You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Wine GuideDocument4 pagesWine GuidepankajkhakareNo ratings yet
- Winemaking Booklet ProsperoDocument44 pagesWinemaking Booklet ProsperoantonioforteseNo ratings yet
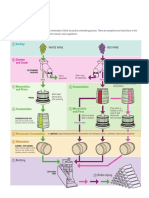
- Doping in Sport - ReviewDocument5 pagesDoping in Sport - ReviewantonioforteseNo ratings yet
- Stone Bee BiologyDocument23 pagesStone Bee BiologyHRTOCNo ratings yet
- Adult Asthma Action PlanDocument2 pagesAdult Asthma Action PlanantonioforteseNo ratings yet
- BeesInDecline PDFDocument48 pagesBeesInDecline PDFanon_635471414No ratings yet
- GINA 2019 Main Report June 2019 Wms PDFDocument201 pagesGINA 2019 Main Report June 2019 Wms PDFAndrada PredaNo ratings yet
- Asthma: Allergy Asthma and Clinical Immunology November 2011Document10 pagesAsthma: Allergy Asthma and Clinical Immunology November 2011Sarmad AlwardiNo ratings yet
- Asthma Diagnosis Monitoring and Chronic Asthma Management PDF 1837687975621Document39 pagesAsthma Diagnosis Monitoring and Chronic Asthma Management PDF 1837687975621hisham hussainNo ratings yet
- Medicine and SurgeryDocument552 pagesMedicine and Surgeryhoneyjoy76100% (2)
- ClematisDocument2 pagesClematisantonioforteseNo ratings yet
- Nuclear Medicine: Science and Safety: Canadian Medical Association Journal October 1996Document3 pagesNuclear Medicine: Science and Safety: Canadian Medical Association Journal October 1996antonioforteseNo ratings yet
- Story of Honey WebDocument11 pagesStory of Honey WebantonioforteseNo ratings yet
- BeesInDecline PDFDocument48 pagesBeesInDecline PDFanon_635471414No ratings yet
- The Backyard Gardener - Simple, Easy, and Beautiful Gardening With Vegetables, Herbs, and FlowersDocument257 pagesThe Backyard Gardener - Simple, Easy, and Beautiful Gardening With Vegetables, Herbs, and Flowersantoniofortese100% (3)
- Elastography of ThyroidDocument8 pagesElastography of ThyroidantonioforteseNo ratings yet
- Fuchsia CareDocument2 pagesFuchsia CareantonioforteseNo ratings yet
- Genitourinary UltrasoundDocument62 pagesGenitourinary Ultrasoundantoniofortese100% (2)
- Growing Fuchsias in The Ground-Barbara's Way: Soil and PositionDocument4 pagesGrowing Fuchsias in The Ground-Barbara's Way: Soil and PositionantonioforteseNo ratings yet
- Floral DiseasesDocument3 pagesFloral DiseasesantonioforteseNo ratings yet
- Alyssum - LobulariaDocument1 pageAlyssum - LobulariaantonioforteseNo ratings yet
- Free 12 X 8 Shed Plan - Step by Step DIYDocument24 pagesFree 12 X 8 Shed Plan - Step by Step DIYgo2top2No ratings yet
- DahliaDocument2 pagesDahliaantonioforteseNo ratings yet
- Achillea - YarrowDocument1 pageAchillea - YarrowantonioforteseNo ratings yet
- Lily-of-the-Valley DocumentDocument1 pageLily-of-the-Valley DocumentantonioforteseNo ratings yet
- Aurinia Saxat1Lis: Golden DustDocument1 pageAurinia Saxat1Lis: Golden DustantonioforteseNo ratings yet
- Acanthus Mollis - Bears BreechesDocument1 pageAcanthus Mollis - Bears BreechesantonioforteseNo ratings yet
- CoreopsisDocument1 pageCoreopsisantonioforteseNo ratings yet
- Ianthus Barbatus: Sweet WilliamDocument4 pagesIanthus Barbatus: Sweet WilliamantonioforteseNo ratings yet
- Gazania - Bright Flowers for Sunny GardensDocument1 pageGazania - Bright Flowers for Sunny GardensantonioforteseNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Microbiology of Wounds: Neal R. Chamberlain, PH.D., Department of Microbiology/Immunology KcomDocument31 pagesThe Microbiology of Wounds: Neal R. Chamberlain, PH.D., Department of Microbiology/Immunology KcomFhaliesha Zakur100% (1)
- 7270 Manuscrito 51969 2 10 20220526Document5 pages7270 Manuscrito 51969 2 10 20220526andreaosorio37No ratings yet
- 5 Useful Tables For Bacterial IdentificationDocument4 pages5 Useful Tables For Bacterial IdentificationVanandiNo ratings yet
- Microbiology Chapter 3Document61 pagesMicrobiology Chapter 3Milad MovahediNo ratings yet
- Jurnal 1Document7 pagesJurnal 1Reza WahyudiNo ratings yet
- Journal of Clinical Microbiology-2019-Munson-e01181-18.full PDFDocument21 pagesJournal of Clinical Microbiology-2019-Munson-e01181-18.full PDFClaudia NeculoiuNo ratings yet
- Bacteria classification guide by growth requirements & structuresDocument7 pagesBacteria classification guide by growth requirements & structuresJorge HammondNo ratings yet
- Lab 3Document6 pagesLab 3Diyana Nabila Abdul WahidNo ratings yet
- Enterobacteriaceae ManualDocument1 pageEnterobacteriaceae ManualMukhtiar AliNo ratings yet
- Discussion MICROBIOLOGY SIMPLE STAINDocument4 pagesDiscussion MICROBIOLOGY SIMPLE STAINKesha Gopal100% (4)
- Daftar PustakaDocument3 pagesDaftar PustakanadaNo ratings yet
- 16 Microbiology MCQsDocument9 pages16 Microbiology MCQsomaromran0% (1)
- Deteksi Keberadaan Bakteri Coliform Pada Depot Air Minum Isi Ulang Di Lingkar Kampus Terpadu Universitas Aisyiyah YogyakartaDocument10 pagesDeteksi Keberadaan Bakteri Coliform Pada Depot Air Minum Isi Ulang Di Lingkar Kampus Terpadu Universitas Aisyiyah YogyakartaVivi AkramNo ratings yet
- Assignment For The Fourth Semesters Botany Department ST MarysDocument3 pagesAssignment For The Fourth Semesters Botany Department ST MarysRimeiaNo ratings yet
- Bacterial CellDocument23 pagesBacterial CellShabeer AhmadNo ratings yet
- Classification of Bacteria On The Basis of ShapeDocument3 pagesClassification of Bacteria On The Basis of ShapeMadelyn AlcalaNo ratings yet
- 1 Structure and Identification of Bacteria 2014 2 1 PDFDocument74 pages1 Structure and Identification of Bacteria 2014 2 1 PDFKaleem AfridiNo ratings yet
- Diagnosticul de Laborator Al: BorreliozelorDocument68 pagesDiagnosticul de Laborator Al: BorreliozelorVLDsNo ratings yet
- Uncovering The Mechanisms of Acinetobacter Baumannii VirulenceDocument12 pagesUncovering The Mechanisms of Acinetobacter Baumannii VirulenceTony ChuNo ratings yet
- Gram Staining Clinical ExerciseDocument10 pagesGram Staining Clinical ExerciseHimani Aggarwal100% (1)
- Bo TanikaDocument90 pagesBo TanikaNedeljkovicL0% (1)
- TN - Gram Positive Vs Gram Negative - Article - JL - V2ksDocument5 pagesTN - Gram Positive Vs Gram Negative - Article - JL - V2ksNGÂN NGUYỄN HOÀNG TRÚCNo ratings yet
- 1 Bacte Mtap 1Document12 pages1 Bacte Mtap 1DENISE MARA�ANo ratings yet
- PONCE - Module 4 - BSN-2 - A18Document6 pagesPONCE - Module 4 - BSN-2 - A18Ponce Kristel Mae ONo ratings yet
- Microgen GN Id Mid65 y Mid641Document20 pagesMicrogen GN Id Mid65 y Mid641Lenin RamirezNo ratings yet
- Bacterial Cell MorphologyDocument4 pagesBacterial Cell MorphologySabrina MascardoNo ratings yet
- Identifying unknown bacteriaDocument2 pagesIdentifying unknown bacteriaAngel JaverNo ratings yet
- Blue - Carba - Prueba Bioquímica para Detección de Carbapenemasas Directamente Desde CultivosDocument3 pagesBlue - Carba - Prueba Bioquímica para Detección de Carbapenemasas Directamente Desde CultivosDavian Araujo PumarejoNo ratings yet
- Annexure 4 Colony Morphology, Shape & Cell Arrangment of MicroorganismDocument6 pagesAnnexure 4 Colony Morphology, Shape & Cell Arrangment of Microorganismdinesh singhNo ratings yet
- Daya Hambat Ekstrak Bunga Ruku-Ruku (Ocimum Sanctum L.) TERHADAP PERTUMBUHAN Streptococcus MutansDocument8 pagesDaya Hambat Ekstrak Bunga Ruku-Ruku (Ocimum Sanctum L.) TERHADAP PERTUMBUHAN Streptococcus Mutanssyarifa husniNo ratings yet