You might also like
- Aula 7 SeleçãoDocument43 pagesAula 7 SeleçãoAndressa Nunes0% (1)
- Seleção zootécnica e funcional em gado de corte: o olho e a máquinaDocument101 pagesSeleção zootécnica e funcional em gado de corte: o olho e a máquinaTainan HalberstadtNo ratings yet
- Glossário de ZootecniaDocument36 pagesGlossário de ZootecniaTrícya CarolinnyNo ratings yet
- Métodos de SeleçãoDocument44 pagesMétodos de Seleçãolucildalima100% (14)
- Gestão de pequenas propriedades leiteiras no sudoeste do ParanáDocument14 pagesGestão de pequenas propriedades leiteiras no sudoeste do ParanáPaulo GodoiNo ratings yet
- 8-Exame Andrológico Do TouroDocument52 pages8-Exame Andrológico Do Tourolucaskkk123No ratings yet
- 4-MANEJO SANITÁRIO VACAS LEITEIRAS DefDocument12 pages4-MANEJO SANITÁRIO VACAS LEITEIRAS DefMarciliana Davantel100% (1)
- SuinoculturaDocument27 pagesSuinoculturaVet_arquivos100% (3)
- A Importância Do Médico Veterinário Na Saúde PúblicaDocument47 pagesA Importância Do Médico Veterinário Na Saúde PúblicaVet_arquivos100% (3)
- Alguns Pontos Importantes em Exames Clínicos de AnimaisDocument4 pagesAlguns Pontos Importantes em Exames Clínicos de AnimaisVet_arquivosNo ratings yet
- Primeiros socorros para pilotos de parapenteDocument4 pagesPrimeiros socorros para pilotos de parapentePaula MeloNo ratings yet
- Diferencial de Seleção e Ganho GenéticoDocument14 pagesDiferencial de Seleção e Ganho Genéticolucildalima100% (5)
- DEP: Como Calcular e Como UtilizarDocument5 pagesDEP: Como Calcular e Como Utilizaravantidigital100% (2)
- Gestão Do Melhoramento Genético Do RebanhoDocument39 pagesGestão Do Melhoramento Genético Do RebanhoALICE CAROLINANo ratings yet
- Melhoramento Genetico em Gado LeiteiroDocument41 pagesMelhoramento Genetico em Gado LeiteiroDelano AlencarNo ratings yet
- Aula Parametros Genéticos PDFDocument20 pagesAula Parametros Genéticos PDFKaio FernandesNo ratings yet
- Aula 1.1 Nocoes de Melhoramento AnimalDocument46 pagesAula 1.1 Nocoes de Melhoramento AnimalDaniel ArrudaNo ratings yet
- EPMURAS na Anhembi MorumbiDocument9 pagesEPMURAS na Anhembi MorumbiLaura ParrasNo ratings yet
- Utilizacao de Escores Visuais de Caracteristicas Morfologicas de Bovinos Nelore Como Ferramenta para o Melhoramento Genetico Animal PDFDocument39 pagesUtilizacao de Escores Visuais de Caracteristicas Morfologicas de Bovinos Nelore Como Ferramenta para o Melhoramento Genetico Animal PDFOdaizi TelesNo ratings yet
- Fichasparacontrolezootecnico PDFDocument35 pagesFichasparacontrolezootecnico PDFElder BiancoNo ratings yet
- Controle Gado CorteDocument30 pagesControle Gado Corteadailson mirandaNo ratings yet
- Melhoramento Genético Animal No Brasil: Fundamentos, História e ImportânciaDocument63 pagesMelhoramento Genético Animal No Brasil: Fundamentos, História e ImportânciaFranciscaGorettiRabelloBrasilNo ratings yet
- Cruzamentos em gado de corte: sistemas, vantagens e resultados de pesquisaDocument68 pagesCruzamentos em gado de corte: sistemas, vantagens e resultados de pesquisaAluisioNeves100% (1)
- Melhoramento genético animalDocument16 pagesMelhoramento genético animalChrisvet100% (2)
- Ciclo Estral BovinosDocument33 pagesCiclo Estral BovinosswaggerleeNo ratings yet
- Manejo Nutricional de Vacas em LactaçãoDocument22 pagesManejo Nutricional de Vacas em Lactaçãomarllinh0No ratings yet
- Diagnóstico de Gestação e de Patologias Uterinas Por Ecografia em Ruminantes, Equinos e SuínosDocument72 pagesDiagnóstico de Gestação e de Patologias Uterinas Por Ecografia em Ruminantes, Equinos e SuínosEduardo De Oliveira CostaNo ratings yet
- Pastoreio Racional VoisinDocument35 pagesPastoreio Racional VoisinEdno Negrini JrNo ratings yet
- Raças e Cruzamentos de Bovinos de LeiteDocument51 pagesRaças e Cruzamentos de Bovinos de LeiteCaroline ItnerNo ratings yet
- Manual Sisgado EsDocument63 pagesManual Sisgado EsCésar Bonifaz100% (1)
- Manual de Exame Ginecológico Da VacaDocument5 pagesManual de Exame Ginecológico Da VacamarciogomesgvNo ratings yet
- Anestro Pós-Parto em Bovinos Mecanismos de Redução Do PeríodoDocument23 pagesAnestro Pós-Parto em Bovinos Mecanismos de Redução Do PeríodoVet_arquivosNo ratings yet
- Clínica Médica de Ruminantes 01Document36 pagesClínica Médica de Ruminantes 01Ana Paula CamposNo ratings yet
- Recomendacoes Tecnicas para Alimentacao de Pequenos RuminantDocument281 pagesRecomendacoes Tecnicas para Alimentacao de Pequenos RuminantRafael LopesNo ratings yet
- Manejo reprodutivo de gado de corteDocument71 pagesManejo reprodutivo de gado de corteMirian FernandesNo ratings yet
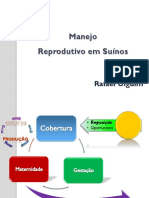
- Manejo Reprodutivo em SuínosDocument55 pagesManejo Reprodutivo em SuínosPedro SantosNo ratings yet
- Periodo Seco Vacas LeiteirasDocument4 pagesPeriodo Seco Vacas LeiteirasCybervetNo ratings yet
- Aspectos Da Reprodução Equina: Inseminação Artificial E Tranferência de Embrião: Revisão de LiteraturaDocument23 pagesAspectos Da Reprodução Equina: Inseminação Artificial E Tranferência de Embrião: Revisão de LiteraturaAdryano CarvalhoNo ratings yet
- Controle zootécnico em bovinosDocument3 pagesControle zootécnico em bovinosGelson LodiNo ratings yet
- Sistema reprodutor eqüinoDocument33 pagesSistema reprodutor eqüinoNeide100% (3)
- Protocolo de Iatf Com Benzoato de Estradiol NoDocument22 pagesProtocolo de Iatf Com Benzoato de Estradiol NoAlana BertoliNo ratings yet
- Revista CRMV - Produção de Embriões Bovinos in Vitro Com Semen SexadoDocument100 pagesRevista CRMV - Produção de Embriões Bovinos in Vitro Com Semen Sexadorodrigoferrazza1662No ratings yet
- 9 Índices Zootécnicos Recomendados de Acordo Com Cada EspécieDocument10 pages9 Índices Zootécnicos Recomendados de Acordo Com Cada EspécieMaria Ercilia FernandesNo ratings yet
- Controle Zootecnico Bovinos de CorteDocument7 pagesControle Zootecnico Bovinos de CorteIdyla AvelarNo ratings yet
- Passo A Passo IATF - CompactadaDocument37 pagesPasso A Passo IATF - CompactadaCabanha Labor Charolês100% (2)
- Ciclos Reprodutivos em Animais DomésticosDocument25 pagesCiclos Reprodutivos em Animais DomésticosAlbertoGonçalvesEvangelistaNo ratings yet
- Reprodução em Equinos ManejoDocument97 pagesReprodução em Equinos Manejoleticia2001delgadoNo ratings yet
- Aula - Formulação de Rações - CompletaDocument21 pagesAula - Formulação de Rações - CompletaJosimar da Silva Chaves100% (1)
- Heterose e HeterozigoseDocument25 pagesHeterose e Heterozigoselucildalima100% (1)
- IA em Suínos: RevisãoDocument90 pagesIA em Suínos: RevisãoJúlio Cesar PereiraNo ratings yet
- Sincronização de Cio Ovinos e CaprinosDocument58 pagesSincronização de Cio Ovinos e CaprinosRodrigo RodriguesNo ratings yet
- Genoma BovinoDocument18 pagesGenoma BovinoFlávio LimaNo ratings yet
- Exterior BovinosDocument135 pagesExterior Bovinosmjm5885No ratings yet
- Maneio Geral de Gado de Leite 22Document28 pagesManeio Geral de Gado de Leite 22Carimo gorges ArmandoNo ratings yet
- Nutrição e manejo de bovinos de corte na fase de criaDocument30 pagesNutrição e manejo de bovinos de corte na fase de criaMilena AlvesNo ratings yet
- Nutrição de búfalas leiteiras: composição, requerimentos e formulação de raçõesDocument16 pagesNutrição de búfalas leiteiras: composição, requerimentos e formulação de raçõesJadir FélixNo ratings yet
- Fibra Detergente Neutro PDFDocument2 pagesFibra Detergente Neutro PDFMigdaliaNo ratings yet
- Anatomia das aves resume ossos, músculos e sistemasDocument32 pagesAnatomia das aves resume ossos, músculos e sistemasAmandaCharaloNo ratings yet
- Suínos - 500 PerguntasDocument245 pagesSuínos - 500 PerguntasMV Meybe ReginaNo ratings yet
- Nanotecnologia aplicada à produção agrícola: uma análise acerca dos aspectos sociais e sustentáveis no âmbito brasileiroFrom EverandNanotecnologia aplicada à produção agrícola: uma análise acerca dos aspectos sociais e sustentáveis no âmbito brasileiroNo ratings yet
- Herdabilidade ResumoDocument7 pagesHerdabilidade ResumoAdemilson JuniorNo ratings yet
- Valor Genético e DEP. Objetivos Com o Texto - by Natália Albieri Koritiaki - MediumDocument18 pagesValor Genético e DEP. Objetivos Com o Texto - by Natália Albieri Koritiaki - MediumSoraya SolNo ratings yet
- A importância da disseminação do conhecimento sobre animais peçonhentosDocument24 pagesA importância da disseminação do conhecimento sobre animais peçonhentosAlan DemarcosNo ratings yet
- (PDF) Demodicose Canina PDFDocument15 pages(PDF) Demodicose Canina PDFVet_arquivosNo ratings yet
- Gênero Leptospira 2015Document52 pagesGênero Leptospira 2015Vet_arquivosNo ratings yet
- Bravecto DemodecicaDocument7 pagesBravecto DemodecicaVet_arquivosNo ratings yet
- Wasava 2015 - Diretrizes para A Vacinação de Cães e Gatos PDFDocument50 pagesWasava 2015 - Diretrizes para A Vacinação de Cães e Gatos PDFmarcelo100% (1)
- Controle Endócrino Do Equilíbrio Hidro-EletrolíticoDocument18 pagesControle Endócrino Do Equilíbrio Hidro-EletrolíticoVet_arquivosNo ratings yet
- Dicionário Inglês-Português de SuinoculturaDocument30 pagesDicionário Inglês-Português de SuinoculturaLeandro CastilhaNo ratings yet
- Curso VendedorDocument138 pagesCurso VendedorVet_arquivosNo ratings yet
- Manual Pos VacinaçãoDocument188 pagesManual Pos VacinaçãoTamara Coutinho Galvão SilvaNo ratings yet
- Carbúnculo Sintomático em BovinoDocument5 pagesCarbúnculo Sintomático em BovinoVet_arquivosNo ratings yet
- Choque Circulatorio-Aspectos Básicos de Fisiopatologia e TerapeuticaDocument22 pagesChoque Circulatorio-Aspectos Básicos de Fisiopatologia e TerapeuticaVet_arquivosNo ratings yet
- As Prostaglandinas Na ReproduçãoDocument26 pagesAs Prostaglandinas Na ReproduçãoVet_arquivos100% (2)
- Timpanismo em RuminAntesDocument6 pagesTimpanismo em RuminAntesVet_arquivos100% (1)
- Sarna Sarcóptica em CãesDocument5 pagesSarna Sarcóptica em CãesMJoanaDuarteNo ratings yet
- Qualidade Do Colostro BovinoDocument5 pagesQualidade Do Colostro BovinoVet_arquivosNo ratings yet
- Retenção de Placenta em VacasDocument7 pagesRetenção de Placenta em VacasVet_arquivos100% (1)
- Causa e Tratamento Do Garrotilho em EquinosDocument5 pagesCausa e Tratamento Do Garrotilho em EquinosVet_arquivos100% (1)
- Sistema Digestivo Dos Rum in AntesDocument6 pagesSistema Digestivo Dos Rum in AntesVet_arquivosNo ratings yet
- Carbúnculo HemáticoDocument5 pagesCarbúnculo HemáticoVet_arquivos100% (1)
- Noções Básicas de FarmacologiaDocument16 pagesNoções Básicas de FarmacologiaVet_arquivos100% (5)
- Doenças Respiratórias Dos BovinosDocument6 pagesDoenças Respiratórias Dos BovinosVet_arquivosNo ratings yet
- Terapêutica de Aves SilvestresDocument6 pagesTerapêutica de Aves SilvestresVet_arquivosNo ratings yet
- Raças de CãesDocument98 pagesRaças de CãesVet_arquivos100% (4)
- Exame Clínico de Enfermidades Do Sist Nervoso em Equinos e RuminantesDocument17 pagesExame Clínico de Enfermidades Do Sist Nervoso em Equinos e RuminantesVet_arquivosNo ratings yet
- Digestão de Celulose Nos VertebradosDocument2 pagesDigestão de Celulose Nos VertebradosVet_arquivosNo ratings yet
- Inovações Técnológicas Na BioclimatologiaDocument24 pagesInovações Técnológicas Na BioclimatologiaVet_arquivos100% (1)
- Métodos Semiológicos de Exame Físico em RuminantesDocument5 pagesMétodos Semiológicos de Exame Físico em RuminantesVet_arquivos100% (3)
- PeelingsDocument12 pagesPeelingsjumaracruzNo ratings yet
- InformaçõesDocument2 pagesInformaçõesManuela ManuNo ratings yet
- Chow ChowDocument9 pagesChow ChowAdestrador Jefferson SantosNo ratings yet
- Doença Da Mancha Branca Na Criação de Camarões UsjtDocument1 pageDoença Da Mancha Branca Na Criação de Camarões UsjtFernanda Ramos SturalliNo ratings yet
- Inspeção de Suínos - RevisãoDocument222 pagesInspeção de Suínos - RevisãoJosé Luís Franco71% (7)
- Trabalho de MorcegosDocument6 pagesTrabalho de MorcegosNayara MeucciNo ratings yet
- Cap. 5 Parte E Filo Dos AnelideosDocument15 pagesCap. 5 Parte E Filo Dos AnelideosJoão L. CasemiroNo ratings yet
- Saúde Da Mulher - Resumo Parcial 1a Prova - Obstetrícia PDFDocument51 pagesSaúde Da Mulher - Resumo Parcial 1a Prova - Obstetrícia PDFEdson Vicente Dias Corrêa100% (1)
- REVISÃO SOBRE A RAIVADocument19 pagesREVISÃO SOBRE A RAIVADaniel StarkNo ratings yet
- 51 Exerc - Cios Sistema Digest - RioDocument9 pages51 Exerc - Cios Sistema Digest - RioAndryo CardosoNo ratings yet
- Fisiologia AnestesicosDocument8 pagesFisiologia AnestesicosJ VichhyNo ratings yet
- Lista de Exercícios de Biologia sobre Vermes e suas DoençasDocument12 pagesLista de Exercícios de Biologia sobre Vermes e suas Doençasricardo1alvesNo ratings yet
- Manual Do CorredorDocument14 pagesManual Do CorredorAnderson FerroBemNo ratings yet
- Determinação dos grupos sanguíneos de alunos e professoresDocument16 pagesDeterminação dos grupos sanguíneos de alunos e professoresMaggy AraújoNo ratings yet
- Desenvolvimento embrionário de anfioxoDocument7 pagesDesenvolvimento embrionário de anfioxoAndressa OliveiraNo ratings yet
- Desenvolvimento Da Dentição MistaDocument25 pagesDesenvolvimento Da Dentição Mistaadrianemarcos50% (2)
- Neuropsicologia Da LinguagemDocument3 pagesNeuropsicologia Da LinguagemPaola Aparecida0% (1)
- Aula Edema Agudo de PulmaoDocument28 pagesAula Edema Agudo de PulmaoMarcos Cunha0% (1)
- Manual Terapias AlternativasDocument29 pagesManual Terapias AlternativasAnalice Carvalho100% (1)
- Alterações Imunológicas - Hipersensibilidade Mediada Por Anticorpos eDocument26 pagesAlterações Imunológicas - Hipersensibilidade Mediada Por Anticorpos eAriel CorciniNo ratings yet
- Sistema Circulatório e RespiratórioDocument5 pagesSistema Circulatório e RespiratórioMichelly Marthinely Tavares50% (2)
- Exame físico cabeça e pescoço: inspeção, palpação e auscultaDocument6 pagesExame físico cabeça e pescoço: inspeção, palpação e auscultaJulhia ManuelaNo ratings yet
- Injetáveis mensais: pontos básicos para profissionais e clientesDocument20 pagesInjetáveis mensais: pontos básicos para profissionais e clientesRohseney SilvaNo ratings yet
- Identificação de fungos causadores de micosesDocument23 pagesIdentificação de fungos causadores de micosesPaula StangherlinNo ratings yet
- Capitulo23 PDFDocument30 pagesCapitulo23 PDFAndersonNo ratings yet
- Caso Clínico de Fratura de Rádio DistalDocument33 pagesCaso Clínico de Fratura de Rádio Distaljglfm6296No ratings yet
- Higiene oral em UCI: avaliação da equipe de enfermagemDocument8 pagesHigiene oral em UCI: avaliação da equipe de enfermagemsamuelNo ratings yet
- Estudos de caso sobre morte materna na Rede Mãe ParanaenseDocument32 pagesEstudos de caso sobre morte materna na Rede Mãe ParanaenseDiogenes Borowsky100% (1)
- Reab Pulm Melhorar VidaDocument10 pagesReab Pulm Melhorar VidaLuciani MoreiraNo ratings yet