You might also like
- Resumen - Reacciones Anapleróticas y RegulaciónDocument4 pagesResumen - Reacciones Anapleróticas y Regulaciónanmenc04No ratings yet
- Regulación Del Ciclo Del Ácido CítricoDocument6 pagesRegulación Del Ciclo Del Ácido CítricoJuan Carlos C RxosarxoNo ratings yet
- Ciclo de KlebsDocument9 pagesCiclo de KlebsIram Fernando Oleta CruzNo ratings yet
- Ciclo de KrebsDocument8 pagesCiclo de KrebsMilka Rafaela Acero Vargas50% (2)
- Inhibidores Del Ciclo de KrebsDocument4 pagesInhibidores Del Ciclo de Krebsingjuferdima2240No ratings yet
- Reacciones AnapleroticasDocument8 pagesReacciones AnapleroticasMiriam SalasNo ratings yet
- Regulación Del Ciclo Del Ácido CítricoDocument6 pagesRegulación Del Ciclo Del Ácido CítricoJuan Carlos C RxosarxoNo ratings yet
- Ciclo de Krebs y Vía AnapleróticasDocument7 pagesCiclo de Krebs y Vía AnapleróticasDelgado Mejía BrandonNo ratings yet
- Ciclo glioxilato produce C4Document1 pageCiclo glioxilato produce C4Abel RiveraNo ratings yet
- Inhibidores Del Ciclo de Krebs: FluoracetatoDocument4 pagesInhibidores Del Ciclo de Krebs: FluoracetatoRicardo Miguel Pezet CahuinNo ratings yet
- Síntesis de Ácidos GrasosDocument19 pagesSíntesis de Ácidos GrasosOriana RiveroNo ratings yet
- Acido CitricoDocument25 pagesAcido CitricoLidda Mariam Calixto Heredia0% (1)
- El CICLO de KREBS y Su RegulaciónDocument9 pagesEl CICLO de KREBS y Su Regulaciónluis emilio pachecoNo ratings yet
- 5.5 Metabolismo de CarbohidratosDocument11 pages5.5 Metabolismo de CarbohidratosGiovanni TavarezNo ratings yet
- Síntesis de Ácidos GrasosDocument16 pagesSíntesis de Ácidos GrasosMona Gallegos100% (1)
- Reacciones AnapleroticasDocument3 pagesReacciones AnapleroticasGio PechNo ratings yet
- Ciclo de Krebs y Fosforilacion OxDocument16 pagesCiclo de Krebs y Fosforilacion Oxjoaquin pinuerNo ratings yet
- Ciclo de KrebsDocument9 pagesCiclo de KrebsMariela Barrantes CorderoNo ratings yet
- Ciclo de KerbsDocument6 pagesCiclo de KerbsRosa JiminNo ratings yet
- La Vía de Síntesis de Los Ácidos Grasos Ocurre en El CitoplasmaDocument7 pagesLa Vía de Síntesis de Los Ácidos Grasos Ocurre en El CitoplasmaLauraSeoaneIvanacNo ratings yet
- Taller 2 2015 Bioquimica ResueltoDocument8 pagesTaller 2 2015 Bioquimica ResueltoGisela AcostaNo ratings yet
- Regulación Del Ciclo de KrebsDocument16 pagesRegulación Del Ciclo de Krebspppineda100801No ratings yet
- Glucolisis y Ciclo de Krebs CorregidoDocument28 pagesGlucolisis y Ciclo de Krebs CorregidoMayra Henriquez0% (1)
- Cuestionario AnaplerosisDocument2 pagesCuestionario AnaplerosisJavier GarciaNo ratings yet
- 2ciclo de KrebsDocument6 pages2ciclo de KrebsSanti GarciaNo ratings yet
- Ciclo Del Ácido Cítrico y GlucolisisDocument18 pagesCiclo Del Ácido Cítrico y GlucolisisJerry Carhuatanta SaavedraNo ratings yet
- Glúcidos S3-1Document45 pagesGlúcidos S3-1SarahNo ratings yet
- Ciclo de Krebs MonografiaDocument6 pagesCiclo de Krebs MonografiaRenato Chuquimbalqui Rivas100% (1)
- Cetogenesis y CetolisisDocument4 pagesCetogenesis y CetolisisLeonardo RestrepoNo ratings yet
- Ciclo de KrebsDocument6 pagesCiclo de Krebskatiyerar78% (9)
- Acetil-CoA, Ciclo de KrebsDocument12 pagesAcetil-CoA, Ciclo de KrebsClauMcClauNo ratings yet
- Ciclo de KrebsDocument7 pagesCiclo de KrebsFranciscoSalvadorNo ratings yet
- Apunte 21 - Metabolismo de Lípidos 2Document23 pagesApunte 21 - Metabolismo de Lípidos 2jhonandyramosNo ratings yet
- Regulacion Del Ciclo de KrebsDocument5 pagesRegulacion Del Ciclo de KrebsDrigo S NavarroNo ratings yet
- Ciclo de KrebsDocument9 pagesCiclo de KrebsFootball Club AllianzNo ratings yet
- CetogénesisDocument2 pagesCetogénesisAnonymous I6RzkOVNo ratings yet
- Ciclo de Krebs 22Document11 pagesCiclo de Krebs 22Mari CastedoNo ratings yet
- Ciclo de Krebs: la vía metabólica clave para la oxidación de nutrientesDocument22 pagesCiclo de Krebs: la vía metabólica clave para la oxidación de nutrientesMaria RadcliffeNo ratings yet
- Reacciones AnapleroticasDocument17 pagesReacciones AnapleroticasMaria RivasNo ratings yet
- Ilovepdf MergedDocument108 pagesIlovepdf MergedJuliana MartinezNo ratings yet
- Material de Estudio. Módulo Fuentes y Destino Metabólico Del PiruvatoDocument4 pagesMaterial de Estudio. Módulo Fuentes y Destino Metabólico Del PiruvatoMajo SNo ratings yet
- CicloGlioxilatoConvierteAcetatoGlucosaDocument16 pagesCicloGlioxilatoConvierteAcetatoGlucosaandreaosoriolNo ratings yet
- Relación Con Otras Rutas MetabólicasDocument4 pagesRelación Con Otras Rutas MetabólicasRECENDIZ BENITEZ LUZ YARETHNo ratings yet
- Apuntes Tema 18Document8 pagesApuntes Tema 18gmolermaNo ratings yet
- Ciclo de Krebs: oxidación de Acetil CoA y producción de ATP, NADH y FADH2Document58 pagesCiclo de Krebs: oxidación de Acetil CoA y producción de ATP, NADH y FADH2santos ArchivaldoNo ratings yet
- Ciclo de Krebs BioquimicaDocument8 pagesCiclo de Krebs BioquimicaYeremi MinauroNo ratings yet
- Ciclo Krebs - metabolismo energético célulasDocument2 pagesCiclo Krebs - metabolismo energético célulasAna Victoria Ramírez Iriarte50% (2)
- Ciclo de Krebs: reacciones y producción de ATPDocument11 pagesCiclo de Krebs: reacciones y producción de ATPjuan pabloNo ratings yet
- Regulación y Papel Del Ciclo de Krebs en MedicinaDocument10 pagesRegulación y Papel Del Ciclo de Krebs en MedicinaMontserrat ValenciaNo ratings yet
- U4. Diapositivas Ciclo de KrebsDocument22 pagesU4. Diapositivas Ciclo de KrebsCesar HernandezNo ratings yet
- Preguntas Ciclo de KrebsDocument2 pagesPreguntas Ciclo de KrebsLennys HenriquezNo ratings yet
- Ciclo de KrebsDocument8 pagesCiclo de KrebsGustavo Adolfo MozNo ratings yet
- Ciclo de KrebsDocument9 pagesCiclo de KrebsJohnHarrisonNo ratings yet
- Ciclo de KrebsDocument10 pagesCiclo de KrebsMariana crystel Cabanillas SalcidoNo ratings yet
- Ciclo de KrebsDocument33 pagesCiclo de KrebsAilen GodoyNo ratings yet
- Ciclo de KrebsDocument6 pagesCiclo de KrebsPilar Rojas RuedaNo ratings yet
- Ciclo Del Ácido CítricoDocument5 pagesCiclo Del Ácido CítricoGENESIS HERNANDEZNo ratings yet
- Ciclo de Krebs: reacciones y funcionesDocument7 pagesCiclo de Krebs: reacciones y funcionesDanny Heras GonzalezNo ratings yet
- La Ciencia Del Metabolismo: Metabolismo sin rodeosFrom EverandLa Ciencia Del Metabolismo: Metabolismo sin rodeosRating: 5 out of 5 stars5/5 (1)
- Cine 4Dx MaracaiboDocument4 pagesCine 4Dx MaracaiboChristopher LeonNo ratings yet
- Reguladores de Fosforilacion OxidativaDocument2 pagesReguladores de Fosforilacion OxidativaChristopher LeonNo ratings yet
- TuberculosisDocument23 pagesTuberculosisChristopher LeonNo ratings yet
- Tamaño Optimo de PlantaDocument40 pagesTamaño Optimo de PlantaChristopher LeonNo ratings yet
- BioeticaDocument12 pagesBioeticaChristopher Leon100% (1)
- Bacterias Anaerobias EstrictasDocument173 pagesBacterias Anaerobias EstrictasChristopher LeonNo ratings yet
- Sida en VenezuelaDocument1 pageSida en VenezuelaChristopher LeonNo ratings yet
- InstrumentoDocument1 pageInstrumentoChristopher LeonNo ratings yet
- Glándulas AnexasDocument2 pagesGlándulas AnexasChristopher LeonNo ratings yet
- Tema4 PicornavirusDocument65 pagesTema4 PicornavirusAndres MarquezNo ratings yet
- Estructura de Los CarbohidratosDocument35 pagesEstructura de Los CarbohidratosMariangel VillasmilNo ratings yet
- Aparato DigestivoDocument103 pagesAparato DigestivoNestor Perdomo CNo ratings yet
- Carta Gantt y Diagrama de Bloques1Document36 pagesCarta Gantt y Diagrama de Bloques1Christopher LeonNo ratings yet
- GramPositivosBacilosDocument115 pagesGramPositivosBacilosChristopher LeonNo ratings yet
- Micoplasmas y ChlamydiasDocument98 pagesMicoplasmas y ChlamydiasChristopher LeonNo ratings yet
- Sida en VenezuelaDocument1 pageSida en VenezuelaChristopher LeonNo ratings yet
- Gusto y OlfatoDocument15 pagesGusto y OlfatoChristopher LeonNo ratings yet
- Diabetes, Expo de Pp1Document5 pagesDiabetes, Expo de Pp1Christopher LeonNo ratings yet
- Tema 8 IDocument34 pagesTema 8 IChristopher LeonNo ratings yet
- Para Tiro IdesDocument52 pagesPara Tiro IdesChristopher LeonNo ratings yet
- Unidad IIDocument93 pagesUnidad IIChristopher LeonNo ratings yet
- DengueDocument1 pageDengueChristopher LeonNo ratings yet
- Cine 4Dx MaracaiboDocument4 pagesCine 4Dx MaracaiboChristopher LeonNo ratings yet
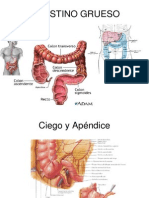
- Intestino GruesoDocument7 pagesIntestino GruesoChristopher LeonNo ratings yet
- Plexo BraquialDocument14 pagesPlexo BraquialChristopher LeonNo ratings yet
- Pliego de CondicionesDocument22 pagesPliego de CondicionesChristopher LeonNo ratings yet
- La Fundicion (Informe)Document31 pagesLa Fundicion (Informe)Christopher LeonNo ratings yet
- Normas COVENIN (Mi Punto)Document10 pagesNormas COVENIN (Mi Punto)Christopher LeonNo ratings yet
- Estirado y TrefiladoDocument11 pagesEstirado y TrefiladoChristopher LeonNo ratings yet