You might also like
- Introducción A La InmunologíaDocument1 pageIntroducción A La InmunologíaYami MoralesNo ratings yet
- Teoria Inmunidad Frente A Los MicroorganismosDocument53 pagesTeoria Inmunidad Frente A Los MicroorganismosAlfredo VidalesNo ratings yet
- Evasión de La Respuesta InmuneDocument2 pagesEvasión de La Respuesta InmuneFredy Alexander Pulido QuevedoNo ratings yet
- INMUNOLOGÍA - Seminario 4 PDFDocument13 pagesINMUNOLOGÍA - Seminario 4 PDFDiego Torres AltamiranoNo ratings yet
- Los Bacilos Gran Negativos Que No Muestran Actividad de La Encima Citocromo OxidasaDocument9 pagesLos Bacilos Gran Negativos Que No Muestran Actividad de La Encima Citocromo OxidasaAldair FrancescoliNo ratings yet
- Mecanismos de Algunas Bacterias para Evadir La Respuesta Inmune EXPO INMUNODocument3 pagesMecanismos de Algunas Bacterias para Evadir La Respuesta Inmune EXPO INMUNOJimena ReyesNo ratings yet
- Evasion Inmunitaria Por Parte de Los ParasitosDocument4 pagesEvasion Inmunitaria Por Parte de Los ParasitosSandra VilloNo ratings yet
- Inmunidad Frente A Bacterias Intracelulares - SalmonellaDocument9 pagesInmunidad Frente A Bacterias Intracelulares - SalmonellaBrian Misari AnchiraicoNo ratings yet
- EnterobacteriaceaeDocument91 pagesEnterobacteriaceaes7vdvm28btNo ratings yet
- Resumo SP4Document12 pagesResumo SP4Carol MouraNo ratings yet
- Bacilos Gramnegativos. EnterobacteriasDocument13 pagesBacilos Gramnegativos. EnterobacteriasAndreaFerreiraNo ratings yet
- Evasión InmunitariaDocument26 pagesEvasión Inmunitariajfloresl11No ratings yet
- Taller - Virulencia y Patogenia Viral y Micótica (Resuelto)Document8 pagesTaller - Virulencia y Patogenia Viral y Micótica (Resuelto)Angie MNo ratings yet
- Metabolismo Vírico. Encuentros en La Biología 2006Document4 pagesMetabolismo Vírico. Encuentros en La Biología 2006Guillermo Domínguez HuertaNo ratings yet
- Inmunidad Antiviral - Bacteriana - Antiparasitaria 2022 ViñaDocument48 pagesInmunidad Antiviral - Bacteriana - Antiparasitaria 2022 ViñaEnfermería UNABNo ratings yet
- Inmunidad Frente A Bacterias ExtracelularesDocument10 pagesInmunidad Frente A Bacterias ExtracelularesLaura RojasNo ratings yet
- Mecanismos de evasión inmune en patógenosDocument23 pagesMecanismos de evasión inmune en patógenosJuan José Gómez Toscanelli60% (5)
- Resumen de Respuesta Inmunologica Ante Virus y Bacterias Grupo 3 Equipo 3 InmunologiaDocument4 pagesResumen de Respuesta Inmunologica Ante Virus y Bacterias Grupo 3 Equipo 3 InmunologiaEdher ArturoNo ratings yet
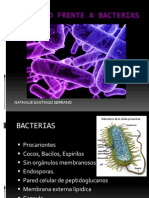
- Inmunidad Frente A BacteriasDocument23 pagesInmunidad Frente A BacteriasNathalie SantiagoNo ratings yet
- Mecanismos de Evasión PDFDocument10 pagesMecanismos de Evasión PDFKatherine Zapata RiosNo ratings yet
- Enterobacteriaceae: familia de bacilos gramnegativosDocument10 pagesEnterobacteriaceae: familia de bacilos gramnegativosAngie Pamela Mirella Colop PuacNo ratings yet
- Mecanismos de Evasion.Document5 pagesMecanismos de Evasion.Liss Tang100% (1)
- Mecanismos de Evasion de La Respuesta InmuneDocument8 pagesMecanismos de Evasion de La Respuesta InmuneNnznznznNo ratings yet
- Respuesta inmune bacterias intracelularesDocument2 pagesRespuesta inmune bacterias intracelulares熊解No ratings yet
- Mecanismos de EvasiónDocument5 pagesMecanismos de EvasiónADRIANA PAOLA GARCIA BARAJASNo ratings yet
- Enfermedades InfecciosasDocument9 pagesEnfermedades InfecciosasJean RojasNo ratings yet
- Gabriel Hernandez Fuentes Respuesta InmunitariaDocument6 pagesGabriel Hernandez Fuentes Respuesta InmunitariaGabriel HernándezNo ratings yet
- Factores de virulencia bacteriana y su regulaciónDocument13 pagesFactores de virulencia bacteriana y su regulaciónKaren SilvaNo ratings yet
- Inmunidad Antibacteriana 2016-1Document39 pagesInmunidad Antibacteriana 2016-1Kiki CarrilloNo ratings yet
- Sesion 9 Sistema InmunologicoDocument94 pagesSesion 9 Sistema InmunologicoJHAMESLEE IVONE JAVIER CUCHILLANo ratings yet
- Características de Los Hogos y BacteriasDocument9 pagesCaracterísticas de Los Hogos y Bacteriaskmai 120No ratings yet
- Inmunidad frente a bacterias: estructura, mecanismos y respuestasDocument8 pagesInmunidad frente a bacterias: estructura, mecanismos y respuestasMilver EscalanteNo ratings yet
- Mecanismos de EvasiónDocument4 pagesMecanismos de EvasiónSairUZárateNo ratings yet
- Enterobacteríaceae. Cap 25, MurrayDocument11 pagesEnterobacteríaceae. Cap 25, Murrayismael AmarilloNo ratings yet
- Grupo F - Cuestionario 04 - Acuña Pérez Juan JesúsDocument7 pagesGrupo F - Cuestionario 04 - Acuña Pérez Juan JesúsJuan Jesús Acuña PėrezNo ratings yet
- Enfermedades infecciosas microbiologíaDocument6 pagesEnfermedades infecciosas microbiologíajose martinezNo ratings yet
- Inmunidad innata y adaptativa frente a parásitosDocument5 pagesInmunidad innata y adaptativa frente a parásitosJAMMILETH LENNY SALVATIERRA GUTIERREZNo ratings yet
- Seminario 5 .Document4 pagesSeminario 5 .Kendra MéndezNo ratings yet
- 8 Inmunidad Frente A Bacterias, Virus, Parasitos, Protozoos y ParazoosDocument29 pages8 Inmunidad Frente A Bacterias, Virus, Parasitos, Protozoos y ParazoosFlora Valencia ManzanaresNo ratings yet
- TEMA Nro 6 Inmunorreaccion Enf InfDocument4 pagesTEMA Nro 6 Inmunorreaccion Enf InfLUIS FERNANDO ORTEGA ESPINOZANo ratings yet
- Inmuno Logia Apuntes Mecanismos de EvasionDocument7 pagesInmuno Logia Apuntes Mecanismos de EvasionEDGAR AXEL GONZALEZ DUENASNo ratings yet
- Inmunidad Contra HongosDocument11 pagesInmunidad Contra HongosMaryori Treyci ILasaca ValenzuelaNo ratings yet
- Mecanismos de Evasión de La Respuesta Inmune en ParásitosDocument7 pagesMecanismos de Evasión de La Respuesta Inmune en ParásitosRina Rubi VillaseñorNo ratings yet
- Ehrlichia CanisDocument5 pagesEhrlichia CanisNaruto ShikamaruNo ratings yet
- 09 - Consolidado Inmunidad Frente A Los MicrobiosDocument46 pages09 - Consolidado Inmunidad Frente A Los MicrobiosJESSELY ROSALÍ MUÑOZ NEYRANo ratings yet
- StreptococcusDocument30 pagesStreptococcusJulio Granado YanceNo ratings yet
- T 10 Respuesta Inmune A Virus y Bacterias UPSJB 2023-IIDocument32 pagesT 10 Respuesta Inmune A Virus y Bacterias UPSJB 2023-IIOswaldo Churi Silva AltamiranoNo ratings yet
- Mecanismos de Evasión de La Respuesta Inmunitaria en Los Parásitos SantyDocument4 pagesMecanismos de Evasión de La Respuesta Inmunitaria en Los Parásitos SantyJORGE SANTIAGO LAZCANO SHERMANNo ratings yet
- Tema 1. Inmunidad Contra BacteriasDocument21 pagesTema 1. Inmunidad Contra BacteriasMarta RelañoNo ratings yet
- 04 Inmunidad y Profilaxis AntiviralDocument30 pages04 Inmunidad y Profilaxis AntiviralRachel Grayson de Todd100% (1)
- Factores Que Facilitan La Invasión MicrobianaDocument3 pagesFactores Que Facilitan La Invasión Microbianaila acostaNo ratings yet
- Inmunidad Frente A Hongos y ParásitosDocument3 pagesInmunidad Frente A Hongos y ParásitosMari CorderoNo ratings yet
- Fisiología y estructura de micobacteriasDocument11 pagesFisiología y estructura de micobacteriasJhonny Neil FloresNo ratings yet
- InmunologíaDocument10 pagesInmunologíaevangelineNo ratings yet
- Mecanismos de Acci N de Los Antimicrobianos PDFDocument9 pagesMecanismos de Acci N de Los Antimicrobianos PDFdatitoxNo ratings yet
- Inmunidad Innata: Douglas R. Mcdonald, Ofer LevyDocument15 pagesInmunidad Innata: Douglas R. Mcdonald, Ofer LevyIsabella GuzmanNo ratings yet
- Respuesta Inmune Contra BacteriasDocument18 pagesRespuesta Inmune Contra BacteriasCrissel Hernández LlacsahuangaNo ratings yet
- Mecanismos de PatogenicidadDocument26 pagesMecanismos de PatogenicidadROBERTO ESAUL GONZALEZ NUNEZNo ratings yet
- 7.clase Inmunidad Virus y Parasitos FMay18 JuevesDocument52 pages7.clase Inmunidad Virus y Parasitos FMay18 JuevesOCTAVIONo ratings yet
- BolPediatr1993 34 289-292Document4 pagesBolPediatr1993 34 289-292Winston Ramos NuñezNo ratings yet
- Webinar 19 MayDocument1 pageWebinar 19 MayWinston Ramos NuñezNo ratings yet
- Preguntas y Respuestas - DermatologíaDocument6 pagesPreguntas y Respuestas - Dermatologíamir-medicina95% (19)
- Fisiología NeonatalDocument1 pageFisiología NeonatalOmar Teran LinaresNo ratings yet
- La Vigilancia Epidemiológica en Salud Pública Es Un Proceso Continuo y Sistemático de ColecciónDocument3 pagesLa Vigilancia Epidemiológica en Salud Pública Es Un Proceso Continuo y Sistemático de ColecciónWinston Ramos NuñezNo ratings yet
- BloqueadoresDocument77 pagesBloqueadoresWinston Ramos NuñezNo ratings yet
- Cuerpos Extraños en ORLDocument2 pagesCuerpos Extraños en ORLAngelica AmesquitaNo ratings yet
- Rol SeptiembreDocument2 pagesRol SeptiembreWinston Ramos NuñezNo ratings yet
- Kit de ClavesDocument18 pagesKit de ClavesWinston Ramos NuñezNo ratings yet
- CLASIFICACIÒNDocument4 pagesCLASIFICACIÒNWinston Ramos NuñezNo ratings yet
- Fiebre ReumáticaDocument2 pagesFiebre ReumáticaJohn Esquivel CisnerosNo ratings yet
- Intoxicaciones y Sus AntídotosDocument2 pagesIntoxicaciones y Sus AntídotosgreyssytorresNo ratings yet
- Indicas PDFDocument1 pageIndicas PDFWinston Ramos NuñezNo ratings yet
- Guía de Práctica Clínica para El Diagnóstico y Tratamiento de La Hipertension ArterialDocument25 pagesGuía de Práctica Clínica para El Diagnóstico y Tratamiento de La Hipertension ArterialWinston Ramos NuñezNo ratings yet
- IndicasDocument1 pageIndicasWinston Ramos NuñezNo ratings yet
- AnemiaDocument13 pagesAnemiaWinston Ramos NuñezNo ratings yet
- Guia Salud RenalDocument25 pagesGuia Salud RenalWinston Ramos NuñezNo ratings yet
- ANEMIADocument13 pagesANEMIAWinston Ramos NuñezNo ratings yet
- Base de DatosDocument4 pagesBase de DatosWinston Ramos NuñezNo ratings yet
- Resumen Guc3adas Ada 2018 2 0Document6 pagesResumen Guc3adas Ada 2018 2 0PUpi GgzNo ratings yet
- Guias Reumatologica de Osteoartritis SAR 2010Document66 pagesGuias Reumatologica de Osteoartritis SAR 2010Haston BillNo ratings yet
- ADA Tratamiento ResuemenDocument9 pagesADA Tratamiento ResuemenCristhian A'DrianzenNo ratings yet
- Factores de Riesgo en Enfermedad Tromboembólica Venosa: Marco Tulio Flores GutiérrezDocument9 pagesFactores de Riesgo en Enfermedad Tromboembólica Venosa: Marco Tulio Flores GutiérrezWinston Ramos NuñezNo ratings yet
- ANTIDIABETICOS ORALES-ampudiablasco2016 PDFDocument13 pagesANTIDIABETICOS ORALES-ampudiablasco2016 PDFaladino abbasNo ratings yet
- Ficha de Recolecion de DatosDocument1 pageFicha de Recolecion de DatosWinston Ramos NuñezNo ratings yet
- Receptores Acoplados A Proteínas GDocument4 pagesReceptores Acoplados A Proteínas GRagurtol AgurtoNo ratings yet
- Guia de Lumbalgia EssaludDocument18 pagesGuia de Lumbalgia EssaludCarlos Emerson Rodriguez MalaverNo ratings yet
- Formulario Quimica Alumnos UdchDocument4 pagesFormulario Quimica Alumnos UdchWinston Ramos NuñezNo ratings yet
- Neumonia Adquirida en La ComunidadDocument32 pagesNeumonia Adquirida en La ComunidadWinston Ramos NuñezNo ratings yet
- Apunte de Transporte A Travéz de La Membrana CelularDocument2 pagesApunte de Transporte A Travéz de La Membrana CelularWinston Ramos NuñezNo ratings yet
- Metabolismo Del Glucógeno y GlucogenolisisDocument4 pagesMetabolismo Del Glucógeno y GlucogenolisisJoshua RichNo ratings yet
- El Adn Como Base de La Vida - Semana 7 de Biologia 2doDocument19 pagesEl Adn Como Base de La Vida - Semana 7 de Biologia 2doAbigailtulcanNo ratings yet
- Unidad 3 Transporte de Membrana GUIA de PREGUNTADocument5 pagesUnidad 3 Transporte de Membrana GUIA de PREGUNTAlauraNo ratings yet
- HKLB AR E1 GyBDocument5 pagesHKLB AR E1 GyBHeaven LozanoNo ratings yet
- Prueba de Ciencias Naturales Grado Sexto Del 2020Document3 pagesPrueba de Ciencias Naturales Grado Sexto Del 2020Eriika Negrete75% (4)
- Biologia Semana 14Document4 pagesBiologia Semana 14carlos MEdinaNo ratings yet
- Hemograma Completo, Perfil Lipídico y TiroidesDocument4 pagesHemograma Completo, Perfil Lipídico y TiroidesNora SmithNo ratings yet
- Cuestionario de Tejido ConectivoDocument4 pagesCuestionario de Tejido ConectivoDiana AgipNo ratings yet
- Inmunidad Adaptativa Frente A Las Bacterias ExtracelularesDocument7 pagesInmunidad Adaptativa Frente A Las Bacterias ExtracelularesJoshue MurrietaNo ratings yet
- Prueba de OctavoDocument4 pagesPrueba de OctavoGeraldine MarambioNo ratings yet
- La SangreDocument13 pagesLa SangrejulietaNo ratings yet
- Manual de La CélulaDocument15 pagesManual de La Célulaomar villaNo ratings yet
- Biologia Celular Y Molecular: Mblga. Dra. Ofelia M. Córdova Paz Soldán 2015Document56 pagesBiologia Celular Y Molecular: Mblga. Dra. Ofelia M. Córdova Paz Soldán 2015MaxNo ratings yet
- Biosíntesis de Nucleótidos de PirimidinaDocument3 pagesBiosíntesis de Nucleótidos de PirimidinaJose Matthew Rivas MoralesNo ratings yet
- RCHJD R3.3 05Document11 pagesRCHJD R3.3 05Jonathan RamirezNo ratings yet
- 4-Citometria de FlujoDocument41 pages4-Citometria de FlujoTatiana MendozaNo ratings yet
- Informe de Laboratorio de B. G #9Document6 pagesInforme de Laboratorio de B. G #9Yureilis Epieyu FajardoNo ratings yet
- Las Células Son Estructuras Increíblemente Complejas y Variadas CAPITULO 2Document34 pagesLas Células Son Estructuras Increíblemente Complejas y Variadas CAPITULO 2acdc29111967No ratings yet
- Curpo Humano y Su Organizacion PDFDocument18 pagesCurpo Humano y Su Organizacion PDFcarol jimena amortegui50% (2)
- 01 GAMETOGÉNESIS IIIIb OOGÉNESISDocument30 pages01 GAMETOGÉNESIS IIIIb OOGÉNESISAdolfo Antonio Ríos AlcortaNo ratings yet
- Ficha 4 CuartoDocument2 pagesFicha 4 CuartoSERGIO OMER REYNA SANTOSNo ratings yet
- Transaminación y Desaminación de ProteínasDocument24 pagesTransaminación y Desaminación de ProteínasCITLALLI MU�OZ ANDRADENo ratings yet
- Biologia, Semana 04Document6 pagesBiologia, Semana 04Angela AbiiNo ratings yet
- Maduración Linfocitos T y BDocument13 pagesMaduración Linfocitos T y BCarlos Shigueta ChavarriaNo ratings yet
- Estructura, función y propiedades de cromosomas, genes, ADN y ARNDocument13 pagesEstructura, función y propiedades de cromosomas, genes, ADN y ARNErick NemhirNo ratings yet
- La CélulaDocument4 pagesLa CélulaKarla Denise Figueroa SanchezNo ratings yet
- Preguntas Sobre La CélulaDocument5 pagesPreguntas Sobre La CélulaJenny Castro JaraNo ratings yet
- Características y Diferencias Entre Células Eucariotas y ProcariotasDocument6 pagesCaracterísticas y Diferencias Entre Células Eucariotas y ProcariotasGavidia JenyferNo ratings yet
- InmunologiaDocument1 pageInmunologiamiraculous ladybugNo ratings yet
- Resumen 1 ParcialDocument17 pagesResumen 1 ParcialLucía Roque RouviereNo ratings yet