You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Family Stress TheoryDocument10 pagesFamily Stress TheoryKarina Megasari WinahyuNo ratings yet
- Onuaguluchi1996 1Document10 pagesOnuaguluchi1996 1IkaSugihartatikNo ratings yet
- AERO241 Example 10Document4 pagesAERO241 Example 10Eunice CameroNo ratings yet
- (Engine International Air Pollution Prevention) : EIAPP CertificateDocument2 pages(Engine International Air Pollution Prevention) : EIAPP CertificateTan DatNo ratings yet
- Endothermic Gas Production Overview: Tmosphere Ngineering OmpanyDocument6 pagesEndothermic Gas Production Overview: Tmosphere Ngineering OmpanyJhon ChitNo ratings yet
- AGIP STD - Valves Specification SheetDocument1 pageAGIP STD - Valves Specification Sheethalim_kaNo ratings yet
- Acuson P10Document2 pagesAcuson P10anon-259218No ratings yet
- Plumbing Breakup M 01Document29 pagesPlumbing Breakup M 01Nicholas SmithNo ratings yet
- Biodegradable Plastic Bag Manufacturing Industry-800655 PDFDocument68 pagesBiodegradable Plastic Bag Manufacturing Industry-800655 PDFAryanNo ratings yet
- Iso 9227Document13 pagesIso 9227Raj Kumar100% (6)
- 10.1.polendo (Additional Patent)Document11 pages10.1.polendo (Additional Patent)Rima AmaliaNo ratings yet
- E10b MERCHANT NAVY CODE OF CONDUCTDocument1 pageE10b MERCHANT NAVY CODE OF CONDUCTssabih75No ratings yet
- Notice of Privacy Practices (Generic)Document3 pagesNotice of Privacy Practices (Generic)cecilled_08No ratings yet
- Lesson 49Document2 pagesLesson 49Андрій ХомишакNo ratings yet
- Hmo Details November 2022 1Document6 pagesHmo Details November 2022 1Saad BelloNo ratings yet
- Classification of Nanostructured Materials: June 2019Document44 pagesClassification of Nanostructured Materials: June 2019krishnaNo ratings yet
- Resume Massage Therapist NtewDocument2 pagesResume Massage Therapist NtewPartheebanNo ratings yet
- PV2R Series Single PumpDocument14 pagesPV2R Series Single PumpBagus setiawanNo ratings yet
- Remote Control Unit Manual BookDocument21 pagesRemote Control Unit Manual BookIgor Ungur100% (1)
- Rigging: GuideDocument244 pagesRigging: Guideyusry72100% (11)
- Industries Visited in Pune & LonavalaDocument13 pagesIndustries Visited in Pune & LonavalaRohan R Tamhane100% (1)
- Missoula County Fairgrounds Phase 2Document10 pagesMissoula County Fairgrounds Phase 2Olivia IversonNo ratings yet
- Dabur Vs PatanjaliDocument4 pagesDabur Vs PatanjalirangarajanNo ratings yet
- Mdx-40a Use en R1 PDFDocument100 pagesMdx-40a Use en R1 PDFMarcos BustamanteNo ratings yet
- Literary Portraiture & Modern Spain: Dr. Rebecca M. Bender (Document6 pagesLiterary Portraiture & Modern Spain: Dr. Rebecca M. Bender (Pedro PorbénNo ratings yet
- Operational Safety and Health Procedures, Practices and RegulationsDocument20 pagesOperational Safety and Health Procedures, Practices and RegulationsDionisa ErnacioNo ratings yet
- f2607-#### f2607 20191203 111644Document2 pagesf2607-#### f2607 20191203 111644คุณชายธวัชชัย เจริญสุขNo ratings yet
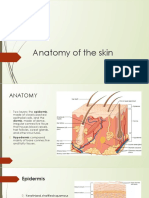
- Anatomy of The SkinDocument28 pagesAnatomy of The Skinay254No ratings yet
- 2015 12 17 - Parenting in America - FINALDocument105 pages2015 12 17 - Parenting in America - FINALKeaneNo ratings yet
- COT RPMS Rating Sheet For MT I IV For SY 2021 2022 in The Time of COVID 19Document2 pagesCOT RPMS Rating Sheet For MT I IV For SY 2021 2022 in The Time of COVID 19Don Mariano Marcos Elementary SchoolNo ratings yet