You might also like
- Nutrient Limitation of Bacterioplankton Growth in Lake Dillon, ColoradoDocument14 pagesNutrient Limitation of Bacterioplankton Growth in Lake Dillon, ColoradoBengt HörbergNo ratings yet
- Gutiérrez Et Al 2011 Marine BiologyDocument15 pagesGutiérrez Et Al 2011 Marine BiologyPato CANo ratings yet
- Phytoplankton DiversityDocument3 pagesPhytoplankton DiversitysgphycoNo ratings yet
- tmp64C4 TMPDocument12 pagestmp64C4 TMPFrontiersNo ratings yet
- Nutrient Cycling in A Microflagellate Food Chain: Nitrogen DynamicsDocument12 pagesNutrient Cycling in A Microflagellate Food Chain: Nitrogen DynamicsFardin XNo ratings yet
- tmp7066 TMPDocument2 pagestmp7066 TMPFrontiersNo ratings yet
- MésoplanctonsDocument34 pagesMésoplanctonserraiouiNo ratings yet
- Rhodes 2010Document11 pagesRhodes 2010AnghelloLozanoDíazNo ratings yet
- American Society of Limnology and OceanographyDocument8 pagesAmerican Society of Limnology and Oceanographygourkanti213No ratings yet
- Vik 2020 Genome ResolvedDocument17 pagesVik 2020 Genome ResolvedConsuelo GazituaNo ratings yet
- Sabadini-Santosetal2014 220719 145824Document11 pagesSabadini-Santosetal2014 220719 145824Eduardo Henrique Mendonca MocoNo ratings yet
- Rotifers in Ecotoxicology: A Review: Terry W. Snell L & Colin R - JanssenDocument17 pagesRotifers in Ecotoxicology: A Review: Terry W. Snell L & Colin R - JanssenroraysideblancoNo ratings yet
- 2016 - Guo Et Al. Effects of Antibiotics On The Growth and Physiology of Chlorophytes, Cyanobacteria and A Diatom PDFDocument14 pages2016 - Guo Et Al. Effects of Antibiotics On The Growth and Physiology of Chlorophytes, Cyanobacteria and A Diatom PDFSacra PsyntergiaNo ratings yet
- tmpB514 TMPDocument36 pagestmpB514 TMPFrontiersNo ratings yet
- Bridging The Gap Between Omics and Earth System Science To Better Understand How Environmental Change Impacts Marine MicrobesDocument15 pagesBridging The Gap Between Omics and Earth System Science To Better Understand How Environmental Change Impacts Marine Microbesmamu43-emailNo ratings yet
- Global Change Biology - 2023 - Hodapp - Climate change disrupts core habitats of marine speciesDocument14 pagesGlobal Change Biology - 2023 - Hodapp - Climate change disrupts core habitats of marine speciestonkadujmovic4No ratings yet
- (1997) - (Sakshaug, 1997) - Parameters of PhotosynthesisDocument34 pages(1997) - (Sakshaug, 1997) - Parameters of PhotosynthesisamensetNo ratings yet
- Global Change Biology - 2023 - Kléparski - Phytoplankton Life Strategies Phenological Shifts and Climate Change in TheDocument17 pagesGlobal Change Biology - 2023 - Kléparski - Phytoplankton Life Strategies Phenological Shifts and Climate Change in Thearber.kaziu12No ratings yet
- Viruses and Nutritent Cycles in The Sea, Wilhelm & Suttle, 1999Document8 pagesViruses and Nutritent Cycles in The Sea, Wilhelm & Suttle, 1999deryhermawanNo ratings yet
- Algae/Bacteria Ratio in High-Rate Ponds Used For Waste TreatmentDocument8 pagesAlgae/Bacteria Ratio in High-Rate Ponds Used For Waste Treatment'Hady' HadiyantoNo ratings yet
- Picocyanobacterial Photosynthetic Efficiency Under Daphnia Grazing PressureDocument8 pagesPicocyanobacterial Photosynthetic Efficiency Under Daphnia Grazing PressureNico MartyniukNo ratings yet
- Frontier 2021Document11 pagesFrontier 2021VideoGamesNo ratings yet
- Photosynthesis and Respiration of Baltic Sea BenthDocument19 pagesPhotosynthesis and Respiration of Baltic Sea BenthAyan DuttaNo ratings yet
- When Microbes and Consumers DetermineDocument11 pagesWhen Microbes and Consumers DetermineLivia TanjungNo ratings yet
- 10 MMahmudi EYH EDL NRBDocument7 pages10 MMahmudi EYH EDL NRBWalid al khaledNo ratings yet
- Jurnal PCA IIDocument12 pagesJurnal PCA IIbiotropic biotropicNo ratings yet
- Biofixation of Carbon Dioxide by Spirulina Sp. and Scenedesmus Obliquus Cultivated in A Three-Stage Serial Tubular Photobioreactor PDFDocument7 pagesBiofixation of Carbon Dioxide by Spirulina Sp. and Scenedesmus Obliquus Cultivated in A Three-Stage Serial Tubular Photobioreactor PDFAna Maria AcevedoNo ratings yet
- Studies On Algae Associated With Coca-Cola and Seven-7up Effluents in Kaduna, NigeriaDocument3 pagesStudies On Algae Associated With Coca-Cola and Seven-7up Effluents in Kaduna, NigeriaIOSRjournalNo ratings yet
- 8573 FullDocument8 pages8573 Fullrivas85No ratings yet
- Ismej 2011132 ADocument12 pagesIsmej 2011132 Aamira yaakopNo ratings yet
- Arandia Gorostidi Et Al 2017 Environmental MicrobiologyDocument13 pagesArandia Gorostidi Et Al 2017 Environmental MicrobiologyPepgpNo ratings yet
- 71317-Article Text-153959-1-10-20111027Document8 pages71317-Article Text-153959-1-10-20111027Queenie De GuzmanNo ratings yet
- Rodriguez Pez Cebra 3BP-2015Document5 pagesRodriguez Pez Cebra 3BP-2015Gabriela RodriguezNo ratings yet
- Biogas Productionfrom Algae Biomass Harvestedat Wastewater TreaDocument5 pagesBiogas Productionfrom Algae Biomass Harvestedat Wastewater TreaNitin PatilNo ratings yet
- BatchMixed Culture of C. Vulgaris Using Settled and DilutedDocument8 pagesBatchMixed Culture of C. Vulgaris Using Settled and DilutedGauden Marie C. BangngayNo ratings yet
- Fmicb 13 907703Document9 pagesFmicb 13 907703Veni DarmaNo ratings yet
- Publications Phyto List1Document93 pagesPublications Phyto List1Hendry WijayantiNo ratings yet
- JPP Volume 9 Issue 7 Page 587 592Document6 pagesJPP Volume 9 Issue 7 Page 587 592Royal GreenNo ratings yet
- Heterotrophic Microbes in the OceanDocument34 pagesHeterotrophic Microbes in the OceanCharity MbiziNo ratings yet
- Mathematical Modelling in Fish PondDocument23 pagesMathematical Modelling in Fish PondMarina SchuhNo ratings yet
- pH effects on growth and lipid accumulation of Nannochloropsis salina algaeDocument7 pagespH effects on growth and lipid accumulation of Nannochloropsis salina algaesamagamaNo ratings yet
- Phaeodactylum Tricornutum: Genome Properties of The DiatomDocument10 pagesPhaeodactylum Tricornutum: Genome Properties of The DiatommbrancovNo ratings yet
- Mathematical Modeling of Dissolved Oxygen in Fish Ponds: Full Length Research PaperDocument14 pagesMathematical Modeling of Dissolved Oxygen in Fish Ponds: Full Length Research PaperBayeNo ratings yet
- Andreas 2018 - Microplastic Pollution Increase THGDocument9 pagesAndreas 2018 - Microplastic Pollution Increase THGRosanne RibeiroNo ratings yet
- The Possession of Coccoliths Fails To Deter Microzooplankton GrazersDocument12 pagesThe Possession of Coccoliths Fails To Deter Microzooplankton GrazerscpavloudNo ratings yet
- 12 - Microalgae Population Dynamics in PhotobioreactorsDocument10 pages12 - Microalgae Population Dynamics in PhotobioreactorsAnthony Emilio Roman VegaNo ratings yet
- Dissolved Organic Matter DOM Release by Phytoplankton in The Contemporary and Future OceanDocument28 pagesDissolved Organic Matter DOM Release by Phytoplankton in The Contemporary and Future OceanFabio RicciNo ratings yet
- Ecology Letters - 2009 - Fierer - Global Patterns in Belowground CommunitiesDocument12 pagesEcology Letters - 2009 - Fierer - Global Patterns in Belowground CommunitiesFernanda LageNo ratings yet
- Desmodesmus Sp. Pigment and FAME Profiles Under Different IlluminationsDocument8 pagesDesmodesmus Sp. Pigment and FAME Profiles Under Different IlluminationsHanna HernandezNo ratings yet
- Beta 2Document8 pagesBeta 2Vanessa NogueiraNo ratings yet
- Rolland2016 Article QuorumSensingAndQuorumQuenchinDocument11 pagesRolland2016 Article QuorumSensingAndQuorumQuenchinlunitajoaNo ratings yet
- Natural Product Reports (2014), 31 (11), 1510-1553. ,: Scifinder®Document14 pagesNatural Product Reports (2014), 31 (11), 1510-1553. ,: Scifinder®Nicolas Ignacio Aliaga MachucaNo ratings yet
- Pasquaud Et Al 20081Document13 pagesPasquaud Et Al 20081MyrubicRubicNo ratings yet
- Animating The Carbon Cycle: Accepted FOR Publication INDocument26 pagesAnimating The Carbon Cycle: Accepted FOR Publication INAahad AmeenNo ratings yet
- Spatial Distribution of Phytoplankton in Hurun BayDocument6 pagesSpatial Distribution of Phytoplankton in Hurun BayHandyra Putra AdytamaNo ratings yet
- Effect of Climate ChangeDocument17 pagesEffect of Climate ChangeHaseen AhmedNo ratings yet
- Maximum Photosynthetic Yield of Green Microalgae in PhotobioreactorsDocument11 pagesMaximum Photosynthetic Yield of Green Microalgae in PhotobioreactorsKhalid SiddiquiNo ratings yet
- Physical Chemical Determinant Properties of Biological Communities in Continental Semi Arid WatersDocument15 pagesPhysical Chemical Determinant Properties of Biological Communities in Continental Semi Arid WatersMeri DoroteuNo ratings yet
- Jost (2010) EcologyDocument6 pagesJost (2010) EcologyMeri DoroteuNo ratings yet
- Biomassa Zoo Manca 2000Document4 pagesBiomassa Zoo Manca 2000Meri DoroteuNo ratings yet
- Jost M Is Measuring Biological DiversityDocument4 pagesJost M Is Measuring Biological DiversityMeri DoroteuNo ratings yet
- Extreme Temperature Trends in The Equatorial Region of Brazil CaseDocument12 pagesExtreme Temperature Trends in The Equatorial Region of Brazil CaseMeri DoroteuNo ratings yet
- Rab AlkesDocument65 pagesRab AlkesHery Eko PrasetyoNo ratings yet
- Lecture 7. Developmental GeneticsDocument25 pagesLecture 7. Developmental GeneticsJimmy BeastNo ratings yet
- BookDocument211 pagesBookAngela KocevskaNo ratings yet
- Illusions The Magic Eye of PerceptionDocument12 pagesIllusions The Magic Eye of PerceptionArctic Illusion100% (2)
- Respiratory Muscle Ultrasonography: Methodology, Basic and Advanced Principles and Clinical Applications in ICU and ED Patients-A Narrative ReviewDocument12 pagesRespiratory Muscle Ultrasonography: Methodology, Basic and Advanced Principles and Clinical Applications in ICU and ED Patients-A Narrative Reviewalejandro RodriguezNo ratings yet
- Bacterial Culture Media: Classifying & Preparing Growth EnvironmentsDocument8 pagesBacterial Culture Media: Classifying & Preparing Growth EnvironmentsawadsonNo ratings yet
- Femael Infertility PDFDocument9 pagesFemael Infertility PDFAyadPalaniNo ratings yet
- Practical 14 PDFDocument3 pagesPractical 14 PDFSiddharth KumraNo ratings yet
- Worksheet Central DogmaDocument3 pagesWorksheet Central DogmaRein Jhonnaley Dioso100% (1)
- Lesson 1 Forensic 5Document7 pagesLesson 1 Forensic 5Marcos DmitriNo ratings yet
- Understanding Meningitis: Causes, Symptoms and TreatmentDocument49 pagesUnderstanding Meningitis: Causes, Symptoms and TreatmentDrAbhilash RMNo ratings yet
- 06 - Cell Nucleus and ChromosomesDocument57 pages06 - Cell Nucleus and Chromosomessabina maharaniNo ratings yet
- Primary Brain TumorDocument33 pagesPrimary Brain Tumoriura echin100% (2)
- Muscular System WebquestDocument7 pagesMuscular System WebquestJxcari50% (2)
- 1.introduction To Physiology, The Cell and General PhysiologDocument3 pages1.introduction To Physiology, The Cell and General PhysiologCLEMENT100% (1)
- Causes and prevention of cancerDocument4 pagesCauses and prevention of cancerosamaNo ratings yet
- Handheld Echocardiography: in DepthDocument11 pagesHandheld Echocardiography: in DepthDokter Annisa SppdNo ratings yet
- Occult Phenomena in The Light of TheologyDocument322 pagesOccult Phenomena in The Light of TheologyJulian West100% (2)
- Part of Brain 2Document2 pagesPart of Brain 2PonCut Teuku AchyarNo ratings yet
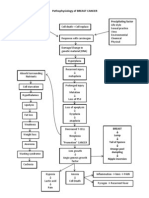
- Pathophysiology of BREAST CANCERDocument1 pagePathophysiology of BREAST CANCERAlinor Abubacar100% (6)
- Asthma Care Quick Reference - Diagnosing and Managing AsthmaDocument18 pagesAsthma Care Quick Reference - Diagnosing and Managing AsthmaMarizka Putri AftriaNo ratings yet
- F4D 24Document2 pagesF4D 24Ivan WongNo ratings yet
- MIDTERM Pilar College Anatomy Physiology Final EditDocument41 pagesMIDTERM Pilar College Anatomy Physiology Final EditYeona BaeNo ratings yet
- MKFP Mat-Xs Clinical 16052012 enDocument8 pagesMKFP Mat-Xs Clinical 16052012 ensuharyantoNo ratings yet
- Full PDFDocument399 pagesFull PDFTeresa Marie Yap CorderoNo ratings yet
- Engebretsen2011, INSULINA EN B BLOQDocument7 pagesEngebretsen2011, INSULINA EN B BLOQFabiola Delia Rosas AlarcónNo ratings yet
- Biology 2000 Paper I Marking SchemeDocument4 pagesBiology 2000 Paper I Marking Schemeapi-2642329075% (4)
- FixationDocument3 pagesFixationficatNo ratings yet
- Cell Signaling Webquest: Part 1: Dropping SignalsDocument3 pagesCell Signaling Webquest: Part 1: Dropping Signalsa60ONo ratings yet
- Renal Arteries PDFDocument18 pagesRenal Arteries PDFServo LedNo ratings yet