You might also like
- BiochimieDocument67 pagesBiochimieana2586No ratings yet
- Biofizica Celulara Cretu CristianDocument3 pagesBiofizica Celulara Cretu CristianlenaileeNo ratings yet
- GlucideleDocument3 pagesGlucideleRodica NancuNo ratings yet
- Ciclul Acizilor TricarboxiliciDocument23 pagesCiclul Acizilor TricarboxiliciandreeaNo ratings yet
- Curs 2 Clasificare. HemoglobinaDocument20 pagesCurs 2 Clasificare. Hemoglobinajacksparrow2No ratings yet
- LP FizioDocument116 pagesLP FizioAndreea FortanNo ratings yet
- Prezentare AnticorpiDocument15 pagesPrezentare AnticorpiOanaDorofteNo ratings yet
- Metabolismul Proteinelor 2Document56 pagesMetabolismul Proteinelor 2Tatiana Manic MunteanuNo ratings yet
- Prezentati Pe Scurt Functiile PolizaharidelorDocument2 pagesPrezentati Pe Scurt Functiile PolizaharidelorDenisa PirvanNo ratings yet
- Totul Despre ProteineDocument9 pagesTotul Despre ProteineCostea MariaNo ratings yet
- UreazaDocument10 pagesUreazaAlexandra BurlacuNo ratings yet
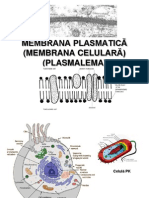
- Membrana PlasmaticăDocument56 pagesMembrana PlasmaticăAndrada Raicu100% (1)
- PinocitozaDocument26 pagesPinocitozaMonicaNo ratings yet
- ATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialDocument8 pagesATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialPascu Alexandru0% (1)
- LP Sange 1 - Fiziologia SangeluiDocument21 pagesLP Sange 1 - Fiziologia SangeluiAnghel Ana-MariaNo ratings yet
- Criterii de Notare Cls A V-ADocument3 pagesCriterii de Notare Cls A V-AAlinaNo ratings yet
- Curs 6 Glicogen, Fru, GalDocument12 pagesCurs 6 Glicogen, Fru, GalMarkus ZahndNo ratings yet
- Tehnici de Colorare in Biologia CelularaDocument11 pagesTehnici de Colorare in Biologia CelularaAlexandra ManuelaNo ratings yet
- 1.5.fiziologia Fibrei MusculareDocument82 pages1.5.fiziologia Fibrei MusculareOana CristinaNo ratings yet
- Ţesuturi Şi Formaţiuni SecretoareDocument1 pageŢesuturi Şi Formaţiuni SecretoareAncuța DanielaNo ratings yet
- BacalaureatDocument35 pagesBacalaureatOprea ViorelNo ratings yet
- Sinteza Si Secretia CelularaDocument6 pagesSinteza Si Secretia Celularaabcfree2006No ratings yet
- Lactat Dehidrogeneza (LDH) - Izoenzime - SynevoDocument4 pagesLactat Dehidrogeneza (LDH) - Izoenzime - SynevoLaurentiu Dinu100% (1)
- Aparitia Celulei EucarioteDocument26 pagesAparitia Celulei EucarioteAlina CatrangiuNo ratings yet
- Referat ConoPidaDocument4 pagesReferat ConoPidalaurentiu112No ratings yet
- Curs 1 - DIGESTIA SI ABSORBTIA ALIMENTELORDocument66 pagesCurs 1 - DIGESTIA SI ABSORBTIA ALIMENTELORmihaelaNo ratings yet
- Celula VegetalaDocument2 pagesCelula VegetalaDorina StamateNo ratings yet
- Microbiologie Bacteriofagul T4Document6 pagesMicrobiologie Bacteriofagul T4Dan Raducu Mhl50% (2)
- Glandele SuprarenaleDocument2 pagesGlandele SuprarenaleAna Maria ToporascuNo ratings yet
- ParatiroideleDocument4 pagesParatiroideleAlexandra DobricăNo ratings yet
- CELULADocument18 pagesCELULASabina PostolacheNo ratings yet
- ParaplastulDocument10 pagesParaplastulLudmila BeregoiNo ratings yet
- Curs 2 Biochimie Clinica PDFDocument9 pagesCurs 2 Biochimie Clinica PDFCristina MurgociNo ratings yet
- Notiuni de Structura A Glucidelor, Lipidelor Si ProteinelorDocument10 pagesNotiuni de Structura A Glucidelor, Lipidelor Si ProteinelorAdrian BăloiNo ratings yet
- DiapedezaDocument10 pagesDiapedezaSoare MihneaNo ratings yet
- Metabolismul LipidelorDocument40 pagesMetabolismul LipidelorEma Virga100% (1)
- LP 1 AdnDocument18 pagesLP 1 AdnGabriela Victoria MartinescuNo ratings yet
- Curs Metabolism PostlicealaDocument19 pagesCurs Metabolism PostlicealaSimona ANo ratings yet
- Branza FetaDocument4 pagesBranza FetaAnaMariaStrNo ratings yet
- Metabolismul Energetic La AnimaleDocument15 pagesMetabolismul Energetic La AnimaleCarmen LucaNo ratings yet
- EnzimologieDocument6 pagesEnzimologieRoxana BoncoiNo ratings yet
- Tesut Conjunctiv Amalinei PDFDocument30 pagesTesut Conjunctiv Amalinei PDFbDashNo ratings yet
- Boli Ale Sistemului Circulator La OmDocument7 pagesBoli Ale Sistemului Circulator La OmTeodoraNo ratings yet
- Digestia Si Absorbtia GlucidelorDocument2 pagesDigestia Si Absorbtia GlucidelorClaudia Grigorii IvanovNo ratings yet
- Catabolismul GlucidelorDocument2 pagesCatabolismul Glucideloririna_gimpireaNo ratings yet
- Absorbţia Apei de Către PlanteDocument2 pagesAbsorbţia Apei de Către PlanteV CNo ratings yet
- DiencefalDocument32 pagesDiencefalBiancaNo ratings yet
- Colesterolul MembranarDocument5 pagesColesterolul MembranarProf. Cristina GheraNo ratings yet
- SingeleDocument47 pagesSingeleOlgutza FloreaNo ratings yet
- FosfolipideleDocument2 pagesFosfolipideleAna RăuNo ratings yet
- Nucleul CelularDocument7 pagesNucleul Celularcandi7No ratings yet
- PoliglucideleDocument11 pagesPoliglucideleDoar Raluca RalucaNo ratings yet
- Curs 6 BiochimieDocument16 pagesCurs 6 BiochimieAlexandra IoanaNo ratings yet
- 1 BioelementeDocument59 pages1 BioelementeMihaela PuscasNo ratings yet
- Structura ProteinelorDocument13 pagesStructura ProteinelorBigangelNo ratings yet
- Structura Primara Si Secundara A ProteinelorDocument9 pagesStructura Primara Si Secundara A ProteinelornosgothsoulNo ratings yet
- Structura ProteinelorDocument7 pagesStructura Proteinelorsilvia constantinNo ratings yet
- Structura ProteinelorDocument8 pagesStructura Proteinelorsilvia constantinNo ratings yet
- Structura ProteinelorDocument7 pagesStructura ProteinelorIoana VoinescuNo ratings yet
- Structura ProteinelorDocument3 pagesStructura ProteinelorElena Cristina U100% (1)
- Olimpiada cl.9Document2 pagesOlimpiada cl.9A BNo ratings yet
- Absolut Superb !Document47 pagesAbsolut Superb !mascaf@yahoo.comNo ratings yet
- Evaluarea Sumativă Vertebrate Cl-7docxDocument8 pagesEvaluarea Sumativă Vertebrate Cl-7docxMarina GuzunNo ratings yet
- ES CL 7 PlanteDocument4 pagesES CL 7 PlanteElena Catana100% (3)
- Riscurile Sarcinii in Adolescenta Pentru Mama Si CopilDocument2 pagesRiscurile Sarcinii in Adolescenta Pentru Mama Si CopilA BNo ratings yet
- Proiectare Biologie 6Document12 pagesProiectare Biologie 6A BNo ratings yet
- Apa Pentru ViataDocument6 pagesApa Pentru ViataA B100% (1)
- Evaluare Sumativa Vertebrate CL.7 2020Document2 pagesEvaluare Sumativa Vertebrate CL.7 2020A BNo ratings yet
- Evaluare PlantedocxDocument3 pagesEvaluare PlantedocxOlga MihailiucNo ratings yet
- Proiectare Biologie 6 2021Document13 pagesProiectare Biologie 6 2021A BNo ratings yet
- Proiect de Lunga Durata Chimie Clasa VIIDocument17 pagesProiect de Lunga Durata Chimie Clasa VIIA BNo ratings yet
- Olimpiada Locala BiologieDocument4 pagesOlimpiada Locala BiologieA BNo ratings yet
- Chimie Clasa VIIDocument18 pagesChimie Clasa VIIA BNo ratings yet
- Proiect de Lunga Durata Biologie Clasa 8Document10 pagesProiect de Lunga Durata Biologie Clasa 8A BNo ratings yet
- Riscurile Sarcinii in Adolescenta Pentru Mama Si CopilDocument2 pagesRiscurile Sarcinii in Adolescenta Pentru Mama Si CopilA BNo ratings yet
- Nerespectarea Normelor Elementare de IgienaDocument1 pageNerespectarea Normelor Elementare de IgienaA BNo ratings yet
- PovesteaapeiDocument13 pagesPovesteaapeiA BNo ratings yet
- Nerespectarea Normelor Elementare de IgienaDocument1 pageNerespectarea Normelor Elementare de IgienaA BNo ratings yet
- Biologie CurriculumDocument28 pagesBiologie CurriculumA BNo ratings yet
- Educatie Pentru Sanatate, Clasa A VI-aDocument7 pagesEducatie Pentru Sanatate, Clasa A VI-aA BNo ratings yet
- GimnospermeDocument13 pagesGimnospermeA B100% (1)
- NU Se Aadmite!!Document1 pageNU Se Aadmite!!A BNo ratings yet
- FluturiDocument1 pageFluturiMihaela MihalacheNo ratings yet