You might also like
- Anatomia Descriptiva Y Funcional Del Aparato RespiratorioDocument37 pagesAnatomia Descriptiva Y Funcional Del Aparato RespiratorioPaola Fabiana del Barrio RuizNo ratings yet
- Vías Aéreas InferioresDocument6 pagesVías Aéreas InferioresJuan AguilaNo ratings yet
- Fisio Transporte de OxigenoDocument18 pagesFisio Transporte de OxigenoAndrés MoraNo ratings yet
- Sistema Respiratorio 2Document9 pagesSistema Respiratorio 2NadiaGavilanCuriNo ratings yet
- Sistema respiratorio: Estructura y función de las vías aéreasDocument5 pagesSistema respiratorio: Estructura y función de las vías aéreasCande Pérez100% (1)
- Circulacion Mayor y Circulacion MenorDocument6 pagesCirculacion Mayor y Circulacion MenorInformes 2022No ratings yet
- PP Respiratorio 08Document41 pagesPP Respiratorio 08Hernán FernándezNo ratings yet
- Cuestionario Aparato RespiratorioDocument7 pagesCuestionario Aparato RespiratorioFederico VeizagaNo ratings yet
- Angiología y EsplacnologíaDocument10 pagesAngiología y EsplacnologíaVicenteFavianCovarrubiasAlcayagaNo ratings yet
- Biofisica de La RespiraciónDocument5 pagesBiofisica de La Respiraciónwandher monteroNo ratings yet
- Introduccion NeumologiaDocument13 pagesIntroduccion NeumologiaDiana PozoNo ratings yet
- Aparato respiratorio: anatomía y fisiologíaDocument32 pagesAparato respiratorio: anatomía y fisiologíaSergio GalvanNo ratings yet
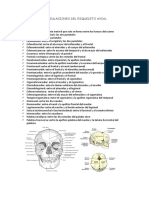
- Articulaciones del esqueleto axialDocument7 pagesArticulaciones del esqueleto axialDaniel LoezaNo ratings yet
- Caja TorácicaDocument10 pagesCaja TorácicaDylan Alexander Lasso YalamaNo ratings yet
- 12 Bronquios y PulmonesDocument45 pages12 Bronquios y PulmonesrubiNo ratings yet
- Sistema RespiratorioDocument3 pagesSistema RespiratorioJose Manuel Mota FigeroaNo ratings yet
- Regiones y planos anatómicosDocument8 pagesRegiones y planos anatómicosBrian HuertaNo ratings yet
- Sangre, Vasos Sanguineos y LinfaticosDocument6 pagesSangre, Vasos Sanguineos y LinfaticosmarleneNo ratings yet
- Arterias CoronariasDocument94 pagesArterias CoronariasROMAINCHONo ratings yet
- Sistema RespiratorioDocument2 pagesSistema RespiratorioMelissa Rivera QuispeNo ratings yet
- Aorta Torácica y AbdomialDocument23 pagesAorta Torácica y AbdomialJonathanNo ratings yet
- Sistema Respiratorio UnapDocument41 pagesSistema Respiratorio UnapruthNo ratings yet
- ResumenesDocument20 pagesResumenesStephano SantistevanNo ratings yet
- Interfase HematogaseosaDocument4 pagesInterfase HematogaseosaLaura Giménez100% (1)
- Vías Aéreas InferioresDocument10 pagesVías Aéreas InferioresPaola NogueraNo ratings yet
- Intercambio GaseosoDocument34 pagesIntercambio GaseosovictorNo ratings yet
- La Arteria AortaDocument3 pagesLa Arteria AortaZin GoneNo ratings yet
- 9 Guía de Estudio 1-9 CorazónDocument12 pages9 Guía de Estudio 1-9 CorazónagustinaNo ratings yet
- Anato Resumen 3Document17 pagesAnato Resumen 3Paola Alexandra Chavez FNo ratings yet
- Resumen T3 - BontragerDocument9 pagesResumen T3 - BontragerLeo MendozaNo ratings yet
- Cuestionario Fisiologia Cap. 1,2,4,5, 25Document5 pagesCuestionario Fisiologia Cap. 1,2,4,5, 25Vinicius SantosNo ratings yet
- Sistema respiratorio: funciones y anatomíaDocument22 pagesSistema respiratorio: funciones y anatomíaBárbara Garcés JarpaNo ratings yet
- PulmónDocument8 pagesPulmónnadallNo ratings yet
- Práctica 8 ExplicaciónDocument9 pagesPráctica 8 ExplicaciónsergifolNo ratings yet
- Vias Respiratorias Superiores e InferioresDocument5 pagesVias Respiratorias Superiores e InferioresDannyel Flores Lott50% (2)
- Control Nervioso de La RespiraciónDocument5 pagesControl Nervioso de La RespiraciónDani Acosta0% (1)
- Práctica 5º ExplicaciónDocument6 pagesPráctica 5º ExplicaciónsergifolNo ratings yet
- El Proceso de La RespiraciónDocument3 pagesEl Proceso de La RespiraciónMario Mauricio Ortega AnayaNo ratings yet
- Exploración RadiológicaDocument7 pagesExploración RadiológicaEder Xavier Cevallos100% (1)
- Neuro Resumen Imprimir PDFDocument52 pagesNeuro Resumen Imprimir PDFAmador IbañezNo ratings yet
- Resumen Del Sistema RespiratorioDocument20 pagesResumen Del Sistema RespiratorioYomike Anthony Murillo LazaroNo ratings yet
- Anatomía y Fisiología Del Aparato RespiratorioDocument29 pagesAnatomía y Fisiología Del Aparato RespiratorioAlexela MendozaNo ratings yet
- Copia de TAREAS SARMIENTODocument75 pagesCopia de TAREAS SARMIENTOValeria Carranza Herrera100% (2)
- El OidoDocument3 pagesEl OidoYudelci Zurita100% (1)
- Construye Un Modelo de PulmonesDocument3 pagesConstruye Un Modelo de PulmonesDaniel RamosNo ratings yet
- Semiología Del Aparato UrinarioDocument26 pagesSemiología Del Aparato UrinarioafdjNo ratings yet
- Apuntes Anatomía COMPLETOSDocument94 pagesApuntes Anatomía COMPLETOSSandra ReinaNo ratings yet
- Anatomía TopográficaDocument9 pagesAnatomía TopográficaMiriam Jimena De Leon PerezNo ratings yet
- Práctica de CorazónDocument18 pagesPráctica de CorazónXimena100% (1)
- Protocolo de Disección de Región Dorsolumbar Anatomia Facultad de MedicinaDocument9 pagesProtocolo de Disección de Región Dorsolumbar Anatomia Facultad de MedicinaCris DevNo ratings yet
- Apuntes Completos AnatomiaDocument104 pagesApuntes Completos Anatomiaeriek.janeiro8017No ratings yet
- Cuestionario de AnatomiaDocument24 pagesCuestionario de Anatomiapatricia100% (1)
- Músculos humanos: tipos, funciones y composiciónDocument5 pagesMúsculos humanos: tipos, funciones y composiciónrafaeltorralbapNo ratings yet
- Control de la respiración por el centro respiratorioDocument4 pagesControl de la respiración por el centro respiratorioJohn Javier GonzalezNo ratings yet
- Músculos Del AntebrazoDocument11 pagesMúsculos Del AntebrazoNathalia SilvaNo ratings yet
- Resumen T9 - BontragerDocument11 pagesResumen T9 - BontragerLeo MendozaNo ratings yet
- Wa0054 1Document18 pagesWa0054 1Francisco BordaNo ratings yet
- 8-Sistema de Nutrición-Sistema RespiratorioDocument10 pages8-Sistema de Nutrición-Sistema RespiratorioLautaro SolisNo ratings yet
- Anatomia y Fisiologia de La RespiracionDocument13 pagesAnatomia y Fisiologia de La RespiracionAXEL YOEL VARGAS MELONo ratings yet
- Aparato Respiratorio y FonadorDocument6 pagesAparato Respiratorio y FonadorMaria Y Sus Manos McvNo ratings yet
- NataciónDocument408 pagesNatacióntorifazio100% (2)
- Neurotape Neurotape: Sports Medicine Center, S.LDocument1 pageNeurotape Neurotape: Sports Medicine Center, S.LFRANCISCO JAVIERNo ratings yet
- TFG Daniel Rivas FloresDocument140 pagesTFG Daniel Rivas FloresFRANCISCO JAVIERNo ratings yet
- 8836 EJERCICIOS-ARTROSIS columnacervical-WEB 2111 PDFDocument2 pages8836 EJERCICIOS-ARTROSIS columnacervical-WEB 2111 PDFLuisa ManzurNo ratings yet
- Terapia acuática: beneficios y centros de tratamientoDocument4 pagesTerapia acuática: beneficios y centros de tratamientoFRANCISCO JAVIERNo ratings yet
- Natació Terapèutica SCMEDocument31 pagesNatació Terapèutica SCMEtaxel0100% (1)
- Tecnica NatacionDocument9 pagesTecnica Natacioncarlosmelissa100% (1)
- Tecnica NatacionDocument9 pagesTecnica Natacioncarlosmelissa100% (1)
- Terapia acuática: beneficios y centros de tratamientoDocument4 pagesTerapia acuática: beneficios y centros de tratamientoFRANCISCO JAVIERNo ratings yet
- Ejercicios de Potenciacion Muscular2tobilloDocument1 pageEjercicios de Potenciacion Muscular2tobilloFRANCISCO JAVIERNo ratings yet
- Informe Final Impacto Cuanti Dic9 2008Document256 pagesInforme Final Impacto Cuanti Dic9 2008FRANCISCO JAVIERNo ratings yet
- Gimnasia en El AguaDocument6 pagesGimnasia en El AguaFRANCISCO JAVIERNo ratings yet
- Boletin 24 - Ejercicios en Agua para OI WEBDocument24 pagesBoletin 24 - Ejercicios en Agua para OI WEBAlicia AltamiranoNo ratings yet
- Revista La HidroterapiaDocument10 pagesRevista La HidroterapiaCarlos Ernesto AriasNo ratings yet
- Columnacervicalmanual1 PDFDocument1 pageColumnacervicalmanual1 PDFFRANCISCO JAVIERNo ratings yet
- Actividades Acuáticas TerapéuticasDocument30 pagesActividades Acuáticas TerapéuticasFabian Andres VeronNo ratings yet
- 03 Ejercicios Columna Lumbar PDFDocument2 pages03 Ejercicios Columna Lumbar PDFFRANCISCO JAVIERNo ratings yet
- Ejercicios CaderaDocument2 pagesEjercicios CaderaTAURUS1313No ratings yet
- Fortalece piernas con 3 ejercicios básicosDocument1 pageFortalece piernas con 3 ejercicios básicosFRANCISCO JAVIERNo ratings yet
- Ejercicios de Williams para abdomen y piernasDocument1 pageEjercicios de Williams para abdomen y piernasFRANCISCO JAVIER0% (1)
- Guia de Ejercicios Lumbar 2Document1 pageGuia de Ejercicios Lumbar 2FRANCISCO JAVIERNo ratings yet
- Ejercicios para HombroDocument2 pagesEjercicios para HombroFRANCISCO JAVIERNo ratings yet
- Preparacion Fisica para Adultos PDFDocument47 pagesPreparacion Fisica para Adultos PDFGabriel Loyo0% (1)
- Preparacion Fisica para Adultos PDFDocument47 pagesPreparacion Fisica para Adultos PDFGabriel Loyo0% (1)
- Ejercicios RodillaDocument2 pagesEjercicios RodillaMayfel ValdezNo ratings yet
- Base Scientific As OsteoporosisDocument19 pagesBase Scientific As OsteoporosisFRANCISCO JAVIERNo ratings yet
- WEB-GUIA-MAYORES Versión Publicación PDFDocument92 pagesWEB-GUIA-MAYORES Versión Publicación PDFRocio Blazquez100% (1)
- Base Scientific As OsteoporosisDocument19 pagesBase Scientific As OsteoporosisFRANCISCO JAVIERNo ratings yet
- Dominiocardiovascular 120719195659 Phpapp02Document6 pagesDominiocardiovascular 120719195659 Phpapp02FRANCISCO JAVIERNo ratings yet
- basesCientificasOsteoporosis PDFDocument21 pagesbasesCientificasOsteoporosis PDFBrighyt Astrid Gonzales GalvezNo ratings yet
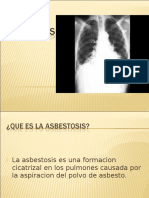
- Asbestosis: causas, síntomas y tratamiento de la enfermedad pulmonar por asbestoDocument24 pagesAsbestosis: causas, síntomas y tratamiento de la enfermedad pulmonar por asbestoHilary MendozaNo ratings yet
- Asfixia Mecanica DocumentoDocument21 pagesAsfixia Mecanica DocumentowendyluzsanchezNo ratings yet
- Semiología y Síndromes CaracterísticosDocument24 pagesSemiología y Síndromes CaracterísticosNelson Rodrigo Quispe VargasNo ratings yet
- RespiraciónDocument3 pagesRespiraciónPam LorasqueNo ratings yet
- Neumonía Connatal o CongénitaDocument17 pagesNeumonía Connatal o CongénitaRodrigo de Sousa100% (1)
- Cuestionario de Sistema CirculatorioDocument5 pagesCuestionario de Sistema CirculatorioSamanta RamirezNo ratings yet
- Aparato respiratorio: estructuras y funcionesDocument18 pagesAparato respiratorio: estructuras y funcionesSebastian Cadavid100% (2)
- Radiologia RespiratoriaDocument96 pagesRadiologia RespiratoriaVictor Raúl Mendoza ConcoNo ratings yet
- Diagnóstico por ImagenDocument155 pagesDiagnóstico por Imagenpetuso2207100% (3)
- Diagnostico de EnfermeriaDocument14 pagesDiagnostico de EnfermeriaMarianna D. Duque CastroNo ratings yet
- Exam - ord.MFH IV - Lun 21.9amDocument6 pagesExam - ord.MFH IV - Lun 21.9amEder Alexander Sánchez PavónNo ratings yet
- Banco de Preguntas Fisiología RespiratoriaDocument11 pagesBanco de Preguntas Fisiología RespiratoriaSergio DextreNo ratings yet
- 5 - Formación de La Energía - Qi, Xue y Jin YeDocument24 pages5 - Formación de La Energía - Qi, Xue y Jin YemonicaNo ratings yet
- Tema 3 PDFDocument13 pagesTema 3 PDFcarmen diazNo ratings yet
- Examen FisiocardioDocument13 pagesExamen FisiocardioMaría Del Barrio EstarliNo ratings yet
- Fisiologia Respiratoria Aplicada A Ventilacion MecanicaDocument47 pagesFisiologia Respiratoria Aplicada A Ventilacion MecanicaGere ChumpitazNo ratings yet
- La Celula de OrganelosDocument4 pagesLa Celula de OrganelosJuan David SuarezNo ratings yet
- Módulo Biología 4 2024Document54 pagesMódulo Biología 4 2024jonathanbernasconi6No ratings yet
- Manejo de SecresionesDocument43 pagesManejo de SecresionesKelly Davila50% (2)
- 2 1 Insuficiencia Respiratoria FarrerasDocument10 pages2 1 Insuficiencia Respiratoria FarrerasDanny Cumpa CapuñayNo ratings yet
- Anexo Fase 3 - LesiónDocument14 pagesAnexo Fase 3 - LesiónPaola MeLoNo ratings yet
- 1 Cuestionario NeumologiaDocument12 pages1 Cuestionario NeumologiaLISBETH ADRIANA HUANCA DURANNo ratings yet
- 10 Preguntas Del ForoDocument2 pages10 Preguntas Del ForoJuanito HernandezNo ratings yet
- Aparatos fonador y respiratorio: órganos y funcionesDocument3 pagesAparatos fonador y respiratorio: órganos y funcionesJulio BarriosNo ratings yet
- Sistema RespiratorioDocument16 pagesSistema RespiratorioJoyas Cosmeticos SolangeNo ratings yet
- Lenguaje 5toDocument4 pagesLenguaje 5toDeNy DíazNo ratings yet
- Manual de Entrenamiento de Salto Libre OperacionalDocument93 pagesManual de Entrenamiento de Salto Libre OperacionalJose50% (2)
- Ppt. 6.-Respuesta Del Sist. Respiratorio Al EjercicioDocument51 pagesPpt. 6.-Respuesta Del Sist. Respiratorio Al EjercicioJulianNo ratings yet
- Respiración en Los Animales para Quinto Grado de SecundariaDocument8 pagesRespiración en Los Animales para Quinto Grado de SecundariaEduardo Hurtado PerezNo ratings yet
- Sistema respiratorio ave corral disecciónDocument3 pagesSistema respiratorio ave corral disecciónJerry VilcaNo ratings yet