You might also like
- Bechamp Was Right All AlongDocument9 pagesBechamp Was Right All AlongYamabushi100% (3)
- Germ Vs Cellular Theory PDFDocument37 pagesGerm Vs Cellular Theory PDFpaulxe100% (3)
- Germ Theory of Disease CausationDocument11 pagesGerm Theory of Disease Causationnashreen83% (6)
- 2017 THE BLUE WAY Visual PDFDocument54 pages2017 THE BLUE WAY Visual PDFAlex KappelNo ratings yet
- Bechamp, Rife and Naessens (The Unhived Mind) PDFDocument4 pagesBechamp, Rife and Naessens (The Unhived Mind) PDFpaulxe100% (1)
- Readers Digest November 2021 PDF RD 2021 PDF EnglishDocument172 pagesReaders Digest November 2021 PDF RD 2021 PDF EnglishIslam Gold100% (1)
- Equine Anesthesia Course NotesDocument15 pagesEquine Anesthesia Course NotesSam Bot100% (1)
- HIV A Virus Like No OtherDocument83 pagesHIV A Virus Like No Otherbadboy6.868510No ratings yet
- AIDS, Cancer and Arthritis: A New Perspective by Phyllis Evelyn Pease (JSE - 21 - 3 - Hodgkinson)Document11 pagesAIDS, Cancer and Arthritis: A New Perspective by Phyllis Evelyn Pease (JSE - 21 - 3 - Hodgkinson)sadunkal100% (1)
- Drug AbuseDocument33 pagesDrug AbuseharshulnmimsNo ratings yet
- Bacteria - Cancer.and - The.origin - Of.life - Part.2 Alan - Cantwell.jr - MDDocument6 pagesBacteria - Cancer.and - The.origin - Of.life - Part.2 Alan - Cantwell.jr - MDSamNo ratings yet
- Subquery ProblemDocument9 pagesSubquery ProblemAbhi RamNo ratings yet
- New Treatments For CancerDocument115 pagesNew Treatments For Cancerfragedu100% (1)
- Immunology Book-Maruthuram CorrectionDocument84 pagesImmunology Book-Maruthuram CorrectionK.VISHNUVARDHAN100% (1)
- History of MicrobiologyDocument5 pagesHistory of MicrobiologyMaloSarkiNo ratings yet
- The Lab-Leak HypothesisDocument32 pagesThe Lab-Leak Hypothesisjavenacosta100% (1)
- Synthesis of The Work of Enderlein, Etc PDFDocument3 pagesSynthesis of The Work of Enderlein, Etc PDFpaulxe100% (1)
- Grain Silo Storage SizesDocument8 pagesGrain Silo Storage SizesTyler HallNo ratings yet
- Historical Development of Medical MicrobiologyDocument84 pagesHistorical Development of Medical MicrobiologyH.K. DL67% (3)
- Open Cholecystectomy ReportDocument7 pagesOpen Cholecystectomy ReportjosephcloudNo ratings yet
- Rabies: Scientific Basis of the Disease and Its ManagementFrom EverandRabies: Scientific Basis of the Disease and Its ManagementRating: 5 out of 5 stars5/5 (1)
- History of MicrobiologyDocument3 pagesHistory of MicrobiologyAngeli ButedNo ratings yet
- Second Thoughts About Viruses, Vaccines, and The HIVAIDS Hypothesis by Robert Young (Young, Robert)Document62 pagesSecond Thoughts About Viruses, Vaccines, and The HIVAIDS Hypothesis by Robert Young (Young, Robert)Maria S100% (1)
- An Introduction to Microbiology for NursesFrom EverandAn Introduction to Microbiology for NursesN. A. SimmonsRating: 3.5 out of 5 stars3.5/5 (4)
- Dr. Oskar Fischer's Mysterious Little Alzheimer's Germ J. of Alzheimer's Disease Editors BlogDocument15 pagesDr. Oskar Fischer's Mysterious Little Alzheimer's Germ J. of Alzheimer's Disease Editors BlogDr. Lawrence Broxmeyer, MD100% (1)
- Ainsworth, The One-Year-Old Task of The Strange SituationDocument20 pagesAinsworth, The One-Year-Old Task of The Strange SituationliliaNo ratings yet
- ResearchDocument16 pagesResearchsk9125770027No ratings yet
- Isitatheoryoralaw? Concept Researched: Overview/Background:: TH THDocument3 pagesIsitatheoryoralaw? Concept Researched: Overview/Background:: TH THAlark SharmaNo ratings yet
- Historia de La Inmunologia Concisa (Ingles)Document15 pagesHistoria de La Inmunologia Concisa (Ingles)AndresPimentelAlvarezNo ratings yet
- History of MicrobiologyDocument6 pagesHistory of MicrobiologySaubie AslamiahNo ratings yet
- Lanado Jan Rey P History of MicrobiologyDocument3 pagesLanado Jan Rey P History of MicrobiologyJAN REY LANADONo ratings yet
- Chapter 1Document20 pagesChapter 1Zoe Alea MartinezNo ratings yet
- Pathogens: A Reconsideration of Koch's Sequence-Based Identification of MicrobialDocument17 pagesPathogens: A Reconsideration of Koch's Sequence-Based Identification of MicrobialGilbert Dela Serna MurilloNo ratings yet
- Centennial Review: The Pathogenesis of TuberculosisDocument6 pagesCentennial Review: The Pathogenesis of TuberculosisAhmad AgielNo ratings yet
- Pharmaceutical MicrobiologyDocument12 pagesPharmaceutical MicrobiologyFarhat KhanNo ratings yet
- Gaston Naessens Somatid Life Cycle 1121Document4 pagesGaston Naessens Somatid Life Cycle 1121Cheshire CatNo ratings yet
- The Concept of MiasmDocument4 pagesThe Concept of MiasmRamesh ShahNo ratings yet
- Award Ceremony SpeechDocument3 pagesAward Ceremony Speechapi-192784092No ratings yet
- Fungi As Human PathogensDocument20 pagesFungi As Human PathogensArdista RaniNo ratings yet
- Helicobacter Pylori and Ulcers:: A Paradigm RevisedDocument8 pagesHelicobacter Pylori and Ulcers:: A Paradigm RevisedNavdeep DhanoaNo ratings yet
- HistoryDocument78 pagesHistorySubrat PandeyNo ratings yet
- Tuberculosis: Disease Research Paper Microbiology - Sat 9:00am BIOL 2164 310 George SofieldDocument9 pagesTuberculosis: Disease Research Paper Microbiology - Sat 9:00am BIOL 2164 310 George SofieldgsofieldNo ratings yet
- History and Story DRAFT ExtraDocument8 pagesHistory and Story DRAFT ExtralouiseNo ratings yet
- Concept of MiasmDocument4 pagesConcept of MiasmCristinaNo ratings yet
- Koch Postulate CoronadoDocument4 pagesKoch Postulate CoronadoCORONADO, Jessa Marie A.No ratings yet
- Is This The Final Solution?Document4 pagesIs This The Final Solution?Orlando GuntherNo ratings yet
- Koch's PostulatesDocument5 pagesKoch's PostulatessaaisNo ratings yet
- Koch Postulate: Why Do We Should Grow Bacteria?: Review ArticleDocument6 pagesKoch Postulate: Why Do We Should Grow Bacteria?: Review Articlesalma sabilaNo ratings yet
- Immunology NotesDocument131 pagesImmunology NotesSuganthiVasanNo ratings yet
- 03.historical Development of MicrobiologyDocument27 pages03.historical Development of MicrobiologyBSNo ratings yet
- Lesson 1 History, Introduction and Scope of ImmunologyDocument5 pagesLesson 1 History, Introduction and Scope of Immunologysomu1No ratings yet
- TOP 26 Significant Events in MicrobiologyDocument5 pagesTOP 26 Significant Events in MicrobiologyRichaella SagaralNo ratings yet
- Germ SummaryDocument4 pagesGerm SummaryKrize Colene dela CruzNo ratings yet
- Unit 3Document19 pagesUnit 3Shruti ThakurNo ratings yet
- History of MicrobiologyDocument63 pagesHistory of MicrobiologyErdem AltunNo ratings yet
- The History, Usage, and Misinformation of Vaccines 1Document20 pagesThe History, Usage, and Misinformation of Vaccines 1api-534716659No ratings yet
- Medical Microbiology, Virology and ImmunologyDocument83 pagesMedical Microbiology, Virology and ImmunologyDora CobuzNo ratings yet
- Tuberculosis and Leprosy in Antiquity An InterpretationDocument12 pagesTuberculosis and Leprosy in Antiquity An InterpretationFredgar22No ratings yet
- The Discovery of Microorganisms: Field of Study Too SmallDocument8 pagesThe Discovery of Microorganisms: Field of Study Too SmallPinky GiriNo ratings yet
- Louis Pasteur, The Father of Immunology?: Kendall A. SmithDocument10 pagesLouis Pasteur, The Father of Immunology?: Kendall A. SmithAbdou MogharbiNo ratings yet
- Leyro JasmineDocument5 pagesLeyro JasmineGokhanScribdNo ratings yet
- The Development-WPS OfficeDocument2 pagesThe Development-WPS OfficeSwati GavaniNo ratings yet
- Plague: From Natural Disease To Bioterrorism: S R, MD, P DDocument9 pagesPlague: From Natural Disease To Bioterrorism: S R, MD, P DJuan Carlos GutiérrezNo ratings yet
- Chapter 1 (1) .PPTX Micro-ParaDocument36 pagesChapter 1 (1) .PPTX Micro-ParaLorelie AsisNo ratings yet
- Anderson. Ecological VisionsDocument24 pagesAnderson. Ecological VisionscaduffNo ratings yet
- MICROBIOLOGYDocument3 pagesMICROBIOLOGYLinaRizaNavalesMonteroNo ratings yet
- MB 0564Document4 pagesMB 0564Bruno GonçalvesNo ratings yet
- Course Syllabus Manufacturing Processes (1) Metal CuttingDocument4 pagesCourse Syllabus Manufacturing Processes (1) Metal CuttingG. Dancer GhNo ratings yet
- Technology For Teaching and Learning 2 OBE SyllabusDocument9 pagesTechnology For Teaching and Learning 2 OBE Syllabusjesreel canalNo ratings yet
- FNCP Improper Waste DisposalDocument2 pagesFNCP Improper Waste DisposalKathleen Daban RagudoNo ratings yet
- Drug Development: New Chemical Entity DevelopmentDocument6 pagesDrug Development: New Chemical Entity DevelopmentDeenNo ratings yet
- Data Sheet FC SIDocument2 pagesData Sheet FC SIAndrea AtzeniNo ratings yet
- Astm A712 PDFDocument3 pagesAstm A712 PDFCristian OtivoNo ratings yet
- VOC & CO - EnglishDocument50 pagesVOC & CO - EnglishAnandKumarPNo ratings yet
- Artesunat Injection 2009Document4 pagesArtesunat Injection 2009Franca ImadiyiNo ratings yet
- 5L ReductionsDocument20 pages5L ReductionsCarlos Javier Orellana OrtizNo ratings yet
- FRM Valuation & Risk Models Dowd, Chapter 2: - Hosted by David Harper Cfa, FRM, Cipm - Published April 22, 2012Document19 pagesFRM Valuation & Risk Models Dowd, Chapter 2: - Hosted by David Harper Cfa, FRM, Cipm - Published April 22, 2012BeastNo ratings yet
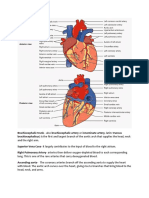
- Brachiocephalic TrunkDocument3 pagesBrachiocephalic TrunkstephNo ratings yet
- Gmail - RedBus Ticket - TN7R20093672Document2 pagesGmail - RedBus Ticket - TN7R20093672Bappa RoyNo ratings yet
- Chan v. ChanDocument2 pagesChan v. ChanjdpajarilloNo ratings yet
- UK FreshTECH Jammer RecipeBook 0Document24 pagesUK FreshTECH Jammer RecipeBook 0Temet NoscheNo ratings yet
- 感應馬達安裝、保養使用說明書31057H402E (英)Document17 pages感應馬達安裝、保養使用說明書31057H402E (英)Rosyad Broe CaporegimeNo ratings yet
- Gloria Pfoltzer Theresa Morris, an Infant, by Mother and Next Friend Christopher Morris, an Infant, by Mother and Next Friend Randy Morris, an Infant, by Mother and Next Friend v. Fairfax County Department of Human Development Susan Manzo Paulette Byrd Florence Hannigan, and Louis Villafane, 966 F.2d 1443, 4th Cir. (1992)Document10 pagesGloria Pfoltzer Theresa Morris, an Infant, by Mother and Next Friend Christopher Morris, an Infant, by Mother and Next Friend Randy Morris, an Infant, by Mother and Next Friend v. Fairfax County Department of Human Development Susan Manzo Paulette Byrd Florence Hannigan, and Louis Villafane, 966 F.2d 1443, 4th Cir. (1992)Scribd Government DocsNo ratings yet
- TFALL CaseStudy-Chandni+Chopra 072020+Document5 pagesTFALL CaseStudy-Chandni+Chopra 072020+Luis Gustavo Heredia VasquezNo ratings yet
- Aircaft Avionics SystemDocument21 pagesAircaft Avionics SystemPavan KumarNo ratings yet
- SAT Subject Chemistry SummaryDocument25 pagesSAT Subject Chemistry SummaryYoonho LeeNo ratings yet
- Global Warmin G and Green House Effect: Submit Ted To:-Mr - Kaush Ik SirDocument24 pagesGlobal Warmin G and Green House Effect: Submit Ted To:-Mr - Kaush Ik SirinderpreetNo ratings yet
- Wago PCB Terminal Blocks and Connectors Catalog 7Document105 pagesWago PCB Terminal Blocks and Connectors Catalog 7alinupNo ratings yet
- Workshop Manual Transporter 2016 19-29Document157 pagesWorkshop Manual Transporter 2016 19-29samueleNo ratings yet