You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- UPCAT Complete Reviewer 2020jjjjjjDocument84 pagesUPCAT Complete Reviewer 2020jjjjjjPolyblank Glitch100% (2)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- HACCP Plan With Flow Chart-1Document23 pagesHACCP Plan With Flow Chart-1Anonymous aZA07k8TXfNo ratings yet
- Itch Mechanisms and ManagementDocument464 pagesItch Mechanisms and ManagementGaurav JaswalNo ratings yet
- (2011) - Multiresistant Gram-Negative Bacteria - The Role of High-Risk Clones in The Dissemination of Antibiotic Resistance. Woodford Et. AlDocument20 pages(2011) - Multiresistant Gram-Negative Bacteria - The Role of High-Risk Clones in The Dissemination of Antibiotic Resistance. Woodford Et. AldlunitaNo ratings yet
- TGFβ signaling in health and diseaseDocument4 pagesTGFβ signaling in health and diseasedlunitaNo ratings yet
- (2010) - New Cucurbitane-Type Triterpenoids From Bryonia Aspera. Sahranavard S, Et Al. Planta Med 2010, 76 1014-1017Document4 pages(2010) - New Cucurbitane-Type Triterpenoids From Bryonia Aspera. Sahranavard S, Et Al. Planta Med 2010, 76 1014-1017dlunitaNo ratings yet
- Bose QC35Document28 pagesBose QC35dlunitaNo ratings yet
- FCQ 4 BQ EXP Proteina - ColagenoDocument1 pageFCQ 4 BQ EXP Proteina - ColagenodlunitaNo ratings yet
- Holism & Reductionism: Issues and DebatesDocument30 pagesHolism & Reductionism: Issues and Debatesh100% (1)
- Parameter Estimation With Term-Wise Decomposition in Biochemical Network GMA Models by Hybrid Regularized Least Squares-Particle Swarm OptimizationDocument8 pagesParameter Estimation With Term-Wise Decomposition in Biochemical Network GMA Models by Hybrid Regularized Least Squares-Particle Swarm OptimizationGek CagatanNo ratings yet
- Myoclonus With DementiaDocument8 pagesMyoclonus With DementiaSantosh DashNo ratings yet
- PK PD en Critical IllnessDocument27 pagesPK PD en Critical IllnessDaniela BolivarNo ratings yet
- Northern Blot Analysis For Detection of RNA in PanDocument8 pagesNorthern Blot Analysis For Detection of RNA in PanFati ZoraNo ratings yet
- PGA102 Presentation 1 (Eng. Ver.)Document18 pagesPGA102 Presentation 1 (Eng. Ver.)Lee Wai KeatNo ratings yet
- Adhesive Systems in Restorative DentistryDocument6 pagesAdhesive Systems in Restorative DentistryStoian SimonaNo ratings yet
- Class Notes 12 - Biology - Reproduction in OrganismDocument8 pagesClass Notes 12 - Biology - Reproduction in OrganismNehaNo ratings yet
- Pathophysiology of Primary Cataract: Saint Paul University Dumaguete College of Nursing S.Y. 2021-2022Document3 pagesPathophysiology of Primary Cataract: Saint Paul University Dumaguete College of Nursing S.Y. 2021-2022zoie ziazzetteNo ratings yet
- BRO Fluorescence SpectrosDocument24 pagesBRO Fluorescence SpectrosWaleed El-azabNo ratings yet
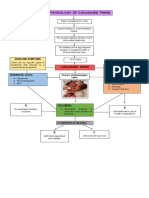
- Pathophysiology of Conjoined TwinsDocument1 pagePathophysiology of Conjoined TwinsPrincess Loreto AlegreNo ratings yet
- Milestone03 Chinonso Nwachukwu 28.05.2023Document18 pagesMilestone03 Chinonso Nwachukwu 28.05.2023Prince NwachukwuNo ratings yet
- Immobilization of Catalase Via Adsorption Into NatDocument6 pagesImmobilization of Catalase Via Adsorption Into NatErwin Guillermo Valdizón WinterNo ratings yet
- Characteristics of A Community PDFDocument15 pagesCharacteristics of A Community PDFAlijanNo ratings yet
- Drug Development ProcessDocument16 pagesDrug Development ProcessNachiket PatelNo ratings yet
- DEVPSY With HighlightsDocument45 pagesDEVPSY With HighlightsFiona ParasNo ratings yet
- Induction Presentation Biology A Level and Gcse.204719226Document25 pagesInduction Presentation Biology A Level and Gcse.204719226Cut TirayaNo ratings yet
- Boger Boenninghausen Characteristics RepertoryDocument12 pagesBoger Boenninghausen Characteristics RepertoryDrx Rustom AliNo ratings yet
- Activity No. 7.1 BloodDocument2 pagesActivity No. 7.1 BloodDree SermanNo ratings yet
- Summative Assessment QuizDocument3 pagesSummative Assessment Quizapi-379149339No ratings yet
- 088 11MS9419 Print PDFDocument11 pages088 11MS9419 Print PDFNehaNo ratings yet
- Astriana Final MRM Project Paper-RevisedDocument92 pagesAstriana Final MRM Project Paper-RevisedNgurah MahasviraNo ratings yet
- Water PotentialDocument35 pagesWater PotentialLuis GarcíaNo ratings yet
- Protein Synthesis ClozeDocument3 pagesProtein Synthesis Clozeapi-309893409No ratings yet
- Answers For Support Worksheet - Option F: 1 A Feature Bacteria Archaea EukaryotaDocument1 pageAnswers For Support Worksheet - Option F: 1 A Feature Bacteria Archaea EukaryotaHellö PëachNo ratings yet
- Beckers World of The Cell 9th Edition Hardin Test Bank DownloadDocument23 pagesBeckers World of The Cell 9th Edition Hardin Test Bank Downloaddaddockstudderyxeq100% (27)
- Gen Bio 2 Q2 Lesson 1.1 Plant Life CycleDocument3 pagesGen Bio 2 Q2 Lesson 1.1 Plant Life CycleRea EscotaNo ratings yet