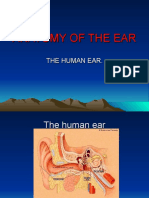
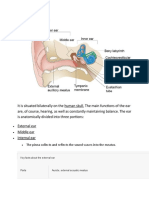
You might also like
- 01.anatomy of The EarDocument52 pages01.anatomy of The EarJumanne Jay100% (1)
- Cleft Lip and Palate Management: A Comprehensive AtlasFrom EverandCleft Lip and Palate Management: A Comprehensive AtlasRicardo D. BennunNo ratings yet
- Central Auditory Processing - Syllabus Fall 2016Document28 pagesCentral Auditory Processing - Syllabus Fall 2016api-349133705No ratings yet
- Physiology of HearingDocument3 pagesPhysiology of HearingKC RajagopalNo ratings yet
- Pseudo BulbarDocument5 pagesPseudo BulbarRegina CaeciliaNo ratings yet
- EXTERNAL EAR DISORDERS AND DEFORMITIESDocument48 pagesEXTERNAL EAR DISORDERS AND DEFORMITIEShemaanandhyNo ratings yet
- Anatomy of The EarDocument8 pagesAnatomy of The EarDarkAngelNo ratings yet
- Drug Induced Ototoxicity - WDocument12 pagesDrug Induced Ototoxicity - WsavitageraNo ratings yet
- Hearing LossDocument31 pagesHearing LossDat boi100% (1)
- TongueDocument10 pagesTongueBhuwanesh SharmaNo ratings yet
- Drooling PDFDocument2 pagesDrooling PDFSuprit SnNo ratings yet
- Embryology of The Ear and Its AnatomyDocument20 pagesEmbryology of The Ear and Its AnatomyElisa NNo ratings yet
- Anatomy of Paranasal SinusesDocument8 pagesAnatomy of Paranasal SinusesPavan TenglikarNo ratings yet
- Lowe Syndrome (Oculocerebrorenal syndrome) A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsFrom EverandLowe Syndrome (Oculocerebrorenal syndrome) A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsNo ratings yet
- Otitis Media With EffusionDocument3 pagesOtitis Media With EffusionAnish RajNo ratings yet
- Osteology of MandibleDocument40 pagesOsteology of MandibleYamini RampriyaNo ratings yet
- GROWTH ASSESSMENT METHODSDocument65 pagesGROWTH ASSESSMENT METHODSdr parveen bathlaNo ratings yet
- Chronic Otitis Media, Cholesteatoma, and Mastoiditis in AdultsDocument14 pagesChronic Otitis Media, Cholesteatoma, and Mastoiditis in AdultschristineNo ratings yet
- 01 - 20131119 Skull IDocument56 pages01 - 20131119 Skull INancyLiao100% (2)
- Special Embryo Tibb TantaDocument15 pagesSpecial Embryo Tibb TantaAhmed GamalNo ratings yet
- Anatomy of The EarDocument8 pagesAnatomy of The EarDarkAngelNo ratings yet
- Inflamed Trachea, (Tracheitis) A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsFrom EverandInflamed Trachea, (Tracheitis) A Simple Guide To The Condition, Diagnosis, Treatment And Related ConditionsNo ratings yet
- Diseases of The EarDocument48 pagesDiseases of The Earabela_amulu100% (1)
- AtelektasisDocument15 pagesAtelektasisMuhammad Ade RahmanNo ratings yet
- The Functional Anatomy of The Cranial NervesDocument29 pagesThe Functional Anatomy of The Cranial NervesRăzvan Moțoc100% (1)
- Malignant Otitis ExternaDocument5 pagesMalignant Otitis Externasergeantchai068No ratings yet
- Otoacoustic Emissions From MedscapeDocument5 pagesOtoacoustic Emissions From MedscapeMin-Joo Esther ParkNo ratings yet
- Ototoxicity LatestDocument37 pagesOtotoxicity LatestSucdi CabdiNo ratings yet
- PPP LarynxDocument65 pagesPPP LarynxAnkur MittalNo ratings yet
- Anatomy of Paranasal SinusesDocument49 pagesAnatomy of Paranasal SinusesNitin SharmaNo ratings yet
- Facial NerveDocument65 pagesFacial NerveprashanthsumsaiarjunNo ratings yet
- Acute Laryngitis: Li SGD 21 LBM 5Document40 pagesAcute Laryngitis: Li SGD 21 LBM 511juni09No ratings yet
- Upper Airway ObstructionDocument31 pagesUpper Airway ObstructiontrimardiyanaisyanNo ratings yet
- Perichondritis PinnaDocument2 pagesPerichondritis PinnaAnish RajNo ratings yet
- External Ear Pathology - PresentationDocument25 pagesExternal Ear Pathology - PresentationNipun MalhotraNo ratings yet
- DYSPHAGIA Lecture NotesDocument84 pagesDYSPHAGIA Lecture Notesmcmak357No ratings yet
- Access Cavity Preparation PDFDocument94 pagesAccess Cavity Preparation PDFtomdienya100% (2)
- Local Anaesthesia and Minor Oral SurgeryDocument33 pagesLocal Anaesthesia and Minor Oral SurgeryAmina AbdullahiNo ratings yet
- ENT Nasal PolypsDocument9 pagesENT Nasal PolypsSTNo ratings yet
- Malignant Otitis Externa 20070821Document0 pagesMalignant Otitis Externa 20070821Suci PramadianiNo ratings yet
- Maxillary SinusDocument25 pagesMaxillary Sinusshreya dasNo ratings yet
- Space InfectionsDocument60 pagesSpace InfectionsDan 04No ratings yet
- Local Complication of Local AnesthesiaDocument22 pagesLocal Complication of Local AnesthesiacobacobatestNo ratings yet
- Ear Infection Assessment and ManagementDocument8 pagesEar Infection Assessment and ManagementArt Christian Ramos100% (1)
- Anatomy of Ear & MastoidDocument137 pagesAnatomy of Ear & MastoidSworupKhadka100% (1)
- Maxillary Sinus Anatomy and PathologyDocument15 pagesMaxillary Sinus Anatomy and PathologyRini RahmawulandariNo ratings yet
- Sailedinitis PDFDocument8 pagesSailedinitis PDFNavatha MorthaNo ratings yet
- Anatomy of The EarDocument10 pagesAnatomy of The EarJP SouaidNo ratings yet
- Thyroglossal Duct CystDocument2 pagesThyroglossal Duct CystPulkitNo ratings yet
- Periodicity DentalGuideDocument52 pagesPeriodicity DentalGuideSalam BataienehNo ratings yet
- Pre-Natal Growth of Craniofacial Complex: Dr. Munizeh KhanDocument36 pagesPre-Natal Growth of Craniofacial Complex: Dr. Munizeh KhanMohsin Habib100% (1)
- Epithelial-Mesenchymal Signalling Regulating Tooth MorphogenesisDocument2 pagesEpithelial-Mesenchymal Signalling Regulating Tooth MorphogenesisalifNo ratings yet
- Stridor in Children: By: Maj Vishal Gaurav Moderator: DR A SethiDocument33 pagesStridor in Children: By: Maj Vishal Gaurav Moderator: DR A SethiVishal GauravNo ratings yet
- Oral AnatomyDocument10 pagesOral AnatomyRobins DhakalNo ratings yet
- Eardrum Repair Surgery DefinitionDocument4 pagesEardrum Repair Surgery DefinitionMelly Selvia ANo ratings yet
- Anatomy of NoseDocument8 pagesAnatomy of Noserizgah jawas100% (1)
- The Effects of Earphone Use On The Incidence of Noise-Induced Hearing Loss An Empirical Study of Senior High School Students in Banda AcehDocument8 pagesThe Effects of Earphone Use On The Incidence of Noise-Induced Hearing Loss An Empirical Study of Senior High School Students in Banda AcehInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Blood Supply To The Maxillary SinusDocument12 pagesBlood Supply To The Maxillary Sinusprostfang100% (2)
- ShobiDocument11 pagesShobiDeshini BalasubramaniamNo ratings yet
- Os Complete Final AnswerDocument198 pagesOs Complete Final AnswerDeshini Balasubramaniam100% (4)
- Lecture №25 Thermal injuries (сo)Document12 pagesLecture №25 Thermal injuries (сo)Deshini BalasubramaniamNo ratings yet
- Lecture №25 Thermal injuries (сo)Document12 pagesLecture №25 Thermal injuries (сo)Deshini BalasubramaniamNo ratings yet
- Bala Graha AfflictionDocument2 pagesBala Graha AfflictionNeeraj VermaNo ratings yet
- Emergency Management of AnaphylaxisDocument1 pageEmergency Management of AnaphylaxisEugene SandhuNo ratings yet
- Mrs. Universe PH - Empowering Women, Inspiring ChildrenDocument2 pagesMrs. Universe PH - Empowering Women, Inspiring ChildrenKate PestanasNo ratings yet
- Consensus Building e Progettazione Partecipata - Marianella SclaviDocument7 pagesConsensus Building e Progettazione Partecipata - Marianella SclaviWilma MassuccoNo ratings yet
- Cot 2Document3 pagesCot 2Kathjoy ParochaNo ratings yet
- Journals OREF Vs ORIF D3rd RadiusDocument9 pagesJournals OREF Vs ORIF D3rd RadiusironNo ratings yet
- Typical T Intersection On Rural Local Road With Left Turn LanesDocument1 pageTypical T Intersection On Rural Local Road With Left Turn Lanesahmed.almakawyNo ratings yet
- Chromate Free CoatingsDocument16 pagesChromate Free CoatingsbaanaadiNo ratings yet
- SDS OU1060 IPeptideDocument6 pagesSDS OU1060 IPeptideSaowalak PhonseeNo ratings yet
- Service Manual: Precision SeriesDocument32 pagesService Manual: Precision SeriesMoises ShenteNo ratings yet
- Alternate Tuning Guide: Bill SetharesDocument96 pagesAlternate Tuning Guide: Bill SetharesPedro de CarvalhoNo ratings yet
- Bula Defense M14 Operator's ManualDocument32 pagesBula Defense M14 Operator's ManualmeNo ratings yet
- United-nations-Organization-uno Solved MCQs (Set-4)Document8 pagesUnited-nations-Organization-uno Solved MCQs (Set-4)SãñÂt SûRÿá MishraNo ratings yet
- 2018 NAMCYA CHILDREN'S RONDALLA ENSEMBLE GuidelinesDocument3 pages2018 NAMCYA CHILDREN'S RONDALLA ENSEMBLE GuidelinesJohn Cedrick JagapeNo ratings yet
- Embryology-Nervous System DevelopmentDocument157 pagesEmbryology-Nervous System DevelopmentGheavita Chandra DewiNo ratings yet
- Excel Solver Optimization ReportDocument9 pagesExcel Solver Optimization ReportMy Duyen NguyenNo ratings yet
- Cover Letter PDFDocument1 pageCover Letter PDFAli EjazNo ratings yet
- Rubric 5th GradeDocument2 pagesRubric 5th GradeAlbert SantosNo ratings yet
- Listening Exercise 1Document1 pageListening Exercise 1Ma. Luiggie Teresita PerezNo ratings yet
- Audi Q5: First Generation (Typ 8R 2008-2017)Document19 pagesAudi Q5: First Generation (Typ 8R 2008-2017)roberto100% (1)
- Kathy Davis - Dancing Tango - Passionate Encounters in A Globalizing World-New York University Press (2015)Document236 pagesKathy Davis - Dancing Tango - Passionate Encounters in A Globalizing World-New York University Press (2015)Csongor KicsiNo ratings yet
- Desana Texts and ContextsDocument601 pagesDesana Texts and ContextsdavidizanagiNo ratings yet
- Oxygen Cost and Energy Expenditure of RunningDocument7 pagesOxygen Cost and Energy Expenditure of Runningnb22714No ratings yet
- Differential Pulse Code ModulationDocument12 pagesDifferential Pulse Code ModulationNarasimhareddy MmkNo ratings yet
- Mpu 2312Document15 pagesMpu 2312Sherly TanNo ratings yet
- Laryngeal Diseases: Laryngitis, Vocal Cord Nodules / Polyps, Carcinoma LarynxDocument52 pagesLaryngeal Diseases: Laryngitis, Vocal Cord Nodules / Polyps, Carcinoma LarynxjialeongNo ratings yet
- Tutorial 1 Discussion Document - Batch 03Document4 pagesTutorial 1 Discussion Document - Batch 03Anindya CostaNo ratings yet
- CFO TagsDocument95 pagesCFO Tagssatyagodfather0% (1)
- Breaking NewsDocument149 pagesBreaking NewstigerlightNo ratings yet
- PEDs and InterferenceDocument28 pagesPEDs and Interferencezakool21No ratings yet