You might also like
- Somatosensory Processes Subserving Perception and ActionDocument51 pagesSomatosensory Processes Subserving Perception and ActionGustavo CabanasNo ratings yet
- Limbic Forebrain - OverviewDocument5 pagesLimbic Forebrain - OverviewHenrique De Figueiredo MariscoNo ratings yet
- Neurobiology RelatedDocument9 pagesNeurobiology RelateddgavrileNo ratings yet
- Cerebellum Ang CognitionDocument14 pagesCerebellum Ang CognitionRenata ZucarinoNo ratings yet
- Three Anti-Correlated Neuronal Networks Managing Brain Activity A ReviewDocument9 pagesThree Anti-Correlated Neuronal Networks Managing Brain Activity A ReviewVioleta-Nella MocanuNo ratings yet
- Cirneci - Three Anti-Correlated Neuronal Networks Managing Brain ActivityDocument8 pagesCirneci - Three Anti-Correlated Neuronal Networks Managing Brain ActivityDanaNo ratings yet
- Religious and Mystical StatesDocument24 pagesReligious and Mystical Statesjosemaria vazquezNo ratings yet
- 54 The Brain From Inside Out 15: Chapter 1. e ProblemDocument20 pages54 The Brain From Inside Out 15: Chapter 1. e ProblemjuannnnNo ratings yet
- 12 - Page - Article - A Proposed Spiritual Axis of The BodymindDocument12 pages12 - Page - Article - A Proposed Spiritual Axis of The Bodymindmuitasorte9No ratings yet
- SASDocument14 pagesSASNicole Ken AgdanaNo ratings yet
- Cingulate Cortex in The Three Limbic SubsystemsDocument13 pagesCingulate Cortex in The Three Limbic SubsystemslooolNo ratings yet
- Cognitive NeuroscienceDocument4 pagesCognitive NeurosciencekhadijaNo ratings yet
- 13 Anatomy of The Nervous SystemDocument78 pages13 Anatomy of The Nervous SystemolivNo ratings yet
- Scientific Basis of Body Mapping and Cortical MapsDocument2 pagesScientific Basis of Body Mapping and Cortical MapsDssalamancazNo ratings yet
- The sense of the physiological condition of the bodyDocument6 pagesThe sense of the physiological condition of the bodyfrompleasureNo ratings yet
- Unit 5 Bio4Er NotesDocument72 pagesUnit 5 Bio4Er NoteskgpunithaNo ratings yet
- Three Anti Correlated Neuronal Networks Managing Brain ActivityDocument8 pagesThree Anti Correlated Neuronal Networks Managing Brain ActivitydgavrileNo ratings yet
- Session #22 SAS - AnaPhy (Lab)Document14 pagesSession #22 SAS - AnaPhy (Lab)G INo ratings yet
- Stochastic Resonance in The Spinal Cord and Somatosensory Cortex of The CatDocument12 pagesStochastic Resonance in The Spinal Cord and Somatosensory Cortex of The CatFrontiersNo ratings yet
- Prescott 2017221412Document23 pagesPrescott 2017221412udanNo ratings yet
- LURIA The Functional Organization of The BrainDocument7 pagesLURIA The Functional Organization of The BrainNatalia HGNo ratings yet
- SNC EEG MalmivuoDocument10 pagesSNC EEG MalmivuoAdam SamenezesNo ratings yet
- Seminar On Concepts of Psychobiology: Kasturba Gandhi Nursing CollegeDocument36 pagesSeminar On Concepts of Psychobiology: Kasturba Gandhi Nursing CollegeNarmadhaNo ratings yet
- Chapter 1 ReadingDocument4 pagesChapter 1 ReadingYousef Ahmad2No ratings yet
- Craig How Do You Feel NowDocument12 pagesCraig How Do You Feel NowKessy ReddyNo ratings yet
- Bushetal 2000 CONTROL COGNITIVO EMOCIONAL PDFDocument8 pagesBushetal 2000 CONTROL COGNITIVO EMOCIONAL PDFtoyetzinNo ratings yet
- Evidence For A Large-Scale Brain SystemDocument14 pagesEvidence For A Large-Scale Brain SystemPoze InstalatiiNo ratings yet
- Brain - Drugs5Document2 pagesBrain - Drugs5swisskomputer3865No ratings yet
- General Organization of Somatosensory SystemDocument16 pagesGeneral Organization of Somatosensory SystemMutaganda Ami fideleNo ratings yet
- Control and RegulationDocument10 pagesControl and RegulationRhoda CatamoraNo ratings yet
- The Central Nervous SystemDocument41 pagesThe Central Nervous Systemhijrah nafeerNo ratings yet
- Emotions in BrainDocument22 pagesEmotions in BrainMensurMrkvićNo ratings yet
- Karolis Etal 2020 BrainLateralizationHumansDocument9 pagesKarolis Etal 2020 BrainLateralizationHumans02chioka44785No ratings yet
- Body OwnershipDocument17 pagesBody OwnershipGaetano CaliendoNo ratings yet
- J Cub 2013 05 041Document10 pagesJ Cub 2013 05 041Eszter SzerencsiNo ratings yet
- FullDocument16 pagesFullЯрослав АнонимNo ratings yet
- MODULE 14 - 15 Psychological Function of Neural Networks and Psychology of PainDocument12 pagesMODULE 14 - 15 Psychological Function of Neural Networks and Psychology of Painartadiel28No ratings yet
- Dum 2016 Cómo El Cerebro Controla El CuerpoDocument6 pagesDum 2016 Cómo El Cerebro Controla El CuerpoJesus GuillenNo ratings yet
- Frontal-Subcortical Circuitry and Behavior PDFDocument11 pagesFrontal-Subcortical Circuitry and Behavior PDFsatyagraha84No ratings yet
- 1 PRECUNEUSselfdanceDocument20 pages1 PRECUNEUSselfdancePaola Andrea GomezNo ratings yet
- Repetition Suppression and Reactivation in Auditory - Verbal Short-Term Recognition MemoryDocument12 pagesRepetition Suppression and Reactivation in Auditory - Verbal Short-Term Recognition Memoryandra_varjogheNo ratings yet
- Luria R., Alexander - Neuropsychology in The Local Diagnosis of Brain DamageDocument17 pagesLuria R., Alexander - Neuropsychology in The Local Diagnosis of Brain DamageNatalia Liseth GAMBA VANEGASNo ratings yet
- Jeff Pressing 1988 Improv-MethodsDocument50 pagesJeff Pressing 1988 Improv-MethodsJoaquín PérezNo ratings yet
- Electrical Chemical Brain StimulationDocument3 pagesElectrical Chemical Brain StimulationNavneet DhimanNo ratings yet
- Churchland - Some Reductive Strategies in Cognitive NeurobiologyDocument31 pagesChurchland - Some Reductive Strategies in Cognitive NeurobiologyPabbyfranNo ratings yet
- CPN 16 018Document14 pagesCPN 16 018Febria Rike ErlianaNo ratings yet
- Functions Neuropshychology IDocument6 pagesFunctions Neuropshychology ICésar ArgumedosNo ratings yet
- Anatomy AssessmentDocument12 pagesAnatomy Assessmentdirshayedilaba24No ratings yet
- Lab 3 CNSDocument8 pagesLab 3 CNSLucky Joy FiguerasNo ratings yet
- Lec2 Cortical AreasDocument11 pagesLec2 Cortical Areasزين العابدين محمد عويشNo ratings yet
- CaapdDocument4 pagesCaapdTimely LianeNo ratings yet
- Pulvermüller and Fadiga (2010) Active PerceptionDocument10 pagesPulvermüller and Fadiga (2010) Active PerceptionMiguel Antonio Barretto GarcíaNo ratings yet
- Frontal Lobe Changes in Alcoholism: A Review of The LiteratureDocument0 pagesFrontal Lobe Changes in Alcoholism: A Review of The Literaturewiwi69No ratings yet
- Oraganisation of The Brain: Studenti (Gr. 13) : - Paun Cristina - Petrescu Andreea Elena - Poenaru Raluca SanzianaDocument10 pagesOraganisation of The Brain: Studenti (Gr. 13) : - Paun Cristina - Petrescu Andreea Elena - Poenaru Raluca SanzianaPaun CristinaNo ratings yet
- Corbetta and Shulman - 2002 - Control of Goal-Directed and Stimulus-Driven AttenDocument15 pagesCorbetta and Shulman - 2002 - Control of Goal-Directed and Stimulus-Driven AttenMario VillenaNo ratings yet
- Anatomy of Neuropsychiatry: The New Anatomy of the Basal Forebrain and Its Implications for Neuropsychiatric IllnessFrom EverandAnatomy of Neuropsychiatry: The New Anatomy of the Basal Forebrain and Its Implications for Neuropsychiatric IllnessNo ratings yet
- Gale Researcher Guide for: Overview of Physiology and NeuropsychologyFrom EverandGale Researcher Guide for: Overview of Physiology and NeuropsychologyNo ratings yet
- Sleep and Brain ActivityFrom EverandSleep and Brain ActivityMarcos G. FrankNo ratings yet
- De Waal & Preston (2017) Mammalian Empathy - Behavioural Manifestations and Neural BasisDocument12 pagesDe Waal & Preston (2017) Mammalian Empathy - Behavioural Manifestations and Neural BasisRenzo Lanfranco100% (2)
- New Measures of Agency From An Adaptive Sensorimotor Task: A1111111111 A1111111111 A1111111111 A1111111111 A1111111111Document13 pagesNew Measures of Agency From An Adaptive Sensorimotor Task: A1111111111 A1111111111 A1111111111 A1111111111 A1111111111Renzo LanfrancoNo ratings yet
- Augusto (2016) Lost in Dissociation. The Main Paradigms in Unconscious Cognition PDFDocument18 pagesAugusto (2016) Lost in Dissociation. The Main Paradigms in Unconscious Cognition PDFRenzo LanfrancoNo ratings yet
- Schizophrenia Risk From Complex Variation of Complement Component 4Document17 pagesSchizophrenia Risk From Complex Variation of Complement Component 4SergioMSaraivaNo ratings yet
- Psychological Testing A Users Guide PDFDocument28 pagesPsychological Testing A Users Guide PDFraginicsekhar7971No ratings yet
- Clinical Assessment of Social Cognitive Function in Neurological DisordersDocument12 pagesClinical Assessment of Social Cognitive Function in Neurological DisordersRenzo LanfrancoNo ratings yet
- The Complete Clinical Assessment in PsychiatryDocument274 pagesThe Complete Clinical Assessment in PsychiatryRenzo Lanfranco100% (4)
- MBCT Booklet Final PDFDocument16 pagesMBCT Booklet Final PDFRenzo Lanfranco67% (3)
- NRN 3776Document14 pagesNRN 3776Renzo LanfrancoNo ratings yet
- Nrurol 2014 193Document8 pagesNrurol 2014 193Renzo LanfrancoNo ratings yet
- Spreading of Pathology in Neurodegenerative Diseases: A Focus On Human StudiesDocument12 pagesSpreading of Pathology in Neurodegenerative Diseases: A Focus On Human StudiesRenzo LanfrancoNo ratings yet
- Blakemore (2012)Document6 pagesBlakemore (2012)Renzo LanfrancoNo ratings yet
- Hypnotic Treatment of Chronic PainDocument30 pagesHypnotic Treatment of Chronic Painelena28_nNo ratings yet
- Kihlstrom (2013)Document10 pagesKihlstrom (2013)Renzo LanfrancoNo ratings yet
- Lieberman (2012)Document5 pagesLieberman (2012)Renzo LanfrancoNo ratings yet
- The Social Brain in Adolescence: Sarah Jayne BlakemoreDocument11 pagesThe Social Brain in Adolescence: Sarah Jayne BlakemoreRenzo LanfrancoNo ratings yet
- Zandian (2007) Cause and Treatment of Anorexia NervosaDocument8 pagesZandian (2007) Cause and Treatment of Anorexia NervosaRenzo LanfrancoNo ratings yet
- Memory Deficits Chapter Provides Guidance on Helping PeopleDocument7 pagesMemory Deficits Chapter Provides Guidance on Helping PeopleRenzo LanfrancoNo ratings yet
- Blakemore & Robbins (2012)Document8 pagesBlakemore & Robbins (2012)Renzo LanfrancoNo ratings yet
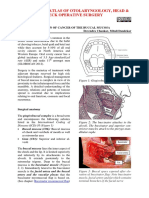
- Surgical Resection of Cancer of The Buccal MucosaDocument21 pagesSurgical Resection of Cancer of The Buccal MucosapradeepNo ratings yet
- Nursing DictionaryDocument353 pagesNursing DictionaryEmmanuel BoadNo ratings yet
- Physical ExaminationDocument117 pagesPhysical Examinationsasmita nayakNo ratings yet
- Tissue Procurement, Processing, and Staining TechniquesDocument10 pagesTissue Procurement, Processing, and Staining TechniquesMuhammad JameelNo ratings yet
- Bioavailabilitas Per Oral: Tradition of Excellence Tradition of ExcellenceDocument63 pagesBioavailabilitas Per Oral: Tradition of Excellence Tradition of ExcellencemiftaNo ratings yet
- Standard and Cardiopulmonary Exercise Testing HandoutDocument17 pagesStandard and Cardiopulmonary Exercise Testing HandoutDarcyNo ratings yet
- Nursing Care PlanDocument5 pagesNursing Care Planruggero07100% (2)
- Lefort 1 OsteotomyDocument16 pagesLefort 1 Osteotomyabhiram.ramesh1290No ratings yet
- NCPDocument10 pagesNCPbabycheska08No ratings yet
- 11.transportation in Plants & AnimalsDocument51 pages11.transportation in Plants & AnimalsReshma ModhiaNo ratings yet
- DR Anu Arasu How Trauma Affects Your HormonesDocument14 pagesDR Anu Arasu How Trauma Affects Your HormonesShalu SaharanNo ratings yet
- Uncontrolled Cell DivisionDocument51 pagesUncontrolled Cell Divisionm43No ratings yet
- Parasitology Lect. (4) : Preparation of Solutions For Wet MountDocument5 pagesParasitology Lect. (4) : Preparation of Solutions For Wet MountAbdul hadiNo ratings yet
- Ent Diseases of The Oral and Pharynx Dr. UyDocument7 pagesEnt Diseases of The Oral and Pharynx Dr. UyAileen EmyNo ratings yet
- Anatomy & Physiology of DengueDocument8 pagesAnatomy & Physiology of DengueYim Yim25% (4)
- Neurology Crossword Puzzle Answer KeyDocument1 pageNeurology Crossword Puzzle Answer KeyRavinaAhujaNo ratings yet
- Functional Hypertrophy: The Secret To Increasing Muscle Without Losing Speed and StrengthDocument2 pagesFunctional Hypertrophy: The Secret To Increasing Muscle Without Losing Speed and Strengthtodd455No ratings yet
- Biochemistry Notes-Study GuideDocument3 pagesBiochemistry Notes-Study Guidegmanb5100% (2)
- Nursing Care Plans for Acute Pain, Hyperthermia, Activity Intolerance, and Infection RiskDocument20 pagesNursing Care Plans for Acute Pain, Hyperthermia, Activity Intolerance, and Infection RiskNursidar Pascual Mukattil80% (5)
- Laboratory Exercise No.8Document3 pagesLaboratory Exercise No.8yumi castroNo ratings yet
- Maxillomandibular Advancement As The Initial Treatment of Obstructive Sleep Apnoea: Is The Mandibular Occlusal Plane The Key?Document9 pagesMaxillomandibular Advancement As The Initial Treatment of Obstructive Sleep Apnoea: Is The Mandibular Occlusal Plane The Key?Jorge Antonio Espinoza YañezNo ratings yet
- Peripheral Testosterone MetabolismDocument8 pagesPeripheral Testosterone MetabolismAgung SentosaNo ratings yet
- 100 Benefits of ExerciseDocument3 pages100 Benefits of ExercisePaul RavenWolf JuniorNo ratings yet
- HemodynamicsDocument115 pagesHemodynamicsناصر دويكاتNo ratings yet
- Understanding 1st, 2nd and 3rd Degree Heart BlocksDocument3 pagesUnderstanding 1st, 2nd and 3rd Degree Heart BlockslhenNo ratings yet
- Mpla 021 Exam May 2017 P1Document4 pagesMpla 021 Exam May 2017 P1Phindile MavimbelaNo ratings yet
- RECALLS - Key points from Preboard 2015 Clinical ChemistryDocument2 pagesRECALLS - Key points from Preboard 2015 Clinical ChemistrySheanalene CastroNo ratings yet
- Inventarisasi Rs Type DDocument63 pagesInventarisasi Rs Type DAnonymous 5wBxtyINo ratings yet
- Autopsy Report: DECEDENT: John AndersonDocument5 pagesAutopsy Report: DECEDENT: John AndersonjilliankadNo ratings yet
- How Ketone Bodies Fuel Brain and HeartDocument3 pagesHow Ketone Bodies Fuel Brain and Heartdaef32No ratings yet