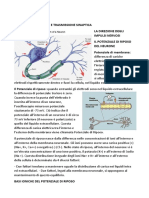
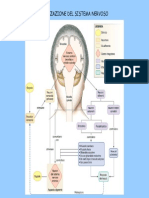
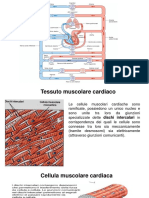
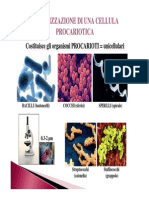
You might also like
- Test Di Fisiologia 1Document47 pagesTest Di Fisiologia 1Matteo Depetris100% (1)
- Fisiologia - Riassunti Kandel "Principi Di Neuroscienze" Cap. 6-15Document32 pagesFisiologia - Riassunti Kandel "Principi Di Neuroscienze" Cap. 6-15Matteo Paolucci100% (15)
- Quiz Neuroanatomia1Document5 pagesQuiz Neuroanatomia1api-3704348100% (1)
- Neuro Chi Rur GiaDocument19 pagesNeuro Chi Rur Giapedrocca1995No ratings yet
- Anestetici LocaliDocument10 pagesAnestetici LocaliChiaraPetraroliNo ratings yet
- Cap Malattie Vascolari Della Retina X58288allp1Document60 pagesCap Malattie Vascolari Della Retina X58288allp1IENEA ANDREEA - CRISTINANo ratings yet
- Potenziale D'azioneDocument8 pagesPotenziale D'azioneMicheleNo ratings yet
- Segnali BioelettriciDocument12 pagesSegnali BioelettriciFrancesco BorealeNo ratings yet
- Potenziale Di MembranaDocument17 pagesPotenziale Di MembranamelissaNo ratings yet
- 2 - BioelettricitàDocument4 pages2 - BioelettricitàFabio BordinNo ratings yet
- 1 Potenziale Di MembranaDocument2 pages1 Potenziale Di MembranaGiacomo Ascari-RaccagniNo ratings yet
- Potenziali D'azioneDocument5 pagesPotenziali D'azioneHghh NgthhNo ratings yet
- Canali Ionici, Fibre NervoseDocument22 pagesCanali Ionici, Fibre Nervoseapi-3704348100% (1)
- Riassuntidi BiofisicaDocument14 pagesRiassuntidi BiofisicaFlavia IovinoNo ratings yet
- CITOLOGIA E ISTOLOGIA - Lezione 7Document2 pagesCITOLOGIA E ISTOLOGIA - Lezione 7Federico TosiniNo ratings yet
- La Pompa Sodio-PotassioDocument19 pagesLa Pompa Sodio-PotassioDavide ValmorbidaNo ratings yet
- Fisiologia 3 - EccitabilitàDocument12 pagesFisiologia 3 - EccitabilitàAnna De Simone100% (2)
- Docsity Domande Triennale Fisiologia UmanaDocument33 pagesDocsity Domande Triennale Fisiologia Umanadestinyema78No ratings yet
- 1 Potenziale Di Equilibrio Di NernstDocument5 pages1 Potenziale Di Equilibrio Di NernstClaudia OrlannoNo ratings yet
- FISIOLOGIAsbobineDocument164 pagesFISIOLOGIAsbobinemarco.ricco.mr00No ratings yet
- CANALI V.D. GENERALE + CANALI AL NaDocument8 pagesCANALI V.D. GENERALE + CANALI AL NaFrancesco BorealeNo ratings yet
- Lezione 10-FisiologiaDocument5 pagesLezione 10-FisiologiaSarah ScuderiNo ratings yet
- Il Potenziale D'azioneDocument6 pagesIl Potenziale D'azioneLeandra EvolaNo ratings yet
- Conduzione NervosaDocument10 pagesConduzione NervosaMiriam LongobardiNo ratings yet
- Appunti Fisiologia IDocument27 pagesAppunti Fisiologia Ikekko88No ratings yet
- Fisiologia PDFDocument75 pagesFisiologia PDFMarioNo ratings yet
- Il Sistema NervosoDocument6 pagesIl Sistema NervosoMartina MusmeciNo ratings yet
- Chimica AnaliticaDocument2 pagesChimica AnaliticaStefano GioiaNo ratings yet
- Appunti FisiologiaDocument30 pagesAppunti FisiologiaWartortle BiancanubbaNo ratings yet
- Trasporto AttivoDocument5 pagesTrasporto Attivogiulia petraccaNo ratings yet
- 1 Edutecnica Semiconduttori 6pDocument6 pages1 Edutecnica Semiconduttori 6pGiovanni AndreoliNo ratings yet
- Anche Ad Altre ComponentiDocument4 pagesAnche Ad Altre Componentifrancesco biancoNo ratings yet
- FisiologiaDocument42 pagesFisiologiaLaura TreggiariNo ratings yet
- Teoria SinapticaDocument5 pagesTeoria SinapticaFran BisoNo ratings yet
- Fisiologia - Fisica Dell'EcgDocument23 pagesFisiologia - Fisica Dell'EcgBenjaminMeghnagi0% (1)
- I Recettori SensorialiDocument37 pagesI Recettori SensorialiMaryna100% (1)
- Dpi Top SecretDocument177 pagesDpi Top SecretratsarcomaNo ratings yet
- Canali Ionici, Enzimi, Molecole CarrierDocument114 pagesCanali Ionici, Enzimi, Molecole Carrierapi-26155364No ratings yet
- Fisiologia 2Document9 pagesFisiologia 2api-3774792No ratings yet
- Appunti Modulo 1-2-3Document84 pagesAppunti Modulo 1-2-3ERIKA BERNOCCHINo ratings yet
- Fisica Del PlasmaDocument10 pagesFisica Del PlasmaWhirl009No ratings yet
- Fisiologia Generale 5Document46 pagesFisiologia Generale 5OrlandoCialliNo ratings yet
- Appunti Di NeurofisiologiaDocument91 pagesAppunti Di NeurofisiologiaSaraNo ratings yet
- Metodi SpettroscopiciDocument23 pagesMetodi SpettroscopicirockerfxNo ratings yet
- NEUROBIOLOGIA La SapienzaDocument30 pagesNEUROBIOLOGIA La Sapienzachiara cimeiNo ratings yet
- Elettromagnetismo PDFDocument112 pagesElettromagnetismo PDFMatteo1980No ratings yet
- Citologia 14 - Potenziale Di MembranaDocument5 pagesCitologia 14 - Potenziale Di MembranaFrancesco TarulloNo ratings yet
- Lezione 04-FisiologiaDocument5 pagesLezione 04-FisiologiaSarah ScuderiNo ratings yet
- Spettroscopia AADocument25 pagesSpettroscopia AAMatteo TreviNo ratings yet
- Potenziale D'azioneDocument30 pagesPotenziale D'azioneRiccardo Mason TroiaNo ratings yet
- Microelettronica Prof D'Alessandro 3Document28 pagesMicroelettronica Prof D'Alessandro 3Giorgio EliseoNo ratings yet
- Proteine Di TrasportoDocument14 pagesProteine Di Trasportorobertosims007No ratings yet
- Trasporti Attraverso La MembranaDocument36 pagesTrasporti Attraverso La MembranaSaraNo ratings yet
- Sistema NervosoDocument15 pagesSistema NervosoBruce WiiNo ratings yet
- Le Giunzioni Nei SemiconduttoriDocument12 pagesLe Giunzioni Nei SemiconduttoripietrNo ratings yet
- FISIOLOGÍADocument26 pagesFISIOLOGÍAAndrea De La FuenteNo ratings yet
- NeurofisiologiaDocument69 pagesNeurofisiologiafabiana900% (1)
- SpettrosDocument34 pagesSpettrosfuliggineNo ratings yet
- Attività Elettrica Del CuoreDocument26 pagesAttività Elettrica Del CuoreStella FilanteNo ratings yet
- 02.cellula Procariotica Cellula Eucariotica PDFDocument40 pages02.cellula Procariotica Cellula Eucariotica PDFDiego Flores EgüezNo ratings yet
- Oboe de GabrielDocument1 pageOboe de GabrielDiego Flores EgüezNo ratings yet
- Vesti La Giubba PDFDocument4 pagesVesti La Giubba PDFDiego Flores EgüezNo ratings yet
- Genetica Medica PDFDocument20 pagesGenetica Medica PDFDiego Flores EgüezNo ratings yet
- PDFDocument3 pagesPDFDiego Flores EgüezNo ratings yet
- 08) Compartimenti Intracellulari PDFDocument27 pages08) Compartimenti Intracellulari PDFDiego Flores EgüezNo ratings yet
- Capitolo5 PDFDocument44 pagesCapitolo5 PDFDiego Flores EgüezNo ratings yet
- Biologia 09-10 - 493-6984742 PDFDocument2 pagesBiologia 09-10 - 493-6984742 PDFDiego Flores EgüezNo ratings yet
- 11 LSS2011PiattiMorfologiaBatterica 42 PDFDocument21 pages11 LSS2011PiattiMorfologiaBatterica 42 PDFDiego Flores EgüezNo ratings yet
- Cellula Glossario PDFDocument2 pagesCellula Glossario PDFDiego Flores EgüezNo ratings yet
- Biologia PDFDocument144 pagesBiologia PDFDiego Flores EgüezNo ratings yet
- BMdC08 2 PDFDocument15 pagesBMdC08 2 PDFDiego Flores EgüezNo ratings yet
- 12 Citoscheletro PDFDocument44 pages12 Citoscheletro PDFDiego Flores EgüezNo ratings yet
- Allegato A Programmi Prove Area SanitariaDocument4 pagesAllegato A Programmi Prove Area SanitariaEla RodriguezNo ratings yet
- Biologia 09-10 - 493-6984742 PDFDocument2 pagesBiologia 09-10 - 493-6984742 PDFDiego Flores EgüezNo ratings yet
- 1364 PDFDocument5 pages1364 PDFDiego Flores EgüezNo ratings yet
- 20130912112644istologia PDFDocument1 page20130912112644istologia PDFDiego Flores EgüezNo ratings yet
- 10.strutture Fondamentali Della Cellula Procariotica (2013) PDFDocument56 pages10.strutture Fondamentali Della Cellula Procariotica (2013) PDFDiego Flores EgüezNo ratings yet
- 1163 - La Membrana Plasmatica PDFDocument8 pages1163 - La Membrana Plasmatica PDFDiego Flores EgüezNo ratings yet
- 2cellula PpsDocument40 pages2cellula PpsDiego Flores EgüezNo ratings yet
- 219 - 16 - Gprotein PDFDocument13 pages219 - 16 - Gprotein PDFDiego Flores EgüezNo ratings yet
- 123 PDFDocument3 pages123 PDFDiego Flores EgüezNo ratings yet
- 10indicatori PDFDocument10 pages10indicatori PDFDiego Flores EgüezNo ratings yet
- Analisi Partitura CiC09 PDFDocument4 pagesAnalisi Partitura CiC09 PDFDiego Flores EgüezNo ratings yet
- Analisis Modelo ESMUCDocument2 pagesAnalisis Modelo ESMUCLuiz Do ValleNo ratings yet
- 1 La Cellula Struttura Master Della Tossicologia PDFDocument28 pages1 La Cellula Struttura Master Della Tossicologia PDFDiego Flores EgüezNo ratings yet
- 03.cellula Eucariotica Virus PDFDocument19 pages03.cellula Eucariotica Virus PDFDiego Flores EgüezNo ratings yet
- Analisi Partitura CiC09 PDFDocument4 pagesAnalisi Partitura CiC09 PDFDiego Flores EgüezNo ratings yet
- Il Glossario Di ChimicaDocument20 pagesIl Glossario Di ChimicaMatteo PeruzzoNo ratings yet
- Esercizi ChimicaDocument4 pagesEsercizi ChimicaMarco DifonzoNo ratings yet
- Catalogo GeneraleDocument88 pagesCatalogo GeneraleLinoSDRNo ratings yet
- DNA Ricombinante, Marcatura Sonda, Elettroforesi, SouthernDocument13 pagesDNA Ricombinante, Marcatura Sonda, Elettroforesi, SouthernemidalbNo ratings yet
- Tavola Conversione Metrico AWGDocument1 pageTavola Conversione Metrico AWGAlessandro InvitiNo ratings yet
- Ecm-1 Fatica IntagliDocument3 pagesEcm-1 Fatica IntagliplutarcoNo ratings yet
- Westex European Brochure ItalianDocument12 pagesWestex European Brochure Italianalejandro orellanaNo ratings yet