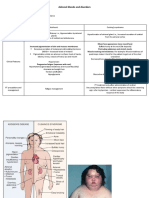
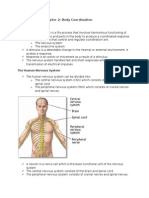
You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Med-Surg Nursing NotesDocument232 pagesMed-Surg Nursing NotesMaria100% (4)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Chapter 18 Common Drugs PDFDocument221 pagesChapter 18 Common Drugs PDFlalallalaNo ratings yet
- Endocrine System Assessment.Document30 pagesEndocrine System Assessment.ramoli1988100% (2)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Self-Assessment Colour Review of Small Animal Soft Tissue SurgeryDocument194 pagesSelf-Assessment Colour Review of Small Animal Soft Tissue Surgerymiliindianu100% (3)
- Therapeutic Parsley Drink Recipe, SymptometryDocument10 pagesTherapeutic Parsley Drink Recipe, SymptometryMalik SayeedNo ratings yet
- NeurotransmittersDocument4 pagesNeurotransmittersSalman Khan100% (1)
- The Dates and Lemon Detox Protocol Sheet Official V-1.2Document20 pagesThe Dates and Lemon Detox Protocol Sheet Official V-1.2Shaun Oneill100% (1)
- Hormone HandoutDocument2 pagesHormone Handoutszisz889No ratings yet
- Treatment of Hypoglycemia in ChildrenDocument16 pagesTreatment of Hypoglycemia in ChildrenKamuiShirouNo ratings yet
- Medicine Alaa Notes (All Files Merged) - Best Pro Choice-.Pdf-1Document120 pagesMedicine Alaa Notes (All Files Merged) - Best Pro Choice-.Pdf-1maimoona suleman100% (6)
- Management of of Clients With Metabolic and EndocrineDocument79 pagesManagement of of Clients With Metabolic and Endocrineagbogoodness100% (1)
- Unit 1 Study GuideDocument6 pagesUnit 1 Study Guideangieswenson100% (4)
- Pheochromocytoma Case Study: 51-Year-Old Woman After AdrenalectomyDocument14 pagesPheochromocytoma Case Study: 51-Year-Old Woman After Adrenalectomydidu91No ratings yet
- Addisons Vs CushingDocument2 pagesAddisons Vs Cushingshifali guptaNo ratings yet
- Endocrine SystemDocument52 pagesEndocrine Systemnurshuhada zainuddinNo ratings yet
- Endocrine System 2Document27 pagesEndocrine System 2Naghman ZuberiNo ratings yet
- Activity 2 MetabsDocument10 pagesActivity 2 MetabsCalvin Keith YadaoNo ratings yet
- Glucocorticoid DeflazacortDocument25 pagesGlucocorticoid DeflazacortChintan ChavdaNo ratings yet
- ChoeDocument116 pagesChoeJihad AnadNo ratings yet
- Stress Dose SteroidsDocument4 pagesStress Dose SteroidsTitien fitria sholihati100% (1)
- Form 4 Science Chapter 2: Body Coordination (38Document13 pagesForm 4 Science Chapter 2: Body Coordination (38Cheah Foo Kit50% (2)
- SPM Science Notes - Body CoordinationDocument12 pagesSPM Science Notes - Body CoordinationSPM98% (50)
- Essentials of Abnormal Psychology 8th Edition Durand Test Bank DownloadDocument38 pagesEssentials of Abnormal Psychology 8th Edition Durand Test Bank DownloadcarrielivingstonocifrtkgnyNo ratings yet
- Adrenal Function, Adrenal Hyperfunction and HypofunctionDocument98 pagesAdrenal Function, Adrenal Hyperfunction and HypofunctionAntony WaithakaNo ratings yet
- Surgery WiseMDDocument61 pagesSurgery WiseMDJameson87No ratings yet
- Lesson 9: Endocrine System: Prepared by Arianne V. JulianDocument34 pagesLesson 9: Endocrine System: Prepared by Arianne V. JulianGisselleNo ratings yet
- Addison Vs CushingsDocument1 pageAddison Vs CushingsChris ZantiraNo ratings yet
- Gender Midterm NotesDocument27 pagesGender Midterm Noteslandrade3440No ratings yet
- MHT-CET Biology Previous Solved Papers - Sample ContentDocument30 pagesMHT-CET Biology Previous Solved Papers - Sample Contentssspd.entNo ratings yet