Professional Documents
Culture Documents
3 - Catálisis Homogénea UT 1
Uploaded by
Cristina ClavijoOriginal Title
Copyright
Available Formats
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentCopyright:
Available Formats
3 - Catálisis Homogénea UT 1
Uploaded by
Cristina ClavijoCopyright:
Available Formats
F.C.A.I. U.N.
CUYO
Catlisis Homognea 1
FACULTAD DE CIENCIAS APLICADAS A LA INDUSTRIA
UNIVERSIDAD NACIONAL DE CUYO
Carrera: Ingeniera Qumica con orientaciones: Petroqumica y Mineralurgia.
CTEDRA: CATLISIS
Unidad Temtica N 1 - REACCIONES CATALTICAS CATLISIS HOMOGNEA
1.1 Fundamentos de catlisis. Catalizadores. Naturaleza y mecanismos.
1.2 Catlisis enzimtica: Generalidades. Catlisis enzimtica. Inhibidores de enzimas
1.3 Catlisis homognea. Catlisis en fase gaseosa.
1.4 Catlisis en fase lquida: Esterificacin e hidrlisis de steres..
1.5 Catlisis cido base: Generalidades. Esterificacin de cidos y alcoholes.
Halogenacin en grupos carbonilos. Saponificacin de steres..
1.6 Polimerizacin de olefinas. Inversin de azcares. Mutarrotacin de la glucosa.
Disociacin del perxido de hidrgeno.
F.C.A.I. U.N. CUYO
Catlisis Homognea 2
Catlisis
De acuerdo con las condiciones en las que se llevan a cabo las reacciones es posible
separar el fenmeno cataltico en tres dominios independientes.
a) Catlisis homognea: Donde todas las especies cinticamente activas,
comprendido el catalizador, constituyen una misma fase, con una velocidad de reaccin
similar en todos los puntos. Se considera tambin en esta rama el caso en que uno de los
reactivos es un gas y que los otros, con el catalizador, pertenecen a una misma fase lquida.
Debido a la solubilidad del gas la transformacin se produce en todo el lquido y no en la
interfaces gas-liquido. La naturaleza de los productos tampoco influye. En este tipo de
catlisis las velocidades son generalmente elevadas, los venenos inofensivos y la
posibilidad de estudio de mecanismos de reaccin ms fcil para poder aislar las especies
intermedias.
El catalizador es una substancia que influye sobre la velocidad de reaccin y puede
alterarse o no durante el proceso. En el caso de que se altere, la substancia se considera
como catalizador solamente si no existe relacin estequiomtrica total entre su peso y el
peso de la substancia transformada. Muchas veces, an las trazas de catalizador son
extremadamente eficaces. La relacin de utilidad, o nmero de molculas convertidas por
molcula de catalizador, puede ser del orden de millones.
En la catlisis homognea se tiene un acceso ms fcil al mecanismo de reaccin y por
consecuencia se puede dominar mejor el proceso cataltico correspondiente. Otra ventaja no
menos despreciable de este tipo de catlisis es la ausencia de efectos de envenenamiento
tan frecuentes en el caso de la catlisis heterognea, y que obliga a tratamientos costosos
de eliminacin de impurezas.
Uno de los inconvenientes de la catlisis homognea es la dificultad de separar el
catalizador del medio reaccionante, lo que presenta un mayor costo que el de los procesos
heterogneos convencionales. En el cuadro siguiente realizamos una comparacin:
b) Catlisis heterognea: El catalizador es insoluble en los sistemas qumicos en los
cuales provoca la transformacin y forma una fase distinta muy a menudo slida. Existen
dos fases y una superficie de contacto. La reaccin se lleva a cabo en esta superficie de
contacto y el fluido es una reserva de molculas por transformar o que ya reaccionaron.
Como la reaccin qumica se pasa en dos dimensiones, al menos uno de los reactivos
debe ser adsorbido qumicamente. La catlisis heterognea est limitada al estudio de
reacciones provocadas en las molculas por el campo de fuerza del slido y se limita a
algunos ngstrom. Debe hacerse notar que la mayor parte de catalizadores slidos son
metales, xidos, sulfuros metlicos o sales (sulfatos silicato, fosfatos) con alta energa
reticular.
c) Catlisis enzimtica: Que recibe su nombre del catalizador, que es una mezcla o
molcula orgnica que generalmente contiene una protena que forma un coloide lioflico.
F.C.A.I. U.N. CUYO
Catlisis Homognea 3
Dada la naturaleza particular del catalizador, la catlisis enzimtica no pertenece clara y
definitivamente al dominio de la catlisis homognea. Est caracterizada por selectividad
muy elevadas y bajas temperaturas.
Se puede afirmar con base, que sin la catlisis enzimtica no sera posible la vida. Es
suficiente decir que el proceso base de la actividad vital, la asimilacin del CO
2
por la
clorofila de las plantas es un proceso fotoqumico y cataltico. La transformacin por las
clulas, de albminas, grasas carbohidratos as como la sntesis de otras molculas son
catalticas. La formacin de las cadenas de RNA, que es la base del cdigo gentico
depende de la presencia de ciertas enzimas.
1. Catlisis Enzimtica
La vida depende de la existencia de unos catalizadores muy potentes y altamente
especficos denominados enzimas. De hecho, todos los procesos metablicos estn
catalizados por enzimas. La tecnologa enzimtica tiene como objetivo la superacin de
todos aquellos inconvenientes que parecen retrasar la aplicacin de las enzimas en estos
procesos a escala industrial, las enzimas son protenas cuya funcin biolgica es catalizar
las reacciones que suceden en las clulas. Esta rea tiene aplicaciones desde tiempos
remotos como la fermentacin, actualmente en diferentes industrias a diferentes niveles, ya
que implica la utilizacin de sistemas enzimticos.
La funcin principal de las enzimas es actuar como catalizadores de las reacciones de
los seres vivos. Todas las reacciones qumicas tienen lugar porque cierta fraccin de la
poblacin de molculas reactantes, poseen la suficiente energa como para alcanzar un
estado activado, llamado estado de transicin, en el que es muy elevada la probabilidad de
que se rompan o se establezcan enlaces para formar los productos.
Existen dos mtodos generales, mediante los cuales puede acelerarse la velocidad de
una reaccin qumica. Uno de ellos es la elevacin de la temperatura (ya que provoca un
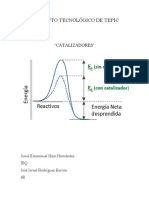
incremento en la velocidad de las molculas); el otro consiste en utilizar un catalizador. El
catalizador se combina con los reactivos de modo transitorio activndolos, produciendo un
estado de transicin de menor energa que en la reaccin no catalizada. Una vez formados
los productos el catalizador queda libre, y puede ser de nuevo utilizado.
Una caracterstica comn de las enzimas es su alto poder cataltico que se debe en
parte a su alta especificidad por los sustratos, la cul puede ser absoluta para un nico
sustrato, o relativa cuando permite la reaccin de compuestos diferentes pero con una
estructura similar.
La especificidad de las enzimas es muy importante para los seres vivos. Cada clula
contiene varios cientos de miles de compuestos diferentes, y existen muchas combinaciones
posibles entre las reacciones qumicas que estos compuestos pueden experimentar. Las
enzimas cuidan de que tengan lugar, de manera especfica, aquellas reacciones que son
esenciales e indispensables para que la clula viva.
Adems de la especificidad y la alta eficiencia, otras dos caractersticas que diferencian
a las enzimas de los catalizadores qumicos, son que: las enzimas pueden saturarse por
sustrato y tienen capacidad para regular su actividad.
Caractersticas de las Enzimas
Las enzimas presentan una serie de caractersticas notables como las siguientes:
- Son protenas que catalizan al reducir la barrera energtica de las reacciones qumicas.
- Influyen solo en la velocidad de reaccin sin alterar el estado de equilibrio.
- Actan en pequeas cantidades.
- Forman un complejo reversible con el sustrato.
- No se consumen en la reaccin, pudiendo actuar una y otra ves.
- Muestran especificidad por el sustrato.
- Su produccin est directamente controlada por genes.
- Son solubles en agua y en alcoholes
- Son precipitadas por sulfato amnico y alcohol concentrado
F.C.A.I. U.N. CUYO
Catlisis Homognea 4
- Tienen carcter coloidal
- Son fcilmente absorbidos
- Son destruidos a 100 C y anulada su accin a 0 C. su mayor actividad se encuentra
entre 37 y 45 C
- El pH influye notablemente en la accin de stas, en general cada una tendr un rango
de pH en el cual se encuentra activa
La actividad cataltica de las enzimas es mucho mayor que los catalizadores
inorgnicos, por lo que podemos decir, que un enzima reduce ms eficientemente la energa
de activacin (Ea) de una reaccin que un catalizador inorgnico, lo que permite que una
reaccin se realice a menor temperatura:
Reaccin H2 O2 H2 O + O2 Ea (Kcal/mol)
a) El perxido de hidrgeno se descompone en: 18
b) El hierro cataltico (Fe) realiza la reaccin: 13
c) El platino (Pt) cataliza la reaccin : 12
d) La catalasa una enzima heptica la realiza: 5
Nomenclatura y Clasificacin de las Enzimas
Muchas enzimas han sido designadas aadiendo el sufijo -asa al nombre del sustrato,
es decir, la molcula sobre la cul ejerce su actividad cataltica. Por ejemplo la ureasa
cataliza la hidrlisis de la urea, y la arginasa cataliza la hidrlisis de la arginina (a urea y
ornitina). Otras enzimas han recibido su nombre en funcin del tipo de reaccin que
catalizan, as la Gliceraldehdo-3P-deshidrogenasa cataliza la oxidacin del la
Gliceraldehdo-3P.
Sin embargo con el descubrimiento de nuevas enzimas esta nomenclatura resulta a
veces confusa. Se subdividen de acuerdo al grupo funcional sobre el que actan en:
Los nombres propuestos en este sistema son los que se emplean cuando es necesaria
una identificacin exacta de las enzimas.
xido Reductasas (reacciones de oxido- reduccin)
Actan sobre ": CH OH "
Actan sobre ": C = O "
Actan sobre ": C = CH "
Actan sobre ": CH NH
2
"
Actan sobre ": CH NH "
Hidrolasas (reacciones de hidrlisis)
Esteres
Enlaces glucosdico
Enlaces pepsdicos
Otros enlaces C N
Anhdridos de cido
Transferasas (transferencia de grupos funcionales)
Grupos de un tomo de C
Grupos aldehdos o cetnicos
Grupos acilos
Grupos glucosilos
Grupos fosfatos
Grupos que contienen azufre
Liasas (Adicin a los dobles enlaces)
: C = C :
: C = O
: C = N
Isomerasas (reaccin de isomerizacin)
Racemasas
Ligasas (Formacin de enlaces con escisin de ATP)
: C O
: C N
: C S
: C C
I. Enzimas Hidrolticas:
Esta clase de enzimas actan normalmente sobre las grandes molculas del
protoplasma, como son la de glicgeno, las grasas y las protenas. La accin cataltica se
expresa en la escisin de los enlaces entre tomos de carbono y nitrgeno (C-N) o carbono
oxigeno (C-O); Simultneamente se obtiene la hidrlisis de una molcula de agua. El
hidrgeno y el oxidrilo resultantes de la hidrlisis se unen respectivamente a las dos
F.C.A.I. U.N. CUYO
Catlisis Homognea 5
molculas obtenidas por la ruptura de los mencionados enlaces. La clasificacin de estas
enzimas se realiza en funcin del tipo de enlace qumico sobre el que actan. A este grupo
pertenecen protenas muy conocidas: la pepsina, presente en el jugo gstrico, y la tripsina y
la quimiotripsina, segregada por el pncreas. Desempean un papel esencial en los
procesos digestivos, puesto que hidrolizan enlaces ppticos, estricos y glucosdicos.
- Carbohidrasas: hidrolizan los hidratos de carbono
Amilasa: almidn maltasa
Sacarasa: sacarosa glucosa y fructosa
Lactasa: lactosa glucosa y galactosa
Maltasa: maltosa glucosa
- Estearasa: hidrolizan los steres en cido y el alcohol correspondiente
Lipasa: grasas cidos grasos y glicerina
Fosfotasas: fosfatos orgnicos cido fosfrico
Clorofilazas: clorofila fitol
- Azolesterasas: hidroliza los steres de amino-alcoholes
- Proteasas: hidrolizan las protenas en proteasas, peptonas y finalmente polipptidos.
- Peptidasas: hidrolizan peptidos en otros ms sencillos, incluso hasta aminocidos.
- Glucosidasas: hidrolizan glucsidos produciendo azcar
- Fosforilasas: descomponen hidratos de carbono, pro inversamente los forman.
- Nucleasas: hidrolizan cidos nuclicos
- Amidasas: actan sobre los enlaces carbono-hidrgeno
II. Enzimas que provocan la oxidacin u oxidasas:
Son las enzimas relacionadas con las oxidaciones y las reducciones biolgicas que
intervienen de modo fundamental en los procesos de respiracin y fermentacin. Extrayendo
dos tomos de hidrgeno, catalizan las oxidaciones de muchas molculas orgnicas
presentes en el protoplasma; los tomos de hidrgeno tomados del sustrato son cedidos a
algn captor.
En esta clase se encuentran las siguientes subclases principales: Deshidrogenasas y
oxidasas. Son ms de un centenar de enzimas en cuyos sistemas actan como donadores,
alcoholes, oxcidos aldehdos, cetonas, aminocidos y muchos otros compuestos y, como
receptores, las propias coenzimas, citocromos, etc.
- Catalasa: perxido de hidrgeno agua y oxgeno
- Peroxidasas: perxidos orgnico agua y oxgeno
- Deshidrogenasas: oxidan eliminando hidrgeno
- Luciferasas: su nombre se debe a que existe en las lucirnagas y acta sobre la
luciferina para producir luz
- Tirosinasa: acta sobre la tirosina produciendo un pigmento negro conocido como
melanina
III. Transferasas (transfieren grupos funcionales)
Estas enzimas catalizan la transferencia de una parte de la molcula (dadora) a otra
(aceptora). Su clasificacin se basa en la naturaleza qumica del sustrato atacado y en la del
aceptor. Tambin este grupo de enzimas actan sobre los sustratos mas diversos,
transfiriendo grupos metilo, aldehdo, glucosilo, amina, sulfat, sulfrico, etc.
IV. Liasas (reacciones de adicin a los dobles enlaces)
Estas enzimas escinden (raramente construyen) enlaces entre tomos de carbono, o
bien entre carbono y oxigeno, carbono y nitrgeno, y carbono y azufre. Los grupos
separados de las molculas que de sustrato son casi el agua, el anhdrido carbnico, y el
amoniaco. Algunas liasa actan sobre compuestos orgnicos fosforados muy txicos,
escindindolos; otros separan el carbono de numerosos sustratos.
V. Isomerasas (reacciones de isomerizacin)
Transforman ciertas sustancias en otras ismeras, es decir, de idntica formula
emprica pero con distinto desarrollo. Son las enzimas que catalizan diversos tipos de
F.C.A.I. U.N. CUYO
Catlisis Homognea 6
isomerizacin, sea ptica, geomtrica, funcional, de posicin, etc. Se dividen en varias
subclases.
Las racemasas y las epimerasas actan en la racemizacin de los aminocidos y en
la epimerizacin de los azcares. Las primeras son en realidad pares de enzimas
especficas para los dos ismeros y que producen un solo producto comn.
Las isomerasas cis trans modifican la configuracin geomtrica a nivel de una doble
ligadura. Los xidosreductasas intramoleculares catalizan la interconversin de aldosas y
cetosas, oxidando un grupo CHOH y reduciendo al mismo tiempo al C = O vecino; en otros
casos cambian de lugar dobles ligaduras.
Por fin las transferasas intramoleculares (o mutasas) pueden facilitar el traspaso de
grupos acilo, o fosforilo de una parte a otra de la molcula,
Algunas isomerasa actan realizando inversiones muy complejas, como transformar
compuestos aldehdos en compuestos cetona, o viceversa. Estas ultimas desarrollan una
oxidorreduccin dentro de la propia molcula (oxido reductasa intramoleculares) sobre la
que actan, quitando hidrgeno, a algunos grupos y reduciendo otros; actan ampliamente
sobre los aminocidos, los hidroxcidos, hidratos de carbono y sus derivados.
VI. Ligasas (formacin de enlaces con consumo de ATP).
Es un grupo de enzimas que permite la unin de dos molculas, libera la energa
necesaria para llevar a cabo la unin de las primeras. Se trata de un grupo de enzimas muy
importantes y recin conocidas, pues antes se pensaba que este efecto se llevaba a cabo
por la accin conjunta de dos enzimas. A este grupo pertenecen enzimas de gran relevancia
reciente, como los aminocido ARNt ligasas conocidas habitualmente con el nombre de
sintetasas de aminocidosARNt o enzimas activadoras de aminocidos que representan el
primer paso en el proceso biosinttico de las protenas, y que forman uniones C-O; las
cido-tiol ligasas.
VII. Enzimas descarboxilantes:
Enzimas que originan un reagrupamiento de la molcula del substrato
- Zimasa: azcares alcohol y dixido de carbono
- Carboxilasa: c. -cetnicos aldehdos y dixido de carbono
VIII. Enzimas que originan un reagrupamiento de la molcula del substrato
- Mutasas
Cofactores Enzimticos (coenzimas)
La actividad de algunas enzimas depende solamente de su estructura como protena,
mientras que otras necesitan, adems uno o ms compuestos no proteicos, llamados
cofactores. El cofactor puede ser un ion metlico o bien una molcula orgnica, llamada
coenzima, aunque algunos enzimas necesitan de ambos. El cofactor puede estar
fuertemente unido a la protena, recibe entonces el nombre de grupo prosttico, o
dbilmente unido y acta bsicamente como un sustrato especfico de la enzima.
El complejo enzima-cofactor catalticamente activo recibe el nombre de holoenzima.
Cuando se separa el cofactor, la protena restante, que por s misma es inactiva
catalticamente, se designa con el nombre de apoenzima.
Holoenzima = Apoenzima + Cofactor
En el grfico se utiliza como cofactor Co
++
; el Co
++
se
une a la enzima, formando la holoenzima, a la cual
se le une el sustrato.
En las enzimas el cofactor puede actuar como:
- Centro cataltico primario: acta con la principal reaccin
- Grupo puente: para reunir el sustrato y la enzima, (complejo activado)
F.C.A.I. U.N. CUYO
Catlisis Homognea 7
- Agente estabilizante de la actividad enzimtica (conformacin).
Por su parte cada una de las coenzimas catalogadas suele contener en su estructura,
alguna vitamina, o molcula derivada de ella. Las coenzimas actan por lo general como
transportadores intermedios de tomos especficos o de electrones.
Activadores Metlicos
Elemento Enzima Activada
Zn
++
Deshidrogenasas, anhidrasa carbnica, ARN y ADN polimerasas.
Mg
++
Fosfohidrolasas, fosfotransferasas, fosfatasas.
Mn
++
Arginasas, peptidasas, quinasas.
Mo Nitratoreductasa, nitrogenasa.
Fe
2+
, Fe
3+
Citocromos, catalasas, ferredoxina, peroxidasas, nitritoreductasa.
Cu
2+
Citocromo oxidasa, tirosinasa, cido ascrbico oxidasa, plastocianina
Ca
2+
K
+
Piruvato fosfoquinasa, ATPasa.
Co Vitamina B
12
hallada en microorganismos y animales, pero no en plantas.
Importante en la fijacin simbitica de nitrgeno.
Ni Ureasa.
Modelos de Actuacin de las Enzimas
Para explicar la actividad cataltica de las enzimas, se ha propuesto un mecanismo
general, en dos etapas, que se representa en la figura siguiente.
E+S ES E+P
K
1 K
2
K
-1
En la primera etapa, la enzima (E) se une a la molcula de sustrato (S), para formar el
complejo enzima-sustrato (ES). En una segunda etapa, el complejo se fragmenta dando
lugar al producto (P) y a la enzima (E), que vuelve a estar disponible para reaccionar con
otra molcula de sustrato. Por lo general, la molcula de enzima es mucho mayor que la del
sustrato por lo que slo una pequea parte de la enzima est implicada en la formacin del
complejo; esta regin que interacciona con el sustrato y en la que tiene lugar la reaccin, se
denomina sitio activo de la enzima.
El sitio activo es una regin tridimensional especfica con una nica distribucin de los
grupos que posibilitan la unin de la enzima al sustrato (estos grupos no tienen por qu ser
necesariamente consecutivos) y reciben el nombre centros catalticos.
El modelo ms conocido sobre el mecanismos de reaccin de las enzimas, es el de
Fischer, quien propuso que la molcula de sustrato se adapta al centro activo de la enzima
del mismo modo que lo hara una llave al encajar en una cerradura.
No obstante, esta hiptesis tiene ciertas limitaciones:
Si el centro activo posee una estructura prediseada para el sustrato, caso de
que sea reversible el proceso, a la vez debe estar perfectamente diseada para que tambin
encaje el producto de la reaccin.
Tampoco explica bien el fenmeno de la inhibicin enzimtica.
Otra hiptesis ms aceptada actualmente es la de enzima flexible o de ajuste inducido
(modelo de Koshland), que sugiere que el sitio activo no necesita ser una cavidad previa
geomtricamente rgida y preexistente, sino que debe tener una disposicin espacial precisa
y especfica que se induce y adapta al sustrato cuando interacciona con l.
Independientemente del modelo, una vez formado el complejo enzima sustrato, mediante un
mecanismo de distorsin, se activan los enlaces que hay que romper y se aproximan los
grupos que hay que enlazar, favoreciendo la formacin del producto resultante de la
reaccin catalizada y quedando la enzima libre para comenzar de nuevo el proceso
cataltico.
1.1. Cintica Enzimtica.
Los principios generales de la cintica de las reacciones qumicas son aplicables a las
reacciones catalizadas por las enzimas, en los seres vivos. No obstante, estas muestran
(adems del fenmeno de la especificidad, antes comentado) un rasgo caracterstico que no
F.C.A.I. U.N. CUYO
Catlisis Homognea 8
se observa en los catalizadores no enzimticos, se trata de la saturacin por el sustrato,
entendida en trminos de ocupacin de los centros activos de todas las molculas de
enzima.
Estudiar el efecto de la concentracin de sustrato sobre la actividad de una enzima no
es sencillo si pensamos que lgicamente la concentracin del sustrato disminuye segn
avanza la reaccin. Una simplificacin
en los experimentos cinticos consiste
en medir la velocidad inicial (V
o
). Si el
tiempo es suficientemente corto la
disminucin de sustrato ser mnima y
podr considerarse casi constante. La
Figura muestra el efecto de distintas
concentraciones de sustrato sobre la
velocidad inicial de la reaccin
catalizada por un enzima, en la cul se
distinguen tres fases:
- Para una concentracin baja de
sustrato, la velocidad es directamente
proporcional a la concentracin del
sustrato (lineal) y la cintica se denomina de primer orden.
- Para una concentracin alta de sustrato, la velocidad de la reaccin se hace
prcticamente constante e independiente de la concentracin de sustrato, la cintica se
considera de orden cero.
- En la zona intermedia de [S] la velocidad del proceso deja de ser lineal, y a esta zona se
la denomina de cintica mixta.
Este comportamiento es caracterstico de la mayora de las enzimas y fue estudiado por
Michaelis y Menten en 1913. La velocidad de una reaccin catalizada nos indica la cantidad
de sustrato consumido o producto formado por unidad de tiempo, en el Sistema
Internacional se designa por U (unidad de actividad enzimtica) y corresponde a los
moles de sustrato consumido en 1 min, o bien a los moles de producto formado en 1 min.
1 U mol S/min mol P/min
La curva que expresa la relacin entre la concentracin de sustrato y la velocidad inicial
tiene la misma forma para la mayora de las enzimas, se trata de una hiprbola rectangular,
cuya expresin algebraica viene dada por la Ec. Michaelis-Menten.
| |
| | S K
S V
V
m
o
+
=
max
Los trminos V
max
(velocidad mxima) y K
m
(constante de Michaelis) son dos
parmetros cinticos de gran importancia, caractersticos de cada enzima, que pueden
determinarse experimentalmente. La velocidad mxima se obtiene cuando la velocidad de
reaccin se hace independiente de la concentracin de sustrato. Este valor depende de la
cantidad de enzima que tengamos. Km nos indica la concentracin de sustrato a la cul la
velocidad de reaccin es la mitad de la velocidad mxima, este parmetro es independiente
de la concentracin de enzima, y es caracterstico de cada enzima segn el sustrato
utilizado (si tiene varios). K
m
tambin indica la afinidad que posee la enzima por el sustrato,
siendo sta mayor, cuanto menor es K
m
. (Cuanto menor sea K
m
menor ser la cantidad de
sustrato, necesaria para alcanzar la mitad de la velocidad mxima, por lo que mayor ser la
afinidad hacia ese sustrato, o dicho de otra forma, si es poco afn necesitar ms sustrato
para alcanzar ese valor de velocidad).
Esta ecuacin puede deducirse matemticamente haciendo la aproximacin del estado
estacionario, segn esta aproximacin la concentracin del complejo ES es constante en el
estado estacionario y por lo tanto las velocidades de formacin y destruccin del complejo
ES son iguales. Adems asumimos que el paso limitante de la reaccin es el segundo y por
lo tanto la velocidad de la reaccin: V
o
= K
2
|ES|.
F.C.A.I. U.N. CUYO
Catlisis Homognea 9
E+S ES E+P
K
1 K
2
K
-1
1.2. V formacin del complejo ES = V destruccin del complejo ES
K
1
|E||S|= K
-1
|ES| + K
2
|ES|
K
1
|E||S|= (K
-1
+ K
2
)
|ES|
La concentracin de enzima libre ser igual a la concentracin total de enzima menos lo
que est unido al sustrato. |E|=| E
t
|-|ES|, as que K
1
| E
t
||S|- K
1
| ES
||S| = (K
-1
+ K
2
)
|ES|
K
1
| E
t
||S|= (K
-1
+ K
2
+
K
1
|S|)
|ES|
Despejando |ES| | |
| |
| |
| |
| | S
K
K K
S E
S K K K
S E K
ES
t t
+
+
=
+ +
=
1
2 1
1 2 1
1
El trmino formado por las tres constantes cinticas es igual a la constante de Michaelis
m
K
K
K K
=
+
1
2 1
De modo que V
o
= K
2
|ES| puede ser expresado como
| |
| | S K
S E K
V
m
t
o
+
=
2
K
2
| E
t
|, ser precisamente la V cuando toda la enzima est unida al sustrato formando
un complejo ES, o sea la enzima est saturada por el sustrato, es decir la V
max
. K
1
| E
t
|=V
max
.
De modo que
| |
| | S K
S V
V
m
o
+
=
max
A partir de esta ecuacin podemos explicar matemticamente las tres fases de la curva
de Michaelis.
A baja [S| (es decir, si Km >>> [S|) el termino K
m
+[S| podemos aproximarlo a K
m
,
quedando una expresin del tipo:
V = k [S|
Esta es una cintica de primer orden que se caracteriza por una variacin lineal de la V
respecto al tiempo y una variacin exponencial de la [S| respecto al tiempo.
- A altas [S| (es decir, Km<<<<[S|) despreciaramos Km frente a [S|, con lo que V = V
max
(que sera constante), la una cintica de orden cero y habramos alcanzado la saturacin por
sustrato.
- El tramo intermedio, en el que la [S| ~ K
m
corresponde a una cintica de orden mixto y
se justifica directamente, con la ecuacin de Michaelis.
En muchos casos es de vital importancia conocer
estos parmetros cinticos. En principio, podran
obtenerse a partir de la curva de Michaelis Menten,
pero existen mtodos grficos ms fiables que
facilitan el clculo de K
m
y la V
max
, todos ellos se
hacen en base a transformaciones matemticas de la
ecuacin de Michaelis y una de las ms utilizadas es
la representacin de Lineweaver-Burk, tambin
conocida como dobles inversos. En esta
representacin se grafica 1/V frente a 1/S, obtindose
una recta cuya interseccin con el eje X es 1/K
m
y
con el eje Y es 1/V
max
, siendo la pendiente K
m
/V
max
.
F.C.A.I. U.N. CUYO
Catlisis Homognea 10
Efecto del pH y de la Temperatura sobre la Actividad Enzimtica.
El pH no afecta la actividad enzimtica directamente sino que modifica la
concentracin de protones. Los protones adems de alterar la estructura de la enzima y el
substrato, pueden participar tambin en la reaccin como substrato o producto. En esos
casos, la concentracin de protones afecta directamente la velocidad de la reaccin.
Cualquier cambio brusco de pH, sabiendo que las enzimas son protenas, puede alterar
el carcter inico de los grupos amino y carboxilo en la superficie proteica, afectando as las
propiedades catalticas de una enzima. A pH alto o bajo se puede producir la
desnaturalizacin de la enzima y en consecuencia su inactivacin.
Un ejemplo es la fosfatasa cida es ms activa a pH 5.0, mientras que la fosfatasa
alcalina lo es a pH 9.
Muchas enzimas tienen mxima actividad cerca de la neutralidad, en un rango de pH
de 6 a 8 (Pepsina 1,5; Tripsina 7,7; Catalasa 7,6; Arginasa 9,7; Fumarasa 7,8; Ribonucleasa
7,8)
Cuando la temperatura sube la velocidad de reaccin aumenta (un aumento de 10 C
incrementa su velocidad entre 1,5 y 5 veces), pero existe una temperatura mxima a la cual
la protena se desnaturaliza dejando de ser funcional. La mayor parte de los enzimas se
desnaturalizan a unos 50C. La ribonucleasa se desnaturaliza a temperaturas superiores a
los 70C.
Reacciones Multisustrato
En este captulo hemos estudiado reacciones del tipo S.P, un mecanismo
relativamente simple con un slo sustrato que podra ajustarse a reacciones
catalizadas por algunas enzimas (Isomerasas, hidrolasas, algunas liasas) pero es
importante tener en cuenta que la gran mayora de las reacciones son multisustrato
y suelen dar varios productos. Cuando una enzima une dos o ms sustratos y libera
mltiples productos, el orden de los pasos pasa a ser una caracterstica importante
del mecanismo de reaccin. Hay distintos mecanismos que explican este tipo de
reacciones, veamos varios casos en los que se utilicen dos sustratos S1 y S2, y se
obtengan dos productos P1 y P2.
Unin ordenada de los sustratos: en este caso un sustrato debe unirse antes de que
el segundo sustrato pueda unirse.
Unin aleatoria de los sustratos: en este caso cualquiera de los dos sustratos puede
ser el primero en unirse a la enzima.
Mecanismo ping-pong: en este caso primero se une un sustrato, se libera el primer
producto, y a continuacin se une el segundo sustrato para as liberar el segundo
producto.
Enzimas Regulatorias
Una caracterstica que diferencia a las enzimas de los catalizadores convencionales,
es la capacidad para regular su actividad. Una enzima puede estar ms o menos activa en
una clula, es decir, su nivel de actividad puede regularse:
- Nivel de sntesis: Que haya ms o menos molculas de la enzima.
- Nivel de actividad: Que las que haya estn ms o menos activas. Puede llevarse a cabo
por factores extrnsecos a la enzima: pH, T, [S|, [I|. O por factores intrnsecos a la propia
enzima, en este caso hablamos de enzimas reguladoras, enzimas que por su propia
naturaleza tienen mecanismos especiales de regulacin. Estn especializadas en regularse
respondiendo de forma muy sensible a seales externas.
En la clula las enzimas trabajan en grupo, en rutas secuenciales para llevar a cabo un
determinado proceso metablico. En cada ruta metablica hay al menos una enzima
reguladora que marca la reaccin global de la ruta. La modulacin de las enzimas
regulatorias puede llevarse a cabo de varias formas:
- Enzimas alostricas. Funcionan mediante la unin reversible no covalente de
compuestos regulatorios llamados moduladores. El modulador puede ser un activador
(modulacin positiva) o un inhibidor (modulacin negativo) y ser el propio sustrato de la
F.C.A.I. U.N. CUYO
Catlisis Homognea 11
reaccin (alosterismo homotrpico) o ser otra sustancia (alsoterismos heterotrpicos).
Normalmente se trata de enzimas multimricas y normalmente el sito activo y el regulatorio
se encuentra en distintas subunidades.
- Enzimas interconvertibles Por modificacin covalente reversible, como por ejemplo por
fosforilacin/defosforilacin o modificacin redox.
- Mediante protenas reguladoras que se unen a la enzima
- Mediante la eliminacin proteoltica de pequeos segmentos peptdicos (esto es
irreversible).
1.3. Inhibidores enzimticos
Existen sustancias que pueden impedir que la enzima desarrolle su actividad cataltica,
rebajando o paralizando la reaccin enzimtica. A estas sustancias se las denomina
inhibidores enzimticos. Teniendo en cuenta que prcticamente todos los procesos celulares
estn catalizados por una enzima, no es de extraar la importancia de muchos inhibidores
enzimticos que actan como frmacos, antibiticos o conservantes, otros pueden ser
txicos e incluso potentes venenos. Por ejemplo la aspirina (acetilsaliclico) inhibe la enzima
que cataliza el primer paso en la sntesis de prostaglandinas, que estn implicados en
procesos como la produccin del dolor.
Hay dos grandes tipos de inhibicin: la reversible y la irreversible. La primera implica
una unin no covalente del inhibidor y por lo tanto siempre puede revertirse. En el caso
irreversible se une de forma covalente a la enzima y la incapacita realmente.
Inhibicin Reversible
Los distintos modelos de inhibicin reversibles implican todos la unin no covalente
del inhibidor con la enzima, pero difieren en los mecanismos por medio de los cuales
reducen la actividad enzimtica y en la forma en que afectan a la cintica de la reaccin.
Entre ellos est la inhibicin competitiva, la acompetitiva y la no competitiva.
- Inhibicin Competitiva
Es una sustancia similar en estructura al sustrato que compite con ste por el sitio
activo de la enzima.
E+S ES E+P
K
1 K
2
K
-1
E+I EI
K
i
Este tipo de inhibicin puede reducirse si se aumenta la concentracin de substrato en
el medio de reaccin.
El succinato deshidrogenasa cataliza la reaccin redox par la que se eliminan dos
tomos de hidrgeno de los tomos de carbono metilnico del succinato:
F.C.A.I. U.N. CUYO
Catlisis Homognea 12
Dos inhibidores competitivos del succinato deshidrogenasa:
- Oxalacetato: COO
-
- CO - CH
3
- COO
-
- Malonato: COO
-
- CH
2
- COO
-
Supongamos la siguiente etapa reversible:
donde I represente el inhibidor y E.I el complejo enzima-inhibidor. La constante de
disociacin de E.I la denominamos K
1
, de tal manera que: | | | || |
1
/ . . K I E I E =
En presencia de inhibidor la ecuacin | | | | | | S E E E .
0
+ = se transforma en:
| | | | | | | | | | | | | | { }
1 0
/ 1 . . . K I E S E I E S E E E + + = + + =
La ecuacin de Estado estacionario para E.S no se altera por la presencia del
inhibidor por lo que an se puede escribir: | | | | | | S K S E E
M
/ . . =
Sustituyendo esta expresin en la ecuacin anterior y despejando:
| |
| |
| |
| | S
K
K
I
E
S E
M
1 1
.
1
0
)
`
+ +
=
Ahora que tenemos una expresin para la concentracin de E.S en funcin de la
concentracin de sustrato, inhibidor y de la enzima inicial, junto con K
M
y K
1
, podemos
obtener una expresin muy til para la velocidad de la reaccin catalizada por una enzima
inhibida.
| |
| | | |
| |
| |
)
`
+ +
= = =
1
0 2
2
1 .
. .
. .
K
I
K S
S E k
S E k v velocidad
M
El valor de la velocidad proporcionado por esta ecuacin ser siempre inferior al de la
ecuacin para la reaccin no inhibida, pero ambas tienen el mismo lmite superior a
concentraciones altas de sustratos. Si la concentracin de I es igual a K
1
, y la concentracin
in de sustrato es igual a K
M
, la velocidad se hace un tercio del valor mximo posible.
Para medir la constante de equilibrio K
1
, se obtienen datos de la velocidad inicial de la
reaccin catalizada por la enzima en funcin de la concentracin de sustrato, a
concentracin de inhibidor constante.
Invirtiendo la ecuacin se obtiene.
F.C.A.I. U.N. CUYO
Catlisis Homognea 13
| | | | | |
| |
)
`
+ + =
1 0 2 0 2
1
. .
.
.
1 1
K
I
S E k
K
E k v
M
Esta ecuacin indica que, a concentracin de
inhibidor constante v
-1
es una funcin lineal de
| |
1
S y la relacin entre la pendiente y la
ordenada en el origen nos proporciona
| |
)
`
+
1
1
K
I
K
M
La relacin entre este grafico y el de la
reaccin no inhibida se ilustra en la figura que
muestra como ambas reacciones tienen la misma
ordenada al origen.
La recta tendr la misma v pero la pendiente
ser ms grande porque K
M
es mayor.
Nuestro modelo de una molcula que se
acompleja con una enzima pero que no reacciona con ella, puede utilizarse para justificar la
estereoselectividad de la accin enzimtica. En muchos casos el enantimero equivocado
se une al centro activo de la enzima con la misma intensidad que el enatimero correcto,
pero al tener la estereoqumica equivocada, la enzima es incapaz de actuar sobre el. As el
ster etlico de D-tirosina se acompleja con quimotripsina con la misma intensidad que con
L-tirosina, pero no tiene lugar reaccin alguna. En este caso, el enantimero equivocado se
comporta como un inhibidor de la enzima.
- Inhibicin A-competitiva
Reacciona con la enzima en un punto
distinto al centro activo, pero slo en el caso de
que sta est unida al sustrato formando el
complejo ES, e impide que la enzima desarrolle
su actividad cataltica.
E+S ES E+P
K
1 K
2
K
-1
ES+I EIS
K
i
Tanto la V
max
como K
m
se alteran en la
misma proporcin, lo que se manifiesta en la
representacin de dobles inversos como rectas
F.C.A.I. U.N. CUYO
Catlisis Homognea 14
paralelas y por lo tanto con la misma pendiente.
El resultado es una recta nueva que se corta en un valor ms grande de ordenadas y
de igual pendiente. Si aumentamos la concentracin de inhibidor obtenemos una familia de
rectas con iguales pendientes y cortes en ordenadas ms grandes.
Como se puede apreciar se produce aparentemente una disminucin de la V
max
y un
descenso de K
m
.
- Inhibicin No Competitiva
Puede combinarse tanto con la enzima libre como con el complejo enzima-sustrato, sin
afectar al sitio activo de la enzima.
Al no unirse al sitio activo, no se afecta la
afinidad de la enzima por el sustrato y en este caso
K
m
no se altera y la V
max
disminuye, como puede
observarse en la representacin de dobles inversos.
- Inhibicin Mixta
Intermedia entre a-competitiva y competitiva, el inhibidor (que no tiene porque
parecerse al sustrato) no se une al centro activo aunque tiene efecto de competitivo. La
unin de uno y otro no son excluyentes, el resultado final depende de cual de los dos
prevalezca.
1/V
1/[S]
0
Sin I
Con I
- 1/Km
1/V
max
1/V
max
Inhibicin no competitiva
1/V
1/[S]
0
Sin I
Con I
- 1/Km
1/V
max
1/V
max
Inhibicin no competitiva
F.C.A.I. U.N. CUYO
Catlisis Homognea 15
Inhibicin Irreversible
El inhibidor se une covalentemente a la enzima y la inactiva de manera reversible.
Casi todos los inhibidores irreversibles son sustancias txicas naturales o sintticas. Se trata
de sustancias que reaccionan con algn grupo funcional importante para la catlisis,
bloquendolo e impidiendo que la enzima desarrolle su actividad. En muchos casos la
interaccin se produce a travs del sitio activo, impidiendo de manera irreversible que el
sustrato ocupe su lugar, tal es el caso del gas Sarn, que inhibe irreversiblemente enzimas
implicadas en la transmisin del impulso nervioso y su inhalacin causa parlisis rpida de
las funciones vitales.
En este caso, cuando afecta al sitio activo, se observara una situacin similar a la
inhibicin competitiva, una disminucin de la V max y un aumento aparente de Km, si bien el
proceso sera completamente irreversible por sustrato que no podra desplazar al inhibidor
del sitio activo. La inhibicin enzimtica por modificacin covalente constituye adems una
importante forma de regulacin metablica.
1.4. Aplicaciones
Las aplicaciones industriales tradicionales se refieren a la produccin de una
transformacin til por alguna enzima, bien sea natural o aadida intencionalmente. Entre
las que podemos citar:
- Fermentacin: La fermentacin alcohlica es un ejemplo conocido de los procedimientos
en que se efectan alteraciones enzimticas, tanto cuando se agrega alguna enzima, como
cuando se aade algn microbio vivo (levadura). El empleo de amilasa en forma de malta es
indudablemente la mayor aplicacin industrial que tiene las enzimas, pero no es del todo
conocida la accin de estas amilasas. La elaboracin de vinagre con alcoholes es un
proceso enzmico producido por un microbio vivo (Acetobacter aceti).
- Curticin: La cantidad de material enzimtico que se usa en la industria de curtidura
representa probablemente la mayor aplicacin industrial de las enzimas despus de la
industria de la fermentacin.
- Fabricacin de queso: Los quesos se fabrican coagulando la casena con rennina, esta
probablemente tiene una accin proteoltica muy dbil que contina en el queso. La rennina
produce un cogulo elstico del que se exprime fcilmente el suero. No es la nica
protenasa que se usa en la elaboracin del queso, pues tambin se emplea mezclas de
rennina con pepsina.
- Elaboracin de pan: La harina de trigo contiene tambin pequeas cantidad de -
amilasa y gran proporcin de -amilasa. La adicin de la -amilasa a la harina,
generalmente en forma de harina de trigo malteado, ocasiona aumento de volumen de la
hogaza. La amilasa agregada hidroliza parte del almidn y lo convierte en maltosa, con lo
E+S ES E+P
K
1 K
2
K
-1
E+I EI
F.C.A.I. U.N. CUYO
Catlisis Homognea 16
cual suministra ms azcar para que fermente la levadura y origina la generacin de mayor
cantidad de dixido de carbono.
- Carbohidrasas Descomponen residuos de azcares de carbohidratos superiores.
Descomponen los almidones y el glucgeno en dextrinas y disocian lentamente las dextrinas
en maltosa y cantidad mnima de glucosa. Destruyen la estructura en cadenas ramificada
del almidn y el glucgeno.
- Productos Mdicos y Farmacuticos En la actualidad el nmero concreto de aplicaciones
es relativamente pequeo. Puesto que las aplicaciones mdicas y farmacuticas de las
enzimas abarcan un amplio espectro de materias, es conveniente dividirlas en tres reas
importantes de inters: terapia enzimtica, uso analtico y productos de compuestos
farmaceticos.
Las aplicaciones mdicas y farmacuticas de enzimas requieren generalmente pequeas
cantidades de enzimas muy purificadas. En parte, esto refleja el hecho de que para una
enzima sea efectiva slo debe modificarse el o los compuestos de inters contenido en un
fluido o tejidos fisiolgicos complejos. Adems, si el destino de una enzima o de un producto
obtenido por mtodos enzimticos es su administracin a un paciente, resuelta evidente que
el preparado debe contener las menores cantidades posibles de material extrao para evitar
probables efectos secundarios.
- Produccin de aminocidos enzimticamente: La produccin de aminocidos mediante
tecnologa con enzimas est adaptada convenientemente. Aunque se pueden sintetizar
empleando un proceso qumico, se debe sealar que en este caso se obtiene una mezcla de
D y L ismeros. Este proceso puede llevarse a cabo mediante el empleo de la enzima
aminoacilasa. Estas son dos reas importantes en el desarrollo de esta tecnologa.
- Antibiticos semi-sintticos: Las penicilinas semisintticas son los principales productos
farmacuticos obtenidos por tecnologa enzimtico.
- Alimentos
Las enzimas son piezas esenciales en el funcionamiento de todos los organismos vivos,
actuando como catalizadores de las reacciones de sntesis y degradacin que tienen lugar
en ellos. La utilizacin de enzimas en los alimentos presenta una serie de ventajas, adems
de las de ndole econmica o tecnolgica. La gran especificidad de accin que tienen las
enzimas hace que no se produzcan reacciones laterales imprevistas. Las enzimas pueden
inactivarse fcilmente cuando se considere que ya han realizado su misin, quedando
entonces asimilados al resto de las protenas presentes en el alimento.
Para garantizar la seguridad de su uso deben tenerse en cuenta algunas
consideraciones: Los microorganismos ideales son aquellos que tienen ya una larga
tradicin de uso en los alimentos (levaduras de la industria cervecera, fermentos lcticos,
etc.).
Las enzimas utilizadas dependen de la industria y del tipo de accin que se desee
obtener, siendo ste un campo en franca expansin. A continuacin se mencionan
solamente algunos ejemplos.
1. Industrias lcteas
2. Panadera
3. Cervecera
4. Fabricacin de zumo
5. -Fabricacin de glucosa y fructosa a partir del maz
6. Refinado de azcar
- Detergentes
La tecnologa de enzimas en los detergentes se desarroll a partir de la dcada de los aos
60, como una herramienta ms de stos para atacar ciertos sustratos (generalmente
proticos) especficos. Las ms comunes son las llamadas proteasas, las cuales degradan
restos de protenas; y las lipasas que pueden atacar restos de sustratos lpidos que son los
que comnmente se adhieren a la ropa y a ellas se les adhieren el resto de la suciedad
como polvo, restos de otros compuestos orgnicos etc. Los detergentes que contienen
enzimas se los llama detergentes biolgicos.
- Energa
Otra actividad que llama a las aplicaciones biotecnolgicas es la produccin de energa,
siendo la ventaja de las fuentes orgnicas renovables. Cada ao crecen unas 200 mil
F.C.A.I. U.N. CUYO
Catlisis Homognea 17
millones de toneladas de biomasa (madera, cereales, etc), de las cuales los humanos
usamos slo un 3%. Por lo tanto, este rubro ofrece un enorme potencial que puede ser
aprovechado. Un ejemplo clsico de biocombustible es el alcohol obtenido por fermentacin
de material rico en azcares y almidn, o de residuos orgnicos varios, incluyendo los
forestales. El principal obstculo para la viabilidad de esta propuesta es el costo, puesto que
el petrleo sigue siendo ms barato. Sin embargo, los avances tecnolgicos estn
permitiendo acortar la brecha.
- Tratamiento de desechos
Pero es el tratamiento de desechos donde la biotecnologa puede tener un mayor impacto a
nivel mundial. Bacterias, microalgas, levaduras, hongos y plantas han mostrado una notable
eficiencia para metabolizar residuos orgnicos, y metales pesados reduciendo hasta 20
veces el costo involucrado en la incineracin de dichos residuos.
Fuentes de Enzimas
Las fuentes de enzimas pueden ser de tipo vegetal, animal y microbiana.
Las enzimas de tipo vegetal, se encuentran las proteasas, carbohidrasas; las cuales
descomponen residuos de azcares de ca - -amilasas
Entre las enzimas de tipo animal estn las esterasa y lipasa, que se producen en la
mucosa gstrica, el pncreas, y en las semillas de ricino. Las fosfotasas, se obtiene de
tejidos animales seo, muscular, tripsina y la quimotripsina se produce en el pncreas.
Las enzimas del tipo microbiano provienen de bacterias y hongos.
Produccin de Enzimas a Gran Escala
La produccin de enzimas para empleo industrial y como alimento se ha desarrollado
en forma independiente en diversas industrias. La fuente original de enzimas de cereales,
principalmente de las distintas clases de malta, es la industria de la malta de cebada.
Las proteasas de las plantas, como la papaina, bromalaeina y ficina, que se emplean
en los EE UU son de importacin, y generalmente los importadores tienen poco control
sobre las condiciones del proceso de produccin.
La industria empacadora de carnes es la fuente principal de las enzimas derivadas del
pncreas, estmago e hgado de los animales. Finalmente, las enzimas de fuentes
microbiolgicas: Bacterias, Hongos y levaduras, se producen en la industria de la
fermentacin.
1.5. Catlisis Enzimtica: Ejemplos
- La Quimotripsina
Es una enzima digestiva, cuya funcin es la de promover la hidrlisis de ciertas
uniones peptdicas en las protenas; enrollada de modo que sus partes hidrfobas se dirigen
a su interior, lejos del agua, y que permite la mxima formacin de puentes de hidrgeno
intramoleculares. No solo cataliza la hidrlisis de las protenas sino que tambin la de
amidas y esteres ordinarios.
La quimotripsina acta en dos etapas; en la primera rompe la cadena peptidica,
actuando como un alcohol. Puede reconocerse esta reaccin como una alcohlisis de una
amida sustituida: La sustitucin nucleoflica del acilo.
Los productos son una amina (parte liberada de la molcula de sustrato) y un ster
de la enzima.
Etapa 1
C NH
O
+
E O H
C O
O
E
+
NH2
Protena Enzima Enzima
Acilada
Parte de la
cadena
Protenica
Amida
Alcohol
Ester Amina
En la segunda etapa hidroliza el ster de la enzima, lo que genera un cido Carboxlico (la
otra parte de la molcula del sustrato) y la enzima que se recupera, lista para entrar
nuevamente en funcin.
F.C.A.I. U.N. CUYO
Catlisis Homognea 18
Etapa 2
C O
O
E
Enzima
Acilada
Ester
+
H2O COOH
+
Resto de la
cadena
Protenica
Acido
Carboxilico
E O H
Enzima
Alcohol
pH
V
7,4
La velocidad de la hidrlisis por
catlisis enzimtica cambia con la
variacin de la acidez del medio
reaccionante. Si se construye el grafico de
la velocidad de hidrlisis en funcin del pH
de la solucin, se obtiene una curva en
forma de campana, a medida que
aumenta el pH, la velocidad alcanza un
mximo, para luego decaer. La velocidad
es mxima para un pH de 7,4 y es ms
lenta tanto en solucin ms cida como
ms bsica.
El anlisis de los resultados experimentales sindica lo siguiente:
La hidrlisis requiere de una base libre, de Kb alrededor de
7
10
, y de una base
protonada, de Kb de aproximadamente
5
10 . 3
. A pH bajo, ambas bases estn
protonadas; a pH elevado, ambas estn libres. La hidrlisis alcanza su velocidad mxima
cuando la base ms dbil est mayormente libre y cuando la ms fuerte se encuentra casi
totalmente protonada.
La quimotripsina no es muy especifica en su accin, al contrario de la mayora de las
enzimas, hidroliza igualmente a protenas, pptidos, amidas simples y steres.
- La pepsina:
Como todos sabemos, muchos nutrimentos como los aminocidos, los minerales y
algunas vitaminas tales como el cido flico y la vitamina B
12
dependen de la acidez
adecuada del estmago para que se puedan digerir y absorber.
El cido gstrico realiza estas funciones de la digestin y de la absorcin, al optimizar el
pH del estmago por medio de la propia enzima digestiva estomacal, llamada la pepsina.
Si, por un lado, encontramos muy poco cido, no pueden ocurrir adecuadamente las
reacciones qumicas normales requeridas para desbaratar y preparar a los nutrimentos para
su absorcin.
Por el otro extremo del pH, demasiada acidez
puede destruir a los tejidos del tracto digestivo
contribuyendo a la formacin de lceras.
Hay una enzima llamada pepsina que es
necesaria para la digestin ptima inicial de las
protenas.
Durante la ingestin de la comida, la secrecin
del cido gstrico dispara la produccin de la
pepsina (niveles de cido bajo, tambin niveles
bajos de pepsina). Esto causar que las protenas
no se desbaraten en aminocidos. La deficiencia consecuente de muchos aminocidos
esenciales puede llevarnos a la depresin, el insomnio, la ansiedad y muchas otras
enfermedades peligrosas a largo plazo.
La pepsina del estmago, presenta un pH ptimo a 2
F.C.A.I. U.N. CUYO
Catlisis Homognea 19
- Importancia del ATP (Trifosfato de adenosina)
Es la principal fuente de energa de los seres vivos y que alimenta casi todas las
actividades celulares, entre ellas el movimiento muscular, la sntesis de protenas, la divisin
celular y la transmisin de seales nerviosas.
Esta molcula se encuentra en todos los seres vivos y constituye la fuente principal de
energa utilizable por las clulas para realizar sus actividades. Se origina por el metabolismo
de los alimentos en unos orgnulos especiales de la clula llamados mitocondrias.
El ATP se comporta como una coenzima, ya que su funcin de intercambio de energa y
la funcin cataltica (trabajo de estimulacin) de las enzimas estn ntimamente
relacionadas.
Los dos puentes entre los grupos fosfato son uniones de alta energa, es decir, son
relativamente dbiles y cuando las enzimas los rompen ceden su energa con facilidad. Con
la liberacin del grupo fosfato del final se obtiene siete kilocaloras de energa disponible
para el trabajo y la molcula de ATP se convierte en
ADP (difosfato de adenosina).
La mayora de las reacciones celulares que
consumen energa estn potenciadas por la conversin
de ATP a ADP, incluso la transmisin de las seales
nerviosas, el movimiento de los msculos, la sntesis
de protenas y la divisin de la clula.
Por lo general, el ADP recupera con rapidez la
tercera unidad de fosfato a travs de la reaccin del
citocromo, una protena que se sintetiza utilizando la energa aportada por los alimentos. En
las clulas del msculo y del cerebro de los vertebrados, el exceso de ATP puede unirse a la
creatina, proporcionando un depsito de energa de reserva.
1.6. Conclusiones
Las enzimas son catalizadores de origen biolgico que cumplen muchos requisitos
para impulsar nuevas industrias qumicas.
La tecnologa enzimtica tiene mltiples aplicaciones, como fabricacin de alimentos,
los progresos que estn realizando actualmente la ingeniera gentica y la biotecnologa
permiten augurar el desarrollo cada vez mayor del uso de las enzimas.
La utilizacin de enzimas en los alimentos presentan una serie de ventajas, adems
de las de ndole econmico y tecnolgico.
Las enzimas utilizadas dependen de la industria y del tipo de accin que se desee
obtener.
Las fuentes de enzimas pueden ser de origen vegetal, animal o microbiano, se
pueden manipular genticamente, la biosntesis de enzimas para optimizar los procesos,
pero se debe tener en cuenta, las respectivas normas.
La produccin de enzimas a gran escala tiene su principal aplicacin en la industria de
la fermentacin.
2. Catlisis en Fase Gaseosa
Las catlisis homogneas en fase gaseosa son relativamente raras, pero existen una
amplia variedad de ejemplos:
El xido ntrico es un catalizador industrial para la oxidacin del dixido de azufre. Se ha
demostrado que la reaccin transcurre mucho ms rpidamente por la secuencia
2 NO + O
2
2 NO
2
NO
2
+ SO
2
NO + SO
3
que por combinacin directa. Inicialmente indicaremos que la primera de estas
reacciones es uno de los pocos ejemplos conocidos de una reaccin trimolecular.
F.C.A.I. U.N. CUYO
Catlisis Homognea 20
Uno de los usos ms importantes del xido ntrico es en el proceso para la fabricacin
del cido sulfrico, este mtodo de obtencin en cmaras de plomo es cada vez menos
utilizado debido a su bajo rendimiento, pero es ms econmico que otros. Se produce un
cido sulfrico de 70% de pureza. Este mtodo se puede expresar en dos reacciones en las
que los xidos de NO NO
2
actan de catalizadores
4 2 2 2
2 2 HNOSO NO NO O SO + + +
(cido nitrososulfrico)
2 4 3 2 4
2 2 3 2 NO NO HSO O H O H HNOSO + + + +
+
en disolucin da disolucin de H
2
SO
4
La formacin del cloruro de hidrgeno a partir de sus elementos est catalizada por
vapor de sodio o de potasio. De la accin de un solo tomo metlico resultan miles de
molculas de HCl.
El efecto cataltico positivo del agua en la reaccin entre el monxido de carbono y el
oxgeno ha sido interpretado de acuerdo con el mecanismo siguiente:
CO + H
2
O CO
2
+ H
2
2 H
2
+ O
2
2 H
2
O
Las descomposiciones del acetaldehdo, de varios teres y del xido nitroso estn
fuertemente catalizadas por el yodo en fase gaseosa.
El Yodo vapor tambin cataliza un nmero de
pirolisis orgnicas. Como ejemplo, podemos encontrar la
descomposicin del acetaldehdo catalizada por yodo, la
cual es una reaccin en cadena:
La aplicacin de la aproximacin del estado
estacionario a las especies intermedias conduce a la
ecuacin de velocidad:
| |
| | | | CHO CH I k
dt
CHO CH d
3
2 1
2
3
=
Donde:
2
2 1
1
1
2
k
k
k
k
|
|
.
|
\
|
=
En el proceso de iniciacin de tomos de yodo, en
donde se propaga la cadena mediante la reaccin [2]. Se
ve que los productos principales (CH
4
y CO), son los mismos que en la reaccin sin
catalizar. El catalizador (yodo), se regenera en la reaccin [6].
Dado que el enlace II es mucho ms dbil que el enlace CC del acetaldehdo, la
iniciacin es mucho ms fcil y E1 es 204 KJ/mol (comparado con 332 KJ/mol para la
reaccin no catalizada). Este valor de E1 da una energa de activacin global de 134 KJ/mol
para la descomposicin catalizada comparada con 198 KJ/mol para la descomposicin no
catalizada. La barrera de energa de activacin, ha descendido por tanto en 64 KJ/mol.
Muchas reacciones gaseosas que parecen homogneas, en realidad se efectan en
las paredes del reactor, y, por tanto, corresponde a catlisis heterogneas. As, el agua
presenta en algunos casos un efecto cataltico negativo, que ha sido interpretado como
debido a su adsorcin sobre las paredes del recipiente, lo que equivale a un
envenenamiento del catalizador. Aquellas pocas reacciones que parecen ejemplos de
catlisis homogneas son realmente reacciones en cadena. Las sustancias que inician las
reacciones en cadenas suelen llamarse sensibilizantes con ms propiedad que
catalizadores.
Hay que verificar si los datos experimentales se ajustan a las ecuaciones cinticas
simples, si no lo hacen, es indicio de efectos heterogneos o bien de la existencia de un
mecanismo en cadena.
F.C.A.I. U.N. CUYO
Catlisis Homognea 21
Un ejemplo experimental sencillo de la accin de un catalizador en fase homognea
gaseosa es el de la descomposicin del ter etlico (comnmente ter). En la reaccin sin
catalizador a 700 C, se obtienen los siguientes productos:
CO H C CH H OC H C
k
+ +
4 2 4 5 2 5 2
2
1
2
ter etlico metano + etileno + monxido de carbono
En la reaccin anterior, la Energa de Activacin (barrera energtica para pasar de
reactivos a productos) es de unos 51,8 Kcal/mol.
Ahora la misma reaccin, pero con catalizador (yodo), en ste caso la desaparicin del
ter etlico es 10.000 veces ms rpida, en donde se generan productos diferentes:
CO CH H C H OC H C
k
+ +
4 6 2 5 2 5 2
'
0
En este caso la Energa de Activacin es de 34 Kcal/mol, por lo que la presencia de
yodo cambi la velocidad y la selectividad de la reaccin. El aumento de velocidad se debe
a la disminucin de la Energa de Activacin. Como las dos reacciones se efectuaron en
condiciones similares se pueden relacionar sus velocidades a partir de las ecuaciones de
Arrhenius:
r catalizado Velocidad
r catalizado con Velocidad
velocidad de Aumento
sin
=
T R
T R
T R
e
k
k
e k
e k
800 . 17
0
'
0
800 . 51
0
000 . 34
'
0
De donde se observa que el aumento en velocidad depende de k
0
y k'
0
y es
exponencialmente proporcional a la diferencia en energas de activacin. Si se asume que k
0
y k
0
' son iguales, entonces el aumento que debera observarse es de 345.000 veces. Este
no es el caso, ya que slo se observ un aumento de 10.000 veces, por lo tanto hay un
factor 34,5 menor que debe ser atribuido a la diferencia en las constantes k y k'
0
. De
acuerdo a la teora cintica de los gases esas constantes dependen del nmero de choques
entre las molculas de reactivo en el momento de la reaccin; en el caso de la reaccin sin
catalizar es el nmero de choques entre molculas de ter etlico, y en el caso de la
reaccin catalizada es el nmero de choques de las molculas de ter etlico con el iodo.
Como la concentracin de catalizador es muy baja (aproximadamente 1%) el nmero
de choques es menor para la reaccin catalizada, y por eso es que se presenta el factor
34,5 favorable a la reaccin sin catalizador:
'
0 0
5 , 34 k k =
Sin embargo esta disminucin en el nmero de choques efectivos se ve ampliamente
compensada por el abatimiento en la energa de activacin que al encontrarse en el trmino
exponencial conduce a un aumento de 345.000 veces en la velocidad. As el aumento neto
observado por la presencia del catalizador es de 10.000 veces.
En la prctica este aumento de velocidad en presencia del catalizador es aprovechado
para obtener la misma velocidad, o ligeramente superior, pero a temperaturas mucho ms
bajas que las utilizadas en el caso de la reaccin sin catalizador.
Un importante ejemplo de reacciones catalizadas en fase gaseosa son las reacciones en
la atmsfera
Las reacciones ms importantes que ocurren en la atmsfera se refieren
principalmente a la destruccin del ozono, catalizada por NO
x
.
El xido ntrico NO destruye el ozono para formar oxgeno O
2
y dixido de nitrgeno
NO
2
. Por esta razn los niveles de ozono no suelen ser elevados en reas urbanas, donde
hay elevados niveles de NO emitido por los vehculos, a diferencia de lo que sucede en las
zonas rurales.
En la alta atmsfera, en las regiones de poca densidad se absorben las radiaciones
de alta energa, del ultravioleta lejano, procedentes del sol que impide que estos lleguen a la
F.C.A.I. U.N. CUYO
Catlisis Homognea 22
tierra haciendo posible la vida en la superficie terrestre. Estas radiaciones dan lugar a
reacciones qumicas diferentes, como las reacciones de fotodisociacin y de ionizacin.
En las Reacciones de Fotodisociacin el sol emite energa radiante dentro de lmites
muy amplios de longitudes de onda, y tienen suficiente energa para ocasionar cambios
qumicos.
La radiacin electromagntica
se puede representar como un flujo de
fotones. Para que ocurra un cambio
qumico se deben de satisfacer dos
condiciones. Primero, debe haber
fotones con suficiente energa para
llevar a cabo un proceso qumico
determinado. Segundo, las molculas
deben absorber estos fotones.
La ruptura de un enlace qumico
que resulta de la absorcin de un fotn
por una molcula se llama
fotodisociacin. Uno de los procesos
ms importantes que ocurren en la
atmsfera superior, es la
fotodisociacin de la molcula de
oxgeno:
O
2 (g)
+ hv (long< 240nm) 2O
-
(g)
E = h v,
en donde h es la
constante de Plank y
v es la
frecuencia de la radiacin
La segunda condicin que se
debe satisfacer antes de que la
disociacin se lleve a cabo, es que el
fotn debe ser absorbido por O
2.
Afortunadamente el O
2
absorbe gran parte de la radiacin
de alta energa de longitud de onda corta, proveniente del espectro solar, antes de que
llegue a la atmsfera inferior. Al hacerlo se forma el oxgeno atmico.
A grandes altitudes, la disociacin del O
2
es muy importante. A 400 Km., solamente el
1% del oxgeno est en forma de O
2
; el otro 99% est en forma de oxgeno atmico. A
130Km, O
2
y O son igualmente abundantes. Por debajo de esta altura, O
2
es ms abundante
que O.
Por consiguiente, los tomos de O sufren colisiones frecuentes con molculas de O
2
.
Estas colisiones llevan a la formacin del ozono, O
3
O
2 (g)
+ O
-
O
3
*
El ozono es la sustancia ms importante que absorbe fotones con longitudes de onda
mayores. Consideremos cmo se forma el ozono en la atmsfera superior y cmo absorbe
los fotones.
El asterisco sobre O
3
significa que la molcula contiene un exceso de energa. Esta
energa se debe eliminar de la molcula en un tiempo muy corto, o se separa de nuevo la
molcula en O
2
y O. Esta descomposicin es el proceso inverso del que form el O
3
. Estas
molculas pueden liberar la energa en exceso chocando con otro tomo u otra molcula y
transfiriendo el exceso de energa a ellas. Representemos el tomo o la molcula con la cul
choca el O
3
como M. (Normalmente M es N
2
u O
2
debido a que stas son las molculas ms
abundantes). La formacin de O
3
y la transferencia de la energa excedente a M se resumen
en las ecuaciones siguientes:
O
2
+O
-
O
3
*
O
3
*
+M O
3
+M*
F.C.A.I. U.N. CUYO
Catlisis Homognea 23
El ozono es capaz de absorber la radiacin solar, lo que resulta en su
descomposicin en O
2
y O.
O
3
+ h (long. entre 240 y 310nm) O
2
+ O
-
+ calor
Este calor calienta la estratosfera u ozonsfera.
El resultado final de este ciclo es que la cantidad de ozono se mantiene constante,
sino fuera por la capa de ozono en la estratosfera, estos fotones de alta energa penetraran
a la superficie de la Tierra. La vida vegetal y animal como la conocemos no poda sobrevivir
en presencia de esta radiacin de energa tan elevada.
Debido a la energa de disociacin del enlace de N
2
, ste no absorbe fcilmente los
fotones, aun cuando stos tengan suficiente energa como para disociar la molcula. El
resultado general es que en la atmsfera superior se forma muy poco nitrgeno atmico
debido a la disociacin de N
2
.
El xido ntrico (NO) ataca al ozono, pero se regenera a tal punto que una de sus
molculas puede destruir muchos miles de las de ozono. En las ciudades, los xidos de
nitrgeno son responsables de la produccin de ozono, por su accin (con luz solar) sobre
ciertos productos de la combustin de hidrocarburos. El ozono que se produce en lo que
llamamos smog es malo para respirar, porque afecta los pulmones, pero el ozono de la
estratosfera es importantsimo para protegernos de la luz ultravioleta. En un caso, el dixido
de nitrgeno reacciona con tomos de oxgeno en zonas donde se produce ozono; en
cambio, cerca de la superficie terrestre no hay suficientes tomos de oxigeno, y el dixido de
nitrgeno, cuando absorbe luz solar, los produce, para que, a su vez, generen ozono. Son
reacciones qumicas muy parecidas pero con efectos opuestos.
Otro grupo de compuestos que pueden destruir el ozono de la estratosfera son los
xidos de nitrgeno (NO
x
), como NO, NO
2
, N
2
O, N
2
O
2
. Estos compuestos provienen de los
gases expulsados por los aviones supersnicos que vuelan a gran altura, as como por
procesos naturales y por otros procesos hechos por el hombre en la Tierra. La radiacin
solar descompone una cantidad considerable de otros xidos de nitrgenos en xido ntrico
(NO), que tambin destruye la capa de ozono de la siguiente manera:
O
3
O
2
+ O
-
NO + O
3
NO
2
+ O
2
NO
2
+ O
-
NO + O
2
Global 2O
3
3O
2
En la que el NO acta como catalizador.
Otro de los responsables de la destruccin de la capa de ozono son los
CLOROFLUOCARBONOS, empleados en la industria de refrigerantes, aire acondicionado,
solventes, esterilizantes, pinturas, desodorantes, insecticidas aerosoles para el cabello etc.
Son sustancias inertes, por tanto una vez liberados a la atmsfera no se destruyen
sino que permanecen all hasta cuando por difusin alcanzan la estratosfera, sufriendo la
radiacin de alta energa que causando la fotlisis o ruptura inducida por la luz.
CFCl
3
CFCl
2
+ Cl
-
CF
2
Cl
2
CF
2
Cl +Cl
-
El cloro al combinarse con una molcula de ozono la destruye, para luego
combinarse con otras molculas de ozono y eliminarlas. El proceso es altamente daino, ya
que en promedio un tomo de cloro es capaz de destruir hasta 100.000 molculas de ozono.
Este proceso se detiene finalmente cuando este tomo de cloro se mezcla con algn
compuesto qumico que lo neutraliza
Cl
-
+ O
3
ClO + O
2
ClO + O
-
Cl
-
+ O
2
El resultado neto de estas reacciones es la conversin de O
3
en O
2
. Debido a que se
utiliza el Cl en la primera etapa de este mecanismo y se forma en la segunda etapa,
F.C.A.I. U.N. CUYO
Catlisis Homognea 24
funciona como un catalizador. La concentracin de O
3
disminuye en las regiones que tienen
ms cantidad de ClO.
3. CATLISIS EN FASE LQUIDA
A diferencia de las reacciones en fase gaseosa, las que son contempladas bajo la
teora cintica de los gases implican colisiones aisladas entre molculas individuales, los
lquidos no son tan sencillos.
La diferencia fundamental entre las reacciones en fase liquida y en fase gaseosa es
que en fase liquida las molculas de reactivo colisionan continuamente con molculas de
disolvente, en muchas reacciones el disolvente origina una ionizacin, por lo que deben
estudiarse las reacciones en las que intervienen iones. La velocidad se vera influenciada por
la constante dielctrica del disolvente.
Otra diferencia es que existe un nmero mayor de choques por unidad de tiempo, la
energa se transfiera en forma rpida y los equilibrios trmicos - vibracional se alcanzan
rpidamente.
La etapa controlante de la velocidad puede ser la velocidad de difusin de las
molculas de reactivo unas hacia otras para producir la colisin y sus velocidades
dependern de la viscosidad del disolvente. En algunas de estas reacciones el disolvente
esta qumicamente implicado en el mecanismo y en algunos casos actuar como catalizador.
3.1. CATLISIS CIDO-BASE
Todos los problemas cinticos tienen un doble aspecto. En primer lugar se trata de
estudiar la reaccin desde un punto de vista experimental para hallar la ecuacin cintica a
la que responde la velocidad. Esta ecuacin nos da las especies qumicas cuya
concentracin influye en la velocidad. En segundo lugar se trata de plantear un mecanismo
(es decir un sistema de ecuaciones diferenciales) cuya solucin nos lleve a la ecuacin
cintica hallada experimentalmente. Entonces decimos que el mecanismo propuesto es
compatible con los datos experimentales.
Desde un punto de vista experimental:
Cuando se estudia una reaccin susceptible de ser catalizada por cidos bases, se
procede generalmente a estudiar la reaccin a pH constante para encontrar una ecuacin
cintica, y a continuacin se estudia la influencia que el pH pueda tener en la constante (
constantes) de velocidad (k) de la ecuacin cintica obtenida.
En el caso ms general se encontrara que k responde a la siguiente expresin:
k = k
0
+ k
H
+
[H
+
] + k
OH
-
[OH
-
]
donde k
0
es una constante independiente del pH, y que correspondera a la reaccin
sin catalizar. Dependiendo de que trmino predomine se habla de catlisis cida especfica,
catlisis bsica especfica.
Muchas veces ocurre que al utilizar un cido dbil para mantener el pH, el cido (AH),
la base conjugada (A
-
), ambos, actan tambin como catalizadores. En el caso ms
general resultara que la constante k vendra dada por la expresin:
k = k
0
+ k
H
+
[H
+
] + k
OH
-
[OH
-
] + k
AH
[AH] + k
A
-
[A
-
]
Cuando los trminos predominantes en la expresin anterior son los correspondientes
a AH A
-
, se habla de catlisis cida bsica general.
F.C.A.I. U.N. CUYO
Catlisis Homognea 25
Las grficas resultantes de medir las constantes de velocidad en funcin del pH, para
los distintos casos que se pueden dar de catlisis cida bsica especfica se dan en forma
grfica en un diagrama de pH contra log de la velocidad, como se muestra en la figura :
a) Caso general, donde la reaccin es
catalizada tanto por cidos como por bases.
La horizontal AB corresponde a la reaccin no
catalizada o bien catalizada por el solvente.
Ejemplo de este caso son la mutorrotacin de
la glucosa y la hidrlisis de los steres.
b) En este caso el segmento AB no
existe, lo que implica que la reaccin no
cataltica (k
o
) no existe, es decir, no hay
efecto del solvente. Como ejemplos podemos
citar la hidrlisis de amidas, y la halogenacin
de cetonas.
c y d) Reacciones catalizadas
nicamente por cidos (c) o bases (d).
Ejemplos para (c): la hidrlisis de orto-sales y
(d) hidrlisis de B-lactonas.
e y f) El mismo caso de (c) y (d) slo
que en ausencia de la influencia del solvente, ejemplos: la hidrlisis del ster diazoactico y
la plimerizacin de nitrosoacetonamina.
Experimentacin de Brnsted
En 1928, BRONSTED demostr que existe una relacin entre el poder cataltico,
medido por los coeficientes catalticos, y la fuerza de los cidos y bases, expresada por sus
constantes de disociacin. Esta relacin para la catlisis con un cido dbil viene dada por
la ecuacin [1] y para el caso de la catlisis por una base dbil ser
[2]
en donde K
A
es la constante de disociacin del cido dbil, y a, b, y son constantes
para una determinada reaccin, disolvente y temperatura. A partir de estas ecuaciones
puede predecirse el efecto cataltico, de un cido o de una base Brnsted-Lowry, sobre la
velocidad especfica de reaccin, si se conoce la constante de disociacin del electrolito
dbil. Las relaciones de las ecuaciones [1] y [2] se mantienen siempre debido a que, tanto el
poder cataltico, como la constante de disociacin de un electrolito dbil, dependen de la
capacidad de un cido dbil para ceder un protn o de una base dbil para aceptarlo.
En general, cuanto mas fuerte es un cido, es mejor catalizador.
Mecanismos
Catlisis especfica (cida):
1er paso: SH+ + B [SHB+] BH+ + S S= H2O
2 paso: BH+ + S PRODUCTOS + SH+
Ejemplos de mecanismo:
No resulta fcil generalizar sobre mecanismos de catlisis cido-base. No obstante,
con mucha frecuencia nos encontraremos etapas del mecanismo que implican la
transferencia de un H
+
de una molcula de reactivo al catalizador , a la inversa.
Este tipo de etapas de transferencia protnica tienen especial importancia en qumica
orgnica, y en mucha de las etapas de los mecanismos de reacciones catalizadas por
enzimas. Aqu vamos a ver algunos ejemplos procedentes de la qumica orgnica:
F.C.A.I. U.N. CUYO
Catlisis Homognea 26
Alquilacin de Parafinas:
Los alquilatos son derivados
de adicin o sustitucin, obtenidos por
reaccin entre olefinas y alcanos, u
otros derivados, y se utilizan en la
preparacin de gasolinas e
intermedios de sntesis. Se utilizan
catalizadores cidos homogneos,
tales como H
2
SO
4
HF. Los
catalizadores son arrastrados con el
producto y separados por decantacin o destilacin. Una parte suele perderse combinada
con oligmeros de los alquenos. Son catalizadores txicos y corrosivos.
Alquilacin de Fenol
La produccin industrial de
alquilfenoles es del orden de
500000tm/a. Estos derivados,
tales como etilfenol, propilfenol,
butilfenol, etc., son usados como
aditivos de gasolinas, herbicidas,
aditivos de polmeros,
surfactantes, lubricantes o
antioxidantes.
Sntesis de Bisfenol - A
El bisfenol-A es un
intermedio en la obtencin de
resinas epoxi y policarbonatos. Se
obtiene mediante condensacin de fenol
y acetona, catalizada por cidos.
4. OTRAS REACCIONES QUE SE VERIFICAN EN FASE
HOMOGENEA
4.1. CATLISIS DE STERES:
Se caracterizan dentro de este grupo:
Se nombran como alcanoatos de alquilo (metanoato de metilo).
Los steres dan sabor y olor a muchas frutas y son los constituyentes mayoritarios de las
ceras animales y vegetales.
La hidrlisis de los steres est catalizada por cidos o bases y conduce a cidos
carboxlicos.
F.C.A.I. U.N. CUYO
Catlisis Homognea 27
Los steres reaccionan con alcoholes con catlisis cida o bsica obtenindose un nuevo
ster sin necesidad de pasar por el cido carboxlico libre. Esta reaccin se denomina
transesterificacin.
Los reactivos de Grignard transforman los steres en alcoholes. La reaccin no se puede
parar y se produce la adicin de dos equivalentes del organometlico.
El hidruro de aluminio y litio los transforma en alcoholes y el DIBAL en aldehdos.
En medios bsicos forman enolatos que condensan generando 3-cetosteres.
Reaccin denominada condensacin de Claisen
Los steres se hidrolizan formando cidos carboxlicos y alcoholes cuando se les calienta
en medios cidos o bsicos. La hidrlisis de los steres es la reaccin inversa a la
esterificacin.
Esterificacin
Los steres se preparan fundamentalmente por accin de los cidos sobre los
alcoholes. Este mtodo bsico puede modificarse usando un cloruro o un anhidro de cido
con un alcohol o aprovechando la reaccin de una sal de un cido con un halogenuro de
alquilo.
La esterificacin de un cido por ebullicin del mismo con un alcohol, en presencia de
un catalizador cido (comnmente cido sulfrico o cloruro de hidrgeno gaseoso),
constituye el mtodo ms frecuentemente usado para la preparacin de un ster.
Reaccin global
El mecanismo de la esterificacin de Fischer es una sustitucin nucleoflica de acilo,
catalizada por un cido, el grupo carbonilo de un cido carboxlico (en contraste con el
cloruro cido) no es suficientemente electroflico para ser atacado por un alcohol. El
catalizador cido protona al grupo carbonilo y lo activa hacia el ataque nucleoflico (le
aumenta la electrofilia). A continuacin, el alcohol ataca al grupo carbonilo protonado para
formar un intermedio tetradrico, que rpidamente, mediante un proceso de intercambio
protnico forma un nuevo intermedio tetradrico que contiene un excelente grupo saliente: el
agua.
La regeneracin del grupo carbonilo provoca la expulsin de agua y la formacin del
ster protonado. Finalmente, el intercambio protnico con una molcula de agua regenera el
catalizador cido.
F.C.A.I. U.N. CUYO
Catlisis Homognea 28
Perfil energtico
Si se sigue el mecanismo desde el final en forma inversa se tiene el mecanismo de la
reaccin de hidrlisis de los steres. Esta reversibilidad es un inconveniente en la
preparacin directa de un ster a partir de un cido; la preferencia por la va del cloruro de
cido se debe a que ambos pasos; la preparacin de cloruro y la del ster con este ltimo,
son esencialmente irreversibles y llegan a la terminacin.
La reaccin entre un cido y un alcohol es reversible y este tipo de reaccin est
entre aquellas que se usaron en el desarrollo de la ley de accin de masas. Esta ley se
puede aplicar a la formacin de acetato de etilo de la siguiente manera:
k =
alcohol cido
agua ster
C x C
C x C
Si comenzamos con un mol de cido actico y un mol de alcohol o proporciones
similares de agua y acetato de etilo, a su debido tiempo el sistema llega al equilibrio en que
un tercio de mol de cido actico y un tercio de mol de alcohol se hallan en presencia de dos
tercios de mol de acetato de etilo y dos tercios de agua. El valor de la constante de
equilibrio, K se puede calcular a partir de los datos.
4
x
x
3
1
3
1
3
2
3
2
= = k
En otras palabras, el rendimiento de ster obtenido por la reaccin de cantidades
equimoleculares de cido actico y alcohol no puede exceder de 66,66 por ciento. Un
rendimiento de este orden puede no ser econmico y puede mejorarse aumentando la
concentracin de una de las sustancias reaccionantes o disminuyendo la concentracin de
una de las sustancias formadas. Si el ster tiene bajo punto de ebullicin y puede ser
destilado del sistema reaccionante casi tan rpidamente como se forma, la esterificacin
CH
3
COOH + C
2
H
5
OH CH
3
COOC
2
H
5
+ H
2
O
F.C.A.I. U.N. CUYO
Catlisis Homognea 29
Acido r-fenilbutrico
r-fenilbutirto de etilo
OH
O
2 2 2
CH CH CH C 5
alcohol
etlico
C H OH
2
+
H SO
2 4
O
OC H
CH CH CH C
2 2 2
5 2
H O
2 +
puede hacerse prcticamente completa por este medio. Sin embargo, la separacin del
ster no siempre es posible y a veces es conveniente reducir la reversibilidad de la reaccin
mediante un agente que elimine el agua. En muchos casos, la eliminacin azeotrpica del
agua mediante el benceno, tolueno o xileno da excelentes resultados, y este mtodo se est
difundiendo rpidamente.
La facilidad de formacin de un ster est estrechamente ligada con la geometra de
las molculas del cido y alcohol usados en su preparacin. Los cidos en los que la cadena
de tomos de carbono se ramifica en el carbono alfa o beta o los cidos con sustituyentes
en los carbonos alfa o beta, son esterificados ms lentamente que los cidos cuyos tomos
de carbono alfa o beta contienen slo hidrgeno. En una molcula de alcohol, la ramificacin
en el carbono carbinlico disminuye claramente la velocidad de esterificacin; por
consiguiente, los alcoholes secundarios forman steres ms lentamente que los alcoholes
primarios y los alcoholes terciarios son los que, se esterifican ms lentamente.
Sin embargo, la esterificacin directa tiene la ventaja de ser una sntesis de un solo
paso. A menudo resulta til si aplicamos nuestros conocimientos de los equilibrios. Si el
cido o el alcohol son baratos y de fcil adquisicin, se utiliza un exceso para desplazar el
equilibrio hacia la formacin de los productos y aumentar as el rendimiento en ster. Por
ejemplo, conviene emplear 8 moles del econmico alcohol etlico para convertir un mol del
muy valioso cido -fenilbutrico ms completamente en el ster:
Si se desea esterificar un cido hay que utilizar un exceso de alcohol y, si es posible,
eliminar el agua de la reaccin.
Sntesis de steres a partir de cloruros y anhidridos de cido
Los steres tambin se pueden sintetizar mediante la reaccin de cloruros de cido o
anhdridos de cido con alcoholes. Como los cloruros de cido y los anhdridos son muchos
ms reactivos hacia el proceso de la sustitucin nucleoflica que los cidos carboxlicos, la
reaccin de esterificacin tiene lugar de forma rpida y sin la presencia de catalizador cido.
Cuando se emplean cloruros de cido y anhdridos para las reacciones de esterificacin hay
que emplear una base, usualmente piridina, para neutralizar el HCl o el cido carboxlico
que se forma en el proceso.
Ejemplos de esterificacin de cidos y alcoholes:
Por el mtodo de Fischer puede prepararse una gran variedad de steres. Los
catalizadores habituales son los cidos sulfrico, clorhdrico y p-toluenosulfnico.
BrCH CO H
2 2
+ C H OH
2 5
+
2
H SO
4
BrCH CO C H
2 2
2
5
+
2
H O
cido bromoactico etanol bromoacetato de etilo
K-O CC
2
butinodiato de potasio
CCO -K
2
+ 2CH OH
3
+
2
H SO
4
metanol
H CO CC
butinodiato de dimetilo
2
3
CCO CH
2 3
F.C.A.I. U.N. CUYO
Catlisis Homognea 30
HO CCO H
2 2
+ 2CH OH
3
H SO
2 4
+
H CO CCO CH
2 2 3 3
+
H O
2
cido etanodioico
metanol etanodioato de dimetilo
Catalizadores para la esterificacin
La eleccin del catalizador depende de varias cosas y vara segn el sistema que se
considere. Los catalizadores ms comunes son los cidos inorgnicos fuertes, si bien en
ocasiones se emplean sales, el gel de slice y resinas de intercambio de cationes.
Entre los cidos minerales, el cloruro de hidrgeno es el catalizador ms usual en
estudios de laboratorio, pero en industrias no se usa por su corrosividad u de ciertos
requisitos del proceso. Puede usarse el gas cido clorhdrico anhidro y el cido clorhdrico
acuoso comercial. En reacciones colaterales se forman cloruros de alquilos cuando es
prolongado el tiempo de reaccin. Los alcoholes terciarios reaccionan con cloruro de
hidrgeno para formar rpidamente cloruros de alquilo. El cido clorhdrico acuoso da
buenos resultados en la esterificacin de cidos aromticos.
Para la mayora de las operaciones industriales se prefiere el uso de cido sulfrico,
pero con algunos alcoholes secundarios, como el ciclohexanol y con alcoholes terciarios
puede ocurrir la deshidratacin y conversin en las oleofinas correspondientes, la
isomerizacin o la polimerizacin. En general con cidos aromticos se requiere mayor
cantidad de cido sulfrico que son los alifticos. Se han recomendado los cidos
bencenosulfnico y p-tolueno sulfnico como catalizadores para la esterificacin de cidos
grasos de cadena larga.
Algunas veces se emplea el cido fosfrico, pero es muy lento. Los cloruros de
magnesio, aluminio, frrico, de cinc, cprico y estnnico producen efectos aceleradores en
la esterificacin. Las sales de mercurio, plata, cobalto, nquel son catalizadores activos
cuando no hay cidos inorgnicos.
Como la esterificacin es un proceso que an usando catalizador puede requerir das,
se ha experimentado una tcnica que utiliza un microondas especial que calienta al reactor,
lo que acelera la velocidad de reaccin.
4.2. Hidrlisis de los steres
Los steres son descompuestos por el agua en sus componentes, cido y alcohol.
Empleando cantidades equimolares de los mismos, se produce una reaccin bimolecular y
reversible cuyo estado final o de equilibrio es el mismo que para la de esterificacin.
La hidrlisis es muy lenta en medio neutro y a temperatura ordinaria. Pero a
temperaturas mayores a 100C se acelera y completa en pocas horas.
La velocidad de hidrlisis puede ser aumentada aadiendo a la mezcla una pequea
cantidad de un cido fuerte que acta como catalizador. Mientras ms fuertes son estos
cidos la hidrlisis es ms rpida.
El mecanismo de hidrlisis cida es el inverso al de esterificacin de cidos
carboxlicos. Es un proceso en equilibrio, pero puede ser desplazado prcticamente hasta su
terminacin mediante el uso de un gran exceso de agua.
La hidrlisis se puede hacer en medio alcalino (saponificacin) y es una tpica
reaccin bimolecular, de segundo orden y no reversible.
Su velocidad es ms de 1000 veces mayor que con los cidos. Debido a que la
energa de actividad es 35% menor para un medio alcalino, con respecto a la hidrlisis
catalizadas por cidas.
Para una misma concentracin, se deduce que la velocidad de saponificacin debe
ser determinada por la concentracin de el ion OH- (mayor en bases fuertes)
O
CH
3
C O C
2
H
5
O
-
CH
3
C O C
2
H
5
OH
+ OH
-
O
-
CH
3
C O C
2
H
5
OH
O
CH
3
C OH + CH
3
O
-
O
CH
3
C OH + CH
3
O
-
O
CH
3
C O
-
+ CH
3
OH
1)
2
3)
F.C.A.I. U.N. CUYO
Catlisis Homognea 31
Los distintos tipos de hidrlisis se dividen en los siguientes tipos:
- Hidrlisis pura con agua sola.
- Hidrlisis con cido acuoso diluido o concentrado.
- Hidrlisis con lcali acuoso diluido o concentrado.
- Fusin con lcali con poco o nada de agua.
- Hidrlisis con enzima como catalizadores
4.3. Hidrlisis cida
Se da por entendido que la solucin es acuosa y acidulada
Un cido mineral acelera ambos procesos protonando le oxgeno carbonlico, con lo
que el carbono del grupo resulta ms susceptible al ataque nucleoflico.
Un mecanismo propuesto para la hidrlisis catalizada especficamente por H
3
O
+
es el
siguiente:
k
1
R
1
COOR
2
(E)
+ H
3
O
+
R
1
COH
+
OR
2
( I
1
) + H
2
O
k
2
k
-1
R
1
COH
+
OR
2
( I
1
) + H
2
O R
1
COOH
2
+
(I
2
) + R
2
OH
R
1
COOH
2
+
(I
2
) + H2O R
1
COOH
(A) + H
3
O
+
La primera etapa es una protonacin del carbonilo. Se forma una especie intermedia
con carga neta positiva. Esta especie intermedia reacciona entonces con agua liberndose
el alcohol. Finalmente, en una tercera etapa muy rpida se libera el cido a partir de un
intermedio, y se regenera el catalizador, H
3
O
+
La velocidad de la reaccin la podemos expresar como v = d[R
2
OH] / dt = k
2
[I
1
] .
Aplicando la aproximacin del estado estacionario al intermedio I
1
, tendramos:
d[I
1
] / dt = k
1
[E][H
3
O
+
] - k
-1
[I
1
] - k
2
[I
1
] ~ 0
[I
1
] = k
1
[E][H
3
O
+
] / ( k
-1
+ k
2
) y substituyendo en la expresin de la velocidad:
v = k
2
k
1
/ ( k
-1
+ k
2
) [E][H
3
O
+
]
Resultando una ecuacin cintica correspondiente a catlisis cida especfica.
Los ejemplos que vamos a ver nos van a servir para desarrollar algunas discusiones
relativas a mecanismos de reaccin. En el ejemplo anterior se puede comprobar que si se
introduce una etapa adicional entre la primera y la segunda, en la que el intermedio I
1
se
convierte en I
1
, siendo esta ltima molcula la que choca con H
2
O en la tercera etapa para
generar R
2
OH e I
2
, el resultado, al aplicar la aproximacin del estado estacionario es la
misma ecuacin cintica obtenida anteriormente. Este resultado se puede generalizar. La
introduccin de etapas que supongan la isomerizacin de especies intermedias no cambia
la ecuacin cintica resultante de la aplicacin del estado estacionario. De forma que la
base
F.C.A.I. U.N. CUYO
Catlisis Homognea 32
identificacin de especies intermedias require mtodos adicionales a los aqu descritos, que
permitan detectar directa indirectamente dichas especies.
4.4. Hidrlisis alcalina
La base promueve la hidrlisis de los steres porque proporciona el reactivo
fuertemente nuclefilo. Esta reaccin es irreversible, porque se forma un anin
carboxilato estabilizado por resonancia que demuestra poca tendencia a reaccionar con
un alcohol.
Observemos los diversos aspectos del mecanismo presentado.
En primer lugar tenemos el ataque del hidrxido sobre el ster, esto nos dice que
presenta una cintica de segundo orden ya que la velocidad de la reaccin depende de la
concentracin del hidrxido y del ster.
Luego el ataque del hidrxido al carbono carbonlico y desplaza al ion alcxido, esto
indica una ruptura del enlace entre el oxgeno y el grupo acilo
Saponificacin De steres
Las grasas y los aceites constituyen los lpidos ms abundantes; ambos grupos estn
constituidos prcticamente por un 100% de triglicridos los que a su vez son steres de
cidos grasos con glicerol. Consecuentemente, dichos cidos representan un alto porcentaje
de la composicin de los triglicridos y de las grasas y aceites.
Estructura molecular de un triglicrido
O
CH
2
O C R
1
O
CH
O C R
2
O
CH
2
O C R
3
Glicerol cidos grasos
Triglicrido
Los cidos grasos pueden ser saturados o insaturados.
El primer grupo est constituido principalmente por cidos de 4 a 24 tomos de
carbono, los de C
4
a C
8
son lquidos a 25C, mientras que los de C
10
en adelante son
slidos. Entre los ms comunes est el cido lurico, que abunda en el aceite de coco, y el
palmtico que se encuentra en los lpidos de la palma. En el siguiente cuadro se muestran
F.C.A.I. U.N. CUYO
Catlisis Homognea 33
los distintos tipos de cidos grasos saturados, como as tambin su frmula qumica y
temperaturas caractersticas.
Cuadro 1-cidos grasos saturados
Nombre trivial Nombre cientfico Frmula Punto de
fusin(C)
Punto de
ebullicin(C)
Butrico
Caproico
Caprlico
Cprico
Lurico*
Mirstico*
Palmtico*
Esterico*
Araqudico
Behnico
Lignocrico
Certico
Butanoico
Hexanoico
Octanoico
Decanoico
Dodecanoico
Tetradecanoico
Hexadecanoico
Octadecanoico
Eicosanoico
Docosanoico
Tetracosanoico
Hexacosanoico
CH
3
(CH
2
)
2
COOH
CH
3
(CH
2
)
4
COOH
CH
3
(CH
2
)
6
COOH
CH
3
(CH
2
)
8
COOH
CH
3
(CH
2
)
10
COOH
CH
3
(CH
2
)
12
COOH
CH
3
(CH
2
)
14
COOH
CH
3
(CH
2
)
16
COOH
CH
3
(CH
2
)
18
COOH
CH
3
(CH
2
)
20
COOH
CH
3
(CH
2
)
22
COOH
CH
3
(CH
2
)
24
COOH
-5,9
-3,4
16,7
31,6
44,2
54,4
63,0
69,4
76,0
79,9
84,2
87,7
164
206
240
271
130
149
167
184
204
-
-
-
*cidos grasos saturados ms comunes en alimentos
En cuanto a los insaturados, son abundantes en los aceites vegetales y marinos. Los
ms comunes se resumen en el siguiente cuadro.
Cuadro 2-cidos grasos insaturados
Nombre trivial Nombre cientfico Frmula Punto de fusin(C)
Palmitoleico
Oleico
Linileico
Linolnico
Araquidnico
Vaccnico
Gadoleico
Ercico
Brasdico
Cetoleico
Hexadeca-9-enoico
Octadeca-9-enoico
Octadeca-9,12-dienoico
Octadeca-9,12,15-trienoico
Eicosa-5,8,11,14-tetraenoico
Octadeca-11-enoico
Eicosa-11-enoico
Docosa-13-enoico
Docosen-13-enoico
Docosen-11-enoico
C
15
H
29
COOH
C
17
H
33
COOH
C
17
H
31
COOH
C
17
H
29
COOH
C
19
H
31
COOH
C
17
H
32
COOH
C
19
H
37
COOH
C
21
H
40
COOH
C
21
H
40
COOH
C
21
H
40
COOH
-0,5
13,0
-5,0
-11,0
-49,5
39,5
23,5
38,0
-
-
Reacciones De Grasas Y Aceites
Los caracteres qumicos de las grasas son los enlaces de Ester y la instauracin.
Como esteres pueden ser hidrolizados, en presencia de cidos, enzimas, o lcalis, a sus
cidos grasos libres, o sus sales y glicerol, respectivamente.
Triesterina + 3 H
2
O acido graso + glicerol
Triesterina + 3 M
+
-
OH sal de acido graso + glicerol
H
+
o enzimas
F.C.A.I. U.N. CUYO
Catlisis Homognea 34
Como los cidos grasos difieren en peso molecular y debido a que pueden estar
presentes substancias que no reaccionan con los lcalis, como alcoholes e hidrocarburos,
grasas diferentes requieren cantidades diferentes de lcalis para saponificarse. por lo tanto,
la cantidad de lcali necesaria para saponificar un peso determinado de grasa puede usarse
como propiedad caracterstica de una grasa particular.
El proceso de hidrlisis bsica de los steres se denomina saponificacin. ste
trmino proviene del latn que significa jabn. Cuando se hidrolizan las grasas o aceites con
NaOH, se obtiene glicerina (propanotriol) y las correspondientes sales sdicas de los cidos
carboxlicos de cadena larga, que son los que conocemos como jabn.
La reaccin qumica involucrada es:
Esta reaccin, al contrario que el proceso de esterificacin de Fischer, es irreversible.
El mecanismo del proceso que permite explicar esto es: el ion hidrxido ataca al carbonilo
del ster formando un intermedio tetrahdrico. Cuando se regenera el grupo carbonilo se
produce la eliminacin del in alcxido y se forma un cido carboxlico. Una rpida
transferencia de protn forma el carboxilato y el alcohol.
Este ltimo paso es muy exotrmico y desplaza los dos equilibrios anteriores del
proceso de saponificacin hacia su terminacin, haciendo que el proceso sea irreversible.
En el segundo paso del mecanismo anterior, se produce la prdida de ion alcxido. En
el estudio de las reacciones de sustitucin (SN1 y SN2) se afirm que las bases fuertes
como el in hidrxido o los alcxidos no son buenos grupos salientes debido a su elevada
basicidad.
Entonces, cmo es posible explicar la eliminacin de un grupo saliente bsico, como
MeO-, en el anterior mecanismo? Las diferencias entre los mecanismos SN1 y SN2 explican
F.C.A.I. U.N. CUYO
Catlisis Homognea 35
por qu estas bases fuertes pueden servir como grupos salientes en la reaccin de
saponificacin, adems que proporcionan la cintica que sigue el proceso.
El mecanismo de la reaccin SN2 tiene lugar en un solo paso, ni muy endotrmico ni
muy exotrmico. El enlace con el grupo saliente est parcialmente roto en el estado de
transicin, de modo que la velocidad de la reaccin es muy sensible a la naturaleza del
grupo saliente. Con un mal grupo saliente, como un alcxido, esta reaccin es muy lenta.
En el mecanismo SN1, el enlace con el grupo saliente se rompe en un segundo paso
del mecanismo. Este segundo paso es muy exotrmico y en este estado de transicin el
enlace con el grupo saliente apenas se ha comenzado a romper. En general, una base
fuerte puede funcionar como grupo saliente si se elimina en un paso muy exotrmico,
convirtiendo un intermedio inestable y con carga negativa, en una molcula estable.
Entonces, la saponificacin sigue un mecanismo de reaccin SN1.
A continuacin, se comparan las grficas de la energa de la reaccin para una
proceso SN2 y para una reaccin de saponificacin de un ster metlico SN1. En la reaccin
SN2 el metxido se aleja en un paso ligeramente endotrmico, y el enlace con el metxido
se rompe casi totalmente en el estado de transicin.
En la saponificacin el alcxido se aleja en un segundo paso exotrmico con un
estado de transicin semejante a los reactivos. El enlace del metxido apenas se ha
comenzado a romper en el estado de transicin.
Mecanismo SN2
Mecanismo SN1
Cintica e Influencia Del Ph
Al seguir la saponificacin un mecanismo de sustitucin nucleoflica
unimolecular(SN1) la cintica que es de esperar es la de una de primer orden, donde slo
dependa de la concentracin del ster:
| | ster k r =
Cuando se estudia experimentalmente una reaccin susceptible de ser catalizada por
cidos bases, se lleva a cabo la reaccin a pH constante para encontrar una ecuacin
cintica, y a continuacin se estudia la influencia que el pH pueda tener en la constante de
velocidad k de la ecuacin cintica obtenida.
Hay bibliografas que sugieren que para cualquier reaccin catalizada por cidos
bases, la constante es una funcin lineal de la concentracin de OH
-:
| |
+ = OH k k k
OH
0 obs ,K
0
= constante independiente del pH, K
OH
= constante
dependiente del pH
Sin embargo, habamos dicho que de acuerdo al mecanismo SN1 la velocidad de la
reaccin deba ser independiente de la concentracin de oxhidrilo, entonces slo puede
significar que la cintica no involucra [OH], con lo cual es de esperar que el segundo trmino
de k de la ecuacin anterior no modifique sustancialmente la velocidad de la reaccin.
Estas observaciones pueden interpretarse si suponemos que la etapa controlante, es
decir la de menor velocidad, es la de formacin del carbocatin de la primera etapa de la
reaccin, seguida de un ataque nucleoflico del OH mucho ms veloz. Esto permite explicar
porque el mecanismo SN1 sigue una cintica de primer orden donde la concentracin de
oxhidrilo no influye demasiado.
F.C.A.I. U.N. CUYO
Catlisis Homognea 36
Un anlisis interesante de hacer, es que ambos mecanismos(SN1 y SN2) coexisten
durante la reaccin siendo uno preponderante sobre el otro. Para nuestro caso la SN1 es
ms importante porque el grupo alcxido acta como buen grupo saliente cuando se elimina
en un paso exotrmico. Pero si aumentamos la [OH], es decir el pH, empieza a tomar
importancia el mecanismo SN2, ya que su cintica depende de esto. Sin embargo, como el
grupo saliente no es bueno y el paso es ligeramente endotrmico su cintica es baja,
significando esto que no es bueno trabajar a pH bsicos extremos.
4.5. Alcoholisis de steres
Un mecanismo similar al anterior se puede plantear para la alcoholisis
tranesterificacin de un ster, substituyendo H
2
O por R
3
-OH. Las etapas que se plantean
son las mismas que en el mecanismo anterior, y es fcil comprobar que v = k
2
k
1
[R
3
-
OH] [E][H
3
O
+
] / ( k
-1
+ k
2
[R
3
-OH]) .
F.C.A.I. U.N. CUYO
Catlisis Homognea 37
5. Catlisis en la polimerizacin de olefinas
5.1. Caractersticas de las poliolefinas:
Bajo Costo: Materia simple y barata, produccin en procesos exotrmicos muy
eficientes y en grandes toneladas.
Livianas: Baja densidad, tomos de bajo peso atmico.
Inocuas e Inertes: Sin grupos funcionales ni elementos pesados.
Verstiles: Termoplsticas, diferentes grados de flexibilidad, elasticidad, dureza.
Aspecto agradable: Transparentes u opacas, coloreables, maleables.
Desechables y Reciclables: Craqueo a ceras, aceites combustibles, monmero.
- Mecanismo de polimerizacin por radicales
Una polimerizacin por radicales se inicia por radicales libres. El catalizador acta en
la etapa de iniciacin, formando un radical libre e inmediatamente produciendo la fijacin de
una primera molcula de monmero.
Utilizacin de catalizador como iniciador
Un iniciador de polimerizacin es un compuesto capaz de producir radicales libres
generalmente por elevacin de temperatura. Su empleo asegura una cantidad dada de
radicales libres a una temperatura mucho ms baja que por iniciacin nicamente trmica.
La molcula de iniciador se divide para dar generalmente dos radicales: I 2 R*
Entonces se podr escribir:
| |
| | I kd
dt
R d
V
i
2
*
= =
Pero todos los radicales formados no son activos. Es necesario utilizar un coeficiente f
llamado eficiencia, que representa la fraccin activa de los radicales producidos, entonces
| | I fkd V
i
2 =
Este coeficiente se calcula en forma experimental, y su valor depende de la
probabilidad de combinacin, posibilidad de dar radicales inactivos en polimerizacin,
posibilidad de terminacin con radical primario.
Iniciadores comunes:
Los hidroperxidos como el hidroperxido de ter-butilo.
Los perxidos: El ms sencillo es el agua oxigenada la cual se emplea en sistema
redox.
Los perxidos de dialquilos como el perxido de diterbutilo.
Los perxidos de diarilos como el perxido benzoilo.
Los persteres: Todos estos compuestos se descomponen bajo la accin de calor y
dan radicales por rotura del enlace.
Los compuestos azoicos: Se descomponen con eliminacin de una molcula de
nitrgeno. El ms conocido es el azobisisobutironitrilo (AZBN).
Estabilidad del iniciador = Tiempo de vida media. La elevacin de temperatura
proporciona la energa E necesaria para la descomposicin del iniciador. La velocidad de
descomposicin es mayor a medida que la temperatura se eleva.
La constante de velocidad de disociacin sigue la ley de Arrhenius
RT E kd
Ae dondeKd R I
/
* 2
=
Cada iniciador, a una temperatura dada, esta caracterizado por su tiempo de vida
media y es el tiempo necesario para que se descomponga la mitad de la cantidad inicial de
iniciador. Este tiempo depende de la temperatura.
En la reaccin de iniciacin, es en donde se produce la adicin de un monmero, una
vez formado, el radical reacciona sobre una molcula del mismo, o sea, es la primera etapa
de la reaccin en cadena. R* + M R-M*
F.C.A.I. U.N. CUYO
Catlisis Homognea 38
- Mecanismo de polimerizacin por coordinacin
Est basada en que cada elemento o tomo central posee un nmero de coordinacin
mxima. Los procedimientos se llaman de coordinacin cuando el catalizador est en forma
de complejo presentando por lo menos un tomo capaz de coordinar la molcula de
monmero.
En la primera etapa, la olefina se polariza por influencia del catalizador, obtenindose
una disociacin del tipo inico.
El crecimiento de las cadenas se efecta por adicin de otras unidades monomricas
por un mecanismo de polimerizacin del tipo aninico.
Iniciacin:
| | | |
5 6 2 5 6 2 3 2 5 6 5 6
H C CH CH XnAl H C CH CH XnAl CH CH CH H C XnAl H C XnAl +
+ + +
Propagacin:
( ) ( ) | |
5 6 2 2 5 6 2 2
2 3 5 6 2
H C CH CH CH CH XnAl H C CH CH CH XnAlCH
CH CH nCH H C CH CH XnAl
n n
= +
+
+
- Polimerizacin metalocnica
A mediados de los aos 50, la industria de los polmeros experiment un gran
desarrollo gracias al descubrimiento de los catalizadores Ziegler-Natta. Estos catalizadores
estn basados en sales de metales de transicin (TiCl
3
, TiCl
4
) que en presencia de un co-
catalizador de alquil aluminio permiten las sntesis de poliolefinas en condiciones no
extremas de presin y temperatura. Tambin logran tener un control sobre las propiedades
del material obtenido junto a un considerable aumento de las actividades.
Hay que notar que la estreoselectividad de los catalizadores es de mucha
importancia para las -olefinas ya que la posicin del grupo sustituyente lateral genera un
centro quiral, lo que lleva a distintas configuraciones espaciales, lo que se traduce en
diversas propiedades fsicas del polmero. En el caso del polipropileno estas diferencias se
deben a la presencia de un grupo metilo en el propileno y a la disposicin del mismo de
acuerdo al plano de simetra establecido por la cadena principal. Si la configuracin
resultante de un polmero muestra a todos los grupos laterales por sobre (o por debajo) del
plano de la cadena principal, la configuracin se denomina isotctica. Si los grupos laterales
quedan alternadamente por encima y por debajo del plano, la configuracin es sindiotctica,
mientras que si la secuencia de posiciones se da en forma aleatoria se dice que posee una
configuracin atctica. Esto determina las propiedades fsicas del producto, as por ejemplo
si el polmero es isotctico, por poseer una estructura ordenada el material resultante va a
tender a ser cristalino, en cambio si es atctico el material resulta altamente amorfo debido
al desorden existente en las cadenas polimricas.
(a) Polipropileno atctico (b) Polipropileno isotctico (c) Polipropileno sindiotctico
F.C.A.I. U.N. CUYO
Catlisis Homognea 39
Sin embargo en los catalizadores Ziegler-Natta, el centro activo del metal ocupa una
posicin en la superficie de un cristal con una estructura de enrejado compatible (sistema
heterogneo), por lo que el centro activo puede ocupar una variedad de sitios del enrejado
del cristal, por lo que los sitios activos no son uniformes, lo que se traduce en distintas
reactividades, lo que impide obtener polmeros con largos de cadena similares llevando a
distribucin de pesos moleculares variados.
Los catalizadores metalocnicos son complejos organometlicos tipo sndwich,
anlogos al ferroceno. Estos catalizadores estn formados por metales de transicin del
grupo IV-B (Ti, Zr, Hf,etc.) donde el tomo metlico central est normalmente ligado a dos
anillos aromticos (del tipo ciclopentadienil o derivados), a travs de una interaccin tipo , y
a ligandos como iones cloruro.
La polimerizacin por metalocenos est causando una gran sensacin en el negocio
de los plsticos, porque resulta lo ms indicado para competir con los polmeros vinlicos
desde que se invent la polimerizacin Ziegler-Natta. La razn es que la polimerizacin
catalizada por metalocenos permite por ejemplo producir polietileno capaz de detener las
balas. Este nuevo polietileno es mejor que el Kevlar para la fabricacin de chalecos a
prueba de balas. Y puede lograrlo porque tiene un peso molecular mucho ms alto que el
polietileno sintetizado con la receta de Ziegler-Natta, hasta seis o siete millones.
Con este nuevo tipo de catalizadores es posible obtener polmeros con distribucin de
pesos moleculares estrecha, as como la obtencin de nuevos materiales gracias a la
posibilidad de incorporar comonmeros uniformemente. Tambin permite hacer polmeros
con tacticidades muy especficas, es decir, se caracterizan por su alta estereoregularidad lo
que permiti obtener polipropileno (PP) altamente sindiotctico, atctico o isotctico,
posibilitando la produccin de PP con nuevas caractersticas.
Variables de operacin
En los procesos de polimerizacin las variables de operacin afectan a las
propiedades del polmero (peso molecular, estereoselectividad, etc) y limitan la actividad del
catalizador.
Temperatura de polimerizacin (Tp): la actividad, la estereoregularidad del sistema
cataltico y el peso molecular del polmero dependen de la temperatura. De manera general
los catalizadores metalocnicos exhiben un rango de temperatura donde la actividad
cataltica es mxima. As algunos sistemas presentan un aumento notorio de la actividad
con la temperatura. Inicialmente este comportamiento fue atribuido a la alta energa
requerida para la polimerizacin. Despus de saber que la especie activa deba poseer un
carcter inico, la fuerte dependencia de la actividad con la temperatura paso a ser
explicada en trminos de la energa de activacin necesaria para la formacin de la especie
activa. Un aumento de Tp promueve una distorsin en la conformacin de los ligantes en los
catalizadores homogneos, lo mismo para sistemas rgidos. El resultado de esto es una
menor capacidad de estereoregularidad del catalizador, reduciendo la tacticidad de los
polmeros formados. El peso molecular de los polmeros tambin disminuye con el aumento
de la Tp. La tasa de desactivacin crece con la Tp favoreciendo los procesos de
transferencia de cadena (finalizacin de cadena) y la desactivacin bimolecular.
Concentracin del catalizador: La concentracin de catalizador no tiene un efecto
significativo en la actividad. El efecto del aumento de la concentracin de catalizador y la
disminucin del peso molecular es ocasionado, posiblemente, por el incremento de
transferencias de cadena
Concentracin de MAO: La actividad cataltica de los compuestos metalocnicos es
fuertemente dependiente de la cantidad de MAO puesta para la activacin. La mayora de
los sistemas presentan una razn optima entre las concentraciones de MAO y de
catalizador.
Concentracin de monmero: El aumento de esta produce una
disminucin de la actividad
La mayor concentracin de monmero tiende a aumentar el peso molecular, aunque
este efecto no es muy marcado.
F.C.A.I. U.N. CUYO
Catlisis Homognea 40
Un metaloceno es un ion metlico con carga positiva, entre medio de dos aniones
ciclopentadienilo, con carga negativa. El anin ciclopentadienilo se forma a partir de la
molcula de ciclopentadieno.
Existe un tomo de carbono con dos hidrgenos cidos, es decir, que pueden
desprenderse con facilidad abandonando los electrones de enlace. Cuando existe
desprendimiento de uno de los hidrgenos, el carbono queda con un par electrnico extra,
de modo que se tiene un anillo con seis electrones y por lo tanto, se volver aromtico (el
anillo en esta forma aninica ser sumamente estable).
Estos iones ciclopentadienilo tienen carga -1, de modo que ante un catin como el
zirconio o el titanio con carga +4, stos se unen a los dos anillos aromticos y para
balancear la carga, se unirn tambin a dos iones cloruro, cada uno con carga -1, para dar
un compuesto neutro.
Esos ligandos extras, los cloruros, ocupan un espacio. Resulta difcil para ellos
deslizarse entre los anillos ciclopentadienilo. Por lo tanto, para hacerles lugar a los cloruros,
los anillos se inclinan entre s, abrindose como el caparazn de una almeja. Esto les da a
los cloruros, espacio para respirar. Observe sto en la figura de abajo:
Varias modificaciones a estos catalizadores se han realizado con el objetivo de
obtener sistemas homogneos con alta actividad y muy buena estereoselectividad. Las
F.C.A.I. U.N. CUYO
Catlisis Homognea 41
modificaciones incluyen distintos tipos de ligandos, el tamao de stos y la presencia de
puentes ligados a los dos anillos que dan al catalizador mayor rigidez y varan la apertura
del anillo. Se ha visto que los ligantes voluminosos aumentan el impedimento estrico para
la coordinacin del monmero. Esto promueve una discriminacin en la orientacin
favoreciendo inserciones donde la repulsin entre los substituyentes del monmero (por
ejemplo el grupo metil en el propileno) de la ltima unidad insertada entre este y los ligantes
sea minimizada. Por otro lado la presencia de grupos sustituyentes voluminosos dificulta la
aproximacin del monmero, reduciendo la velocidad de propagacin, o sea, la actividad
cataltica disminuye. Adems la presencia de sustituyentes dadores de electrones (como el
grupo metilo) sobre los anillos aromticos promueve un aumento en la actividad cataltica.
Por ejemplo:
Se diferencia del bis-clorozirconoceno porque cada anillo ciclopentadienilo tiene
fusionado un anillo aromtico de seis carbonos, mostrado en rojo. Este sistema de dos
anillos formado a partir de un anillo ciclopentadienilo fusionado con un anillo fenilo, se
denomina ligando indenilo. Por otra parte, hay un puente etileno, representado en azul, que
une los anillos ciclopentadienilo, el superior y el inferior. Estas dos caractersticas hacen de
este compuesto, un excelente catalizador para la obtencin de polmeros isotcticos.
Los voluminosos ligandos indenilo, dispuestos en posiciones opuestas como estn,
dirigen los monmeros entrantes, as slo pueden reaccionar cuando se encuentren en la
direccin correcta, para dar polmeros isotcticos. Ese puente etileno mantiene fijos a los
dos anillos indenilo. Sin el puente, giraran y podran no situarse en el lugar correcto para
dirigir la polimerizacin isotctica.
El efecto de los grupos sustituyentes, como por ejemplo Me, Et, etc, en el grupo
ciclopentadienil se estudi en la polimerizacin de polietileno, y se verific que los grupos
voluminosos producen una desactivacin menor en la reaccin en funcin del tiempo. De
acuerdo con otro estudio, se conoce que los efectos electrnicos son ms importantes que
los efectos estricos, pero a medida que los grupos sustituyentes se tornan ms
voluminosos, los efectos estricos comienzan a predominar sobre los electrnicos. El
balance entre efectos estricos y electrnicos de los sustituyentes determina el aumento o
disminucin en la actividad debido a la estabilizacin de la especie propagadora.
Hemos hablado sobre lo que son los metalocenos y por qu son capaces de hacer
polmeros con una tacticidad especfica. Pero no hemos dicho nada acerca de cmo ocurre
realmente la polimerizacin. Por eso ahora mismo vamos a tratarlo. Para lograr que nuestro
complejo zirconoceno catalice una polimerizacin, lo primero que debemos hacer es agregar
una pizca de algo llamado MAO. En realidad es la abreviatura de metil aluminoxano.
Los catalizadores metalocenos as descritos no tienen actividad sino que necesitan del
metilaluminoxano (MAO) como co-catalizador Los aluminoxanos consisten en unidades
oligomricas de [-Al(Me)O-] que pueden estar en forma lineal y/o cclica. Los cloruros del
zirconoceno son lo que llamamos lbiles. Esto quiere decir que pueden desprenderse
fcilmente Por lo tanto el MAO puede reemplazarlos con algunos de sus grupos metilo
Funciones del MAO en la polimerizacin:
F.C.A.I. U.N. CUYO
Catlisis Homognea 42
Alquilacin del metaloceno, produccin de la especie catinica y estabilizacin de
estas especies.
Eliminacin de impurezas del sistema reaccional, como agua u oxgeno.
Reactivacin de especies inactivas o durmientes del metaloceno. Tambin puede
participar en la terminacin de la cadena
En la siguiente figura puede observarse el complejo al cual se llega (se representa
parte del catalizador para ahorrar espacio) cuando esto ocurre:
El zirconio cargado positivamente es
estabilizado, ya que los electrones del enlace carbono-hidrgeno son compartidos con el
zirconio (a-asociacin agstica). Pero necesita ms de una asociacin agstica para ser
satisfecho, y esa necesidad es la que facilita la
entrada al monmero olefnico, ya que su doble
enlace carbono-carbono posee electrones para
compartir:
La estabilidad del complejo no es muy
prolongada ya que los electrones del enlace zirconio-
carbono metlico se desplazan para formar un enlace
entre el carbono metlico y uno de los carbonos del
alqueno (propileno en este caso). Entre tanto, el par
electrnico que haba participado del enlace alqueno-
complejo metlico se desplaza para formar un enlace
entre el zirconio y uno de los carbonos del propileno:
Como puede observarse en la figura, el proceso
ocurre a travs de un estado de transicin de cuatro
miembros, y el zirconio termina
perdiendo un ligando pero con
una asociacin agstica con un
enlace C-H del monmero
propileno. Entonces, otro
monmero propileno puede
F.C.A.I. U.N. CUYO
Catlisis Homognea 43
aparecer y reaccionar del mismo modo que lo hizo el primero.
El propileno se coordina con el zirconio... y luego los electrones cambian de posicin:
Un segundo monmero propileno se ha
agregado a la cadena. Obsrvese que se
obtiene un polmero isotctico; esto se debe a
que el monmero propileno siempre se acerca
al catalizador con su grupo metilo dirigido en el
sentido opuesto del ligando indenilo.
Si el grupo metilo se dirigiera hacia el
ligando indenilo, ambos se toparan, evitando
que el propileno se acercara lo suficiente al
zirconio para formar el complejo. De modo que
slo cuando el metilo apunta en sentido
contrario respecto al ligando indenilo, el
propileno puede acomplejarse con el zirconio.
Cuando se agrega el segundo
monmero, ste debe aproximarse desde el
otro lado y tambin sus grupos metilo deben
apuntar en sentido contrario a los anillos
indenilo. Esto significa que el grupo metilo
apunta hacia arriba en lugar de hacerlo hacia
abajo. Dado que el segundo propileno se
adiciona desde el lado opuesto al primero,
debe dirigirse en direccin opuesta para que
los grupos metilo terminen del mismo lado de
la cadena polimrica.
La cadena polimrica termina cuando un enlace Zr H se forma antes de la adicin de
otra molcula olefnica:
Los catalizadores de zirconoceno/MAO son entre 10 y 100 veces ms efectivos que
los catalizadores de Ziegler para la polimerizacin de eteno. Cada tomo de Zr forma unas
46.000 cadenas polimricas por hora. Los clculos indican que una unidad de eteno es
insertada en la cadena cada 3 x 10
-5
segundos, una velocidad de reaccin muy elevada.
F.C.A.I. U.N. CUYO
Catlisis Homognea 44
6. Reacciones Catalticas especiales
6.1. AUTOCATLISIS
Una forma de catlisis en la que uno de los productos de la reaccin sirve como
catalizador para la reaccin.
La reaccin ms sencilla es: A+R R+R
Proceso mediante el cual un compuesto qumico induce y controla una reaccin
qumica sobre s mismo. Los compuestos autocatalticos no son catalizadores en
sentido estricto ya que su estructura qumica resulta alterada durante el proceso. No
obstante el compuesto no se destruye pudiendo mantener sus propiedades
autocatalticas.
Autocatlisis no debe confundirse con multiplicacin a travs de produccin
incluyendo la propia reproduccin en la cual el crecimiento de una poblacin es una
propiedad agregada de la capacidad productiva de sus miembros individuales. En la
autocatlisis, las propiedades sirven como sus propios catalizadores.
Un conjunto de reacciones qumicas pueden ser llamadas colectivamente
autocatalticas si un nmero de esas reacciones producen, como productos de reaccin, la
catlisis suficiente para las otras reacciones que forman el conjunto, es autosostenida dando
un extra de energa y molculas de alimento. Un conjunto autocataltico es una coleccin de
entidades, cada una de las cuales es capaz de catalizar la creacin de otras dentro del
conjunto, formando un todo, el conjunto es capaz de catalizar su propia rplica. De esta
forma el conjunto es un todo, el cual se dice que es cataltico. Los conjuntos autocatalticos
fueron originalmente, los ms concretos definidos en trminos de entidades los cuales
replican molculas.
Tipos de autocatalisis
Uno de los tipos de autocatlisis ms importantes es la catlisis biolgica o
enzimtica. Las enzimas, se encuentran entre los catalizadores ms importantes y tienen
una funcin esencial en los organismos vivos donde aceleran reacciones que de otra forma
requeriran temperaturas que podran destruir la mayora de la materia orgnica. La pepsina,
la tripsina y otras enzimas poseen la propiedad peculiar denominada autocatlisis que les
permite originar su propia formacin a partir de un precursor inerte denominado cimgeno.
Como consecuencia, estas enzimas se pueden reproducir en un tubo de ensayo. Las
enzimas son muy eficaces. Cantidades pequeas de una enzima pueden realizar a bajas
temperaturas lo que podra requerir reactivos violentos y altas temperaturas con mtodos
qumicos ordinarios. Por ejemplo, unos 30g de pepsina cristalina pura son capaces de
digerir casi dos toneladas mtricas de clara de huevo en pocas horas. Watson y Crick,
bilogos, consideraron un conjunto autocataltico a la forma en que funciona el metabolismo
en principio; por ejemplo, una protena ayuda a sintetizar otra protena y as siguiendo.
Luego del descubrimiento de la doble hlice, el dogma central del mapa gentico fue
formulado, en el cual a ADN es trascripto al ARN el cual es traducido a protena.
As tambin, varios modelos del origen de la vida estn basados en el desarrollo
molecular de un conjunto autocataltico. Muchos de estos modelos que han emergido de
estudios de sistemas complejos predicen que la vida surgi no de una molcula en particular
sino de un conjunto autocataltico, sin embargo no todas las molculas son capaces de
autoreproducirse.
Mecanismo de reaccin
A= B+C
El esquema ms simple para una reaccin autocatalizada es:
A+B---> B+B+C
Al cabo de un tiempo t la expresin de la velocidad es.
Dx/dt= k1 (a-x) x
Puede observarse que la reaccin presentar un mximo para x=a/2. Sin embargo, si
no hay absolutamente nada de B en el momento inicial, x=0 y la velocidad ser cero, a
F.C.A.I. U.N. CUYO
Catlisis Homognea 45
menos que se tenga una pequea cantidad de B por otro procedimiento, la reaccin no
puede ocurrir.
Suponemos una cantidad finita de B, representada como b, la ecuacin de la
velocidad se transforma en:
Dx/dt= k1(a-x)(b+x)
Integrando por partes se obtiene:
Ln a(b+x)= k1t
a+b b(a-x)
Las consecuencias de la ecuacin en la concentracin de A se muestra en las curvas
sigmoideas caractersticas de las reacciones autocatalizadas. Se ilustra tambin que b
influye en el tiempo necesario para alcanzar la velocidad mxima aunque no lo hace con la
curva.
Variacin de la concentracin de A con el tiempo para distintos valores de b,
manteniendo constantes a y k1.
Reacciones Autocataliticas
Las reacciones autocatalticas proceden lentamente al comienzo porque hay poco
producto presente, la tasa de reaccin aumenta progresivamente mientras la reaccin
avanza y luego disminuye nuevamente mientras las concentraciones de reactantes
disminuyen.
En una reaccin autocataltica ha de existir inicialmente algo de producto R para que
transcurra la reaccin. Empezando con una concentracin pequea de R, vemos
cualitativamente que la velocidad aumentar a medida que se vaya formando R. En el otro
extremo, cuando ha desaparecido prcticamente todo el componente A, la velocidad ha de
tender a cero. Este comportamiento se indica en la Fig. A1, la cual muestra que la velocidad
vara a lo largo de una parbola cuyo mximo corresponde a concentraciones iguales de A y
de R.
Flg. A1. Curvas conversin-tiempo y concentracin-velocidad pare reacciones
autocataliticas de la ecuacin
R A
A
C KC
dt
dC
ra = = .
Ejemplos
Tin pest es un ejemplo de una transformacin alotrpica, el cual causa el deterioro de
los objetos de lata a bajas temperaturas. Este fenmeno tambin ha sido llamado tin
disease, or tin leprosy (Lpre d'etain). Fue largamente observado en los tubos de los
F.C.A.I. U.N. CUYO
Catlisis Homognea 46
rganos en las catedrales medievales de Europa en climas fros. Lo que los miembros de
estas iglesias notaron fue que tan pronto como el metal comenzaba a descomponerse el
proceso se aceleraba, pareca autoalimentarse y continuaba inclusive a altas temperaturas.
A 13.2C o menos, la lata pura tena el hbito de destruirse desde el altropo beta (blanco)
al altropo alfa (gris). Eventualmente se descompona a polvo, de ah su nombre.
Lo que suceda es que la descomposicin se catalizaba a s misma, ese es el porqu
la reaccin pareca dispararse una vez comenzada, la mera presencia de tin pest conduca a
ms tin pest. Los objetos de lata a bajas temperaturas simplemente se desintegraban.
Reaccin de Permanganato con cido Oxlico
Esta reaccin es muy familiar para los estudiantes, donde se usan valoraciones con
autoindicador, realizndose luego de calentar la solucin a 70C. Es frecuente que despus
de la primera adicin de permanganato de la bureta se tenga la impresin de haberse
pasado del punto de equilibrio, pero en un minuto el color rosa desaparece y el
permanganato de las siguientes adiciones reacciona muy rpidamente. La reaccin est
catalizada por uno de los productos, el in Mn2+. En su ausencia la reaccin inicial
transcurre lentamente.
La adicin de unos pocos cristales de MnSO
4
acelera notablemente la reduccin por
cido oxlico de permanganato violeta a manganeso (II) el cual es incoloro. Cuando no hay
in manganeso II agregado, la reaccin procede inicialmente lentamente, pero gradualmente
se hace rpida debido a que el producto autocataliza la reaccin. El intermediario rojo es
[Mn (C
2
O
4
)
3
]
3-
. La ecuacin para la oxidacin de oxalato en solucin cida de permanganato
es:
2 MnO
4
-
(aq ) + 5 H
2
C
2
O
4
(aq ) + 6 H
3
O
+
(aq ) --> 2Mn
2+
(aq ) + 10 CO
2
(aq ) + 14 H
2
O
En un beaker de 150ml se coloc 75ml de disolucin de cido oxlico y 5 ml de
disolucin de cido sulfrico, el color de la disolucin es muy clara como transparente, y el
beaker se puso caliente. Se agreg 25 ml de disolucin de permanganato, y la disolucin se
puso muy oscura casi negra, en un tiempo de 1.38 seg, volvi a hacer la disolucin clara
como la del inicio. Se repiti el experimento, pero en este caso el tiempo fue 1.30 seg. Se
repiti de nuevo el experimento pero en lugar de aadir permanganato de potasio, se aadi
2ml de Mn2 +, y posteriormente el KMnO4, el color oscuro que caracterizaba esta disolucin
desapareci en 1.10 seg, se repiti el experimento y el tiempo en que duro la disolucin en
aclararse fue en 1.15 seg.
Anlisis
Las reacciones que se hicieron en el laboratorio son formas de autocatlisis, ya que
un mismo producto de la reaccin cataliza a esta, en la disolucin de cido oxlico y cido
sulfrico su temperatura aumenta debido a que son dos cidos fuertes y liberan energa en
forma de calor, caracterizndose as como una reaccin exotrmica por lo tanto una
reaccin espontnea, al agregarle el permanganato la disolucin se volvi oscura como
negra, porque el permanganato tiene como caracterstica fsica un color morado intenso,
este le da un aporte de energa a la reaccin de cido oxlico y cido sulfrico, y dicha
disolucin se incolora en 1.38 seg., lo que quiere decir que la reaccin se complet.
Todos las disolucin se hacen en fase liquida, por consiguiente son catlisis
homognea, ya que los reactivos y el catalizador estn en esta fase, en una disolucin de
cido oxlico, cido sulfrico se agreg Mn2 + y permanganato, la reaccin se aclaro en un
tiempo mas corto que la disolucin que no se le agreg Mn2 +, debido a que el Mn2 + es un
catalizador y hace que la reaccin se complete mas rpido.
Reaccin Redox:
MnO
4
+ 5C
2
O
4
H
2
+ 8H
2
O + ------------ Mn
2
+ + 10CO
2
+ 12H
2
O
Equilibrada:
2MnO
4
+ 5C
2
O
4
H
2
+ 16H
2
O + --------------2Mn
2
+ + 10CO
2
+ 24H
2
0
Autocatlisis Biolgica: Antes del surgimiento de las enzimas con base proteica, las
cuales son el principio de las catlisis biolgicas hoy en da, se crea que alguna sustancia,
tal como el ARN poda servir como un reproductor y autocatalizador.
En un principio, las reacciones de autosntesis de biomolculas dependan totalmente
de la radiacin emitida por el sol, sobre todo de la luz, calor y radiacin gamma.
F.C.A.I. U.N. CUYO
Catlisis Homognea 47
Hidrlisis de esteres: la hidrlisis de un ester produce un alcohol y un cido orgnico
en medio cido. Una vez que se form una porcin de producto, es el cido orgnico el que
permite continuar con la reaccin, catalizando la hidrlisis.
Fermentacin: Los ejemplos ms importantes de reacciones autocatalticas son las
reacciones de fermentacin, debidas a la accin de un microorganismo sobre una
alimentacin orgnica. Frecuentemente varios microorganismos compiten para dar
diferentes productos. Otro tipo de reaccin que tiene comportamiento autocataltico es la
reaccin exotrmica (la combustin de un gas) que se efecta adiabticamente entrando al
sistema los reactantes a una temperatura demasiado baja para la combustin. En una
reaccin de este tipo, denominada autotrmica, el calor puede considerarse como el
producto que mantiene la reaccin.
As, con flujo en pistn la reaccin se detiene mientras que con flujo en mezcla
completa la reaccin se automantiene debido al calor generado que eleva la temperatura de
los reactantes hasta que puedan reaccionar. Las reacciones autotrmicas son de gran
importancia en los sistemas en fase gaseosa catalizados por slidos, y se estudian mas
adelante.
Diseo aplicado en reacciones autocatalticas
Cuando un reactante desaparece de acuerdo con una ecuacin cintica de primero o
segundo orden en un reactor discontinuo, al principio su velocidad de desaparicin es rpida
ya que la concentracin del reactante es elevada, y despus disminuye progresivamente a
medida que el reactante se va consumiendo.
Sin embargo, en una reaccin autocataltica, al principio la velocidad es pequea
debido a que hay poco producto presente, aumenta hasta un valor mximo a medida que se
va formando producto, y despus desciende nuevamente hasta un valor bajo a medida que
el reactante se consume. En la Fig. A2 se representa el comportamiento caracterstico.
Fig. A2. Curva -caracterstica velocidad-concentracin para reacciones autocataliticas,
Las reacciones que presentan este tipo de curvas velocidad-concentracin dan lugar a
interesantes problemas de optimizacin, a la vez que proporcionan un buen ejemplo del
mtodo general de diseo. Por estas razones vamos a estudiarlas con ms detalle,
considerando exclusivamente las curvas 1 /(-ra) frente a X, con su mnimo caracterstico,
como se muestra en la Fig.A2.
F.C.A.I. U.N. CUYO
Catlisis Homognea 48
Fig. A3. Para las reacciones autocataltlcas. El flujo en mezcla completa es ms eficaz
para conversiones bajas y el flujo en pistn para conversiones altas.
Comparacin del reactor de flujo en pistn con el de mezcla completa, sin
recirculacin.
Para cualquier curva velocidad-conversin, la comparacin de las reas de la Fig. A3
muestra cul es el reactor ms adecuado (el que tiene un volumen ms pequeo) para un
fin determinado. Se encuentra que:
1. Para conversiones bajas .el reactor de mezcla completa resulta ms adecuado que
el reactor de flujo en pistn.
2. Para conversiones suficientemente altas el reactor de flujo en pistn es el ms
adecuado.
Estas conclusiones difieren de las que hemos encontrado para las reacciones
ordinarias de orden n (n > 0) para las cuales el reactor de flujo en pistn era siempre ms
eficaz que el reactor de mezcla completa. Hemos de indicar tambin que, como inicialmente
ha de estar presente algn producto en la alimentacin para que se efecte la reaccin
autocataltica, no podra operar un reactor de flujo en pistn con una alimentacin de
reactante puro; en tal caso habra que aadirle continuamente a la alimentacin algn
producto, presentndose as una buena oportunidad para emplear un reactor con
recirculacin.
Reactor con recirculacin. Cuando se trata una sustancia en un reactor con
recirculacin para obtener una conversin final conocida XA,, se puede pensar que ha de
existir una relacin de recirculacin determinada que sea ptima y que haga mnimo el
volumen del reactor o del tiempo espacial.
Sistema de reactores mltiples. Para que el volumen total sea mnimo, hemos de
elegir un sistema en el cual la mayor parte del proceso se efecte con la composicin
correspondiente al punto de velocidad mxima, o cerca de el. Utilizando un reactor de
mezcla completa, este punto puede alcanzarse directamente sin pasar por las
composiciones intermedias de velocidades ms bajas. Si queremos alcanzar conversiones
ms elevadas hemos de desplazarnos progresivamente desde esta composicin hasta la
composicin final; esto requiere un reactor de flujo en pistn.
En consecuencia, para que la conversin de una sustancia sea mayor que la
correspondiente a la del punto de mxima velocidad, el mejor dispositivo ser un reactor de
mezcla completa que opere a la velocidad mxima, seguido de un reactor de flujo en pistn,
como se indica en la Fig. A4,
Este dispositivo es mejor que un solo reactor de flujo en pistn, un solo reactor de
mezcla completa, o un reactor con recirculacin.
Reactores con separacin y recirculacin del reactante no convertido. Si el
reactante no convertido puede separarse de la corriente de producto y retornar al reactor,
todo el proceso puede efectuarse a una composicin para cualquier grado de conversin. En
este caso, el mejor procedimiento es utilizar un reactor de mezcla completa que opere con la
composicin del punto de mxima velocidad de reaccin, como se indica en la Fig. A4.
El volumen del reactor es ahora el mnimo absoluto, menor que cualquiera de los
sistemas estudiados anteriormente. Sin embargo, la economa global, incluyendo el coste de
separacin y recirculacin, determinar cual es el dispositivo ptimo global.
F.C.A.I. U.N. CUYO
Catlisis Homognea 49
Fig. A4. a) La mejor disposicin de reactor mltiple, y b) la mejor disposicin cuando el
reactante no convertido puede ser separado y recirculado
Ejemplo - Flujo en pistn y en mezcla completa para una reaccin autocataltica
Se han de analizar varias disposiciones de reactores para la transformacin de A en R. La
alimentacin contiene 99 % de A y 1% de R; el producto ha de constar de 10 % de A y 90 %
de R.
La transformacin se efecta por medio de la reaccin elemental A+RR+R de coeficiente
cintico k = 1 litro/(mol)-(min). La concentracin de las sustancias activas es:
C
AO
+ C
RO
= C
A
+ C
R
= C
O
= 1 mol/litro,
Desde el principio hasta el fin.
Cul es el tiempo de permanencia en el reactor que conducir a un producto en el cual C
R
=
0,9 mol / litro? a) en un reactor de flujo en pistn, b) en un reactor de mezcla completa, y c)
en el sistema de tamao mnimo, sin recirculacin.
Si la corriente de salida del reactor es ms rica en A que la deseada, supngase que puede
separarse en dos corrientes, una que contiene el producto deseado (90 % de R y 10 % de
A) y otra que contiene 1 mol/litro de componente A puro. Despus la corriente de A puno se
hace recircular.
d) En estas condicionas, calclese el tiempo de permanencia mnimo si el reactor funciona
en las condiciones de mxima eficacia.
F.C.A.I. U.N. CUYO
Catlisis Homognea 50
Solucin. Aunque la expresin cintica de esta reaccin es lo suficientemente sencilla para
que pueda resolverse analticamente, utilizaremos el procedimiento grfico general
desarrollado en este capitulo. Por otra parte, es ms conveniente utilizar concentraciones en
lugar de conversiones.
En la Tabla 6-E6 se dan los datos necesarios para construir la curva de 1/(- r
A
) frente a C
A
, y
en la Fig. 6-E6, no construida a escala, se indica como se obtienen los correspondientes
valores, midiendo en cada caso el rea comprendida bajo la curva 1/ (- r
A
) frente a C
A
y las
ordenadas extremas.
Para un producto en el que C
A
= 0,1 mol/litro y C
R
= 0,9 mol/litro, de la Fig. 6-E6 tenemos:
a) Para flujo en pistn: = 6,8 min.
b) Para flujo en mezcla completa = 9,9 min.
TABLA GE6
C
A
C
B
-r
A
1/-r
A
0.99 0.01 0,0099 101.01
0.95 0,05 0.0475 21.05
0.90 0.10 0.09 11.11
0.70 0,30 0.21 4.76
0.50 0,50 0.25 4.00
0,30 0,70 0.21 4.76
0.10 0.90 0.09 11.11
c) Para el dispositivo reactor ms eficaz, sin recirculacin, necesitamos un reactor de mezcla
completa
(T = 2,0 min.) Seguido de un reactor de flujo en pistn (T = 2,2 min.), con tiempo espacial
total:
T
m+p
= 4,2min
d) Con separacin de producto y recirculacin, el dispositivo ms eficaz tiene:
Tmin = 2.0 min.
7. Bibliografa
- CHEMICAL KINETICS (2 Edicin en Ingls) Keith J. Laidler (1965) McGraw-Hill
- Pginas 450- 456
- -CHEMICAL KINETICS AND REACTION MECHANISMS James H. Espenson (1981)
McGraw-Hill Pginas 198-199
- QUMICA FSICA Daz Pea y Roig. Alhambra. Pginas 1133-138
- http://mazinger.sisib.uchile.cl/repositorio/ap/ciencias_quimicas_y_farmaceuticas/ap-
fisquim-farm11/c17.3.html
- http://omega.ilce.edu.mx:3000/sites/ciencia/volumen2/ciencia3/059/htm/sec_6.htm
- http://www.uam.es/personal_pdi/ciencias/juansqui/catalisisweb.htm
- LOGAN, S.R. Fundamentos de Cintica Cumica. Ed. Addison Wesley. 2000.
- GUERASIMOV, YA. Y OTROS, Curso de Qumica Fsica. Ed.MIR 1977
- AVERY, H.E, Cintica Qumica Bsica y Mecanismos de Reaccin. Ed. Revert. 1997
- BREWSTER, R. Q; McEWEN, W.E., Qumica Orgnica, Ed. Prentice Hall, 1996.
- MORRISON, R. T.; BOYD, R. N.. Qumica Orgnica. Ed: Addison Wesley.
- WALAS, S.N. Cintica de las reacciones qumicas. Agular 1965
- Internet: www. monografas.com y otras.
- www.sinorg.uji.es
- QUMICA ORGNICA (3 EDICIN), R.T. MORRISON Y R.N. BOYD (1978), ALLYN AND
BACON, INC.
- Qumica Orgnica (3 Edicin), FOX, WHITESEN .
- LOUIS AND MARY FIESER. PG.949-951 JABONES TEORIA
- HENRY RAKOFF PG.281 HIDRILISIS DE ESTERES
- BREWTER Y MC EWEN PG.282 PROPIEDADES QUIMICAS DE LAS GRASAS
- NOLLER. PG.152
- CIENCIA EN ESPAOL.HTM
F.C.A.I. U.N. CUYO
Catlisis Homognea 51
- Cintica de reacciones; Steanley Walas
- Cintica qumica bsica y mecanismos de reaccin; H.E. Avery
- Chang, Raymond; Qumica General
- http://www.jmarcano.com/recursos/contamin/catmosf2c.html
- http://ciencia.nasa.gov/headlines/y2000/ast12dec_1.htm
- http://html.rincondelvago.com/contaminacion-atmosferica.html
- http://www.ciencia-hoy.retina.ar/hoy36/clorofl1.htm
- www.omega.ilce.edu.mx:3000/sites/ ciencia/volumen2/ciencia3/059/htm/sec_5.htm
- http://www.elementos.buap.mx/nu
- P Gasesa y J. Hubble, tecnologa de las enzimas, Editorial acribis. S.A., Segunda Edicin,
Zaragoza Espaa, 1990.
- Perry, Manual del Ingeniero Qumico, sexta Edicin,Tomo VI, Editorial McGraw-Hill, 1992.
- Kirk, Raymond, Enciclopedia de Tecnologa Qumica, Tomo VI, primera Edicin, Editorial
Uteha, Espaa 1962.
- Ocano, Enciclopedia de la Ciencia y La tecnologa Tomo V, ediciones Ocano, Barcelona
1980.
- http:://www.icp.csis.es/biocatlisis/web3lineas.html
- http:www.mty.itesm.mx/data/cd/html
- http//www.unav.es/memoria/8-99/ingenieria.html
- GUERASIMOV, V y Otros: Curso De Qumica Fsica, Ed. Mir, 2 ed. 1977
- Internet: www.chemfinder.com
- www.chemweb.com
- www.spectragalactic.com
- www.probes.com/handbook
- www.barguell.com/.../Proyectos%20Curriculares/ Bachillerato /BAC%20Biologia %202%
BA.doc - Resultado Suplementario
- www.fi.uba.ar/materias/6304/planificacion.pdf
- Raymond Chang Qumica Gral. Kirk-Othmer Enciclopedia de tecnologa Qumica.
- Cotton y wilkinson Qumica Inorgnica y Avanzada.
- www.ehu.es/biomoleculas/ENZ/ENZ2.htm
- Enciclopedia Kirk tomo 16
- www.wikipedia.com
- Qumica Organica Morrison y Boyd
- Qumica Orgnico Fox Whitellsen
- http://www.udc.es/dep/qf/Qo/PrOrg.htm.
- Raymond Chang. Qumica sptima edicin. Mc Graw Hill. Capitulo 18.
- https://www.me.gov.ve/SegundaEtapa/Glosario/cienciasnaturaleza.htm
- Levenspiel: Ingeniera de las reacciones qumicas-Ed Reverte 1974
- etbiochem.com
You might also like
- Unidad V.catalisis en Sistemas Homogeneos y HeterogeneosDocument19 pagesUnidad V.catalisis en Sistemas Homogeneos y Heterogeneosgreey escorciaNo ratings yet
- CATALISIS (Final2)Document28 pagesCATALISIS (Final2)Liliana LiliNo ratings yet
- CatalizadorDocument23 pagesCatalizadoralfredo pcNo ratings yet
- Unidad 6.1 - Cinética Enzimática-PresentaciónDocument74 pagesUnidad 6.1 - Cinética Enzimática-PresentaciónCamila EscobarNo ratings yet
- CatalizadoresDocument7 pagesCatalizadoresJOSU� EMMANUEL HAM HERN�NDEZ100% (1)
- CatalisisDocument21 pagesCatalisisBeatriz EchevarriaNo ratings yet
- Nutricion y Metabolismo CelularDocument31 pagesNutricion y Metabolismo CelularCarlos Amaranto0% (1)
- Maria Veronica FisicoquimicaDocument16 pagesMaria Veronica FisicoquimicaBrenda araujooNo ratings yet
- CATÁLISISDocument6 pagesCATÁLISISAdrian GarciaNo ratings yet
- 2º Biología BachilleratoDocument6 pages2º Biología BachilleratoJorge MéndezNo ratings yet
- Catalisis HomogeneaDocument36 pagesCatalisis HomogeneaJulia PeñaNo ratings yet
- EnzimasDocument11 pagesEnzimasvanesaNo ratings yet
- Enzimas WordDocument14 pagesEnzimas WordYaz ReyesNo ratings yet
- Ordenado Tema Del Primer SeminarioDocument22 pagesOrdenado Tema Del Primer SeminarioMadheleyn del Carmen Alvarez ArmasNo ratings yet
- CatalizadoresDocument22 pagesCatalizadoressofiaNo ratings yet
- Cinética de Las ReaccionesDocument6 pagesCinética de Las ReaccionesNazarith RodríguezNo ratings yet
- Catalisis 9CDocument21 pagesCatalisis 9CfernandoNo ratings yet
- UntitledDocument3 pagesUntitledstefy galoNo ratings yet
- Practica No. 3. Actividad Enzimatica - AmilasaDocument12 pagesPractica No. 3. Actividad Enzimatica - AmilasanicoleNo ratings yet
- ENZIMASDocument46 pagesENZIMASVane Rubii CamposNo ratings yet
- Catalizadores No BiologicosDocument9 pagesCatalizadores No BiologicosYuritziNo ratings yet
- LAB3 QMCDocument5 pagesLAB3 QMCNikol Lopez ValdezNo ratings yet
- Catalisis Heterogenea y Cinetica Electrodica (Resumen)Document4 pagesCatalisis Heterogenea y Cinetica Electrodica (Resumen)Fabio Alejandro Rodriguez RaudaNo ratings yet
- Transparencias Tema 05Document36 pagesTransparencias Tema 05Alberto José Martín SánchezNo ratings yet
- TEMA 4bioqDocument9 pagesTEMA 4bioqAlba Montes FernandezNo ratings yet
- Guia Catalisis EmilioDocument29 pagesGuia Catalisis EmilioOscar SiviraNo ratings yet
- Catalizador HomogéneoDocument4 pagesCatalizador HomogéneoJGerardo D Hernández100% (1)
- Marco Teorico)Document24 pagesMarco Teorico)Jhoan BorhorquezNo ratings yet
- EnzimasDocument73 pagesEnzimasAnonymous BXO5lCb5x4No ratings yet
- Que Es Un CatalizadorDocument5 pagesQue Es Un CatalizadorRonaldNo ratings yet
- Bloque IIDocument54 pagesBloque IImaritaviNo ratings yet
- Aspectos Generales y Propiedades de Las EnzimasDocument37 pagesAspectos Generales y Propiedades de Las Enzimasronald stevn alvarez osorioNo ratings yet
- Bioquímica Ambiental - Clase7 PDFDocument19 pagesBioquímica Ambiental - Clase7 PDFMelany FranciaNo ratings yet
- Apuntes Guia TP2 2022Document8 pagesApuntes Guia TP2 2022raul roblesNo ratings yet
- Unidad 4 Fisicoquimica 2Document32 pagesUnidad 4 Fisicoquimica 2Adán Fonseca HernandezNo ratings yet
- Unidad IV. Investigación FisicoquímicaiiDocument32 pagesUnidad IV. Investigación FisicoquímicaiiCarlos FernandoNo ratings yet
- EnzimasDocument8 pagesEnzimasrkhgn7khjvNo ratings yet
- Taller de Repaso de EnzimasDocument8 pagesTaller de Repaso de EnzimasDani CardonaNo ratings yet
- Taller de Repaso de EnzimasDocument8 pagesTaller de Repaso de EnzimasDani CardonaNo ratings yet
- CatalisisDocument19 pagesCatalisisEduardo OcelotlNo ratings yet
- Catalizador - EcuRed PDFDocument3 pagesCatalizador - EcuRed PDFYori Edison Vasquez SalamancaNo ratings yet
- Bioquímica Unidad 2 Parte IDocument63 pagesBioquímica Unidad 2 Parte IDarisber Vanessa Davis RNo ratings yet
- Practica de Catalisis (Investigacion)Document4 pagesPractica de Catalisis (Investigacion)Jas AcostaNo ratings yet
- Unidad 4 Fisicoquimica2Document56 pagesUnidad 4 Fisicoquimica2Martha C. ArroyoNo ratings yet
- Catalisis AmbientalDocument13 pagesCatalisis AmbientalJhonatan MalaraNo ratings yet
- CATALISISDocument15 pagesCATALISISAndrés Santa CruzNo ratings yet
- Bioquimica InformeDocument5 pagesBioquimica InformeAlexia BernalesNo ratings yet
- Resumen de CatalisisDocument5 pagesResumen de CatalisisLUIS ANGEL VAZQUEZ VALDOVINOSNo ratings yet
- EnzimasDocument29 pagesEnzimasLuis MarquinezNo ratings yet
- EnzimasDocument23 pagesEnzimasMaruby MontonerNo ratings yet
- Información BiologiaDocument3 pagesInformación BiologiaEmerson Tiago Camacho DíazNo ratings yet
- Catalizadores No BiológicosDocument9 pagesCatalizadores No BiológicosYuritziNo ratings yet
- Catalisis BifasicaDocument8 pagesCatalisis BifasicaJuan David Lopez OsorioNo ratings yet
- Examen de BioquímicaDocument3 pagesExamen de BioquímicaAndy DuarteNo ratings yet
- Presentación Catálisis - Fernandez BernalDocument14 pagesPresentación Catálisis - Fernandez BernalAldair fernandezNo ratings yet
- Tema 7 BioqDocument10 pagesTema 7 BioqAlexandra María Marín BernalNo ratings yet
- Investigacion CatalizadoresDocument5 pagesInvestigacion CatalizadoresLeslie Priscila VelNo ratings yet
- Actividad Enzimática de La CatalasaDocument10 pagesActividad Enzimática de La Catalasaalfredo andaluzNo ratings yet
- Diplomatura en Gestion Calidad Alimentos Bromato 2017Document12 pagesDiplomatura en Gestion Calidad Alimentos Bromato 2017Cristina ClavijoNo ratings yet
- Pe 2021Document2 pagesPe 2021Cristina ClavijoNo ratings yet
- Drama NavideñoDocument7 pagesDrama NavideñoCristina ClavijoNo ratings yet
- Drama NavideñoDocument7 pagesDrama NavideñoCristina ClavijoNo ratings yet
- Sermon Ministerio Del Niño Sabado 14 de Diciembre 2019Document1 pageSermon Ministerio Del Niño Sabado 14 de Diciembre 2019Cristina ClavijoNo ratings yet
- Ejercicio de Flujo CompresibleDocument1 pageEjercicio de Flujo CompresibleCristina ClavijoNo ratings yet
- Cuadernillo Quimica UncuyoDocument113 pagesCuadernillo Quimica UncuyoCristina Clavijo100% (1)
- Análisis de Redes Hidráulicas NEWDocument26 pagesAnálisis de Redes Hidráulicas NEWCristina ClavijoNo ratings yet
- Hornos para MineralurgiaDocument9 pagesHornos para MineralurgiaCristina ClavijoNo ratings yet
- Cantera de Aridos Don RamonDocument4 pagesCantera de Aridos Don RamonCristina ClavijoNo ratings yet
- Act 3 Unidad Vi Serrano Veliz PeriniDocument5 pagesAct 3 Unidad Vi Serrano Veliz PeriniCristina ClavijoNo ratings yet
- Paritaria Universidad 2014Document12 pagesParitaria Universidad 2014Cristina ClavijoNo ratings yet
- Diplomado en Petroleo y GasDocument6 pagesDiplomado en Petroleo y GasCristina ClavijoNo ratings yet
- IIA Cantera Arenera BecarDocument39 pagesIIA Cantera Arenera BecarCristina Clavijo100% (1)
- UD AromaticosDocument2 pagesUD AromaticosCristina ClavijoNo ratings yet
- Estequiometria PDFDocument16 pagesEstequiometria PDFHerlan FloresNo ratings yet
- Auxiliar ES - 2018-3TDocument66 pagesAuxiliar ES - 2018-3Tjsolisadv9289No ratings yet
- Señales de Los TiemposDocument15 pagesSeñales de Los TiemposCristina ClavijoNo ratings yet
- Misionero Infantes JennyDocument21 pagesMisionero Infantes JennyCristina ClavijoNo ratings yet
- Calendario 2019Document31 pagesCalendario 2019Cristina ClavijoNo ratings yet
- Catálisis Homogénea UT 1Document60 pagesCatálisis Homogénea UT 1Cristina ClavijoNo ratings yet
- El Ultimo Viaje de ValentinaDocument321 pagesEl Ultimo Viaje de ValentinaCristina ClavijoNo ratings yet
- AutosDocument244 pagesAutosmmcode13No ratings yet
- Canfield Jack - Chocolate Caliente para El AlmaDocument352 pagesCanfield Jack - Chocolate Caliente para El Almaamigoandres2001No ratings yet
- 2013-SIEF XI Memorias (Final)Document616 pages2013-SIEF XI Memorias (Final)Cristina ClavijoNo ratings yet
- Sermon 5c2ba Sabado4Document7 pagesSermon 5c2ba Sabado4Cristina ClavijoNo ratings yet
- Fluidos Ebook - Flujo Entre Placas ParalelasDocument2 pagesFluidos Ebook - Flujo Entre Placas ParalelasCristina ClavijoNo ratings yet
- Mecánica de Fluídos 2 - Wendor Chereque MoránDocument246 pagesMecánica de Fluídos 2 - Wendor Chereque Moránjohannitah100% (1)
- Modulo09 - U2 Flujo LaminarDocument12 pagesModulo09 - U2 Flujo LaminarmasterjackersNo ratings yet
- PRACT N°3 FACŢORES QUE AFECTAN LA ACTIVIDAD ENZIMATICADocument11 pagesPRACT N°3 FACŢORES QUE AFECTAN LA ACTIVIDAD ENZIMATICAKim Hyun JoongNo ratings yet
- Reporte 7 CineticaDocument13 pagesReporte 7 CineticaGiovanni100% (2)
- Proceso de Obtencion Butadieno (Informe)Document84 pagesProceso de Obtencion Butadieno (Informe)KevinSuarezHuanasca100% (1)
- Síntesis de antibióticos β-lactámicos semisintéticos mediante la penicilina acilasa.Document3 pagesSíntesis de antibióticos β-lactámicos semisintéticos mediante la penicilina acilasa.Cecyteg Gto FisicaNo ratings yet
- El Dimetil ÉterDocument9 pagesEl Dimetil ÉterLuis CchcNo ratings yet
- EnzimasDocument8 pagesEnzimasPao MoralesNo ratings yet
- Informe de AluminaDocument15 pagesInforme de AluminaCarlos Cardenas VilcaNo ratings yet
- Una Revisión Sobre La Pirólisis de Residuos Plásticos.Document65 pagesUna Revisión Sobre La Pirólisis de Residuos Plásticos.wilhianNo ratings yet
- Introduccion Al Diseño de ReactoresDocument16 pagesIntroduccion Al Diseño de ReactoresRosa HernandezNo ratings yet
- Ejercicios 2 ParcialDocument2 pagesEjercicios 2 ParcialAlinne VillegasNo ratings yet
- Nanoparticulas de Oro FaradayDocument8 pagesNanoparticulas de Oro FaradayLizeth Paola TellezNo ratings yet
- Fermentación Laboratorio 2Document12 pagesFermentación Laboratorio 2Henry Perez33% (3)
- Seminario Productos Derivados Del Benceno, Tolueno y XilenoDocument16 pagesSeminario Productos Derivados Del Benceno, Tolueno y XilenoBrendaNo ratings yet
- Informe Cinética EnzimáticaDocument3 pagesInforme Cinética EnzimáticaEli CortésNo ratings yet
- Practicas 2o ParcialDocument5 pagesPracticas 2o ParcialAngel CastellanosNo ratings yet
- Capítulo 4 PI-140 B Fenómenos de Transporte UNIDocument50 pagesCapítulo 4 PI-140 B Fenómenos de Transporte UNIJaire MoroteNo ratings yet
- Cuestionario 2Document2 pagesCuestionario 2Rubencito F FernandezNo ratings yet
- Enzimas 2020Document54 pagesEnzimas 2020catalinaNo ratings yet
- PruebaDocument99 pagesPruebaFranz Gustavo Vargas MamaniNo ratings yet
- Análisis de Anfetaminas PDFDocument75 pagesAnálisis de Anfetaminas PDFsmatalab100% (3)
- AIBNDocument61 pagesAIBNViridianaGarciaNo ratings yet
- Revision Bibliografica QuimicaDocument7 pagesRevision Bibliografica QuimicaItza Manuela Santos DiazNo ratings yet
- Tema 6 EstequiometriaDocument11 pagesTema 6 EstequiometriaLuisa Gallo Martínez100% (3)
- Práctica de Cultivo y Aislamiento de BacteriasDocument3 pagesPráctica de Cultivo y Aislamiento de BacteriasYARIT SALOME SANCHEZ MORENONo ratings yet
- TFG GIQuimicaDocument18 pagesTFG GIQuimicaMorelvys DiazNo ratings yet
- Trabajo de Grado Audreys SalazarDocument65 pagesTrabajo de Grado Audreys SalazarMichelle Carriel CiriloNo ratings yet
- 2.-Constante de Equilibrio Quimico de Un Sist HeterogeneoDocument8 pages2.-Constante de Equilibrio Quimico de Un Sist Heterogeneo.:("*"BLacK BuLLeT"*"):.94% (16)
- Ciencia y Tecnología ProblematicaDocument7 pagesCiencia y Tecnología ProblematicaYajaira MacadanNo ratings yet
- EnzimasDocument10 pagesEnzimasLeader Leyva HernandezNo ratings yet