You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Pharmacogenomics 121004112431 Phpapp02Document53 pagesPharmacogenomics 121004112431 Phpapp0288AKK100% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- 4.management of Vertical Discrepancies (2) 2Document97 pages4.management of Vertical Discrepancies (2) 2Arun Joy100% (2)
- Kelley Eidem Recipes For CancerDocument4 pagesKelley Eidem Recipes For CancerRohinee SinghNo ratings yet
- Evidence-Based Nursing: The 5 Steps of EBNDocument13 pagesEvidence-Based Nursing: The 5 Steps of EBNRanjith RgtNo ratings yet
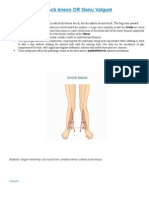
- Genu ValgumDocument2 pagesGenu ValgumPurohit_R0% (1)
- Who Reccomendation MalnutritionDocument123 pagesWho Reccomendation MalnutritionAgaipNo ratings yet
- Moderatemalnutrition Consultation Programmaticaspects MM ReportDocument18 pagesModeratemalnutrition Consultation Programmaticaspects MM ReportCatalina SuciuNo ratings yet
- Giving Children Best Start Intergrating Nutrition 1000daysDocument12 pagesGiving Children Best Start Intergrating Nutrition 1000daysCatalina SuciuNo ratings yet
- Carlo Agostoni Oscar Brunser Issues in Complementary FeedingDocument278 pagesCarlo Agostoni Oscar Brunser Issues in Complementary FeedingCatalina SuciuNo ratings yet
- 2 - Changizi 2006 The Structures of Letters and SymbolsDocument24 pages2 - Changizi 2006 The Structures of Letters and SymbolsCatalina SuciuNo ratings yet
- Giving Children Best Start Intergrating Nutrition 1000daysDocument12 pagesGiving Children Best Start Intergrating Nutrition 1000daysCatalina SuciuNo ratings yet
- Happiness Hypothesis Ch4Document13 pagesHappiness Hypothesis Ch4Catalina SuciuNo ratings yet
- Criminal Justice Policy Review-2006-Lanza-Kaduce-127-43 PDFDocument18 pagesCriminal Justice Policy Review-2006-Lanza-Kaduce-127-43 PDFCatalina SuciuNo ratings yet
- Focus-Group Interview and Data Analysis: Fatemeh RabieeDocument6 pagesFocus-Group Interview and Data Analysis: Fatemeh RabieeCatalina SuciuNo ratings yet
- Introduction of The ResearchDocument3 pagesIntroduction of The Researchcristine baldazoNo ratings yet
- Radiological Findings of 2019-Ncov Pneumonia PMH ExperienceDocument33 pagesRadiological Findings of 2019-Ncov Pneumonia PMH ExperienceHarin NavalanNo ratings yet
- Course List For The Bachelor of Pharmacy (Honours) - My - UQ - The University of Queensland, AustraliaDocument2 pagesCourse List For The Bachelor of Pharmacy (Honours) - My - UQ - The University of Queensland, AustraliaLy Huyen TranNo ratings yet
- 25 Icebreaker Published by Mekar Publishing v2 Part 1Document16 pages25 Icebreaker Published by Mekar Publishing v2 Part 1Fakhrul Anwar AhmadNo ratings yet
- 1) CORRECT - Assess For Patency of The NG Tube Muscle Spasms Associated With VomitingDocument39 pages1) CORRECT - Assess For Patency of The NG Tube Muscle Spasms Associated With VomitingJujuNo ratings yet
- Nursing Care Plan Clubfoot or Talipes EquinovarusDocument6 pagesNursing Care Plan Clubfoot or Talipes EquinovarusMarzie LugtuNo ratings yet
- Paederus Dermatitis: A Case SeriesDocument5 pagesPaederus Dermatitis: A Case SeriesrositaNo ratings yet
- Legalizing MarijuanaDocument3 pagesLegalizing MarijuanaAlyssa MabalotNo ratings yet
- CHECKLIST Trach Care and Suctioning.Document6 pagesCHECKLIST Trach Care and Suctioning.Mickey MoraNo ratings yet
- ICU Liberation ABCDEF Bundle Implementation Both Spontaneous Awakening Breathing Trials SBT SATDocument63 pagesICU Liberation ABCDEF Bundle Implementation Both Spontaneous Awakening Breathing Trials SBT SATbejoNo ratings yet
- Cerebroplacental Ratio in Fetal Well Being Assessment in SGA and AGA FetusDocument11 pagesCerebroplacental Ratio in Fetal Well Being Assessment in SGA and AGA FetuspolygoneNo ratings yet
- Green Back TanjyDocument20 pagesGreen Back TanjymymicroNo ratings yet
- Medsurg (112) Rle: Care of Patients With Problems of The Hematologic SystemDocument5 pagesMedsurg (112) Rle: Care of Patients With Problems of The Hematologic SystemChelsea Faith SarandiNo ratings yet
- Loeffler SyndromeDocument19 pagesLoeffler SyndromeYama Sirly PutriNo ratings yet
- LFD Opd-PediaDocument2 pagesLFD Opd-PediaBaccay GeremieNo ratings yet
- ReseachDocument3 pagesReseachWen TingNo ratings yet
- 2 Marks Q & ADocument11 pages2 Marks Q & AJayaprakash JayaramanNo ratings yet
- Shouldice NotesDocument3 pagesShouldice NotesDisha TutejaNo ratings yet
- Fecal Occult BloodDocument5 pagesFecal Occult BloodTriXie Sorrilla0% (1)
- Encopresis and EnuresisDocument10 pagesEncopresis and EnuresisRoci ArceNo ratings yet
- Guia Tiva 2018Document14 pagesGuia Tiva 2018John Bryan Herrera DelgadoNo ratings yet
- Medical Claim Form - FLEX StaffDocument1 pageMedical Claim Form - FLEX StaffHerawaty Syaiful GusmentaNo ratings yet
- GOLD Global Strategy For Diagnosis, Management, and Prevention of COPD 2014Document102 pagesGOLD Global Strategy For Diagnosis, Management, and Prevention of COPD 2014DanielTanNgNo ratings yet
- AplDocument37 pagesApladitiya tegarNo ratings yet
- Mehmood Ali BukhariDocument1 pageMehmood Ali Bukharinoorulain bukhariNo ratings yet