Msculos e receptores musculares . Por Karine Kavalco, 1998. .
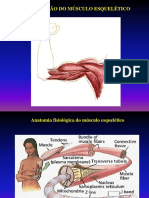
Msculo esqueltico O msculo de vertebrados est sob controle voluntrio e tem um aspecto de estriado. A base estrutural da contrao em msculo estriado de vertebrados - o processo gerador de fora mais bem conhecido - constitudo de clulas multinucleadas que so delimitadas por uma membrana celular eletricamente excitvel (Stryer, 1996). O msculo estriado esqueltico formado por feixes de clulas cilndricas muito longas e multinucleares, que apresentam estriaes transversais. Tm contrao rpida, vigorosa e sujeita ao controle voluntrio (Junqueira & Carneiro, 1996). Uma clula muscular contm muitas miofibrilas paralelas, cada uma com cerca de 1 mm de dimetro, que esto imersas no citossol (Stryer, 1996). Segundo Junqueira & Carneiro (1990), os feixes de fibras esto organizados e envolvidos por uma membrana, o epimsio. O epimsio composto por tecido conjuntivo que adentra o limite da fibra envolvendo feixes menores. Neste ponto o chamamos perimsio. Por sua vez, cada fibra muscular envolvida por uma camada muito fina constituda pela lmina basal da fibra muscular e por fibras reticulares do endomsio. Esses envoltrios fazem com que as fibras permaneam unidas e que no haja dissipao de foras durante a contrao muscular. Essa funo importante porque a maioria das fibras no se estende de uma extremidade a outra da fibra. ainda por este tecido conjuntivo que acontece a transmisso da fora de contrao do msculo a tendes, ossos, ligamentos, etc. Cada fibra muscular apresenta perto de seu centro uma terminao nervosa motora, a chamada placa motora.
Miofibrilas O citoplasma da fibra muscular apresenta-se preenchido principalmente por fibrilas paralelas - as miofibrilas. As miofibrilas so cilndricas e preenchem completamente o interior da fibra muscular. Ao microscpio tico aparecem com estriaes transversais, pelas alternncias de faixas claras e escuras. A faixa escura anisotrpica ( banda A), enquanto a faixa clara (banda I) isotrpica. No centro de cada banda I aparece uma linha transversal escura - a linha Z. A estriao da miofibrila devida repetio de unidades iguais, chamadas sarcmeros. Cada sarcmero formado pela parte da miofibrila que fica entre duas linhas Z sucessivas e contm uma banda A separando duas semibandas I. A banda A apresenta uma zona mais clara no seu centro, a banda H. A disposio dos sarcmeros coincide nas vrias miofibrilas da fibra muscular. Conseqentemente, as bandas formas um sistema de estriaes transversais, paralelas, que caracterstico das fibras musculares estriadas. Da linha Z partem os filamentos finos (actina) que correm at o bordo externo da banda H. Os filamentos grossos (miosina) ocupam a regio central do sarcmero. Como resultado dessa disposio, a banda I formada somente pela parte dos filamentos finos que no so invadidos pelos filamentos grossos. A banda A formada principalmente por filamentos grossos e a banda H somente pelos filamentos grossos. Na regio lateral da banda A, os filamentos finos e grossos se interdigitam. Um corte transversal nessa regio mostra uma disposio simtrica tal que cada filamento grosso fica rodeado por seis filamentos finos, num arranjo hexagonal (Junqueira & Carneiro, 1990). As miofibrilas do msculo estriado contm quatro protenas principais: miosina, actina, tropomiosina e troponina. Os filamentos grossos so formados de miosina e as outras trs protenas
so encontradas nos filamentos finos. A miosina e a actina, juntas, representam 55% do total de protenas do msculo estriado (Junqueira & Carneiro, 1990). Miosina: a miosina tem trs atividades biolgicas importantes: so o principal constituinte dos filamentos grossos; uma enzima e uma ATPase; e liga-se actina polimerizada. ATP + H 2O ADP + Pi + H+. Essa equao global, que ocorre em uma srie de etapas independentes, fornece a energia livre para contrao muscular. A miosina pode ser vista como uma mecanoenzima porque ela catalisa a converso de energia qumica a mecnica. uma protena grande, constitudas por seis cadeias polipeptdicas: duas cadeias pesadas idnticas e dois pares de cadeias leves. A molcula consiste de duas cabeas unidas a um basto muito longo. O basto so duas a-hlices formado pelas cadeias pesadas. A cadeia pesada de cada cabea liga-se a duas cadeias leves diferentes. Actina: a principal constituinte dos filamentos finos. A actina um monmero chamada actinaG, pela sua forma globular; quando a fora inica aumenta, esta polimeriza-se numa forma fibrosa , a actinaF. Como a miosina, esta protena tambm uma ATPase; o ciclo ATP-ADP da actina participa na montagem e desmonte do filamento. Todas as unidades de actina de uma filamento tm o mesmo sentido. Triponina e tropomiosina: a actina e a miosina so mantidas sob controle pela tropomiosina e pelo complexo de troponina. Estas esto localizadas no filamento fino. A tropomiosina um basto filamentoso que alinha-se quase paralelamente ao eixo do filamento fino. A troponina um complexo de trs cadeias polipeptdicas: TnC, TnI e TnT. Cada complexo de troponina regula, atravs da tropomiosina, as interaes com actina (Stryer, 1996).
Sarcolema e Sarcoplasma O sarcolema a membrana celular da fibra muscular. No entanto, consiste de uma membrana celular verdadeira, a membrana plasmtica, e de uma fina camada de material polissacardico similar ao da membrana basal que circunda os capilares sangneos; finas fibrilas colgenas tambm esto presentes na camada mais externa do sarcolema. Nas extremidades das fibras musculares, essas camadas superficiais do sarcolema se fundem com as fibras tendinosas que, por sua vez, se renem em feixes para formar os tendes musculares, para ento se inserirem nos ossos (Junqueira & Carneiro, 1990). Segundo Junqueira & Carneiro (1990), as miofibrilas esto suspensas no interior da fibra muscular numa matriz denominada sarcoplasma, que composta dos constituintes intracelulares habituais (corresponde ao citoplasma das demais clulas do corpo). O lquido sarcoplasmtico contm grandes quantidades de potssio, magnsio, fosfato e enzimas. Tambm existe grande nmero de mitocndrias, que se localizam entre e paralelamente s miofibrilas, o que indica grande necessidade de ATP formado naquelas organelas para que ocorra contrao das miofibrilas.
Fibras vermelhas e fibras brancas De acordo com a estrutura e composio bioqumica, as fibras musculares esquelticas podem ser classificadas como fibras do tipo I e fibras do tipo II. As fibras do tipo I so chamadas fibras lentas e as do tipo II so chamadas fibras rpidas. De cor vermelho-escura, as fibras do tipo I so ricas em sarcoplasma, com muita mioglobina; estas fibras so adaptadas para contraes contnuas e a
energia produzida a partir da fosforilao oxidativa de cidos graxos. As fibras do tipo II, porm as adaptadas para contraes rpidas e descontnuas; essas fibras contm pouca mioglobina e so de cor vermelho-clara. As fibras do tipo II podem ainda ser divididas em tipo A, B e C. Essa classificao feita de acordo com caractersticas funcionais e bioqumicas, como por exemplo, a estabilidade da ATPase actomiosina que elas contm. As fibras mais rpidas so do tipo II B e necessitam principalmente da gliclise como fonte de energia, como afirmam Junqueira & Carneiro (1990).
Biologia molecular da contrao Na dcada de 1950, Andrew Huxley e Ralph Niedergerke, e Hugh Huxley e Jean Hanson, independentemente, propuseram um modelo de filamentos deslizantes, baseados em estudo com raios X e microscopia tica e eletrnica. As caractersticas essenciais deste modelo so: Os comprimentos dos filamentos grossos e finos no mudam durante a contrao muscular; Ao contrrio, o comprimento do sarcmero diminui durante a contrao porque os dois tipos de filamento superpem-se mais. Os filamentos grossos e finos deslizam uns sobre os outros na contrao; A fora de contrao gerada por um processo que move ativamente um tipo de filamento ao longo dos filamentos vizinhos do outro tipo (Stryer, 1996). O encurtamento do sarcmero resultante do deslizamento dos filamentos de miosina pelos filamentos de actina sem mudana no seu comprimento. Este modelo do filamento deslizante, proposto pela primeira vez em 1954, foi crucial para a compreenso do mecanismo contrtil (Alberts et al., 1997). Segundo Alberts et al. (1997), a base estrutural da interao geradora de fora para o movimento visvel em micrografias eletrnicas de elevada ampliao. Os filamentos de miosina so vistos com numerosas extenses laterais, ou pontes transversais, com 13 nm de comprimento, que fazem contato com filamentos de actina adjacentes. Estas estruturas so formadas pelas cabeas de miosina. Quando o msculo se contrai, os filamentos de actina e de miosina so puxados uns por sobre os outros pelas pontes transversais que aturam ciclicamente, como uma srie de finos remos. Conforme foi anteriormente exposto, a cabea globular, ou domnio motor da miosina liga-se ao filamentos de actina e hidrolisa ATP. Cada molcula de actina num filamento de actina tem a capacidade de ligar uma cabea de miosina, formando um complexo que exibe a polaridade estrutural do filamento. Tais complexos, quando observados ao microscpio eletrnico aps colorao negativa, revelam uma morfologia regular e caracterstica: cada cabea de miosina forma uma projeo lateral e a imagem sobreposta de vrias dessas projees cria um aspecto de flechas ao longo do filamento. A extremidade penetrante da flecha de miosina corresponde extremidade menos de crescimento lento do filamento de actina (extremidade menos termina na linha medial - linha M - da zona H). A extremidade farpada da flecha corresponde extremidade mais de crescimento rpido (extremidade mais termina na linha Z). As cabeas da miosina se dispem em direes opostas em ambos os lados de uma regio central exposta do filamento de miosina. Uma vez que as cabeas devem interagir com os filamentos de actina na regio de sobreposio, estes devem estar dispostos com polaridades opostas nos sois lados do sarcmero. Conforme cita Stryer (1996), julga-se que o ciclo ATP-ADP na miosina produza movimento direcional da seguinte maneira:
1. No msculo em repouso, as cabeas da miosina so incapazes de interagir com as unidades de actina nos filamentos finos por causa da interferncia estrica da tropomiosina. Nesse estado, os produtos da hidrlise, ADP e Pi , ainda esto unidos miosina. 2. Quando o msculo estimulado, a tropomiosina muda de posio. As cabeas de miosina podem assim distender-se para fora do filamento grosso e ligar-se s unidades de actina nos filamentos finos. 3. A ligao da miosina-ADP-Pi , actina leva liberao de Pi. A dissociao subsequente do ADP induz uma grande alterao na conformao de miosina. A mudana na orientao de miosina em relao actina constitui o pulso de fora da contrao. O ADP liberado da miosina ao final do pulso de fora. 4. A subsequente ligao do ATP miosina leva rpida liberao de actina. A cabea de miosina novamente desprendida do filamento fino. 5. Finalmente, o ATP ligado hidrolisado da cabea livre da miosina, o que a recompe para a prxima interao com o filamento fino. A interao molecular geradora de fora recm-descrita, ocorre somente quando o msculo esqueltico recebe um sinal vindo do sue nervo motor. O sinal dispara um potencial de ao na membrana plasmtica da clula muscular e, esta excitao eltrica se espalha rapidamente por uma srie de dobras membranosas, os tbulos transversos, que se estendem desde a membrana plasmtica para dentro, envolvendo cada miofibrila. O sinal atravessa um pequeno espao e alcana ento o retculo sarcoplasmtico, um conjunto de vesculas achatadas e anastomosadas que circundam, como uma rede, cada miofibrila (Alberts et a., 1997). Alberts et al. (1997), descrevem a juno mioneural: na juno, grandes canais liberadores de Ca++ estendem-se como pilares a partir da membrana do retculo sarcoplasmtico, conectandose com a membrana do tbulo transverso (T) no lado oposto. Quando as protenas da membrana do tbulo T, sensveis voltagem, so ativadas pelo potencial de ao que est chegando, elas provocam a abertura de alguns dos canais de Ca ++, provavelmente por acoplamento mecnico direto. Os ons Ca++ so liberados do retculo sarcoplasmtico (onde esto armazenados em alta concentrao) na fenda da juno mioneural provocando a abertura de mais canais amplificando, assim, a resposta. A entrada de grande quantidade de Ca ++ no citoplasma inicia a contrao das miofibrilas. Todas as miofibrilas contraem ao mesmo tempo porque o sinal proveniente da membrana plasmtica da clula muscular transmitido, para cada sarcmero, num intervalo de milissegundos. O aumento da concentrao do Ca++ transitrio pois o on recaptado para o retculo sarcoplasmtico por uma Ca ++-ATPase existente na membrana do retculo. Em 30 milissegundos a concentrao do Ca++ citoplasmtico alcana nveis de repouso e as miofibrilas relaxam. A dependncia de Ca++ para a contrao do msculo esqueltico dos vertebrados e, portanto sua dependncia dos comandos motores transmitidos via nervos, inteiramente devida a uma grupo de protenas acessrias intimamente associadas aos filamentos de actina. Misturando-se miosina com filamentos de actina puros num tubo de ensaio, verifica-se que a atividade de ATPase da miosina estimulada mesmo na ausncia de Ca ++, numa miofibrila normal, por outro lado, na qual os filamentos de actina esto associados com protenas acessrias, a estimulao da atividade ATPsica da miosina dependente de Ca ++. A forma muscular da tropomiosina uma destas protenas acessrias, uma molcula com formato de basto (apresentada anteriormente) e que se liga no sulco da hlice de actina. Outra protena acessria importante envolvida na regulao pelo Ca++ do msculo esqueltico de vertebrados a troponina, formada por um complexo de trs polipeptdios - troponinas T, I e C. O complexo da troponina apresenta uma forma alongada, na qual as subunidades C e I formam uma regio globular e a T, uma longa cauda. A cauda da troponina T liga-se tropomiosina e, imagina-se que seja a responsvel pelo posicionamento do complexo no
filamento fino. A troponina I liga-se actina e, quando adicionada troponina T e tropomiosina, forma um complexo que inibe a interao da actina com a miosina, mesmo na presena de Ca++ (Alberts et al.,1997). . . Receptores musculares: Fuso Muscular e rgo Tendinoso de Golgi As terminaes musculares so mecanoceptores da sensibilidade tecidual profunda, que detectam deformao mecnica (Guyton, 1992). Os msculos e tendes tm uma quantidade abundante de dois tipos especiais de receptores: os fusos musculares e os rgos tendinosos de Golgi. A principal diferena entre o fuso muscular e o rgo tendinoso de Golgi que enquanto o fuso detecta o comprimento relativo do msculo e o rgo tendinoso detecta a tenso muscular. Fuso muscular: Cada fuso se localiza ao redor de trs a dez pequenas fibras musculares intrafusais, afiladas em suas extremidades e que se inserem nas bainhas das fibras musculares esquelticas extrafusais adjacentes. A poro receptora do fuso muscular est localizada na parte mdia entre suas duas extremidades, onde as fibras musculares intrafusais no tm quaisquer elementos contrteis. H dois tipos diferente de fibras intrafusais: fibras com bolsa nuclear (ncleos ficam agregados no interior de um saco expandido na poro central da rea receptora) e as fibras com cadeia nuclear (possuem ncleos alinhados em cadeia ao longa da rea receptora). O fuso pode ser excitado de duas maneiras diferentes: pelo estriamento de todo o msculo e pela contrao das pores terminais das fibras intrafusais. Observa-se dois tipos de terminaes sensoriais na regio receptora do fuso muscular: terminao primria (no centro da zona receptora, uma fibra grande envolve as pores centrais das fibras intrafusais formando esta terminao que transmite impulsos para a medula numa velocidade de 100m/s) e a terminao secundria (duas fibras nervosas sensoriais menores inervam a regio receptora em cada lado da terminao primria; podem ser chamadas tambm de terminaes em buqu de flores). Quando a poro receptora do fuso muscular estirada lentamente, o nmero de impulsos transmitidos, tanto pela terminao primria como pela secundria, aumenta quase diretamente em proporo ao grau de estiramento, e as terminaes continuam a transmitir esses impulsos durante muitos minutos. Esse efeito chamado de resposta esttica do receptor do fuso e significa simplesmente que tanto a terminao primria como a secundria continuam a transmitir seus sinais durante todo o tempo que o receptor permanecer estirado. Como apenas as fibras intrafusais do tipo com cadeia nuclear so inervadas por terminaes tanto primria como secundria, acredita-se que essas fibras com cadeias nuclear sejam responsveis pela resposta esttica. Quando o comprimento do receptor do fuso aumenta subitamente, a terminao primria (mas no a terminao secundria) estimulada. Esse estmulo excessivo da terminao primria chamado de resposta dinmica, que significa que a terminao primria responde de forma extremamente ativa rpida velocidade de modificao do comprimento. Imediatamente aps ser sustado o aumento de comprimento do receptor, a freqncia de descarga dos impulsos volta a nvel muito baixo, correspondente resposta esttica. Ao contrrio, quando h o encurtamento do receptor do fuso, essa modificao diminui momentaneamente a freqncia de impulsos provenientes da terminao primria. Portanto, a terminao primria envia ao sistema nervoso central sinais de grande intensidade alertando-o sobre qualquer alterao do comprimento da rea receptora do fuso (Guyton, 1992). rgo tendinoso de Golgi: situam-se dentro dos tendes musculares e imediatamente adiante de suas inseres nas fibras musculares. um rgo estimulado pela tenso produzida por esse pequeno feixe de fibras musculares. O rgo tendinoso tm uma resposta dinmica e uma resposta esttica, respondendo com intensidade quando a tenso do msculo aumenta subitamente, mas dentro de uma pequena frao de segundo ele se acomoda em um nvel inferior de disparo constante, que quase diretamente proporcional tenso muscular. Assim, o rgo tendinoso de
Golgi proporciona ao sistema nervoso uma informao instantnea do grau de tenso de cada pequeno segmento de cada msculo. As fibras do tipo Ib transmitem sinais tanto para reas localizadas da medula como para reas cerebrais distantes, tais como cerebelo e crtex cerebral por meio de vias longas, tais como os feixes espinocerebelares e outros feixes para o crtex cerebral. O sinal local na medula excita interneurnio inibitrio nico que, por sua vez, inibe o motoneurnio anterior. Esse circuito local inibe diretamente o msculo individual, sem afetar os msculos adjacentes. Por vezes, o reflexo causado pela estimulao do rgo tendinoso quando submetido a um aumento da tenso muscular (os sinais so transmitidos para a medula espinhal para causar efeitos reflexos no prprio msculo estimulado) inteiramente inibitrio, exatamente contrrio ao reflexo do fuso muscular. Outra provvel funo do reflexo do rgo de Golgi a de equalizar as foras contrteis das fibras musculares dispersas, isto , as fibras que esto exercendo tenso excessiva so inibidas, enquanto as que esto exercendo tenso muito baixa tornam-se mais excitadas (Guyton, 1992).
Bibliografia Consultada Alberts, B; Bray, D; Lewis, J; Raff, M; Roberts, K; Watson, J. D. (1997) Biologia Molecular da Clula.Traduzido por Amauri Braga. 3.ed. Porto Alegre: Artes Mdicas. Guyton, A.C. (1992) Tratado de Fisiologia Mdica. 8.ed. Rio de Janeiro: Guanabara Koogan. Junqueira, L.C.; Carneiro, J. (1990) Histologia bsica. 7.ed. Rio de Janeiro: Guanabara Koogan. Stryer L.(1996) Bioqumica. 4.ed. Rio de Janeiro, Guanabara Koogan.
You might also like
- Fisiologia Do Musc EsqueléticoDocument32 pagesFisiologia Do Musc EsqueléticosalvadoorjrNo ratings yet
- ARTIGO - Fisiologia Do Músculo EsqueléticoDocument32 pagesARTIGO - Fisiologia Do Músculo EsqueléticoKeyla SilvaNo ratings yet
- Tecidos Musculares 40Document3 pagesTecidos Musculares 40Raquel ChequeleiroNo ratings yet
- Tecido Muscular - ResumoDocument6 pagesTecido Muscular - ResumoHelena MarcondesNo ratings yet
- Resumo Tecido Muscular - JunqueiraDocument4 pagesResumo Tecido Muscular - JunqueiraLucas Thiago BorgesNo ratings yet
- #Resumo#Document10 pages#Resumo#Monitoriadefisio100% (2)
- Tecido MuscularDocument4 pagesTecido MuscularDeadmanNo ratings yet
- Medresumos 2016 - OmfDocument16 pagesMedresumos 2016 - OmfMirianNo ratings yet
- Cabulas Dos Musculos 1Document1 pageCabulas Dos Musculos 1api-26429188No ratings yet
- FisiologiaDocument38 pagesFisiologiaAleida SantosNo ratings yet
- Movimento humano ATP músculos esqueléticos contraçãoDocument7 pagesMovimento humano ATP músculos esqueléticos contraçãoJoyceMedeirosNo ratings yet
- Sistema Muscular - FisiologiaDocument5 pagesSistema Muscular - FisiologiaKauan GabrielNo ratings yet
- Unidade Motora e Suas SindromesDocument39 pagesUnidade Motora e Suas SindromesCibele BragaNo ratings yet
- Movimento e características do tecido muscularDocument3 pagesMovimento e características do tecido muscularcedaspy cedaspyNo ratings yet
- Bioquímica da contração muscular: filamentos deslizantesDocument10 pagesBioquímica da contração muscular: filamentos deslizanteshellensabinoNo ratings yet
- Apg 20 - Semestre 2Document4 pagesApg 20 - Semestre 2Bruna BuenoNo ratings yet
- Tecido MuscularDocument7 pagesTecido MuscularRebeca SoaresNo ratings yet
- Aula 021659031827Document12 pagesAula 021659031827Jurandi AraujoNo ratings yet
- OpenAccess Lorente 9788580393996 08Document12 pagesOpenAccess Lorente 9788580393996 08Larissa Medicina veterináriaNo ratings yet
- Fisiologia muscular - estrutura e contraçãoDocument33 pagesFisiologia muscular - estrutura e contraçãoCaio RosignoliNo ratings yet
- Resumo Contração MuscularDocument11 pagesResumo Contração MuscularThiago PontesNo ratings yet
- Sistema muscular: tipos, estrutura e contraçãoDocument10 pagesSistema muscular: tipos, estrutura e contraçãoSuellen OliveiraNo ratings yet
- Junção NeuroDocument33 pagesJunção NeuroFafnir Kesk100% (2)
- Resumo Tecido MuscularDocument5 pagesResumo Tecido MuscularLaryssa AmorimNo ratings yet
- Ergonomia 5Document16 pagesErgonomia 5Jane CastroNo ratings yet
- Tecido MuscularDocument6 pagesTecido MuscularLeo RamosNo ratings yet
- Fisiologia MuscularDocument17 pagesFisiologia MuscularMaria Baião BasílioNo ratings yet
- 13 Tec MuscularDocument47 pages13 Tec MuscularmalubastosNo ratings yet
- Ergonomia 5Document16 pagesErgonomia 5Nicolle HenriquesNo ratings yet
- Sistema muscular: estrutura e contraçãoDocument18 pagesSistema muscular: estrutura e contraçãoGessica Fabiana Cardoso0% (1)
- HistologiaPL - Tecido MuscularDocument35 pagesHistologiaPL - Tecido Muscularviolleta gomia100% (1)
- Trabalho Do Sistema MuscularDocument18 pagesTrabalho Do Sistema MuscularkynhoNo ratings yet
- Tecido MuscularDocument22 pagesTecido MuscularAna Clara FrotaNo ratings yet
- Fisiologia Do Sistema MuscularDocument17 pagesFisiologia Do Sistema MuscularKaillany Andrade NaivaNo ratings yet
- Sistema muscular: Mioblastos e formação das fibras muscularesDocument43 pagesSistema muscular: Mioblastos e formação das fibras muscularesIvanCostaPassosNo ratings yet
- Músculo esquelético: estrutura e contraçãoDocument5 pagesMúsculo esquelético: estrutura e contraçãomariany100% (2)
- Trabalho de BiologiaDocument5 pagesTrabalho de BiologiaemileNo ratings yet
- Tecido MuscularDocument5 pagesTecido MuscularLívia TipografiasNo ratings yet
- Resumo Fisiologia MuscularDocument6 pagesResumo Fisiologia MuscularDaniel SilvaNo ratings yet
- Tecido MuscularDocument9 pagesTecido MuscularIsadora RuthNo ratings yet
- Resumo - Módulo 1 - Fisiologia Do ExercícioDocument23 pagesResumo - Módulo 1 - Fisiologia Do ExercícioLarissa PammelaNo ratings yet
- Trabalho FisiologiaDocument2 pagesTrabalho FisiologiaFábio PereiraNo ratings yet
- Resumo Tecido Muscular Histologia e Fisiologia Da ContraçãoDocument10 pagesResumo Tecido Muscular Histologia e Fisiologia Da ContraçãoIgor BarbosaNo ratings yet
- Arquitetura muscular e contraçãoDocument120 pagesArquitetura muscular e contraçãoMagno DuarteNo ratings yet
- Músculo Estriado Esquelético e suas Características HistológicasDocument1 pageMúsculo Estriado Esquelético e suas Características HistológicasDavi CastroNo ratings yet
- Fechamento Tutoria 12Document12 pagesFechamento Tutoria 12vanessa da silva caetanoNo ratings yet
- HjjnhiDocument5 pagesHjjnhiZeka Zeki ZokoNo ratings yet
- Estrutura e componentes do músculo estriado esqueléticoDocument9 pagesEstrutura e componentes do músculo estriado esqueléticoRafael FontesNo ratings yet
- Tecidos musculares estriados e lisosDocument6 pagesTecidos musculares estriados e lisosvanuzaNo ratings yet
- Tecidos musculares: estrutura e funçãoDocument8 pagesTecidos musculares: estrutura e funçãoJOAO VICTOR COSTA LUSTOSANo ratings yet
- 1contração Do Músculo EsqueléticoDocument63 pages1contração Do Músculo EsqueléticoKelly Coelho RochaNo ratings yet
- Unid 6Document26 pagesUnid 6marcelisousadasilvaNo ratings yet
- Resenha Condromalacia PatelarDocument2 pagesResenha Condromalacia PatelarThiago Oliveira de SousaNo ratings yet
- Fisiologia Da Contrac3a7c3a3o Muscular - RoDocument36 pagesFisiologia Da Contrac3a7c3a3o Muscular - RoMarcelo CostaNo ratings yet
- Aula 3 Contração MuscularDocument51 pagesAula 3 Contração MuscularBila BernardesNo ratings yet
- Contraà à o MuscularDocument5 pagesContraà à o Muscularvitoriamylena9No ratings yet
- Tecido MuscularDocument9 pagesTecido MuscularSilmara SilvaNo ratings yet
- Fundamentos Quânticos Das Práticas IntegrativasFrom EverandFundamentos Quânticos Das Práticas IntegrativasNo ratings yet
- O Poder Do Quântico - Consciência ThetaDocument28 pagesO Poder Do Quântico - Consciência ThetaLuciani Moreira100% (8)
- O Poder Do Quântico - Consciência ThetaDocument28 pagesO Poder Do Quântico - Consciência ThetaLuciani Moreira100% (8)
- Mapa Das ComarcasDocument1 pageMapa Das ComarcasLuciani MoreiraNo ratings yet
- Cronico DegenerativoDocument84 pagesCronico DegenerativoLuciani MoreiraNo ratings yet
- Reprogramando suas crenças limitantes em 4 passosDocument55 pagesReprogramando suas crenças limitantes em 4 passosPasor Mauricio Brito67% (3)
- Ebook Receitas de Café Da Manhã Low Carb PDFDocument39 pagesEbook Receitas de Café Da Manhã Low Carb PDFQue LanceNo ratings yet
- Desperte o Seu PotencialDocument34 pagesDesperte o Seu PotencialLuciani MoreiraNo ratings yet
- O Melhor Ano MulheresDocument26 pagesO Melhor Ano MulheresCris SalesNo ratings yet
- Aula 4 6 Ato Administrativo Val Eficácia - Atributos - Extinção Desfazimento e Sanatória PDFDocument188 pagesAula 4 6 Ato Administrativo Val Eficácia - Atributos - Extinção Desfazimento e Sanatória PDFLuciani MoreiraNo ratings yet
- Ficha de Controle de ProntuáriosDocument2 pagesFicha de Controle de ProntuáriosLuciani MoreiraNo ratings yet
- RPGDocument64 pagesRPGLuciani Moreira100% (1)
- FACIPE+ +DA+I+ +PONTO+04+ +poder Dever+AdministrativoDocument8 pagesFACIPE+ +DA+I+ +PONTO+04+ +poder Dever+AdministrativoTacy NascimentoNo ratings yet
- Manual SeiDocument54 pagesManual SeiLuciani MoreiraNo ratings yet
- Método Pilates No Tratamento de Processos Álgicos Da Coluna Vertebral - Uma Revisão BibliográficaDocument13 pagesMétodo Pilates No Tratamento de Processos Álgicos Da Coluna Vertebral - Uma Revisão BibliográficaLuciani MoreiraNo ratings yet
- 2016 Nutrição Perda de Peso AdultoDocument2 pages2016 Nutrição Perda de Peso AdultoLuciani MoreiraNo ratings yet
- Obesidade Na Infancia e Adolescencia Manual de OrientacaoDocument120 pagesObesidade Na Infancia e Adolescencia Manual de OrientacaoLidiany BarrosNo ratings yet
- Art. 205 A 2014Document7 pagesArt. 205 A 2014Luciani MoreiraNo ratings yet
- Art. 205 A 2014Document7 pagesArt. 205 A 2014Luciani MoreiraNo ratings yet
- Orientação Diabetes MellitusDocument1 pageOrientação Diabetes MellitusLuciani MoreiraNo ratings yet
- Alimentos Que Ajudam A Dormir MelhorDocument1 pageAlimentos Que Ajudam A Dormir MelhorLuciani MoreiraNo ratings yet
- Anamnese CardiológicaDocument12 pagesAnamnese CardiológicaIsabelle Lopes GontijoNo ratings yet
- Segurança Do PacienteDocument10 pagesSegurança Do PacienteLuciani MoreiraNo ratings yet
- Requisitos boas práticas processamento produtos saúdeDocument17 pagesRequisitos boas práticas processamento produtos saúdeLuciani MoreiraNo ratings yet
- Lista de refeições e alimentos do Centro Universitário FranciscanoDocument2 pagesLista de refeições e alimentos do Centro Universitário FranciscanoLuciani Moreira100% (1)
- Sinais VitaisDocument12 pagesSinais VitaisLuciani Moreira100% (1)
- Ética e HumanizaçãoDocument12 pagesÉtica e HumanizaçãoLuciani MoreiraNo ratings yet
- Lei #11.091Document36 pagesLei #11.091Luciani MoreiraNo ratings yet
- Re Sumo Adm Rec Human OsDocument4 pagesRe Sumo Adm Rec Human OsLuciani MoreiraNo ratings yet
- Manual Tecnico Limpeza HospitalarDocument25 pagesManual Tecnico Limpeza HospitalarJOAQUIMCOXIMNo ratings yet
- Apresentação 1Document1 pageApresentação 1Luciani MoreiraNo ratings yet
- Guia 1 de AntroDocument7 pagesGuia 1 de AntroSao TrầnNo ratings yet
- Terminologia AnatômicaDocument1 pageTerminologia AnatômicaAlineNo ratings yet
- Fichas Trabalho 1a10Document16 pagesFichas Trabalho 1a10Joaquim BarbosaNo ratings yet
- Órgãos Yin e Yang na Medicina Tradicional ChinesaDocument1 pageÓrgãos Yin e Yang na Medicina Tradicional ChinesathelmalowenNo ratings yet
- Vertebrados: répteis, aves e mamíferos - exercíciosDocument7 pagesVertebrados: répteis, aves e mamíferos - exercíciosmary5stela5camillatoNo ratings yet
- O trato gastrintestinalDocument8 pagesO trato gastrintestinalCarolina StrongerNo ratings yet
- Estômago, intestino e sistema digestórioDocument13 pagesEstômago, intestino e sistema digestórioAna Leonor Fernandez CardozoNo ratings yet
- Tecido EpitelialDocument27 pagesTecido EpitelialNeila carine Silva MeiraNo ratings yet
- Lista de Alguns Livros de BiologiaDocument17 pagesLista de Alguns Livros de BiologiaRodrigo Dumas Saavedra100% (1)
- Sistema muscular: músculos, funções e anatomiaDocument51 pagesSistema muscular: músculos, funções e anatomiaAnnalethycia Pereira100% (2)
- Relações EcológicasDocument6 pagesRelações EcológicasAna100% (1)
- Relações ecológicas entre espéciesDocument1 pageRelações ecológicas entre espéciesPaulo HenriqueNo ratings yet
- Sistema TegumentarDocument44 pagesSistema TegumentarAnonymous 85j0VSCv100% (2)
- Estrutura e funções dos ossosDocument2 pagesEstrutura e funções dos ossosAlice Sodre0% (2)
- E Book Biologia de Animais Marinhos 2Document112 pagesE Book Biologia de Animais Marinhos 2ufracbbacharel2022No ratings yet
- Introdução À GenéticaDocument28 pagesIntrodução À GenéticaKatiuscia HasegawaNo ratings yet
- Histologia do Aparelho Reprodutor MasculinoDocument2 pagesHistologia do Aparelho Reprodutor MasculinoAna Carolina OliveiraNo ratings yet
- Trabalho AnatomiaDocument8 pagesTrabalho AnatomiaFlávia BelmonteNo ratings yet
- Teste Nº1 - Organização e Sist. ReprodutorDocument6 pagesTeste Nº1 - Organização e Sist. ReprodutorJoana RibeiroNo ratings yet
- Apostila Sistema Respiratorio by Taina Carvalho 4 220914 172602Document45 pagesApostila Sistema Respiratorio by Taina Carvalho 4 220914 172602sophia carolineNo ratings yet
- Nervos Do Corpo Humano - Toda MatériaDocument3 pagesNervos Do Corpo Humano - Toda MatériaRodrigo FerrazNo ratings yet
- Sistema TegumentarDocument5 pagesSistema TegumentarSara AleixoNo ratings yet
- Respiração, Aeração Ou VentilaçãoDocument23 pagesRespiração, Aeração Ou VentilaçãoBruno FelipeNo ratings yet
- Biologia Dos Vertebrados PDFDocument64 pagesBiologia Dos Vertebrados PDFFCiências100% (4)
- TECIDOSDocument1 pageTECIDOSMaurim OlendinoNo ratings yet
- HexapodaDocument10 pagesHexapodaAmanda PratesNo ratings yet
- Músculos faciais da mímica e expressãoDocument2 pagesMúsculos faciais da mímica e expressãocabfgNo ratings yet
- Termoregulação TesticularDocument5 pagesTermoregulação TesticularDiego Moraes Soares MoraesNo ratings yet
- TESTE 11ºbiologia - Módulo A3 - Distribuição Nos AnimaisDocument5 pagesTESTE 11ºbiologia - Módulo A3 - Distribuição Nos AnimaisIsaura SilvaNo ratings yet
- Avaliação Ciência Sistema Reprodutor Métodos Contracepção DSTsDocument5 pagesAvaliação Ciência Sistema Reprodutor Métodos Contracepção DSTsMichele MoreiraNo ratings yet