoneurona Sndrome de Guillain-Barr Miastenia gravis Distrofias musculares Miopatas Parlisis diafragmtica Fatiga muscular Enfermedades pared torcica Cifoescoliosis Fibrotrax Toracoplastia Espondilitis anquilopoytica Sndrome de obesidad hipoventilacin Tabla 1. Causas de hipoventilacin alveolar crnica. No se mencionan las enfermedades obstructivas traqueales y bronquiales que en algn momento de su evolucin pueden cursar con hipoventilacin.
sos estmulos o situaciones. Dentro de cada uno de estos grupos existen una serie de tcnicas que proporcionan informacin global sobre la funcin objeto de estudio y otras tcnicas que permiten un anlisis ms especfico de los diferentes factores implicados en la misma. De todo ello puede desprenderse que el diseo de un estudio de funcin pulmonar en un paciente con sospecha de un cuadro de hipoventilacin crnica depender en cada caso de el problema objeto del anlisis (diagnstico, determinacin del grado de disfuncin) y de las circunstancias de aplicacin de las tcnicas (laboratorio de funcin pulmonar, consulta ambulatoria) (4). El presente captulo de este libro est dedicado al estudio de la exploracin funcional de los sndromes de hipoventilacin y se estructura en tres partes. En la primera, se realiza una breve descripcin de las pruebas de funcin ventilatoria empleadas en la exploracin de los sndromes de hipoventilacin ya que su estudio pormenorizado ha sido efectuado en ante-
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
157 riores captulos. En la segunda parte, se estudian las alteraciones que pueden encontrase en los sndromes y enfermedades ms relevantes. Por ltimo, en la tercera y ltima parte, se aborda la evaluacin diagnstica global y se propone un algoritmo de actuacin.
2. EXPLORACIN DE LA FUNCIN VENTILATORIA En el control del sistema respiratorio intervienen mecanismos voluntarios y reflejos que van a determinar la duracin de la inspiracin y la espiracin (5). Si la mecnica ventilatoria es normal, la ventilacin minuto y los valores de la PaCO2 como reflejo de la ventilacin alveolar constituyen un ndice del impulso ventilatorio. Es necesario determinar el rendimiento mecnico del aparato respiratorio y valorar si la respuesta ventilatoria se establece al nivel que corresponde a sus caractersticas mecnicas antes de comenzar el estudio del control de la ventilacin. Puesto que existen diferencias en el estudio de la respiracin durante la vigilia y el sueo vamos a separar su estudio en ambas situaciones (Tabla 2).
A-Vigilia Conduccin neural Electromiograma diafragmtico Conduccin neuromuscular Patrn ventilatorio Presin de oclusin Presin inspiratoria mxima Control qumico de la respiracin Respuesta ventilatoria a la hipercapnia Respuesta ventilatoria hipoxia Prueba de esfuerzo B-Sueo Polisomnografa Pulsioximetra nocturna Anlisis transcutneo de gases Capnografa Tabla 2. Mtodos de exploracin del control de la ventilacin.
2.1. Estudios en vigilia Con el fin de facilitar el estudio del control respiratorio durante la vigilia puede efectuarse una divisin en tres componentes, en funcin del lugar dnde se localice la alteracin:
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
158 1) Conduccin neural: referida a los impulsos enviados a los msculos respiratorios desde los centros respiratorios. 2) Conduccin neuromuscular: considerada como la respuesta de los msculos respiratorios a los impulsos nerviosos. 3) Control qumico de la respiracin evaluado a travs de la respuesta al estmulo qumico con CO2 o con O2. 2.1.1. Conduccin neural La actividad de los centros respiratorios no puede ser medida directamente, por lo que se han utilizado varios mtodos indirectos como la electromiografa del diafragma. El EMG diafragmtico refleja la medida de la conduccin neural desde los centros respiratorios a los nervios motores del diafragma (la actividad elctrica del diafragma guarda relacin directa con la actividad nerviosa del nervio frnico). En estudios con animales de experimentacin se ha visto que la actividad elctrica del diafragma se modificaba poco por las cargas elctricas o de resistencia al flujo, lo que hace que sea una buena medida aunque existan alteraciones mecnicas pulmonares o torcicas (7). Sin embargo, es una medida difcil de reproducir, no estandarizada, molesta para el paciente y al alcance de muy pocos centros, por lo que su utilizacin ha quedado muy restringida, siendo poco empleada en la prctica clnica habitual (8). 2.1.2. Conduccin neuromuscular La respuesta de los msculos respiratorios a los impulsos nerviosos puede determinarse a travs de un anlisis del patrn ventilatorio, ya que la ventilacin constituye la manifestacin externa del impulso ventilatorio, o mediante la presin de oclusin que mide la presin generada por los msculos respiratorios en respuesta al estmulo nervioso. a. Anlisis del patrn ventilatorio El patrn ventilatorio es expresin del rendimiento del control de la respiracin y de los cambios que se establecen en l para compensar las alteraciones morfolgicas o funcionales del aparato respiratorio.(9) La frecuencia, la profundidad y el patrn respiratorio reflejan una compleja interrelacin de mecanismos reguladores qumicos y neurohumorales que impulsan la funcin del aparato respiratorio. En el complejo control del sistema respiratorio intervienen mecanismos voluntarios y reflejos (qumicos, mecnicos...etc) que van a determinar el patrn respiratorio (10-11). Clsicamente, el patrn ventilatorio se analiza en trminos de frecuencia respiratoria (f) y volumen corriente (VT) mediante la formula de ventilacin minuto (VE): VE = f x VT
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
159 Clark y Von Euler (12) enunciaron el concepto de que el movimiento respiratorio es el resultado de un mecanismo "driving-timing". Este mecanismo, aceptado actualmente, considera que la ventilacin es el resultado de la interaccin de dos sistemas: uno "generador", que sera responsable del "impulso" o "drive" respiratorio y uno "oscilador" o "timing", responsable de la secuencia inspiracin-espiracin. De esta forma puede relacionarse la ventilacin con el estmulo respiratorio. Milic Emili introduce los siguientes parmetros (13-14): Ti: tiempo inspiratorio Ttot: duracin total del ciclo respiratorio Ti/Ttot: relacin entre tiempo inspiratorio y tiempo total ( fraccin de tiempo del ciclo total que se utiliza para la inspiracin, por lo que sera una medida del timing). VT/Ti: flujo inspiratorio medio VT/Ti (MIF), representa el volumen corriente alcanzado durante la inspiracin y viene a traducir el impulso neuromuscular o estmulo inspiratorio inicial (drive). Como la frecuencia y el tiempo respiratorio son inversamente proporcionales y el volumen corriente es el producto del flujo inspiratorio por el tiempo, la ecuacin que representa la ventilacin minuto (VE) puede expresarse as: VE: Ti/Tot x VT/Ti dnde el primer multiplicando se relaciona con el mecanismo de terminacin de la inspiracin y el segundo con el estmulo inspiratorio inicial. Existen diferentes mtodos de medida del patrn ventilatorio: Determinacin mediante mscara facial o pieza bucal. El paciente respira de forma tranquila a travs de una de ellas que est conectada a un espirmetro convencional o a un neumotacgrafo. Pletismografa inductiva respiratoria. Consiste en la colocacin de unas bandas torcicas y abdominales que van a registrar los cambios de volumen (15). Tanto la VE como el VT/Ti, aunque representan una medida del impulso central, resultan muy artefactadas por factores mecnicos torcicos y pulmonares. Este hecho ha determinado la bsqueda de medidas mas especficas como la presin de oclusin en los primeros 100 milisegundos (P0.1) que puede considerarse una expresin del impulso respiratorio neuromuscular.
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
160 b. Presin de oclusin La P0.1 es la presin negativa medida en la boca, producida como consecuencia de la contraccin de los msculos inspiratorios, durante los primeros 100 milisegundos de una inspiracin realizada contra una va area ocluida. Refleja la transformacin mecnica del impulso procedente de los centros respiratorios y se considera una determinacin muy til para la valoracin del impulso inspiratorio central por las razones que se exponen a continuacin (7,8,16,17): El periodo de 100 milisegundos resulta tan breve que ni siquiera resulta perceptible, lo que evita toda respuesta correctora (corticales) mientras el individuo respira contra la oclusin. Su medicin se realiza en ausencia de flujo areo y a nivel de capacidad residual funcional (equilibrio de las fuerzas de retraccin elstica), por lo que va a estar poco influenciada por alteraciones en la mecnica pulmonar y los datos obtenidos van a corresponder solo a la presin generada por los msculos inspiratorios. nicamente en el caso de alteraciones importantes de la FRC, la P0.1 puede experimentar cambios significativos. Por tanto, la P0.1 depende fundamentalmente de la informacin inspiratoria generada por los centros respiratorios, por la transmisin nerviosa y por las propiedades de los msculos inspiratorios. Aunque pudiera deducirse que en enfermedades que cursen con debilidad de los msculos inspiratorios podra verse afectada, este hecho solo se produce en situaciones extremas. En diversos estudios se ha podido comprobar que su medida guarda buena correlacin con la ventilacin minuto y el EMGdi (18). El valor normal de P0.1 es de -2.6 cm H2O con un rango entre 1.5-5 cm H2O. Se trata de una medida reproducible, aunque la variabilidad interindividual, interobservador e interda puede llegar a ser importante (cifras de hasta el 11% y el 13,9 %) (19). Este hecho hace que su determinacin deba realizarse de forma muy meticulosa teniendo en cuenta estos factores. La P0.1 se encuentra habitualmente elevada en las enfermedades pulmonares crnicas como reflejo de una "lucha" de los centros respiratorios contra la mecnica pulmonar alterada. Una P0.1 elevada refleja una actividad inspiratoria aumentada que podra desembocar a la larga en la aparicin de fatiga. Por el contrario, la P0.1 se encuentra disminuida en los cuadros de hipoventilacin de causa central. c. Presin inspiratoria mxima (PImax) La medida de la PI max es una tcnica sencilla y que proporciona una valiosa informacin acerca de la actividad de los msculos respiratorios. Habitualmente se efecta la medicin partiendo de volumen residual (RV) y, aunque existen valores de referencia, se consideran normales cifras por encima de 75 cm de H2O. Esta tcnica por si sola no permite diferenciar la afectacin neuromuscular intrnseca, de la secundaria a hiperinsuflacin pulmonar por lo que su interpretacin debe hacerse teniendo en cuenta el estudio de los volmenes pulmonares, sobre todo el RV.
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
161 2.1.3. Estudios de quimiosensibilidad El estudio del control qumico de la respiracin se efecta midiendo la respuesta de los centros respiratorios a la estimulacin con mezclas pobres en O2 (estimulacin hipxica) o enriquecidas en CO2 (estimulacin hipercpnica) y valorando las modificaciones que se producen en ventilacin, patrn ventilatorio y presin de oclusin (20-22). a. Respuesta ventilatoria al CO2 El CO2 es uno de los principales estmulos para la respiracin, por lo que la respuesta en condiciones estandarizadas a su aumento en el aire inspirado, permite cuantificar la sensibilidad de los centros respiratorios a este estmulo. La respuesta ventilatoria a las variaciones de la PaCO2 es lineal en un amplio rango de valores. Para su determinacin existen dos mtodos: Mtodo del estado estable (Lloyd y Cunningham) (23): el paciente respira una mezcla constituida por O2 al 21%, CO2 al 6 o al 8% y nitrgeno. Se realiza un control continuo de los niveles de CO2 en aire espirado hasta que se estabilizan, hecho que tiene lugar generalmente entre 8-10 minutos. Mtodo de respiracin en circuito cerrado o rebreathing (Read) (24): el paciente reinspira en una bolsa que contiene una mezcla gaseosa conteniendo una concentracin conocida de CO2 (habitualmente un 7%) y que ha sido enriquecida con oxgeno. Es una tcnica que resulta ms rpida y que permite alcanzar un mayor estmulo, por lo que prcticamente ha desplazado al mtodo del estado estable Con cualquiera de los dos mtodos se mide el incremento de la respuesta ventilatoria y de la P0.1 con relacin al valor basal. El resultado se describe mediante la pendiente de respuesta que relaciona la ventilacin y la presin de oclusin con la PCO2 teleespiratoria
A) Respuesta ventilatoria a la hipoxia -Valor medio pendiente VE/SatO2 -1,6 l/min./% (rango entre -0,18 y -4,90 l/min/%.) -Valor medio pendiente P0.1/SatO2 -0,06 cmH2O/ % (rango entre -0,01 y -0,23 cmH2O/%) B) Respuesta ventilatoria a la hipercapnia -Valores de la respuesta ventilatoria (VE/pCO2) a la hipercapnia - Baja: < 1.5 l./min./mmHg. - Intermedia: 1.5-5 l./min./mmHg. - Elevada: >5 l./min./mmHg. - Valor medio respuesta P0.1/ pCO2 0,49 cmH2O/mmHg (rango entre 0,12 y 2,64 cmH2O/mmHg. Tabla 3. Valores de respuesta ventilatoria a la hipoxia y la hipercapnia.
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
162 (VE/PCO2; P0.1/PCO2). En la tabla -3- se recoge la significacin de los valores de la respuesta ventilatoria obtenidos. El 80% de los individuos sanos presentan una respuesta ventilatoria que se sita en valores intermedios. Sin embargo, la magnitud de la respuesta ventilatoria es muy variable (desde individuos que responden con incrementos de VE del orden de 6-7 litros/min. por cada mmHg de aumento de pCO2, a otros que solo lo hacen 1 litro). De promedio, la respuesta es de 3 litros por cada mmHg de aumento, lo que se traduce en un incremento de la VE de 2 litros (VE normal en reposo 4 litros), lo que viene a indicar que la capacidad de correccin de la PaCO2 es bastante grande en general (25). Existen numerosas causas que pueden aumentar o reducir la respuesta ventilatoria a la hipercapnia, siendo las ms comunes las recogidas en la tabla 4. Los valores de la pendiente de P0.1 muestran tambin una gran variabilidad, con una media de 0,49 cm H2O / mmHg, situndose el rango considerado como normal entre 0,12 y 2,64 cm H2O / mmHg (Tabla 3) b. Respuesta ventilatoria a la hipoxia La hipoxemia, al igual que la hipercapnia, se acompaa de un incremento tanto en la VE como en la P0.1. En este caso la relacin no es lineal sino curvilnea, existiendo por el contrario, una relacin inversa lineal entre el grado de desaturacin y el incremento en las medidas de la ventilacin. La respuesta a la hipoxemia depende, sobre todo, de los quimioreceptores carotdeos. Por otra parte, la magnitud de la respuesta ventilatoria ante una disminucin de PaO2 depende tambin de la PaCO2 y aumenta a medida que lo hace la concentracin de CO2 en sangre arterial. Aunque existen varios procedimientos de medida, el ms utilizado es la estimulacin hipxica isocpnica progresiva. Consiste en la reinspiracin en una bolsa llena con aire, conectada a un filtro destinado a la absorcin de CO2. De esta forma se produce una cada progresiva en la saturacin de O2, mantenindose estables los niveles de CO2. La prueba finaliza cuando la saturacin de O2 cae por debajo del 80% 26 Al igual que ocurra con la hipercapnia, la respuesta ventilatoria a la hipoxemia muestra una gran variabilidad (7,26,27). Los valores medios para la pendiente VE/SatO2 son de -1,6 l/min./%, con un rango que oscila entre -0,18 y -4,90 l/min/%. Los cambios de la P0.1 tambin son muy variables, siendo el valor medio de -0,06 cmH2O/%, con un rango que se sita entre -0,01 y -0,23 cmH2O/%. En la tabla 4 se destacan aquellas situaciones que con mayor frecuencia cursan bien con una respuesta aumentada, bien con respuestas disminuidas. 2.2. Estudios durante el sueo Durante el sueo se producen una serie de alteraciones fisiolgicas entre las que destacan cambios sensoriales y metablicos, prdida del tono muscular, disminucin de la presin arterial y de la frecuencia cardiaca y modificaciones en el comportamiento del aparato respiratorio. Los cambios que el sueo origina sobre el control respiratorio contribuyen a una gran variedad de trastornos que, bien pueden ser enteramente responsables de una entidad clnica como el sndrome de apnea e hipopnea durante el sueo (SAHOS), bien pueden agra-
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
163
A) Respuesta hipercpnica Disminucin Atletas entrenados. Envejecimiento. Algunos grupos raciales. Frmacos: narcticos, barbitricos... Enfermedades neurolgicas: (encefaltis). Enfermedades metablicas: (mixedema). Obesidad-hipoventilacin Hipoventilacin primaria. Enfermedades neuromusculares. EPOC Hipercapnia Aumento: Concentraciones bajas de HCO3. Frmacos: teofilinas, salicilatos, tiroxina, progesterona. Hipertiroidismo. Aumento de resistencias de vas altas. Aumento de la impedancia del sistema respiratorio B) Respuesta a la hipoxemia Aumento: Situaciones que aumenten la respuesta ventilatoria al CO2. Existencia de niveles elevados de PaCO2. Disminucin: Hipoxemia prolongada (cardiopatas congnitas, grandes alturas...). Envejecimiento. Mixedema, opiceos, disfuncin del sistema nervioso autnomo. Endarterectoma carotdea Tabla 4. Cambios en la respuesta ventilatoria a la hipoxia y la hipercapnia.
var una patologa existente contribuyendo a la progresin de la misma, como sucede en el caso de cuadros como la enfermedad pulmonar obstructiva crnica (EPOC), las enfermedades neuromusculares o el sndrome de obesidad hipoventilacin (27-28). El sueo produce cambios a nivel de la ventilacin y de los volmenes pulmonares. La ventilacin disminuye en todas las etapas del sueo (sobre todo en las fases REM), debido a la perdida de la influencia del estado de vigilia (control automtico) y a los cambios que se producen en la sensibilidad de los centros respiratorios. En sujetos normales, durante las
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
164 fases REM del sueo se produce un descenso aproximado del 10% en la FRC. Esta disminucin probablemente est relacionada con la perdida del tono de los msculos intercostales y por hipoventilacin debido al menor impulso neuronal (29-30). Los mtodos de exploracin de las alteraciones ventilatorias durante el sueo van desde exploraciones sencillas como la pulsioximetra, la medida transcutnea de CO2 y la gasometra arterial nocturna hasta exploraciones ms completas como la poligrafa y la polisomnografa, que permiten registrar parmetros tanto ventilatorios como neurofisiolgicos (4). El estudio en profundidad de las alteraciones respiratorias durante el sueo y de los mtodos de exploracin de las mismas escapa de los objetivos de este captulo y se aborda con amplitud en otros apartados de estas monografas.
3. SNDROMES CLNICOS DE HIPOVENTILACIN En esta seccin se aborda de forma ms detallada el estudio clnico y funcional de los cuadros ms relevantes que, en la clnica habitual, pueden cursar en algn momento de su evolucin con hipoventilacin alveolar. En la Tabla 1 se encuentran recogidos estos procesos que van desde las alteraciones a nivel del control central de la ventilacin hasta las enfermedades de la caja torcica, pasando por los diferentes cuadros en los que la afectacin de los msculos respiratorios en el seno de una enfermedad neuromuscular condiciona el desarrollo de insuficiencia respiratoria. En la figura 1 se encuentra esquematizada la patogenia de la insuficiencia respiratoria que se produce en los diferentes grupos de patologas que pueden cursar con hipoventilacin alveolar crnica.
Figura 1. Patogenia de la insuficiencia respiratoria en los cuadros clnicos que pueden cursar con hipoventilacin alveolar crnica.
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
165 3.1. Sndrome de Hipoventilacin Alveolar Primaria Es una enfermedad de etiologa desconocida que est caracterizada por la existencia de hipercapnia crnica e hipoxemia, en ausencia de enfermedad neurolgica identificable, fatiga o enfermedad muscular o defectos mecnicos de la ventilacin (31). En su patogenia interviene un fallo en el sistema de control respiratorio metablico o automtico. Durante el da, por el efecto de la activacin reticular de las neuronas respiratorias, los pacientes son capaces de mantener una respiracin rtmica, aunque a un nivel ms bajo de lo normal. Durante el sueo, la respiracin pasa a depender del control metablico, cesa la respiracin rtmica y tienen lugar perodos de apnea e hipopnea central, con la consiguiente aparicin de alteraciones gasomtricas y el desarrollo progresivo de policitemia e hipertensin pulmonar que, de mantenerse en el tiempo, llevar a la alteracin gasomtrica en vigilia (32). Como enfermedad aislada es un cuadro raro, pero se considera que participa en cierto grado en otras enfermedades, acentuando la hipoventilacin que se produce en patologas neuromusculares, el sndrome de obesidad-hipoventilacin e incluso en la EPOC. Las caractersticas funcionales del sndrome de hipoventilacin alveolar primaria son las siguientes (32): Espirometra, pletismografa, compliance, resistencias de la va area y presiones musculares normales. Disminucin de la PaCO2 con la hiperventilacin voluntaria Presin de oclusin muy disminuida e incluso ausente. Respuesta ventilatoria anormal ante la estimulacin hipercpnica o hipoxmica Respuesta ventilatoria al ejercicio anormal, provocando elevacin de la PaCO2 y descenso de la PaO2. Como en estadios iniciales la PaCO2 puede ser normal en vigilia, los estudios nocturnos, en especial la poligrafa-polisomnografa, cobran especial importancia en su diagnstico. 3.2. Hipoventilacin Alveolar Central En este cuadro, a diferencia de la hipoventilacin idioptica, existe una causa neurolgica conocida. Entre sus etiologas figuran enfermedades como la encefalitis, los accidentes cerebrovasculares y los tumores, traumatismos y la administracin crnica de frmacos (barbitricos, morfina, hipnticos, etc) o la exposicin a diversos txicos. Los datos clnicos y los hallazgos que pueden encontrarse en el laboratorio de exploracin funcional respiratoria son similares a los que han sido descritos en el cuadro idioptico, aunque es caracterstico que en estos casos el paciente no es capaz de aumentar de forma voluntaria su nivel de ventilacin (30-32).
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
166 3.3. Sndrome de ObesidadHipoventilacin Es un cuadro caracterizado por obesidad importante e hipoventilacin alveolar y constituye la expresin mxima de las consecuencias que la obesidad tiene sobre la funcin respiratoria. En la patogenia de la insuficiencia respiratoria de este sndrome pueden intervenir diversos factores. En primer lugar, el acumulo de grasa en las musculatura y en la caja torcica ocasiona una disminucin de la elasticidad del sistema respiratorio. Adems, los volmenes pulmonares disminuyen, de forma ms acusada la FRC, lo que lleva a la aparicin de microatelectasias y alteracin de las relaciones ventilacin-perfusin. Por ltimo, existe una alteracin a nivel de los centros respiratorios con disminucin de quimiosensibilidad al CO2 y una mayor incidencia de SAHOS (33-34). Las caractersticas funcionales del sndrome de obesidad-hipoventilacin pueden incluir las siguientes alteraciones: Patrn funcional de tipo restrictivo con disminucin de los volmenes dinmicos: capacidad vital inspiratoria, volumen de reserva espiratorio y de la FRC. El volumen residual permanece normal o slo ligeramente descendido. El estudio de elasticidad pulmonar pone de manifiesto una presin elstica normal con un coeficiente de retraccin elevado y una distensibilidad disminuida. La PI max puede encontrarse disminuida, con PE max normal Presin de oclusin en general aumentada como reflejo del incremento de las resistencias elsticas del trax, Hiporespuesta de la presin de oclusin y de la ventilacin frente a estmulo qumico con CO2. Este hecho traduce una hiposensibilidad de los centros respiratorios que podra ser importante para explicar la patogenia del sndrome. Los estudios nocturnos (poligrafa, polisomnografa) son obligados para descartar la existencia de un SAHS asociado y valorar de forma adecuada la repercusin que el sueo tiene en el deterioro gasomtrico. 3.4. Hipoventilacin en enfermedades musculares Las enfermedades musculares constituyen una de las causas mas frecuentes de hipoventilacin. Cualquier enfermedad que afecte a la musculatura respiratoria puede provocar una alteracin de la ventilacin, cuyo grado estar condicionado por la gravedad del proceso de base (35). En estos enfermos puede apreciarse, antes de producirse hipercapnia, una disminucin de la capacidad vital y de la presin inspiratoria mxima. Mencin aparte hay que hablar de un tipo especfico de hipoventilacin por fallo muscular respiratorio que es el que se produce a causa del desarrollo de fatiga muscular. Esta fatiga puede ocurrir por factores como el aumento mantenido del trabajo respiratorio, por disminucin de la eficacia de los
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
167 msculos respiratorios, o por falta de los nutrientes energticos necesarios para la actividad muscular. Un ejemplo tpico del desarrollo de fatiga muscular lo constituye la EPOC evolucionada, patologa en la que pueden converger los tres factores anteriormente comentados. Las indicaciones ms importantes para la evaluacin de la funcin de los msculos respiratorios en clnica son cuantificar su debilidad en enfermedades pulmonares, neuromusculares o de la pared torcica y el estudio de la disnea y/o la alteracin gasomtrica no atribuible a patologa cardiopulmonar. El estudio detallado de las pruebas de funcin muscular escapa de los objetivos de esta monografa y remitimos al lector al volumen III de esta serie de monografas dedicadas al estudio de la funcin respiratoria. Sin embargo, a grandes rasgos se pueden dividir estas exploraciones como se describe a continuacin (36). a) Evaluacin de la fuerza global de los msculos respiratorios. Varias de las pruebas funcionales utilizadas rutinariamente dependen del esfuerzo, y por tanto pueden ser empleadas para controlar la fuerza de los msculos respiratorios. La capacidad vital (VC) ha sido utilizada con este fin ya que sus resultados dependen de la fuerza de los msculos inspiratorios y espiratorios, y de la compliance toracopulmonar. La VC es un ndice poco sensible para evidenciar una disminucin de escasa magnitud de la fuerza de los msculos respiratorios debido a la forma de la curva volumen-presin del sistema toracopulmonar. La ventilacin voluntaria mxima, fcil de realizar con un espirmetro estandard, es un ndice que proporciona una informacin global del grado de resistencia muscular. Las presiones estticas mximas - PI max y la PE max- son el mtodo ms sencillo para explorar la fuerza mxima que pueden desarrollar en forma integrada los msculos inspiratorios y espiratorios (36-37). b) Evaluacin del diafragma: la presin generada a travs de este msculo o presin transdiafragmtica (Pdi) se calcula a partir de la diferencia de la presin abdominal y la presin pleural medidas de forma simultanea No solo interesa medir tambin la Pdi generada para producir el volumen corriente, sino tambin la Pdi mxima generada durante una contraccin isomtrica a FRC, que representa una estimacin de la fuerza que desarrolla el diafragma contra los msculos abdominales. La Pdi mxima ha sido utilizada para el diagnstico de la parlisis diafragmtica bilateral y la relacin Pdi/Pdi max ha adquirido significacin para explorar la fatigabilidad del diafragma. Una forma ms sencilla que permite valorar la contraccin del diafragma y de los msculos inspiratorios en su conjunto es la denominada maniobra de sniff, que se realiza mediante una inspiracin voluntaria brusca y de corta duracin, efectuada con uno o ambos orificios nasales abiertos (Pdi max sniff) (37). c) Electromiografa: el anlisis espectral del electromiograma (EMG) puede ser utilizado para el estudio de la fatiga muscular ya que los cambios electromiogrficos preceden a la aparicin de los fenmenos mecnicos de agotamiento. Las caractersticas funcionales de la afectacin respiratoria en las enfermedades neuromusculares pueden incluir las siguientes alteraciones:
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
168 La espirometra y la pletismografa muestran una alteracin funcional restrictiva con disminucin de la capacidad vital (VC) y de los volmenes dinmicos. El VR puede estar normal o incrementado si existe debilidad de los msculos espiratorios. Resulta de gran utilidad la determinacin seriada de la capacidad vital, de forma que un descenso de la misma por debajo de 15 ml/kg resulta muy indicativo de riesgo de fallo ventilatorio, sobre todo si se ha objetivado una tendencia decreciente clara. Puede resultar de utilidad realizar la medida de la VC sentado o en bipedestacin y en dcubito supino ya que las variaciones posturales de esta sugieren el diagnstico de parlisis diafragmtica bilateral. En esta situacin el cambio de posicin erecta al dcubito supino se asocia con una reduccin de la VC mayor del 30%. El patrn respiratorio est caracterizado por una respiracin ms rpida y superficial. Este hecho hace que en fases iniciales de la enfermedad se observen incluso descensos en las cifras de PaCO2. La respuesta inicial del sistema de control de la respiracin es aumentar el impulso inspiratorio central, pero este mecanismo compensador acaba agotndose. Recientemente se ha demostrado que a partir de la determinacin de la P0.1 es posible realizar una estimacin de la presin esofgica respirando a volumen corriente en enfermos neuromusculares y, por tanto, determinar el ndice tensin-tiempo de los msculos respiratorios (38). Con referencia a las presiones musculares, cifras inferiores a 20 cm H2O en la PImax son indicativas de debilidad muscular con hipoventilacin alveolar asociada, por lo que la hipercapnia es la norma en estos casos. La relacin entre los parmetros de fuerza muscular y la VC guarda una relacin sigmoidea (similar a la curva de saturacin de la hemoglobina). As, descensos de la fuerza muscular de hasta el 50% pueden cursar con VC normal y, una vez que la presin muscular desciende a valores inferiores al 50%, la cada de la VC se torna ms rpida (Figura 2) (37). Este hecho hace que sea importante la realizacin de estudios durante el ejercicio, y sobre todo durante el sueo, en pacientes con enfermedades neuromusculares en los que la cada de los volmenes y las presiones musculares sean moderados, con el fin de detectar precozmente las alteraciones gasomtricas e instaurar el tratamiento oportuno. 3.5. Hipoventilacin en las alteraciones de la pared torcica En los cuadros como la cifoescoliosis, la espondilitis anquilopoytica y las secuelas de tuberculosis (fibrotrax, toracoplastias) es frecuente la aparicin en fases avanzadas de insuficiencia respiratoria crnica (39). Estas situaciones provocan una disminucin de los volmenes pulmonares y un incremento del trabajo respiratorio debido a la escasa movilidad de la caja torcica (40). En las enfermedades de la caja torcica, sobre todo la cifoescoliosis, existe una disminucin muy importante del VRE, indicando que la respiracin normal se hace partiendo casi del VR (41). En los casos secundarios a secuelas de tuberculosis es frecuente la coexistencia de un trastorno obstructivo como consecuencia de la distorsin del rbol bronquial.
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
169
Figura 2. Relacin entre la capacidad vital (VC) y la presin inspiratoria mxima (PImax). Se observa que puede existir un descenso importante de la PI max sin cambios significativos en la VC.
La hipoventilacin secundaria a enfermedades toracgenas resulta de fcil diagnstico ya que suma a las evidentes alteraciones radiolgicas, el hallazgo funcional de una alteracin ventilatoria de tipo restrictivo. No existe afectacin a nivel de centros reguladores ni de msculos (salvo en los casos de cifoescoliosis secundaria a enfermedades neuromusculares como por ejemplo la enfermedad de Duchenne) por lo que no es necesario ampliar el estudio funcional respiratorio 4. EVALUACIN DIAGNSTICA El diagnstico de estos procesos requiere de una historia clnica cuidadosa, de tcnicas de imagen (radiografa de trax) y sin duda, lo ms importante, de la exploracin funcional respiratoria. Las pruebas habitualmente utilizadas figuran en la Tabla-2- y van desde la realizacin de una gasometra arterial hasta pruebas de elevada complejidad como pueden ser la medida de presiones transdiafragmticas. 4.1. Estudio bsico El estudio bsico inicial de un paciente con sospecha clnica de un sndrome de hipoventilacin debe constar de la realizacin de una espirometra forzada con prueba broncodilatadora, la determinacin de volmenes pulmonares, la medida de presiones musculares mximas, la medida del patrn ventilatorio y una gasometra arterial en condiciones basales. Son pruebas esenciales, pero de gran importancia por su simplicidad, aceptacin por parte del paciente e informacin que aportan. En esta apartado incluimos tambin los estudios durante el sueo, desde la sencilla pulsioximetra y la gasometra arterial nocturna, hasta la polisomnografa convencional.
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
170 4.2. Estudio avanzado Dentro de este apartado se incluye el estudio del impulso ventilatorio central y de la respuesta a estmulos hipxico e hipercpnico, las pruebas de esfuerzo a las que se dedica otro captulo de esta monografa y la exploracin sofisticada de la funcin muscular (EMG, Pdi, etc). Estas pruebas estn slo al alcance de centros de referencia y su utilizacin queda restringida a cuadros de difcil diagnstico y estudios de investigacin. 4.3. Aplicacin clnica: algoritmo prctico El primer paso para valorar el estado del control ventilatorio se realiza a travs de la medida de los gases sanguneos en sangre arterial en condiciones de vigilia y en reposo. El hallazgo de una PaCO2 elevada junto con una cifra de bicarbonato tambin elevada confirma la existencia de una situacin de hipoventilacin alveolar crnica. La primera medida en estos casos es descartar la existencia de alteraciones mecnicas mediante la realizacin de espirometra y pletismografa. La existencia de un cuadro funcional de tipo obstructivo orientar hacia la existencia de una EPOC, mientras que la demostracin de alteraciones de tipo restrictivo debe hacer completar el estudio con la medida de presiones musculares mximas con el objetivo de descartar una enfermedad neuromuscular. Si no existen alteraciones mecnicas, o bien la alteracin es inferior al grado de hipoventilacin encontrado y s, adems el gradiente alvolo-arterial de oxgeno es normal pero los gases se normalizan tras la realizacin de una hiperventilacin voluntaria, puede sospecharse un cuadro de hipoventilacin de tipo central. La determinacin de la presin de oclusin basal y en respuesta a los estmulos qumicos servir para confirmar este diagnstico. Si tanto la PaO2 como la PaCO2 en vigilia son normales pero los datos clnicos (somnolencia, hipertensin pulmonar, poligobulia, disnea) orientan a la existencia de un sndrome de hipoventilacin, el estudio debe completarse con la realizacin de pruebas nocturnas (pulsioximetra, gasometra nocturna, poligrafa, polisomnografa) o una prueba de esfuerzo en funcin de la sospecha diagnstica.
5. RESUMEN La bomba ventilatoria es un rgano vital cuya funcin depende del estmulo neural, la maquinaria contrctil y las estructuras de la pared torcica. Puede fracasar por alteraciones en la generacin o transmisin del estmulo inspiratorio central, defectos estructurales de la pared torcica, patologas que afectan al msculo estriado o como consecuencia de un desequilibrio entre el aporte energtico al msculo y el trabajo externo. Las pruebas funcionales disponibles para evaluar cada uno de los aspectos mencionados son generalmente complejas, tiene diversas limitaciones metodolgicas, pero permiten obtener un conocimiento ms objetivo de la alteracin ventilatoria que presenta el paciente.
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
171 6. BIBLIOGRAFA 1. Weinberger SE, Schwartzstein RM, Weis JW. Hipercapnia. N Engl J Med 1989;321:1223-1231. 2. Glauser FL, Fairman RP, Bechard D. The causes and evaluation of chronic hypercapnea.Chest 1987;91:755-759. 3. Strumpf DA, Millman RP, Hill NS. The management of chronic hypoventilation. Chest 1990;98:474-480 4. Arnedillo Muoz A, Snchez Juanes MJ, Rodrguez Gonzlez-Moro JM, de Lucas Ramos P. La exploracin funcional en Neumologa: indicaciones y modalidades.Medicine 1994;6(65):563-575. 5. Nunn JF. Control of breathing. En: Nunn JF Editor. Nunns applied respiratroy physiology. 4 th edition. Cambridge: Butterworth-Heinemann LTD 1993;pp 90-116. 6. Caruana-Montaldo B, Gleeson K, Zwillich CW. The control of breathing in clinical practice. Chest 2000;117:205-225 7. Cherniack NS, Lederer DH,Althose MD,Kelsen SG. Occlusion pressure as a technique in evaluating respiratory control. Chest 1976 ;70 (Suppl): 137-141. 8. Burki NK. Measurements of ventilatory regulation. Clin Chest Med 1989;10:215-226. 9. Marn trigo JM. Inters del estudio del patrn ventilatorio. Arch Bronconeumol 1989;25:91-92. 10. Remmers JE. A century of control of breathing. Am J Respir Crit Care Med 2005;172:6-11. 11. de Miguel Poch, Palomera Frade J, Villena Garrido V, Gaite Alvaro A. Alteraciones en el control de la ventilacin. Par 1991;98:39-52. 12. Clark FJ, Von Euler C. On the regulation of depth and rate of breathing. J Physiol 1972;222:267-295. 13. Milic Emili J, Grunstein MM. Drive and Timming components of ventilation. Chest 1976;70:114-117. 14. Milic-Emili J. Recents advantages in clinical assessment of control of breathing. Lung 1982;160:1-17. 15. Chadha TS, Watson H, Birch S et al. Validation of respiratory inductive plethysmography using different calibration procedures. Am Rev Respir Dis 1982;125:644-649.
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
172 16. Whitelaw WA, Derenne J, Milic-Emili J. Occlusion pressure as a measure of respiratory center outpout in conscious man. Respir Physiol 1975 ;23 :181-199. 17. Grippi MA, Metzger LF, Krupinski AV, Fishman AP . Pruebas funcionales pulmonares en AP Fishman: Tratado de Neumologa 2 Edicin. Ed Doyma. Barcelona 1991. Vol 3. pp 2289-2235. 18. Garay SM, Rappaport DM. Control of respiration: ventilatory response testing en Miller A. Pulmonary function test in clinical and occupational lung disease. Ed Grune-Straton. Orlando 1986. 19. Whitelaw WA, Derenne JP. Airway occlusion pressure. J Appl Physiol 1993; 74: 14751483. 20. Berger AJ. Regulation of respiration (1). N Engl J Med 1977 ;297 :92-97. 21. Berger AJ. Regulation of respiration (2). N Engl J Med 1977;297 :138-143. 22. Berger AJ. Regulation of respiration (3). N Engl J Med 1977; 297:194-201. 23. Lloyd BB, Cunningham DJC. A quantitative approach to the regulation of human respiration. En Cunningham DJC, Lloyd BB (eds). The regulation of human respiration. Oxford, Blackwell 1963 pp 331-349. 24. Read DJC. A clinical method for assessing the ventilatory response to carbon dioxide. Austral Ann Med 1966;16:20-32. 25. Jones RL,Neary JM, Ryan TG. Valores normales de la respuesta de la ventilacin hipercpnica : efectos de la edad y capacidad ventilatoria. Respiration 1994 ;4 :183-188 (edicin espaola). 26. Rebuck AS, Campbell EJM. A clinical method for assesing the ventilatory response to hypoxia. Am Rev Respir Dis 1974 ;109 :345-350. 27. Lourenco RV. Clinical methods for the study of regulation of breathing. Chest 1976;70:109-112. 28. Phillipson EA. Control of breathing during sleep Am Rev Respir Dis 1978;118:909-939. 29. Douglas NJ, White DP, Weil JV, Pickett CK, Zwillich CW. Hypercapnic ventilatory response in sleeping adults. Am Rev Respir Dis 1982; 126: 758762. 30. Garca Ro F Control de la respiracin.. Arch Bronconeumol 2004;40(supl 5):14-20. 31. American Thoracic Society. Idiopathic congenital central hypoventilation sndrome: diagnosis and management. Am J Respir Crit Care Med 1999;160:368-373.
EXPLORACION FUNCIONAL DE LOS SNDROMES DE HIPOVENTILACIN
173 32. Calverley PMA Control of breathing. Eur Respir Mon 2005;31:44-56 33. Cherniack NS, Altose MD. Respiratory responses to loading. In: Hornbein TK, ed. The Regulation of Breathing Part II. New York, Marcel Dekker Inc, 1981; pp. 905964. 34. de Lucas Ramos P, Rodrguez Gonzlez-Moro, Rubio Socorro Y. Obesidad y funcin pulmonar . Arch Bronconeumol 2004;40(supl 5): 27-31 35. Scano G, Gigliotti F, Duranti R, Gorini M, Fanelli A, Marconi G. Control of breathing in patients with neuromuscular diseases. Monaldi Arch Chest Dis 1993;48:87-91. 36. Fiz JA, Morera J. Exploracin funcional de los msculos respiratorios. Arch Bronconeumol 2000;36:391-410. 37. Galdiz Iturri JB. Aplicacin clnica de las pruebas de funcin muscular respiratoria. JM Pino Garca y F Garca Ro. Estudio de la funcin respiratoria. Msculos respiratorios. Sanitaria 2000. Madrid 2005 pp 149-174 38. Garcia-Ro F, Pino JM, Ruiz A, Daz S, Prados C, Villamor J. The accuracy of noninvasive estimates of respiratory muscle effort during spontaneous breathing in restrictive diseases J Appl Physiol 2003;95:1542-1549. 39. Pozzi E, Gulotta C. Classification of chest wall diseases. Monaldi Arch Chest Dis 1993;48:65-68. 40. Han JN, Gayan Ramirez G, Dekhuijzen R, Decramer M. Respiratroy function of the rib cage muscles. Eur Respir J 1993;6:722-728. 41. Baydur A, Milic-Emili J. Respiratory mechanics en kyphoscoliosis. Monaldi Arch Chest Dis 1993;48:69-79.
CAPTULO 9
SNDROME DE HIPERVENTILACIN
M TERESA RAMREZ PRIETO
Servicio de Neumologa. Hospital Universitario La Paz. Paseo de la Castellana 261-28046 Madrid
PALABRAS CLAVE: Sndrome de hiperventilacin, hipocapnia, alcalosis respiratoria, acidosis metablica, pH, disnea, taquipnea, tetania, taquicardia, provocacin mediante hiperventilacin, ansiedad, agorafobia, trastorno psiquitrico, cafena, -bloqueantes, benzodiacepinas, antidepresivos tricclicos, control de la respiracin, yoga, PCO2, cuestionario para hiperventilacin, cuestionario Nijmegen.
SNDROME DE HIPERVENTILACIN
176 NDICE DE CONTENIDOS: 1. Introduccin 2. Epidemiologa 3. Fisiopatologa 4. Diagnstico 5. Tratamiento 6. Conclusiones 7. Abreviaturas y unidades de medida 8. Bibliografa
SNDROME DE HIPERVENTILACIN
177 1. INTRODUCCIN El sndrome de hiperventilacin (SH) es una entidad no bien definida y que se vincula, al menos en parte, a los trastornos de ansiedad (1). Desde un punto de vista fisiolgico, se evidencia una respuesta ventilatoria excesiva, lo que conlleva un importante descenso del dixido de carbono alveolar (hipocapnia) y un incremento del pH en sangre (alcalosis respiratoria). Esta situacin mantenida en el tiempo, desencadena la aparicin de sntomas como disnea, palpitaciones, vrtigo, temblor distal, parestesias, dolor precordial, inquietud y disminucin del nivel de conciencia (2). El trmino sndrome de hiperventilacin, fue empleado por primera vez por Kerr et al (l3) en 1937, refirindose a una serie de sntomas inducidos por la hiperventilacin voluntaria e idnticos a los observados en las crisis de neurosis ansiosa. En sus estudios comprobaron que la hiperventilacin era el fenmeno comn y determinante de los sntomas de ambos procesos. Un ao despus, Soley y Shock (4) confirmaron la relacin entre el sndrome de hiperventilacin y la hiperventilacin voluntaria. En 1953 (5), Lewis estableci la diferencia entre la hiperventilacin aguda, caracterizada por ataques de sudoracin fra, sensacin de disnea, palpitaciones y tetania, de la hiperventilacin crnica, en la que los sntomas son ms leves y variados, percibidos por los pacientes como disnea, dolor opresivo abdominal y torcico, crisis de palpitaciones... Aos despus, Lum (6) encontr que la hiperventilacin crnica era ms frecuente que la aguda, guardando una ntima relacin con la ansiedad. Esta apreciacin fue crucial para el adecuado manejo y tratamiento de los sntomas asociados, donde los psicotropos juegan un papel fundamental. Ms recientemente, Dent el al (7) replantean el concepto de sndrome de hiperventilacin y sugieren el trmino desrdenes con hiperventilacin, refirindose a la compleja interaccin entre trastornos respiratorios, psiquitricos y psicolgicos, por lo que la hiperventilacin propia de enfermedades como el asma, las neumonas, el dolor o los trastornos del sistema nervioso central, formaran parte de l. En general, el trmino sndrome de hiperventilacin, ha sufrido importantes modificaciones desde su primera descripcin, donde se le consideraba similar a la neurosis obsesiva, hasta la introduccin del concepto de hiperventilacin crnica, el cual carece de los sntomas agudos y no siempre se encuentra relacionado con crisis de ansiedad. Los estudios realizados en los aos 80 en Noruega, Blgica y Gran Bretaa, establecen que el 35-83% de los SH son de origen ansioso (8-11) y que el 5-11% restante, son debidos a otros trastornos mdicos (12-14).
2. EPIDEMIOLOGA La incidencia o prevalencia del sndrome de hiperventilacin, nunca ha sido estudiada en la poblacin general. Los datos recogidos se encuentran relacionados con poblaciones muy concretas, como la publicada por McKell (15) que describe una prevalencia del 5,8% de hiperven-
SNDROME DE HIPERVENTILACIN
178 tilacin en un grupo de 500 pacientes sometidos a estudios gastroenterolgicos, o Rice (14) que observa hiperventilacin en un 10,7% de los ingresados a cargo del Servicio de Medicina Interna, siendo slo de un 4,5% en los ingresados en el Servicio de Urgencias (16). La hiperventilacin puede ocurrir tambin en nios, siendo muy frecuentemente empleada como arma para manipular a sus cuidadores y familiares, como fue el caso de una epidemia observada en una escuela elemental, que afect a numerosos nios sanos. Tras diversos estudios, se excluyeron enfermedades de base orgnica, lo que llev a solucionarlo con tratamiento psicolgico (17). La evolucin del sndrome de hiperventilacin infantil es variable, pero hasta un 40% lo conservan en la edad adulta (18). La relacin hombre/mujer de este sndrome en la poblacin general, es variable, aunque se encuentra en un rango de 1:1,1 en hombres frente al 1:6,5 en mujeres, con edades comprendidas entre los 15 y 55 aos (12). 3. FISIOPATOLOGA En el control de la ventilacin, el CO2 y el O2 van a actuar como estmulos qumicos, mediante la activacin de los quimiorreceptores tanto centrales como perifricos. En sujetos sanos, la presin parcial de CO2 (PCO2), va a ser el ms importante, mientras que la presin parcial de O2 (PO2), va a necesitar cadas de 40-50 mmHg para inducir una respuesta ventilatoria, siendo slo los quimiorreceptores perifricos sensibles a estos cambios. Tras la estimulacin de los quimiorreceptores perifricos, se observa una activacin de los centros respiratorios situados en el sistema nerviosos central, dando lugar a la correspondiente respuesta cardiaca, vasomotora, muscular autonoma y simptico-mimtica. Sobre los receptores centrales van a actuar tambin el hipotlamo (cetro de la termorregulacin, locomotor y emocional) y el cortex. Desde el sistema nervioso central, siguiendo la va bulboespinal, va a estimularse la motoneurona espinal, dando lugar a la respiracin autonmica. No obstante, existe otra va alternativa para la respiracin voluntaria a travs tracto piramidal. La respiracin en humanos no es slo una respuesta a las necesidades metablicas de O2 o CO2, es tambin una actividad mecnica, de hecho, si esta desarrollndose esta ltima funcin, el control qumico se va a interrumpir (19). As, cuando se desarrolla una actividad fsica importante, la ventilacin se incrementa sustancialmente en escasos segundos, antes incluso de que los quimiorreceptores detecten cambios en la PCO2, denominndose respuesta anticipada. Una repuesta similar se puede observar en los pacientes con sndrome de hiperventilacin sometidos a situaciones de estrs (20). Otra caracterstica distintiva de los centros respiratorios, es el fenmeno del volante por el que una vez estimulados, no podrn interrumpir su actividad de forma brusca, precisando un tiempo aproximado de 48 segundos para volver a su situacin basal. Este hecho fue puesto en evidencia por Swanson et al (21) que, tras exponer a siete sujetos
SNDROME DE HIPERVENTILACIN
179 sanos, en situacin de hiperventilacin voluntaria, a ambiente enriquecido con CO2 durante la inspiracin, hasta lograr valores de CO2 normales, encontraron un tiempo fijo de 22 segundos para la recuperacin de la frecuencia ventilatoria tras cesar la hiperventilacin. Otro grupo (22), llev a cabo un estudio similar pero modificando las concentraciones de CO2, observando que el tiempo de recuperacin guardaba una relacin inversamente proporcional a la PaCO2. Lo cual significa que, a menor concentracin de CO2, los centros respiratorios permanecen ms tiempo hiperactivos, una vez que han sido estimulados. Estos hallazgos, podran justificar el mayor grado de excitabilidad propia de las estructuras neuronales de los pacientes con hiperventilacin, de hecho, en estos casos, tras una hiperventilacin voluntaria, necesitan una mayor cantidad de tiempo para recuperar su PaCO2 (23). Hormonas como la progesterona, la norepinefrina y la tirosina son capaces de estimular los centros respiratorios. Ms an, la secrecin renal de epinefrina en los pacientes con sndrome de hiperventilacin, alcanza valores en torno a 15,8 17,5 g/24 horas, mientras que en sujetos sanos no sobrepasan los 6 g/24 horas. Esto podra, en parte, justificar los problemas de la hiperventilacin y justificar la indicacin de los adrenrgicos, especialmente por el hecho de que dichas drogas van a ser capaces de suprimir la actividad de los quimiorreceptores perifrficos (24). Otras drogas, como el cido-acetilsaliclico y la cafena, estimulan la ventilacin, hasta el punto de que la cafena va a ser capaz de desencadenar ansiedad y crisis de pnico (12). El alcohol, a su vez, va a ser capaz tambin de desencadenar crisis de hiperventilacin (25). Existen tres formas bsicas para el estudio de la capacidad de los centros respiratorios para mantener concentraciones constantes de CO2: 1) anlisis durante carrera libre, 2) estimulacin del sistema de feedback mediante el enriquecimiento del aire inspirado con CO2, 3) interferencia en el sistema de control a travs de la hiperventilacin voluntaria. Mediante el anlisis del CO2 durante la carrera libre, Vansteenkiste et al (26) observaron que, en 21 de los 37 sujetos estudiados por sospecha de hiperventilacin, la concentracin PCO2 descenda 0,25 kPa tras recuperar la frecuencia respiratoria. Igualmente, Hormbrey et al (27) evidenciaron irregularidades en la respiracin con fluctuaciones en el tiempo inspiratorio y espiratorio, as como en las concentraciones de la PCO2. Han (28) estudi el patrn respiratorio de 201 sujetos sanos frente a 580 con sospecha del sndrome de hiperventilacin, encontrando importantes irregularidades en el patrn respiratorio de estos ltimos, como exceso de suspiros y pausas al final de la inspiracin y la espiracin. La curva de respuesta al CO2 puede ser de gran utilidad en el estudio del sndrome de hiperventilacin, donde es caracterstico una respuesta ventilatoria invertida pues, tras administrar CO2, la respuesta ventilatoria en lugar de aumentar, se enlentece (29). Este fenmeno es anlogo al observado en los pacientes con hiperventilacin a los que se hace respirar en bolsas cerradas. Debido a que las crisis de pnico se asocian con la hiperventilacin, se ha estudiado el comportamiento del CO2 durante dichas crisis, coincidiendo los distintos grupos (30,31) en los
SNDROME DE HIPERVENTILACIN
180 datos obtenidos, donde destaca un leve desplazamiento hacia la izquierda (5 mmHg) del CO2, siendo similar a la observada en los pacientes con sndrome de hiperventilacin. Diversos autores han estudiado la homeostasis del CO2 tras la hiperventilacin voluntaria. La primera descripcin de los test de provocacin de la hiperventilacin, fue llevada a cabo por Weiman y Korschinsky (32), los cuales mantenan a los pacientes en estado de hiperventilacin durante 20 minutos en cuatro tiempos consecutivos, encontrando una fuerte correlacin entre la presin alveolar de CO2, los sntomas clnicos y los signos propios de tetania. A partir de ese momento, el reconocimiento de los sntomas se ha empleado como criterio diagnstico del sndrome de hiperventilacin. Posteriormente, Beumer and Hanrdonk (2,3) desarrollaron un test ms prctico, en el que se analizaba las concentraciones de CO2 a los 3 y 5 minutos tras interrumpir una hiperventilacin voluntaria de tres minutos. 4. DIAGNSTICO Existe una gran confusin en cuanto a los criterios diagnsticos del sndrome de hiperventilacin. Esto es debido a la ausencia de la tcnica que permita confirmar las sospechas con suficiente especificidad y sensibilidad. En general, si aceptamos la definicin clsica del sndrome de hiperventilacin, habrn de cumplirse los siguientes criterios: 1) presencia de hiperventilacin con descenso de los niveles de la PaCO2, 2) exclusin de enfermedades de base orgnica que se acompaen de hiperventilacin y 3) clnica justificable por la hipocapnia. En este sentido, el espectro de sntomas asociados a la hiperventilacin es extremadamente inespecfico y variable, pues puede afectar a cualquier rgano debido al descenso de los niveles de CO2 o a la excesiva respuesta del sistema adrenrgico. En la Tabla 1 quedan reflejados los sntomas ms comunes y su frecuencia. Respecto al primer criterio, es relativamente fcil analizar las concentraciones de CO2 tanto a nivel hospitalario como ambulante. La principal limitacin se encuentra en los casos con fobia o ansiedad, pues dichos procesos son fluctuantes y no siempre posibles de evidenciar en el momento de la extraccin de las muestras. La determinacin del CO2 de forma ambulante, mediante determinacin transcutnea, no es de fcil accesibilidad, y resulta poco precisa en episodios de hiperventilacin de escasa duracin, pues son necesarios dos minutos, como mnimo, para que los capnigrafos puedan hacer una determinacin fiable. En este sentido, se estn desarrollando nuevos test dirigidos al anlisis, no slo de las concentraciones de la PaCO2, sino de los cambios en los patrones respiratorios. La mayor limitacin de dichos test radica en su incapacidad para distinguir entre los sntomas debidos a la hipocapnia y aquellos relacionados con otros factores como el disconfort mecnico tras la hiperventilacin mantenida o el propio estrs de la prueba. En todo caso, debern descartarse aquellas entidades orgnicas capaces de generar hiperventilacin (Tabla 2), para lo cual es imprescindible una historia clnica meticulosa, pues son numerosas las posibilidades y no siempre fciles de diferenciar. En cuanto al tercer criterio, identificar los sntomas debidos a la hipocapnia, resulta el ms complejo, pues no es fcil discernir entre los sntomas debidos a la hipocapnia per se o al
SNDROME DE HIPERVENTILACIN
181
Sistema
Sntoma
Opresin torcica
Frecuencia relativa (%)
51-90 43-88 28-96 3-45 24-95 19-74 35-71 58-81 1-37 37-86 32-55 39-81 18-85 6-46 12-50 35-83 45-80 54-88 19-86 52-100 30-98
Respiratorio
Taquipnea Disnea Agarrotamiento muscular
Neuromuscular
Parestesias Hipertona digital Desconexin del medio Mareo
Cerebral
Temblor Cefalea Rubor facial Palpitaciones
Cardiocirculatorio
Dolor precordial Arritmia Epigastralgia
Gastrointestinal Nausea Hiperhidrosis General Astenia Insomnio Nerviosismo Psicolgico Ansiedad
Tabla 1. Sntomas asociados con el sndrome de hiperventilacin. Modificado de Folgering H (17).
SNDROME DE HIPERVENTILACIN
182
Asma Enfermedades pulmonares intersticiales Insuficiencia cardiaca congestiva Embolismo pulmonar Neumotrax Hipertermia Dolor intenso Cirrosis heptica Angina Abuso de cafena Intoxicacin por cido acetil saliclico Tumor cerebral Accidentes cerebrovascular Insuficiencia renal grave Acidosis metablica Embarazo Estimulacin vestibular Hernia diafragmtica gigante Anemia grave Tabla 2. Entidades clnicas acompaadas de hiperventilacin.
1. Historia clnica detallada. 2. Descartar entidades orgnicas que se acompaen de hiperventilacin. 3. Detectar los sntomas y signos propios del sndrome de hiperventilacin: taquipnea, taquicardia, opresin torcica, suspiros frecuentes, sudor fro, agarrotamiento de manos y pies. 4. Pruebas de laboratorio: espirometra forzada, gasometra arterial basal, respuesta ventilatoria al estmulo hipercpnico, test de hiperventilacin con determinacin de las concentraciones de PCO2 al final de la espiracin.
Tabla 3. Protocolo diagnstico para el sndrome de hiperventilacin. Tomado de Folguerin (17).
sndrome de hiperventilacin. En este sentido, Roll y Zetterquist (33), fueron los primeros en disear un test con capacidad para reconocer dichos sntomas. Posteriormente, Hornsveld et al (34) demostraron la disociacin entre los sntomas referidos por los pacientes y los niveles de PCO2 al final de la espiracin. Sin embargo, Vansteenkiste (26) encontr una buena concordancia entre los sntomas referidos por los pacientes sometidos a 1 minuto de hiperventilacin voluntaria y los descritos durante las supuestas crisis de hiperventilacin. En el grupo de agorafbicos estudiado por Bass et al (35), se observ que, los sntomas pro-
SNDROME DE HIPERVENTILACIN
183 vocados por varios minutos de hiperventilacin voluntaria y los referidos durante una crisis de pnico, eran idnticos. De esto se concluye que, referido a los criterios clsicos para el diagnstico del sndrome de hiperventilacin, quizs sera necesario redefinirlos o elaborar un nuevo protocolo que permitiera, de forma ms exacta, llegar al diagnstico del mismo. Un posible protocolo es el propuesto por Folgerin (14) (tabla 3). Segn este protocolo, el primer paso para el diagnstico es una adecuada historia clnica, donde deber quedar reflejada el carcter, la intensidad y duracin de los sntomas, los posibles factores precipitantes (fobias, ansiedad) y, si es posible, invitar al paciente a reproducir un episodio. Para la realizacin de la historia clnica, puede ser til alguno de los cuestionarios elaborados a tal efecto. En concreto, el realizado por Doorn et al (36), denominado cuestionario Nijmegen (Tabla 4), fue validado con 163 pacientes y 100 controles. Dicho cuestionario consta de 16 preguntas, con cuatro posibles respuestas, medidas de 0 (nunca) a 4 (muy frecuente). Un resultado superior a 23, detecta los trastornos respiratorios con un valor predictivo del 80%. Dicho cuestionario ha sido validado posteriormente por Dixhoorn (37) y por Vansteenkiste (26), vindose que, a parte de ser de gran utilidad para el diagnstico del sndrome de hiperventilacin, podra emplearse con buena rentabilidad para monitorizar la respuesta al tratamiento. Una vez establecida la sospecha clnica, ser imprescindible descartar todas aquellas enfermedades de base orgnica que pudieran presentar hiperventilacin (Tabla 2), para lo que se llevarn a cabo las medidas diagnsticas precisas: radiografa simple de trax, tomografa axial computarizada, anlisis de sangre y orina completos... Una exploracin fsica minuciosa puede ser de gran utilidad, pues suele ser rigurosamente normal en el sndrome de hiperventilacin y, sin embargo, en caso de existir una enfermedad orgnica de base, se podran detectar signos propios de cada uno de ellas. Una vez superado los pasos previos, sern los exmenes en el laboratorio de exploracin funcional respiratoria, los que permitan confirmar la sospecha clnica. As, habr de determinarse en primer lugar la PCO2 al final de la espiracin, para lo cual se podr emplear un espirmetro o un pletismgrafo. La mayor limitacin de estas tcnicas, es la necesidad de emplear boquillas o mascarillas, que pueden modificar el patrn respiratorio, por lo que la pletismografa inductiva puede ser una alternativa. En esta tcnica, se emplearn una banda torcica y otra abdominal para realizar las determinaciones, por lo que se generarn menos interferencias en el patrn respiratorio. Con esta tcnica, sin embargo, va a ser muy difcil calibrar la amplitud de los movimientos con el volumen corriente, y casi imposible mantener dicha calibracin. Los patrones respiratorios sern evaluados en trminos de; ventilacin minuto, nmero de suspiros, regularidad o irregularidad en el patrn respiratorio y modificaciones en la PCO2 al final de la espiracin. Para analizar el comportamiento del CO2, existen distintos tipos de pruebas de provocacin de la hiperventilacin (HVPT). Estas pruebas, van a tener como objetivo reproducir los sntomas referidos por los pacientes mediante una hiperventilacin
SNDROME DE HIPERVENTILACIN
184
Sntomas
Dolor torcico Tensin emocional Visin borrosa Mareo Confusin Taquipnea Disnea Opresin torcica Dolor abdominal Temblor distal Imposibilidad para inspiracin profunda Rigidez de dedos y/o manos Manos fras Taquicardia Ansiedad
Puntuacin
Total:
Tabla 4. Cuestionario Nijmegen para la deteccin del sndrome de hiperventilacin. Tomado de Doorn (36).
SNDROME DE HIPERVENTILACIN
185 voluntaria, de varios minutos de duracin, en los que el paciente ser monitorizado en el laboratorio de exploracin funcional (39,40). La mayor limitacin para las HVPT, radica en su incapacidad para distinguir entre los sntomas producidos por la hipocapnia, por el estrs de la prueba o el disconfort que genera la hiperventilacin. Una de las HVPT ms empleada, consiste en mantener una hiperventilacin voluntaria de 1 3 minutos, hasta que la PCO2 al final de la espiracin descienda por debajo de 20 mmHg (2,6 KPa). En ese momento, se analizarn los cambios en la frecuencia respiratoria, en la PaCO2 y su capacidad de recuperacin hasta valores basales. Los datos as obtenidos, se contrastarn con los sntomas y signos percibidos por los pacientes. El grupo de Gardner (41), describi otro tipo de HVPT en la que estimulaba la hiperventilacin mediante una marcha de 5-10 minutos en tapiz rodante. Tras el ejercicio, se meda la PCO2 al final de la espiracin, debiendo ser superior a 4 kPa (30 mmHg) a los cinco minutos de cesar el ejercicio en los sujetos sanos. Aunque las HVPT pueden ser de utilidad en el estudio del sndrome de hiperventilacin, ninguna ha demostrado suficiente sensibilidad y especificidad como para establecer su diagnstico. Dada la arbitrariedad en este sentido, algunos grupos (42) han elaborado un protocolo diagnstico, basado en signos asociados a los trastornos en la ventilacin que pudieran justificar la hiperventilacin (Tabla 5), sin valorar los sntomas clnicos. Segn este grupo, la identificacin de 3 o ms sntomas permitir la aproximacin diagnstica del sndrome de hiperventilacin. Dadas las limitaciones de las HPVT, y ya que la mayora de las crisis de hiperventilacin se desarrollan en la vida diaria del paciente, el grupo de Hornsveld (2), desarroll dos estudios paralelos basados en la monitorizacin continua de los niveles de CO2 mediante cateterizacin arterial. En el primer estudio se monitorizaron, de forma ambulante, los cambios en la PaCO2 de 28 suje1. Niveles bajos de PCO2 arterial/ final de espiracin. 2. Patrn respiratorio irregular. 3. Taquipnea (>12/min). 4. Niveles PetCO2 bajos en condiciones basales. 5. Descenso de la ventilacin minuto/frecuencia respiratoria cuando la PetCO2 se eleva 0,7-1,2 kPa. 6. Prueba de provocacin: la determinacin del capniograma al minuto de la ventilacin voluntaria muestra uno de los siguientes datos; no cambios en PetCO2. 7. No cambios en la frecuencia respiratoria. 8. Ausencia de recuperacin de la PetCO2 tras tres minutos de cesar la hiperventilacin hasta un mnimo del 90% del valor basal. Tabla 5. Signos de trastornos en la ventilacin responsables de hiperventilacin. Modificado de Folgerin (34).
SNDROME DE HIPERVENTILACIN
186 tos diagnosticados de agorafobia. En el segundo estudio, el anlisis se llev a cabo en sujetos con sospecha de sndrome de hiperventilacin y con una prueba de HPVT positiva. Los resultados obtenidos fueron llamativos pues, en el grupo con diagnstico de agorafobia, tan slo en un caso se evidenci un leve descenso la PaCO2, mientras que en el grupo con estudio de HPVT positivo, slo en 7 de las 22 crisis referidas, hubo un leve descenso de la PaCO2. En ambos grupos, no hubo diferencias significativas en la incidencia de falsos positivos y falsos negativos. La ausencia de un gold standard para el diagnstico del sndrome de hiperventilacin, hace que no se pueda ni excluir ni confirmar sin tener en cuenta todos los datos clnicos y funcionales antes expuestos, siendo necesarios ms estudios en este campo.
5. TRATAMIENTO La aproximacin teraputica al sndrome de hiperventilacin, se fundamenta en tres campos; fisioterapia, psicoterapia y tratamiento farmacolgico. La eleccin de cada uno de ellos depender de la gravedad del caso, siendo necesario, en ocasiones, el empleo de los dos y siempre con el objetivo de erradicar los sntomas. El primer paso en el tratamiento, ser la adecuada explicacin al paciente de su proceso, pues el reconocimiento de los sntomas y su origen facilita el control de los mismos. En este sentido, la capacidad para reproducir los sntomas mediante alguna da las HPVT, ser un indicador de buena respuesta al tratamientor (42). El empleo de ciertas estrategias como, respiracin en bolsa cerrada, tcnicas para aumentar el espacio muerto o el entrenamiento de la respiracin, pueden ser de gran utilidad en los procesos agudos. El entrenamiento de la respiracin es una de las modalidades teraputicas ms empleadas y con mejores resultados. Su fundamento consiste en; 1) reducir la hiperventilacin durante la fase aguda, 2) relajacin y autocontrol mediante ejercicios respiratorios similares a los empleados en Yoga. Muchos terapeutas, han insistido en la eficacia de la tcnica de respiracin mediante el empleo de la parte superior de la caja torcica, lo cual supone realizar dos tercios de la inspiracin con el diafragma y el tercio restante, con los msculos intercostales y escalenos. No obstante, otros terapeutas, como el grupo de Troyer (43), han considerado a la respiracin diafragmtica una falacia pues, si este tipo de respiracin es la consecuencia de la estimulacin de las gamma-motoneurona, al realizar la respiracin abdominal, se podra modificar el patrn respiratorio pero no la frecuencia respiratoria. Para llegar a esta conclusin, realizaron un estudio en el que monitorizaron, tanto en sujetos sometidos a respiracin abdominal como en sujetos sanos, los cambios en la presin pleural, la presin gstrica, la actividad electromiogrfica de los msculos intercostales y el diafragma, as como los movimientos de la caja torcica y el abdomen. Con este estudio demostraron que, aunque con la respiracin abdominal se incrementaba la amplitud de movimientos de la pared anterior del abdomen, al mismo tiempo se incrementaba la actividad electromiogrfica de los msculos intercostales, sin diferencias en el control de la frecuencia respiratoria respecto del grupo control.
SNDROME DE HIPERVENTILACIN
187 En el control de la frecuencia respiratoria, otros grupos se han valido de ejercicios similares a los del yoga (44), donde se realizan movimientos respiratorios a muy baja frecuencia, trescuatro por minuto. Tras un buen entrenamiento, los pacientes alcanzan una ventilacin minuto muy baja, con valores ms elevado de PCO2. Este tipo de respiracin, ha sido empleado en un programa multidisciplinar (42-46), demostrando ser muy eficaz en el control de los sntomas durante crisis de pnico, gracias a la capacidad de mantener los niveles de PCO2. Diversos autores han analizado la utilidad de frmacos, como los -bloqueantes-adrenrgicos, las benzodiacepinas o los antidepresivos tricclicos en el control de los sntomas del sndrome de hiperventilacin, con resultados prometedores. As, los -bloqueantes-adrenrgicos, con su capacidad para controlar la actividad del sistema nervioso simptico, podran controlar los sntomas al incrementar los niveles de PaCO2, como as puso en evidencia el estudio caso-control llevado a cabo por Folgering (47), y ratificado posteriormente por el mismo grupo con un estudio no controlado (48). No obstante, el efecto de los -bloqueantes-adrenrgicos, aunque inicialmente mejoran los sntomas al mantener elevadas las cifras de la PaCO2, no se van a mantener una vez interrumpido el tratamiento. Estos resultados parecen concordantes con los publicados por Van den Ven (49), que estudio los efectos del bisoprolol en 60 pacientes con sndrome de hiperventilacin, mostrndose eficaz en el control de los sntomas, frecuencia y gravedad de los ataques. Otros frmacos, como los antidepresivos tricclicos, aunque menos estudiados, parecen capaces de reducir la intensidad de los sntomas (50). Igualmente, las benzodiacepinas, gracias a su capacidad para relajar la musculatura y deprimir los centros respiratorios, podran mejorar la intensidad de las crisis de hiperventilacin (51). 6. CONCLUSIONES Clsicamente, el sndrome de hiperventilacin, se conoce como un complejo de sntomas inducidos por el incremento de la frecuencia respiratoria y reproducible mediante maniobras de hiperventilacin voluntaria. No existe ningn sntoma clnico especfico del mismo, de hecho puede afectar a cualquier rgano, sin embargo, los ms frecuentes son la taquipnea, la taquicardia, la opresin torcica, la rigidez de miembros y el mareo. Aunque dichos sntomas, parecen guardar una clara relacin con el descenso en las cifras de la PaCO2, tambin pueden desencadenarse ante cambios en la misma, pero mantenindose dentro de los rangos de la normalidad. El diagnstico de esta entidad ofrece grandes dificultades pues, aunque en los ltimos aos se han desarrollado tcnicas diagnsticas, como la provocacin de hiperventilacin o la monitorizacin ambulante de las cifras de la PaCO2, ninguna ha demostrado suficiente sensibilidad y especificidad. En este sentido, se han elaborado protocolos diagnsticos (tabla 3), donde se valorarn los resultados de estas pruebas y los datos clnicos referidos por el paciente, excluyendo siempre cualquier otra enfermedad orgnica que pudiera justificar la hiperventilacin.
SNDROME DE HIPERVENTILACIN
188 Desde un punto de vista teraputico, la combinacin de psicoterapia, rehabilitacin respiratoria, con especial nfasis en el control de la frecuencia respiratoria, y frmacos, como los -bloqueantes, antidepresivos tricclicos o las benzodiacepinas, parecen ofrecer los mejores resultados. No obstante, de todo lo expuesto se deduce que, el sndrome de hiperventilacin es un gran desconocido, que se ha empleado durante mucho tiempo como cajn de sastre para todos aquellos procesos clnicos que cursaban con hiperventilacin aadida y sin otra causa justificable ms que los trastornos por ansiedad, siendo necesarios ms estudios que permitan un mayor entendimiento y control del mismo.
SNDROME DE HIPERVENTILACIN
189 7. BIBLIOGRAFA 1. Woodhead MA, Mcfarlane JT, McCracken JS, Rose DH, Finch RG. Prospective study of the aetiology and outcome of pneumonia in the community. Lancet 1987;1:671-4 2. Hornsveld H, Garssen B. Hyperventilation syndrome: an elegant but scientifically untenable concept. The Netherlands Journal of Medicine 1997;50:13-20 3. Kerr WJ, Dalton JW, Gliebe PA. Some physical phenomena associated with the anxiety states and their relation to hyperventilation. Ann Inter Med 1937;11:961-992 4. Soley M, Shock N. The aetiology of effort syndrome. Am J Med Sci 1938;196:840-851 5. Lewis BJ. The hyperventilation syndrome. Ann Intern Med 1953;38:918-927 6. Lum LC. Hyperventilation, the tip and the iceberg. J Psychosom Res 1975;19:375-383 7. Dent R, Yates D, Higginbottam T. Does the hyperventilation syndrome exist?. Thorax 1983;38:223 8. Cowley DS, Roy-Byrne. Hyperventilation and panic disorder. Am J Med 1987;83:929-937 9. Garssen B, Van Veenendall W, Bloemink R. Agoraphobia and the hyperventilation syndrome. Behav Res Ther 1983;21:643-649 10. de Ruiter C, Garssen B, Rijken H, Kraaimaat F. The hyperventilation syndrome in panic disorder, agoraphobia, and generalized anxiety disorder. Behav Res Ther 1989;27:447-452 11. Rapee R. Differential response to hyperventilation in panic disorder and generalized anxiety disorder. J Abnom Psychol 1986;95:24-28 12. Brashear RE. Hyperventilation syndrome. Lung 1938;161:257-273 13. Lum LC. The syndrome of habitual chronic hyperventilation. In: Hill OW, ed. Modern trends in psychosomatic medicine. London: Butterworths, 1976:196-230 14. Rice RL. Sympton patterns of the hyperventilation syndrome. Am J Med 1950;8:691-700 15. McKell TE, Sullivan AJ. The hyperventilation syndrome in gastroenterology. Gastroenterology 1947;9:6-16
SNDROME DE HIPERVENTILACIN
190 16. Dlle W. Hyperventilation und hyperventilation syndrome. Med Klin 1964;59:695-699 17. Folgering H. Lung biology in health and disease. Control of breathing in health and disease. Altose M.D, Kawakami Y. Edit. The hyperventilation syndrome. Marcel Dekker, Inc.;1999:633-660 18. Herman SP, Stickler GB, Lucas AR. Hyperventilation syndrome in children and adolescents: a long term follow-up. Pediatrics 1981;67:183-187 19. Phillipson EA, McClean PA, Sullivan CE, Zamel N. Interaction of metabolic and behavioural respiratory control during hypercapnia and speech. Am Rev Respir Dis 1978;117:903-909 20. Wasserman K, Hansen JE, Sue DY, Whipp BJ, Casaruby R. Principles in exercise testing and interpretation. Malvern, PA:Lea &Febiger, 1994 21. Swason GD, Wards DS, Bellville JW. Posthyperventilation isocpnica hyperpepnea. J Appl Physiol 1976;40:592-596 22. Folgerin H, Durlinger M. The time-course of post-hyperventilation breathing in man depends on PaCO2. J Appl Physiol 1983;54:809-813 23. Beumer HM, Hardonk HJ. Symptomatic und behandlung des Hyperventilationssyndroms. Munch Med Wochenschr 1971;113:1255-1258 24. Folgerin H, Ponte J, Sadig T. Adrenergic mechanisms and chemoreception in the carotid body of the cat and rabbit. J Physiol (Lond) 1982;325:1-21 25. Roelofs SM, DIkkenberg GM. Hyperventilation and anxiety: alcohol withdrawal symptoms decreasing with prolonged abstinence. Alcohol 1987;4:215-220 26. Vansteenkiste J, Kochette F, Demedts M. Diagnostic tests of hyperventilation syndrome. Eur Respir J 1991;4:393-399 27. Hormbrey J, Jacobi MS, Patil CP, Saunders KB. CO2 response and pattern of breathing in patients with symptomatic hyperventilation, compared to asthmatic and normal subjects. Eur Respir J 1988;1:846-852 28. Han JN. Breathing patterns in anxiety disorders; is there a hyperventilation syndrome? Dissertation, University of Leuven, Belgium, 1995 29. Folgerin H, Colla P. Some anomalies in the control of PaCO2 in patients with a hyperventilation syndrome. Bull Eur Physiopathol Resp 1978;14:505-512 30. Woods SW, Charney DS, Loke J, Goodman WK, Redmond DE, Heninger GR. Carbon dioxide sensitivity in panic anxiety. Arch Gen Psychiatry 1986;43:900-909
SNDROME DE HIPERVENTILACIN
191 31. Zandbergen J, Pols H, de Loof C, Griez EJL. Ventilatory response to CO2 in panic disorder. Psychiatry Res 1991;39:13-19 32. Weimann G, Korskinsky H. Durchfhdrung und Bewertung des Hyperventilations-versuchs. Med Klin 1970;65:56-62 33. Roll MI, Zetterquist A. Acute chest pain without obvious organic cause before the age of 40: response to forced hyperventilation. J Int Med 1990;228:223-228 34. Hornsveld H, Garssen B, Fiedeldij DpM, van Spiegel P, de Haes H. Double blind placebo controlled study on the hyperventilation provocation test: questioning the concept validity of the hyperventilation syndrome. Dissertation, University of Amsterdam, The Netherlands, 1996;8:113-121 35. Bass C, Lelliott P, Marks I. Fear talk versus voluntary hyperventilation in agophobics and normal. Psychiatr Med 1989;19:669-676 36. Van Doorn P, Colla P, Folgering H. Een vragenlijst voor hyperventilatieklachten. Psycholoog 1983;18:573-577 37. van Dixhoorn J, Duivenvoorden HJ. Efficacy of Nijemegen questionnaire in recognition of the hyperventilation syndrome. J Psychosom Res 1985;29:199-206 38. Folgering H, Jongmans M, Cox N, Dekhuijzen P. Treatment of hyperventilation during pulmonary rehabilitation. Eur Respir J 1991;4:222s 39. Hornsveld H, Garssen B, Fieldij Dop M, van Spiegel P. Symptom reporting during voluntary hyperventilation and mental load: implications for diagnosis hyperventilation syndrome. J Psychosom Res 1990;34:687-697 40. Lewis RA, Howell JBL. Definition of hyperventilation syndrome. Bull Eur Physiopathol Respir 1986;22:201-205 41. Gardner WN. Diagnosis and organic causes of symptomatic hyperventilation. In: Timmons BH, Ley R, eds. Behavioural and psychological approaches to breathing disorders. New York: Plenum 1994;6:99-112 42. Salkovskis PM, Jones DR, Clark DM. Respiratory control in treatment of panic attacks: replication and extension with concurrent measurement of PCO2. Br J Psychiatry 1986;148:526-532 43. de Troyer A, Samspon MG. Activation of the parasternal intercostals during breathing efforts in human subjects. J Appl Physiol 1982;52:524-529 44. Stanescu DC, Nemeyer B, Veriter C, Marechal C. Pattern of breathing and ventilatory response to CO2 in subjects practising hatha-yoga. J Appl Physiol 1981;57:1625-1629
SNDROME DE HIPERVENTILACIN
192 45. Clark DM, Salkovskis PM, Chalkley AJ. Respiratory control as a treatment for panic attacks. J Behav Ther Exp Psychiatry 1985;16:23-30 46. van Doorn P, Folgering H, Colla P. Control of the end-tidal PCO2 in the hyperventilation syndrome: effects of biofeedback and of breathing instructions compared. Clin Respir Physiol 1982;18:829-836 47. Folgering H, Cox A. Beta-Bloker therapy with metroprolol in the hyperventilation syndrome. Respiration 1981;41:33-38 48. Folgering H, Rutten H, Roumen Y. Beta-Blockade in the hyperventilation syndrome; a retrosprective assessment of symptoms and complaints. Respiration 1983;44:19-25 49. van de ven LLM, Mouthaan BJ, Hoes MJAJM. Treatment of the hyperventilation syndrome with bisoprolol: a placebo-controlled clinical trial. J Psychosom Res 1995;39:1007-1013 50. Hoes MJAM, Colla P, Folgering H. Clomipramine treatment of hyperventilation. Pharmakopsychiatry 1980;13:25-28 51. Aronson PR. Evaluation of psychotropic drug therapy in chronic hyperventilation syndrome. J New Drugs 1966;6:305-307
You might also like
- Conceptos Y Estrategias En Terapia RespiratoriaFrom EverandConceptos Y Estrategias En Terapia RespiratoriaRating: 4 out of 5 stars4/5 (1)
- Control de La Respiracion PDFDocument10 pagesControl de La Respiracion PDFa_ba_soloNo ratings yet
- Artìculo de InvestigaciònDocument11 pagesArtìculo de Investigaciònkatherin pichardoNo ratings yet
- TRABAJO Regulacion VentilatoriaDocument15 pagesTRABAJO Regulacion VentilatoriasierramenesesfranyersondanielNo ratings yet
- Cap. 41 GuytonDocument3 pagesCap. 41 GuytonNt Gaff100% (2)
- Regulacion de La RespiracionDocument17 pagesRegulacion de La RespiracionLaurita LpNo ratings yet
- Tema 8.1.x. Manual SEPAR - La Respiración Durante El SueñoDocument11 pagesTema 8.1.x. Manual SEPAR - La Respiración Durante El SueñoDavid BudauNo ratings yet
- Manual RespiratorioDocument18 pagesManual RespiratorioArmando MozquedaNo ratings yet
- Control de La RespiracionDocument4 pagesControl de La RespiracionHannia AguilarNo ratings yet
- Fisiología Respiratoria: El Control de La RespiraciónDocument5 pagesFisiología Respiratoria: El Control de La RespiraciónJorge LopezParraNo ratings yet
- Regulación de La Respiración PDFDocument11 pagesRegulación de La Respiración PDFpropranolol123100% (1)
- Practica 15 Funcion PulmonarDocument9 pagesPractica 15 Funcion PulmonarAitana PiñaNo ratings yet
- Regulación de Las Funciones Del Aparato Respiratorio TocaraDocument6 pagesRegulación de Las Funciones Del Aparato Respiratorio TocarafranciscoNo ratings yet
- La Respiración Durante El SueñoDocument11 pagesLa Respiración Durante El SueñosoNo ratings yet
- Fisiologia Respiratoria en El NiñoDocument89 pagesFisiologia Respiratoria en El NiñoJaviNo ratings yet
- Guia Taller Ventilacion Mecanica EsucomexDocument34 pagesGuia Taller Ventilacion Mecanica Esucomexalyzabeth23No ratings yet
- Regulacion de La RespiracionDocument3 pagesRegulacion de La RespiracionAlessandra Maldonado ChavezNo ratings yet
- Respiraciones Periódicas y Otras Alteraciones Del SueñoDocument27 pagesRespiraciones Periódicas y Otras Alteraciones Del SueñosoNo ratings yet
- Fisiologia II 2013Document32 pagesFisiologia II 2013Alan AguilarNo ratings yet
- Binomio Pulmon-CerebroDocument6 pagesBinomio Pulmon-CerebroLizbeth EmpanaditaNo ratings yet
- Sistema de control respiratorio: estructura y relacionesDocument8 pagesSistema de control respiratorio: estructura y relacionesWilford ChaguaNo ratings yet
- Fisioterapia-Respiratoria-en-Pediatria--2017Document53 pagesFisioterapia-Respiratoria-en-Pediatria--2017Ezequiel SainzNo ratings yet
- Respiracion WLCDocument19 pagesRespiracion WLCWilson LizanaNo ratings yet
- Práctica 11Document5 pagesPráctica 11Francisco Gutierrez PardoNo ratings yet
- Farmacoterapia Del Sistema RespiratorioDocument18 pagesFarmacoterapia Del Sistema RespiratorioLiz Ramoz0% (1)
- Chiroque Sem9Document31 pagesChiroque Sem9Marie Claire Pacheco ParedesNo ratings yet
- Perspectiva Del Cuidado RespiratorioDocument151 pagesPerspectiva Del Cuidado RespiratorioDARLYN STHEFANY RODRIGUEZ MAYORGANo ratings yet
- Silabo - Fisiologia Humana 2014Document11 pagesSilabo - Fisiologia Humana 2014USMP FN ARCHIVOSNo ratings yet
- RESPIRATORIODocument4 pagesRESPIRATORIOEVa SandryNo ratings yet
- Mecánica respiratoria: volúmenes pulmonaresDocument23 pagesMecánica respiratoria: volúmenes pulmonaresEduardo Andrés Cifuentes SilvaNo ratings yet
- Practica #03 "Espirometría. Volúmenes y Capacidades Pulmonares. Interpretación."Document21 pagesPractica #03 "Espirometría. Volúmenes y Capacidades Pulmonares. Interpretación."Jesus Guido Orestes Ojeda TorresNo ratings yet
- Fisioterapia RespiratoriaDocument9 pagesFisioterapia RespiratoriaArturo LoSa100% (1)
- Regulación de La Respiración - Organización Morfofuncional de Su Sistema de ControlDocument11 pagesRegulación de La Respiración - Organización Morfofuncional de Su Sistema de ControlCarlos AvilaNo ratings yet
- UNIDAD IV Funcion PulmonarDocument4 pagesUNIDAD IV Funcion PulmonarCarlos Erpel SandovalNo ratings yet
- °3 - Regulación de La Respiración PDFDocument20 pages°3 - Regulación de La Respiración PDFKarolay Fiestas PaicoNo ratings yet
- Fisiologia RespiratoriaDocument6 pagesFisiologia RespiratoriaPedro ZamoraNo ratings yet
- Procedimientos 4Document145 pagesProcedimientos 4Esteban SalinasNo ratings yet
- Regulacion Durante El Ejercicio Proc 1Document15 pagesRegulacion Durante El Ejercicio Proc 1JechîNo ratings yet
- Modulo MorfofisiologiaDocument214 pagesModulo MorfofisiologiaJohn VidesNo ratings yet
- Insuf. Respiratoria NeonatalDocument11 pagesInsuf. Respiratoria NeonatalJoshua FaudoaNo ratings yet
- Protocolo EspirometriaDocument24 pagesProtocolo EspirometriaAngieNo ratings yet
- Farmacología respiratoriaDocument35 pagesFarmacología respiratoriaLuis Manuel100% (1)
- Mód CN A 5 - 3bgu P4Document66 pagesMód CN A 5 - 3bgu P4Alisson GuzhñayNo ratings yet
- Contenido Semana 4 Salud LaboralDocument23 pagesContenido Semana 4 Salud Laboralpedro enrique monardez camposNo ratings yet
- Tos - Acercamiento A La Fisiopatología, Enfoque y Tratamiento - Rev Colomb Neumol 2009Document7 pagesTos - Acercamiento A La Fisiopatología, Enfoque y Tratamiento - Rev Colomb Neumol 2009ErnestoNo ratings yet
- Regulación PulmonarDocument6 pagesRegulación PulmonarIvaniuska GuerraNo ratings yet
- Insuficiencia Respiratoria AgudaDocument13 pagesInsuficiencia Respiratoria AgudaEliud SánchezNo ratings yet
- Fisiologia de Los Sistemas EspecialesDocument6 pagesFisiologia de Los Sistemas EspecialesVictor RiveraNo ratings yet
- 5.guia de Actividades Modulo 1 - 230817 - 081457Document9 pages5.guia de Actividades Modulo 1 - 230817 - 081457Mabel PatiñoNo ratings yet
- Practica de Fisio de Regulacion RespiratoriaDocument20 pagesPractica de Fisio de Regulacion RespiratoriaChristian Andre Ocaña ParedesNo ratings yet
- Historia clínica en Neumología: Anamnesis y exploración físicaDocument27 pagesHistoria clínica en Neumología: Anamnesis y exploración físicaPUBG Mobile TutorialesNo ratings yet
- Control de La VentilaciónDocument2 pagesControl de La VentilaciónRonaldo GuaoNo ratings yet
- Atrapamiento aéreo EPOCDocument9 pagesAtrapamiento aéreo EPOCCristian BenayNo ratings yet
- Regulaciòn de La RespiraciònDocument6 pagesRegulaciòn de La RespiraciònAndresiño Lima ArqqueNo ratings yet
- Facultad Ciencias de La Salud Escuela Profesional de OdontologiaDocument29 pagesFacultad Ciencias de La Salud Escuela Profesional de OdontologiaAndrea SugashimaNo ratings yet
- Determinación de la capacidad pulmonar y espirometríaDocument69 pagesDeterminación de la capacidad pulmonar y espirometríaGuillermo ChirinosNo ratings yet
- Generación del Ritmo RespiratorioDocument69 pagesGeneración del Ritmo RespiratorioJose Antonio TorresNo ratings yet
- Fisiología Del S. Respiratorio 4Document38 pagesFisiología Del S. Respiratorio 4Shierlley SantiNo ratings yet
- Reporte 1: Laboratorio de Fisiología Ii María de Los Ángeles Benítez Escobedo 4C #06 Dr. José Anselmo Hernández IbarraDocument7 pagesReporte 1: Laboratorio de Fisiología Ii María de Los Ángeles Benítez Escobedo 4C #06 Dr. José Anselmo Hernández Ibarra9r4hp288bsNo ratings yet
- Guia 8 Hormonas y ReceptoresDocument8 pagesGuia 8 Hormonas y ReceptoresJhon GutierrezNo ratings yet
- Leucemias 1Document90 pagesLeucemias 1Thalia LRNo ratings yet
- Consenso Fasgo 2017 Hipertension y EmbarazoDocument21 pagesConsenso Fasgo 2017 Hipertension y EmbarazoFranklin Vasquez BurgosNo ratings yet
- Devolucion de TurnoDocument1 pageDevolucion de TurnoThalia LRNo ratings yet
- 140-Article Text-539-8-10-20191001Document6 pages140-Article Text-539-8-10-20191001Sheila Cruz RodriguezNo ratings yet
- ConaremeDocument107 pagesConaremeThalia LRNo ratings yet
- Emergencias OncologicasDocument57 pagesEmergencias OncologicasThalia LRNo ratings yet
- Administracion Financiera Capitulo 1 Fundamentos Del Capital de TrabajoDocument34 pagesAdministracion Financiera Capitulo 1 Fundamentos Del Capital de TrabajoHaroldo Medrano L0% (1)
- Energia Solar XD 2Document1 pageEnergia Solar XD 2Thalia LRNo ratings yet
- 04 - Fracturas Del CalcaneoDocument54 pages04 - Fracturas Del CalcaneoThalia LRNo ratings yet
- Cambios Anátomo Fisiológicos en La Infancia y La Adolescencia 07Document42 pagesCambios Anátomo Fisiológicos en La Infancia y La Adolescencia 07Thalia LR0% (1)
- 3 Manejo Basico de La Via AereaDocument36 pages3 Manejo Basico de La Via AereaPablo PachNo ratings yet
- Ecogafía y angiografía ocular enDocument27 pagesEcogafía y angiografía ocular enThalia LRNo ratings yet
- Giardia Lamblia - Ciclo de EvolucionDocument2 pagesGiardia Lamblia - Ciclo de EvolucionThalia LRNo ratings yet
- 6 NeuropsicologiaDocument28 pages6 NeuropsicologiaLuis Ivan Pineda KunihiroNo ratings yet
- PG 65-69Document5 pagesPG 65-69Thalia LRNo ratings yet
- Diabetes: clasificación, factores y epidemiologíaDocument28 pagesDiabetes: clasificación, factores y epidemiologíarock2312No ratings yet
- Matriz EducativaDocument2 pagesMatriz EducativaThalia LRNo ratings yet
- Anatomia Del Aparato Genital FemDocument36 pagesAnatomia Del Aparato Genital FemThalia LRNo ratings yet
- Anexo N1Document3 pagesAnexo N1Thalia LRNo ratings yet
- CorClase 31 - Cancer de EndometrioDocument28 pagesCorClase 31 - Cancer de EndometrioThalia LRNo ratings yet
- CA Vulva VaginaDocument34 pagesCA Vulva VaginaThalia LRNo ratings yet
- EVALUACI+ôN DEL ESTADO NUTRICIONALDocument46 pagesEVALUACI+ôN DEL ESTADO NUTRICIONALThalia LRNo ratings yet
- Cri Cri CriDocument32 pagesCri Cri CriThalia LRNo ratings yet
- Cacervixfinal 090810205016 Phpapp01Document38 pagesCacervixfinal 090810205016 Phpapp01juancarlos_sebastiánNo ratings yet
- Traumahepatico 100405051810 Phpapp01Document23 pagesTraumahepatico 100405051810 Phpapp01Thalia LRNo ratings yet
- Em Briolo GiaDocument18 pagesEm Briolo GiaThalia LRNo ratings yet
- CorClase 32 - Cancer de OvarioDocument31 pagesCorClase 32 - Cancer de OvarioThalia LRNo ratings yet
- Manual de Semiología en Quechua PDFDocument41 pagesManual de Semiología en Quechua PDFThalia LRNo ratings yet
- TQ 18Document42 pagesTQ 18Thalia LRNo ratings yet
- BTC IntroduccionDocument9 pagesBTC IntroduccionThalia LRNo ratings yet
- Hidrólisis de Las Proteínas Concentradas en La Harina de QuinuaDocument3 pagesHidrólisis de Las Proteínas Concentradas en La Harina de QuinuaSebastian Castillo VillNo ratings yet
- Tema 6 Señales QuímicasDocument20 pagesTema 6 Señales QuímicasMar Ruiz Sainz-PardoNo ratings yet
- Sistemas Del CuerpoDocument5 pagesSistemas Del CuerpoPapelería El CentroNo ratings yet
- Presentaciones Sobre GeneticaDocument61 pagesPresentaciones Sobre GeneticaAnpapigua GodiNo ratings yet
- Determinación de hierro sérico por método colorimétricoDocument3 pagesDeterminación de hierro sérico por método colorimétricoAngelesEstelaRuizNo ratings yet
- Guía de Teórico-Coloquios 2016 Biologica GeneticaDocument33 pagesGuía de Teórico-Coloquios 2016 Biologica GeneticaFernando RomeroNo ratings yet
- PediadatosDocument45 pagesPediadatosArturo Melgar Pliego0% (2)
- Valores CriticosDocument35 pagesValores CriticosYohanny Mercedes Rodriguez MoretaNo ratings yet
- La MasturbacionDocument15 pagesLa MasturbacionNena Aguilar100% (1)
- ECCEMAsDocument18 pagesECCEMAsapi-3705495100% (1)
- Fisiología de La FaringeDocument13 pagesFisiología de La FaringeBrooke Wilson0% (1)
- Postura y respiración recursos prácticosDocument42 pagesPostura y respiración recursos prácticosYaneth CabezasNo ratings yet
- DrenajeLinfáticoCuelloDocument9 pagesDrenajeLinfáticoCuelloThaylor Mart Ibañez SandovalNo ratings yet
- Absorción y metabolismo del etanol en el cuerpo humanoDocument20 pagesAbsorción y metabolismo del etanol en el cuerpo humanoConnor CocoNo ratings yet
- Colgajo Cirugia OralDocument10 pagesColgajo Cirugia Oralruben442No ratings yet
- Explicacion TutoriaDocument5 pagesExplicacion TutoriaANANo ratings yet
- Componentes ProteicosDocument3 pagesComponentes ProteicoscitlalyNo ratings yet
- PORCINOSDocument34 pagesPORCINOSHumberto Morales Ramirez100% (2)
- IMVIC: Identificación bacterias mediante pruebas Indol, Rojo de metilo, Voges-Proskauer y CitratoDocument3 pagesIMVIC: Identificación bacterias mediante pruebas Indol, Rojo de metilo, Voges-Proskauer y CitratoOskr RodríguezNo ratings yet
- Protein AsDocument29 pagesProtein AsGabii PachecoNo ratings yet
- CLOREMIADocument37 pagesCLOREMIAMario Perez RumboNo ratings yet
- Evidencia III BIOQUIMICA Equipo 5Document7 pagesEvidencia III BIOQUIMICA Equipo 5Lynn PedrazaNo ratings yet
- Sistema urinario: riñones, nefrona y funciónDocument5 pagesSistema urinario: riñones, nefrona y funciónnancyNo ratings yet
- Tiens AcumagicDocument59 pagesTiens Acumagicelizabeth1199100% (3)
- InsulinoterapiaDocument7 pagesInsulinoterapiaJaviera Andrea Lisperguier CNo ratings yet
- ImagenesDocument7 pagesImagenesMirna VelizNo ratings yet
- Homeostasis y EstresDocument31 pagesHomeostasis y EstresYomar Irazabal DominguezNo ratings yet
- El TalloDocument4 pagesEl TalloReybenguaricapaNo ratings yet
- Los vicios juveniles y sus graves consecuenciasDocument10 pagesLos vicios juveniles y sus graves consecuenciasAnonymous 3UMOUUNo ratings yet
- Exudado NasalDocument3 pagesExudado NasalLaura Rodríguez LópezNo ratings yet