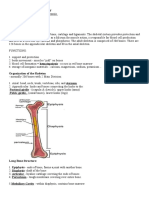
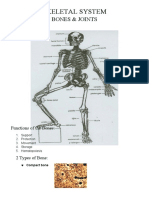
You might also like
- 08 Vertebra, Ribs, and SternaDocument3 pages08 Vertebra, Ribs, and SternaKaye Coleen Cantara Nuera100% (1)
- Vertebrate Systems SummaryDocument29 pagesVertebrate Systems SummaryHazel Grace Bellen100% (1)
- BIO 342 Comparative Vertebrate Anatomy Lecture Notes 2 - Vertebrate Skeletal Systems & SkullDocument76 pagesBIO 342 Comparative Vertebrate Anatomy Lecture Notes 2 - Vertebrate Skeletal Systems & SkullSteph VeeNo ratings yet
- Compa SkeletalDocument37 pagesCompa SkeletalJamie LiwanagNo ratings yet
- Veterinary OsteologyDocument279 pagesVeterinary OsteologyTatenda Mageja100% (2)
- Skeletal SystemDocument62 pagesSkeletal SystemAlyza AlcazarinNo ratings yet
- Skeletal SystemDocument52 pagesSkeletal SystemHarish musictryNo ratings yet
- Vertebral Column and Ribs PartsDocument22 pagesVertebral Column and Ribs PartsDaniele Talens CaguiatNo ratings yet
- Relationship To Other Organ SystemsDocument11 pagesRelationship To Other Organ SystemsVern NuquiNo ratings yet
- Osteology: The Study of BonesDocument67 pagesOsteology: The Study of BonesChen Romaquin Jimenez ThesisNo ratings yet
- REVISED Skeletal System of The Frog 2 (PAOLO)Document42 pagesREVISED Skeletal System of The Frog 2 (PAOLO)Paolo Naguit100% (2)
- Zoology MidtermsDocument14 pagesZoology MidtermsJULIANNE ANACTANo ratings yet
- 2.1 (A) - Support & Locomotion in Humans & AnimalsDocument59 pages2.1 (A) - Support & Locomotion in Humans & AnimalsFauziah Abu Bakar100% (1)
- Comparative Anatomy of Vertebrate SkeletonDocument53 pagesComparative Anatomy of Vertebrate SkeletonYlane Lee100% (1)
- Skeletal System Animal ScienceDocument16 pagesSkeletal System Animal Sciencealexenneth canilaNo ratings yet
- 2ND Grading Laboratory SheetsDocument16 pages2ND Grading Laboratory SheetsAllecia Leona Arceta SoNo ratings yet
- Lecture 3Document39 pagesLecture 3Anne XxNo ratings yet
- Bio85 Skeletal SystemDocument35 pagesBio85 Skeletal SystemvinlavisoNo ratings yet
- Sekolah Menengah Kebangsaan Permas Jaya: Prepared By:-Azam SenseiDocument59 pagesSekolah Menengah Kebangsaan Permas Jaya: Prepared By:-Azam SenseiazamsenseiNo ratings yet
- 09+AppendicularDocument45 pages09+AppendicularnayoonNo ratings yet
- Sekolah Berasrama Penuh Integrasi Gopeng Prepared By:Muhd Fazli Bin DollahDocument60 pagesSekolah Berasrama Penuh Integrasi Gopeng Prepared By:Muhd Fazli Bin DollahrameshkasinathanNo ratings yet
- Chapter 1-Musculoskeletal SystemDocument113 pagesChapter 1-Musculoskeletal SystemWang Ming YaNo ratings yet
- Frog Skeletal System GuideDocument22 pagesFrog Skeletal System GuideXella RiegoNo ratings yet
- Laboratory 3 SkeletalDocument9 pagesLaboratory 3 SkeletalKyla InoferioNo ratings yet
- Skeletal Urogenital Lecture ComparativeDocument136 pagesSkeletal Urogenital Lecture ComparativeLei Jenevive UmbayNo ratings yet
- Anatomy BonesDocument11 pagesAnatomy BonesDunkin DonutNo ratings yet
- Premedical Biology: Motor MechanismDocument44 pagesPremedical Biology: Motor MechanismsheenaNo ratings yet
- Session 14 Skeletal SystemDocument17 pagesSession 14 Skeletal SystemScience Of LifeNo ratings yet
- Chapter 1: Introduction: Phylum Chordata: The Big FourDocument6 pagesChapter 1: Introduction: Phylum Chordata: The Big FourKariza AbuNo ratings yet
- 7 Unikonta - Deuterostomes & ChordatesDocument5 pages7 Unikonta - Deuterostomes & ChordateskahiauNo ratings yet
- Skeleton Ikan IIDocument15 pagesSkeleton Ikan IIYvonee XaviersNo ratings yet
- Chordata Introduction: Phylum OverviewDocument2 pagesChordata Introduction: Phylum OverviewTrisha RosalesNo ratings yet
- The Vertebral Column, Cranium, Jaw, Ribs, and Intramuscular Bones Comprise The SkeletonDocument17 pagesThe Vertebral Column, Cranium, Jaw, Ribs, and Intramuscular Bones Comprise The SkeletonDewi NurhayatiNo ratings yet
- 2.1 (A) - Support & Locomotion in Humans & AnimalsDocument61 pages2.1 (A) - Support & Locomotion in Humans & Animalsliming8112No ratings yet
- Endoskeleton of Frog (With Diagram) : The Endoskeleton Is Described Under Two Broad HeadsDocument13 pagesEndoskeleton of Frog (With Diagram) : The Endoskeleton Is Described Under Two Broad HeadsMarielle AcojidoNo ratings yet
- Group2 - BSHM 1 BDocument43 pagesGroup2 - BSHM 1 BSAPILAN DENMARNo ratings yet
- Skeletal SystemDocument23 pagesSkeletal SystemrosabelalebsNo ratings yet
- 2.1 (A) - Support & Locomotion in Humans & AnimalsDocument59 pages2.1 (A) - Support & Locomotion in Humans & Animalsliefeng81100% (8)
- Skeletal Muscles Have Muscular & Tendinous PortionsDocument14 pagesSkeletal Muscles Have Muscular & Tendinous PortionsBasim MaqsoodNo ratings yet
- Bones that Make Up the Human Skeletal SystemDocument7 pagesBones that Make Up the Human Skeletal Systemhannah peloniaNo ratings yet
- Anatomia Da ColunaDocument59 pagesAnatomia Da ColunaPaula Duarte MarquesNo ratings yet
- Skeletal System 1Document31 pagesSkeletal System 1Hannan AliNo ratings yet
- Comparative Anatomy: (Chordates) Skeletal SystemDocument66 pagesComparative Anatomy: (Chordates) Skeletal SystemPAU FRANCISCONo ratings yet
- Anatomical IntroductionDocument17 pagesAnatomical Introductionllazoozllbraa 7372No ratings yet
- Osteocytes - Bone Cells Classifications of Bones According To SizeDocument17 pagesOsteocytes - Bone Cells Classifications of Bones According To SizeA CNo ratings yet
- Skeletal Systems in VertebratesDocument26 pagesSkeletal Systems in Vertebratesgoldennyeche080No ratings yet
- Anatomy and Physiology: Chapter 7 - SkeletonDocument3 pagesAnatomy and Physiology: Chapter 7 - Skeletonexpediant2No ratings yet
- Joints of Axial SkeletonDocument24 pagesJoints of Axial SkeletonPraney SlathiaNo ratings yet
- Act08 The Skeletal System DiscussionDocument8 pagesAct08 The Skeletal System DiscussionSofia A.No ratings yet
- Exercise 4: Skeletal SystemDocument8 pagesExercise 4: Skeletal SystemstevemacabsNo ratings yet
- Understanding Cartilage and Bone HistologyDocument28 pagesUnderstanding Cartilage and Bone HistologyMuhammad AwaisNo ratings yet
- 3-General-osteologyDocument90 pages3-General-osteologyvbh8222No ratings yet
- SkeletalDocument21 pagesSkeletalKeith HiadgeNo ratings yet
- Chapter 7 & 8 Notes - A&PDocument8 pagesChapter 7 & 8 Notes - A&PAshley MarieNo ratings yet
- Skeletal System UpdatedDocument55 pagesSkeletal System UpdatedAlthea OcaslaNo ratings yet
- CHNCGDocument7 pagesCHNCGDANIEL LANCE NEVADONo ratings yet
- Skeletal System Zoo 102Document14 pagesSkeletal System Zoo 102Rosselle NoyoalNo ratings yet
- Human Evolution - A Very Short IntroductionDocument1 pageHuman Evolution - A Very Short IntroductionRoberto Oscar LandaNo ratings yet
- Dinosaur: Dinosaurs Are A Diverse Group of Animals of TheDocument5 pagesDinosaur: Dinosaurs Are A Diverse Group of Animals of ThebabublsNo ratings yet
- Basic GeologyDocument39 pagesBasic Geologyduncmcleod100% (1)
- Veronica Castillo Et Al. (2017) - AAPG Chapter 8. Perla Field. The Largest Discvery Ever in LATAMDocument12 pagesVeronica Castillo Et Al. (2017) - AAPG Chapter 8. Perla Field. The Largest Discvery Ever in LATAMprimeradsNo ratings yet
- Proboscideans: Evolution of Elephants and Their RelativesDocument14 pagesProboscideans: Evolution of Elephants and Their RelativesAli NabavizadehNo ratings yet
- Introduction to Oil & Gas: Energy Sources and Future ProspectsDocument44 pagesIntroduction to Oil & Gas: Energy Sources and Future ProspectsRabiathul EleenaNo ratings yet
- 1 An Introduction Basin AnalysisDocument29 pages1 An Introduction Basin AnalysisMuhamadKamilAzharNo ratings yet
- 28 Orders of InsectsDocument5 pages28 Orders of InsectsJoelben NaresNo ratings yet
- Arachnidae of The World Bibliography, Avon 2015Document1,043 pagesArachnidae of The World Bibliography, Avon 2015LUisNo ratings yet
- Instant Download Social Welfare Politics and Public Policy 8th Edition Dinitto Test Bank PDF Full ChapterDocument32 pagesInstant Download Social Welfare Politics and Public Policy 8th Edition Dinitto Test Bank PDF Full ChapterDonnaMoorefspo100% (4)
- Lesson 9 History of The EarthDocument4 pagesLesson 9 History of The EarthRazen SisonNo ratings yet
- Mineral Vein December 2017Document5 pagesMineral Vein December 2017api-275611550100% (1)
- Ceballos Et Al 2015 PDFDocument6 pagesCeballos Et Al 2015 PDFGuillermo Acosta PérezNo ratings yet
- 1979 Hassan The Geologist and ArchaeologyDocument5 pages1979 Hassan The Geologist and ArchaeologySERGIO GIRALDO ARIASNo ratings yet
- Artigo Tafonomia 2Document11 pagesArtigo Tafonomia 2Manoela NogueiraNo ratings yet
- Glaciogenic reservoirs and hydrocarbon systemsDocument2 pagesGlaciogenic reservoirs and hydrocarbon systemsAHMEDNo ratings yet
- T TP 2550624 Ordering The Stages of Fossilisation ks2 Activity - Ver - 1Document2 pagesT TP 2550624 Ordering The Stages of Fossilisation ks2 Activity - Ver - 1deensraja3No ratings yet
- Nuril Trisnawati Te2Document3 pagesNuril Trisnawati Te2Nuril Trisnawati83% (6)
- Mythe To Mitcheldean Mains Reinforcement, Gloucestershire - PotteryDocument24 pagesMythe To Mitcheldean Mains Reinforcement, Gloucestershire - PotteryWessex ArchaeologyNo ratings yet
- Cranial shape evolution in birdsDocument17 pagesCranial shape evolution in birdsHugoBenitezdNo ratings yet
- The Princess and The FrogDocument3 pagesThe Princess and The Frogyayuk setyaningrumNo ratings yet
- Geology Western AustraliaDocument261 pagesGeology Western Australiajavier riosNo ratings yet
- NEW INSECT ORDER DISCOVEREDDocument30 pagesNEW INSECT ORDER DISCOVEREDRafael CedroNo ratings yet
- Fossil NotesDocument8 pagesFossil Notesapi-293690555100% (1)
- Jurassic Park-Character ListDocument3 pagesJurassic Park-Character ListSabina-Andreea AlexandruNo ratings yet
- Biological and Cultural Evolution: From Australopithecus To Homo SapiensDocument4 pagesBiological and Cultural Evolution: From Australopithecus To Homo SapiensAlexander IgnacioNo ratings yet
- Tortonian Scleractinian Corals From The Island of Gavdos (South Greece)Document9 pagesTortonian Scleractinian Corals From The Island of Gavdos (South Greece)anaptdrNo ratings yet
- Sedimentary Geology: An Introduction To Sedimentology and StratigraphyDocument19 pagesSedimentary Geology: An Introduction To Sedimentology and Stratigraphysaptak19900% (1)
- What Would Happen If Dinosaurs Roamed The Earth TodayDocument15 pagesWhat Would Happen If Dinosaurs Roamed The Earth TodayTrisha MateoNo ratings yet