You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Logic of English - Spelling Rules PDFDocument3 pagesLogic of English - Spelling Rules PDFRavinder Kumar80% (15)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Immediate Life Support PDFDocument128 pagesImmediate Life Support PDFShilin-Kamalei Llewelyn100% (2)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Philippine Housing Industry: Regulatory ReviewDocument42 pagesPhilippine Housing Industry: Regulatory ReviewAl MarzolNo ratings yet
- Pathophysiology of Tethered Cord Syndrome and SimilarDocument10 pagesPathophysiology of Tethered Cord Syndrome and SimilarfadhlialbaniNo ratings yet
- Print Jurnal 1Document1 pagePrint Jurnal 1fadhlialbaniNo ratings yet
- Tumor Lysis Syndrome OLDocument37 pagesTumor Lysis Syndrome OLfadhlialbaniNo ratings yet
- MRI CorrelateDocument7 pagesMRI CorrelatefadhlialbaniNo ratings yet
- Altered Iron Metabolism in HIV Infection - Mechanisms, Possible Consequences, and Proposals For Management.Document1 pageAltered Iron Metabolism in HIV Infection - Mechanisms, Possible Consequences, and Proposals For Management.fadhlialbaniNo ratings yet
- 0-DM Problem Chart 2Document1 page0-DM Problem Chart 2fadhlialbaniNo ratings yet
- Transposition of The Great Arteries: What Is It?Document3 pagesTransposition of The Great Arteries: What Is It?fadhlialbaniNo ratings yet
- 7 - Quality of Life, Depressed Mood, and Self-Esteem in Adolescents WithDocument6 pages7 - Quality of Life, Depressed Mood, and Self-Esteem in Adolescents WithfadhlialbaniNo ratings yet
- Mellits and Cheek FormulaDocument5 pagesMellits and Cheek FormulafadhlialbaniNo ratings yet
- Pediatrics-2008-Inattention, Hyperactivity, and School Performance in School Age Children With Complex CHDDocument11 pagesPediatrics-2008-Inattention, Hyperactivity, and School Performance in School Age Children With Complex CHDfadhlialbaniNo ratings yet
- The Impact of Severe Congenital Heart Disease On Physical and Psychosocial Functioning in AdolescentsDocument7 pagesThe Impact of Severe Congenital Heart Disease On Physical and Psychosocial Functioning in AdolescentsfadhlialbaniNo ratings yet
- Etiology Malnutrisi HIVDocument8 pagesEtiology Malnutrisi HIVfadhlialbaniNo ratings yet
- 1 - Psychological Aspects in Children and Adolescent With Congenital Heart Disease and Their ParentsDocument18 pages1 - Psychological Aspects in Children and Adolescent With Congenital Heart Disease and Their ParentsfadhlialbaniNo ratings yet
- Pediatrics-1993 - Psychosocial Risks of Chronic Health Conditions in Childhoentod and AdolescDocument5 pagesPediatrics-1993 - Psychosocial Risks of Chronic Health Conditions in Childhoentod and AdolescfadhlialbaniNo ratings yet
- Evaluasi Anak Dengan Ambigous GenitaliaDocument12 pagesEvaluasi Anak Dengan Ambigous GenitaliafadhlialbaniNo ratings yet
- Infantile Spasms - Pediatric NeurolDocument1 pageInfantile Spasms - Pediatric NeurolfadhlialbaniNo ratings yet
- Pathogenesis of Encephalitis BookDocument19 pagesPathogenesis of Encephalitis BookfadhlialbaniNo ratings yet
- Evaluation of Risk Factors For Severe PneumoniaDocument8 pagesEvaluation of Risk Factors For Severe PneumoniafadhlialbaniNo ratings yet
- Infantile Spasm - EmedicineDocument13 pagesInfantile Spasm - EmedicinefadhlialbaniNo ratings yet
- Infantile Spasms - MenkesDocument4 pagesInfantile Spasms - MenkesfadhlialbaniNo ratings yet
- Guidelines American Academy of Neurology 2011Document4 pagesGuidelines American Academy of Neurology 2011fadhlialbaniNo ratings yet
- Treatment Guidelines For GuillainDocument15 pagesTreatment Guidelines For GuillainfadhlialbaniNo ratings yet
- WHO HIV Clinical Staging GuidelinesDocument49 pagesWHO HIV Clinical Staging Guidelinesdonovandube8235No ratings yet
- Infantile Spasm - EmedicineDocument13 pagesInfantile Spasm - EmedicinefadhlialbaniNo ratings yet
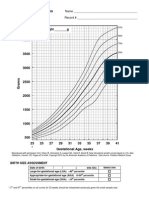
- Growthcurves Gestational AgeDocument4 pagesGrowthcurves Gestational AgefadhlialbaniNo ratings yet
- Altered Iron Metabolism in HIV Infection - Mechanisms, Possible Consequences, and Proposals For Management.Document1 pageAltered Iron Metabolism in HIV Infection - Mechanisms, Possible Consequences, and Proposals For Management.fadhlialbaniNo ratings yet
- Management of Elevated Serum Ferritin LevelsDocument2 pagesManagement of Elevated Serum Ferritin LevelsfadhlialbaniNo ratings yet
- Royals - LordeDocument14 pagesRoyals - LordefadhlialbaniNo ratings yet
- Early Developmental Delays - A Cross Validation StudyDocument5 pagesEarly Developmental Delays - A Cross Validation StudyfadhlialbaniNo ratings yet
- Prevalence of Undetected Developmental Delays in Iranian ChildrenDocument10 pagesPrevalence of Undetected Developmental Delays in Iranian ChildrenfadhlialbaniNo ratings yet
- Engineering Graphics and Desing P1 Memo 2021Document5 pagesEngineering Graphics and Desing P1 Memo 2021dubethemba488No ratings yet
- 9francisco Gutierrez Et Al. v. Juan CarpioDocument4 pages9francisco Gutierrez Et Al. v. Juan Carpiosensya na pogi langNo ratings yet
- Rebecca Young Vs CADocument3 pagesRebecca Young Vs CAJay RibsNo ratings yet
- MCS 033 NotesDocument7 pagesMCS 033 NotesAshikNo ratings yet
- IndianJPsychiatry632179-396519 110051Document5 pagesIndianJPsychiatry632179-396519 110051gion.nandNo ratings yet
- Materials System SpecificationDocument14 pagesMaterials System Specificationnadeem shaikhNo ratings yet
- DHBVNDocument13 pagesDHBVNnitishNo ratings yet
- GE 110HP DC Trolley MotorDocument10 pagesGE 110HP DC Trolley MotorAnthony PetersNo ratings yet
- CA-2 AasthaDocument4 pagesCA-2 AasthaJaswant SinghNo ratings yet
- What Is SCOPIC Clause - A Simple Overview - SailorinsightDocument8 pagesWhat Is SCOPIC Clause - A Simple Overview - SailorinsightJivan Jyoti RoutNo ratings yet
- KBC Autumn Regatta 2023 Saturday Race ScheduleDocument2 pagesKBC Autumn Regatta 2023 Saturday Race SchedulezainNo ratings yet
- Florida v. DunnDocument9 pagesFlorida v. DunnJustice2No ratings yet
- Buddhism & Tantra YogaDocument2 pagesBuddhism & Tantra Yoganelubogatu9364No ratings yet
- Entrepreneurial Skills and Intention of Grade 12 Senior High School Students in Public Schools in Candelaria Quezon A Comprehensive Guide in Entrepreneurial ReadinessDocument140 pagesEntrepreneurial Skills and Intention of Grade 12 Senior High School Students in Public Schools in Candelaria Quezon A Comprehensive Guide in Entrepreneurial ReadinessAngelica Plata100% (1)
- Chapter 3C Problem Solving StrategiesDocument47 pagesChapter 3C Problem Solving StrategiesnhixoleNo ratings yet
- Psychology and Your Life With Power Learning 3Rd Edition Feldman Test Bank Full Chapter PDFDocument56 pagesPsychology and Your Life With Power Learning 3Rd Edition Feldman Test Bank Full Chapter PDFdiemdac39kgkw100% (9)
- MC Donald'sDocument30 pagesMC Donald'sAbdullahWaliNo ratings yet
- Name: Kartikeya Thadani Reg No.: 19bma0029Document4 pagesName: Kartikeya Thadani Reg No.: 19bma0029Kartikeya ThadaniNo ratings yet
- Equivalence ProblemsDocument2 pagesEquivalence ProblemsRomalyn GalinganNo ratings yet
- Field Assignment On Feacal Sludge ManagementDocument10 pagesField Assignment On Feacal Sludge ManagementSarah NamyaloNo ratings yet
- Employer'S Virtual Pag-Ibig Enrollment Form: Address and Contact DetailsDocument2 pagesEmployer'S Virtual Pag-Ibig Enrollment Form: Address and Contact DetailstheffNo ratings yet
- Ignorance Is The Curse of God. Knowledge Is The Wing Wherewith We Fly To Heaven."Document3 pagesIgnorance Is The Curse of God. Knowledge Is The Wing Wherewith We Fly To Heaven."Flori025No ratings yet
- Lista Agentiilor de Turism Licentiate Actualizare 16.09.2022Document498 pagesLista Agentiilor de Turism Licentiate Actualizare 16.09.2022LucianNo ratings yet
- Diane Mediano CareerinfographicDocument1 pageDiane Mediano Careerinfographicapi-344393975No ratings yet
- CGP Module 1 FinalDocument19 pagesCGP Module 1 Finaljohn lexter emberadorNo ratings yet
- Feuerhahn Funeral Bullet 17 March 2015Document12 pagesFeuerhahn Funeral Bullet 17 March 2015brandy99No ratings yet
- Atelierul Digital Pentru Programatori: Java (30 H)Document5 pagesAtelierul Digital Pentru Programatori: Java (30 H)Cristian DiblaruNo ratings yet