You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Charcoal RefDocument7 pagesCharcoal RefAmit KumarNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Activated Carbon For Purification of AlcoholDocument28 pagesActivated Carbon For Purification of AlcoholcmdcscribdNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Copper RoleDocument8 pagesCopper RoleAmit KumarNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
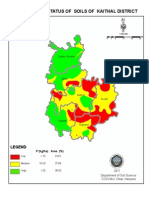
- Phosphorus Status of Soils of Kaithal District: LegendDocument1 pagePhosphorus Status of Soils of Kaithal District: LegendAmit KumarNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Role of Cu and ZN in Beef CycleDocument12 pagesRole of Cu and ZN in Beef CycleAmit KumarNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Mecovet Dose ReferenceDocument4 pagesMecovet Dose ReferenceAmit KumarNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Presentation NutritionDocument82 pagesPresentation NutritionAmit KumarNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Erythrocyte replacement and Methylcobalamin deficiency anemiaDocument1 pageErythrocyte replacement and Methylcobalamin deficiency anemiaAmit KumarNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Drugs ListDocument195 pagesDrugs ListAmit Kumar100% (3)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- Flood FillDocument1 pageFlood FillshubhamNo ratings yet
- IEQ CompleteDocument19 pagesIEQ Completeharshal patilNo ratings yet
- VivsayamDocument87 pagesVivsayamvalarumsakthi100% (2)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- WL 318 PDFDocument199 pagesWL 318 PDFBeckty Ahmad100% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- HSC 405 Grant ProposalDocument23 pagesHSC 405 Grant Proposalapi-355220460100% (2)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Oral Nutrition Support NotesDocument28 pagesOral Nutrition Support Notesleemon.mary.alipao8695No ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Plate-Load TestDocument20 pagesPlate-Load TestSalman LakhoNo ratings yet
- Rectifiers and FiltersDocument68 pagesRectifiers and FiltersMeheli HalderNo ratings yet
- Interactive Architecture Adaptive WorldDocument177 pagesInteractive Architecture Adaptive Worldhoma massihaNo ratings yet
- Convocation ProgramDocument125 pagesConvocation ProgramZirak TayebNo ratings yet
- Soil LiquefactionDocument12 pagesSoil LiquefactionKikin Kikin PelukaNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Madeleine Ker - TakeoverDocument91 pagesMadeleine Ker - Takeover66677785100% (1)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- 07 Raction KineticsDocument43 pages07 Raction KineticsestefanoveiraNo ratings yet
- Who will buy electric vehicles Segmenting the young Indian buyers using cluster analysisDocument12 pagesWho will buy electric vehicles Segmenting the young Indian buyers using cluster analysisbhasker sharmaNo ratings yet
- Ro-Buh-Qpl: Express WorldwideDocument3 pagesRo-Buh-Qpl: Express WorldwideverschelderNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Apu Trilogy - Robin Wood PDFDocument48 pagesThe Apu Trilogy - Robin Wood PDFSamkush100% (1)
- Conjoint Analysis Basic PrincipleDocument16 pagesConjoint Analysis Basic PrinciplePAglu JohnNo ratings yet
- Design of Fixed Column Base JointsDocument23 pagesDesign of Fixed Column Base JointsLanfranco CorniaNo ratings yet
- An Online ECG QRS Detection TechniqueDocument6 pagesAn Online ECG QRS Detection TechniqueIDESNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Stability Calculation of Embedded Bolts For Drop Arm Arrangement For ACC Location Inside TunnelDocument7 pagesStability Calculation of Embedded Bolts For Drop Arm Arrangement For ACC Location Inside TunnelSamwailNo ratings yet
- Madu Rash Tak AmDocument4 pagesMadu Rash Tak AmAdv. Govind S. TehareNo ratings yet
- Elements of ClimateDocument18 pagesElements of Climateእኔ እስጥፍNo ratings yet
- Descripcion Unidad 9, Dos CiudadesDocument13 pagesDescripcion Unidad 9, Dos CiudadesGabriela ValderramaNo ratings yet
- What Is DSP BuilderDocument3 pagesWhat Is DSP BuilderĐỗ ToànNo ratings yet
- Rotary Twin Scew Brochure UK HRDocument20 pagesRotary Twin Scew Brochure UK HRNguyễn Hữu DũngNo ratings yet
- Lightwave Maya 3D TutorialsDocument8 pagesLightwave Maya 3D TutorialsrandfranNo ratings yet
- Interpreting Piping and Instrumentation DiagramsDocument41 pagesInterpreting Piping and Instrumentation DiagramsFredric Tun100% (2)
- The Templist Scroll by :dr. Lawiy-Zodok (C) (R) TMDocument144 pagesThe Templist Scroll by :dr. Lawiy-Zodok (C) (R) TM:Lawiy-Zodok:Shamu:-El100% (5)
- Diia Specification: Dali Part 252 - Energy ReportingDocument15 pagesDiia Specification: Dali Part 252 - Energy Reportingtufta tuftaNo ratings yet
- Internship ReportDocument18 pagesInternship ReportRathan Kumar SMNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)