You might also like
- Evolution of Flow Cytometry As A Drug Screening PlatformDocument3 pagesEvolution of Flow Cytometry As A Drug Screening PlatformdavdavdavdavdavdavdaNo ratings yet
- Rheinfelden 28th February/3rd March 1638!: Imperialist Brief 1Document1 pageRheinfelden 28th February/3rd March 1638!: Imperialist Brief 1davdavdavdavdavdavdaNo ratings yet
- Cd4's and IL-10Document12 pagesCd4's and IL-10davdavdavdavdavdavdaNo ratings yet
- GustavPlayBook PDFDocument48 pagesGustavPlayBook PDFdavdavdavdavdavdavdaNo ratings yet
- GneisenauDocument4 pagesGneisenaudavdavdavdavdavdavdaNo ratings yet
- Rheinfelden 28th February/3rd March 1638!: Protestant Brief 1Document1 pageRheinfelden 28th February/3rd March 1638!: Protestant Brief 1davdavdavdavdavdavdaNo ratings yet
- TAL Enemy BattalionDocument2 pagesTAL Enemy BattaliondavdavdavdavdavdavdaNo ratings yet
- Iphone 6s Manual User GuideDocument196 pagesIphone 6s Manual User GuideSamuel ShepherdNo ratings yet
- Ni 2987Document11 pagesNi 2987davdavdavdavdavdavdaNo ratings yet
- GustavPlayBook PDFDocument48 pagesGustavPlayBook PDFdavdavdavdavdavdavdaNo ratings yet
- SiA Playbook FINAL-1Document56 pagesSiA Playbook FINAL-1davdavdavdavdavdavdaNo ratings yet
- Intracellular signaling pathwaysDocument2 pagesIntracellular signaling pathwaysdavdavdavdavdavdavdaNo ratings yet
- BLK Qtrly Commentary 2015 q3 AmersDocument14 pagesBLK Qtrly Commentary 2015 q3 AmersdavdavdavdavdavdavdaNo ratings yet
- Sfo PlaybookDocument40 pagesSfo PlaybookdavdavdavdavdavdavdaNo ratings yet
- Indg 330Document3 pagesIndg 330davdavdavdavdavdavdaNo ratings yet
- Expression of A-Defensin 1-3 in T CellsDocument8 pagesExpression of A-Defensin 1-3 in T CellsdavdavdavdavdavdavdaNo ratings yet
- John Guest Speedfit Catalog Auguet 2014 EN PDFDocument48 pagesJohn Guest Speedfit Catalog Auguet 2014 EN PDFdavdavdavdavdavdavdaNo ratings yet
- Ni 2499Document11 pagesNi 2499davdavdavdavdavdavdaNo ratings yet
- Science 2013 Millien 792 6Document6 pagesScience 2013 Millien 792 6davdavdavdavdavdavdaNo ratings yet
- NM 3670Document6 pagesNM 3670davdavdavdavdavdavdaNo ratings yet
- Shipping Guide IX Final 2009Document28 pagesShipping Guide IX Final 2009davdavdavdavdavdavdaNo ratings yet
- Bassoe PaperDocument9 pagesBassoe PaperdavdavdavdavdavdavdaNo ratings yet
- French Clothing in The TYWDocument14 pagesFrench Clothing in The TYWdavdavdavdavdavdavda100% (1)
- 00 - All OOBs (Orders of Battle) in One DocumentDocument244 pages00 - All OOBs (Orders of Battle) in One Documentairfix199975% (4)
- Leflunomide & AdenosineDocument10 pagesLeflunomide & AdenosinedavdavdavdavdavdavdaNo ratings yet
- JC SupplementalDocument5 pagesJC SupplementaldavdavdavdavdavdavdaNo ratings yet
- Piis2213858713701657 2Document14 pagesPiis2213858713701657 2davdavdavdavdavdavdaNo ratings yet
- Embj 201386030 FullDocument17 pagesEmbj 201386030 FulldavdavdavdavdavdavdaNo ratings yet
- MHC Class I and Class I-like gene expression in malignant T cellsDocument5 pagesMHC Class I and Class I-like gene expression in malignant T cellsdavdavdavdavdavdavdaNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Brain and Cranial NervesDocument82 pagesThe Brain and Cranial NervesImmanuel Ronald LewisNo ratings yet
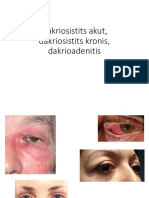
- Dakriosistits Akut, Dakriosistits Kronis, DakrioadenitisDocument37 pagesDakriosistits Akut, Dakriosistits Kronis, DakrioadenitisyuliasmindeNo ratings yet
- NAT Reviewer ScienceDocument3 pagesNAT Reviewer ScienceRandy100% (1)
- Digestive System EmbryologyDocument104 pagesDigestive System EmbryologyAlistair WalkerNo ratings yet
- English Exercise Book 3 Eso A: Ies Canónigo ManchónDocument36 pagesEnglish Exercise Book 3 Eso A: Ies Canónigo ManchónrobertoquinteiroNo ratings yet
- George Orwell's Animal Farm: A concise study guideDocument41 pagesGeorge Orwell's Animal Farm: A concise study guidetracy zvenhamo100% (1)
- Heaven & Earth Spa Treatments Under 40Document2 pagesHeaven & Earth Spa Treatments Under 40Janhavi SoniNo ratings yet
- 24XX Dying Sun 1.1 SPREADSDocument2 pages24XX Dying Sun 1.1 SPREADSEvandro FormozoNo ratings yet
- Open and Closed Packed Positions of JointsDocument1 pageOpen and Closed Packed Positions of JointsRaymond100% (4)
- Histology PlacentaDocument26 pagesHistology Placentakhiks34100% (2)
- CounselorDocument45 pagesCounselorChristian Jett SowoneulNo ratings yet
- Live Broadcasting Raising Dragons in The Interstellar (Unknown) (Z-Library)Document702 pagesLive Broadcasting Raising Dragons in The Interstellar (Unknown) (Z-Library)mddgrcvfNo ratings yet
- Jump To Navigationjump To Search: Kermode BearDocument4 pagesJump To Navigationjump To Search: Kermode Bearswahyudi9No ratings yet
- Inactivation of Viruses: June 2011Document13 pagesInactivation of Viruses: June 2011Iqbal Muhammad Yaa-BegitulahNo ratings yet
- Forensic Hair AnalysisDocument31 pagesForensic Hair AnalysisJauhar FirdausNo ratings yet
- Xii - Vivential ExamDocument4 pagesXii - Vivential ExamKatherin Rodriguez ZavaletaNo ratings yet
- 100 Ways To Tell If You Are FilipinoDocument4 pages100 Ways To Tell If You Are FilipinoAbygail Pasco MagbagNo ratings yet
- 14 Sop Got Electric ShockDocument3 pages14 Sop Got Electric ShockRio SimatupangNo ratings yet
- Fishes of The MaldivesDocument391 pagesFishes of The MaldivesIbrahim Asad100% (1)
- Emperor Penguins Are Birds That Live in Antarctica. They Are The Largest Species of PenguinDocument8 pagesEmperor Penguins Are Birds That Live in Antarctica. They Are The Largest Species of PenguinThe Children's NurturyNo ratings yet
- Stem Cell Dan Produknya Di UnnesDocument56 pagesStem Cell Dan Produknya Di UnnesemyNo ratings yet
- Ion ChannelsDocument17 pagesIon ChannelsFrancia ToledanoNo ratings yet
- How to Make Homemade BaconDocument11 pagesHow to Make Homemade Baconpilapil_j100% (1)
- YDh.1.Mitaksara - Tr.gharpure - Pt2mitāk Arā of Vijñāneśvara On Yājñavalkya-Dharmaśāstra, PT 2 of Gharpure's Translation.Document240 pagesYDh.1.Mitaksara - Tr.gharpure - Pt2mitāk Arā of Vijñāneśvara On Yājñavalkya-Dharmaśāstra, PT 2 of Gharpure's Translation.Tim LubinNo ratings yet
- Kindergarten classroom weekly activitiesDocument13 pagesKindergarten classroom weekly activitiesMarian Triguero SaldiNo ratings yet
- Mountain Terrors IntroductionDocument13 pagesMountain Terrors IntroductionGeraldine VasquezNo ratings yet
- Autopsy of Heart External ExaminationDocument20 pagesAutopsy of Heart External ExaminationchinnnababuNo ratings yet
- Atlas Games2Document2 pagesAtlas Games2Beki LokaNo ratings yet
- Animal Kingdom Digital NotesDocument20 pagesAnimal Kingdom Digital NotesKshreeNo ratings yet
- Ivan Petrovic PavlovDocument3 pagesIvan Petrovic PavlovNazreen Ayein Shukor100% (1)