You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Surinamensoids CichlidsDocument4 pagesThe Surinamensoids CichlidsGil Iack XimenesNo ratings yet
- BIODIVERSITI f2 2017Document54 pagesBIODIVERSITI f2 2017farah6214100% (1)
- Excretory Products and Their EliminationDocument9 pagesExcretory Products and Their EliminationBiju MylachalNo ratings yet
- Lab Report Cells Seen Under The MicroscopeDocument5 pagesLab Report Cells Seen Under The Microscopeorlandoe2560% (1)
- Synonym (Taxonomy)Document5 pagesSynonym (Taxonomy)Constantin TodiNo ratings yet
- Application Form For Master by Research/Ed.D/Phd Programme of The University of Nottingham Ningbo ChinaDocument5 pagesApplication Form For Master by Research/Ed.D/Phd Programme of The University of Nottingham Ningbo ChinaΧαράλαμποςΧατζηκανέλλουNo ratings yet
- Xanthomonas Axonopodis PV AlliiDocument1 pageXanthomonas Axonopodis PV AlliiΧαράλαμποςΧατζηκανέλλουNo ratings yet
- Menu International DinnerDocument8 pagesMenu International DinnerΧαράλαμποςΧατζηκανέλλουNo ratings yet
- Caplow TriadsDocument7 pagesCaplow TriadsΧαράλαμποςΧατζηκανέλλουNo ratings yet
- RSPB 20091076Document7 pagesRSPB 20091076ΧαράλαμποςΧατζηκανέλλουNo ratings yet
- Mosquito Special Reference Albopictus, Novalbopictus And: Oviposition Traps, TO SubalbatusDocument8 pagesMosquito Special Reference Albopictus, Novalbopictus And: Oviposition Traps, TO SubalbatusΧαράλαμποςΧατζηκανέλλουNo ratings yet
- Test Procedures For Insecticide Resistance Monitoring in Malaria Vector MosquitoesDocument40 pagesTest Procedures For Insecticide Resistance Monitoring in Malaria Vector MosquitoesΧαράλαμποςΧατζηκανέλλουNo ratings yet
- Muscles of MasticationDocument18 pagesMuscles of MasticationgauravNo ratings yet
- Lect 2-Intro ContinuedDocument42 pagesLect 2-Intro ContinuedSleeping BeautyNo ratings yet
- JKSSB Class IV 27 Feb Paper JK Latest InfoDocument12 pagesJKSSB Class IV 27 Feb Paper JK Latest InfoIdrees AhmedNo ratings yet
- Distinguishing Skulls of Lions (Panthera Leo) and Tigers (Panthera Tigris)Document6 pagesDistinguishing Skulls of Lions (Panthera Leo) and Tigers (Panthera Tigris)brentlion100% (3)
- Colonial Structure of Communication PDFDocument2 pagesColonial Structure of Communication PDFRichardNo ratings yet
- Animal Structure Ruminant and MonogastricDocument23 pagesAnimal Structure Ruminant and MonogastricQuinnee VallejosNo ratings yet
- SassafasDocument462 pagesSassafashasla18No ratings yet
- Land AnimalsDocument52 pagesLand AnimalsVinuta AilNo ratings yet
- ds87 Intro Monohybrid CrossesDocument7 pagesds87 Intro Monohybrid Crossesapi-110789702No ratings yet
- Sexual Reproduction in Flowering Plants: Pre Fertilisation EventsDocument16 pagesSexual Reproduction in Flowering Plants: Pre Fertilisation EventsYashwanth SrinivasaNo ratings yet
- Zoology - Ijzr - Diversity of Spiders Around - Priya SharmaDocument8 pagesZoology - Ijzr - Diversity of Spiders Around - Priya SharmaTJPRC PublicationsNo ratings yet
- Captive Care of North American Water TurtlesDocument160 pagesCaptive Care of North American Water TurtlesAnthony Lopez SahagunNo ratings yet
- Stictia: Last Updated: 23 November, 2017Document10 pagesStictia: Last Updated: 23 November, 2017PedroVaNo ratings yet
- Reptiles & Amphibians: Second Report of Ophiophagy in A Cat-Eyed Snake (Leptodeira SP.) in Costa RicaDocument2 pagesReptiles & Amphibians: Second Report of Ophiophagy in A Cat-Eyed Snake (Leptodeira SP.) in Costa Ricaapi-309280225No ratings yet
- Szovenyi Natcroat 2012 2Document16 pagesSzovenyi Natcroat 2012 2Ir MaNo ratings yet
- 2019 Indiana Dunes Birding Festival GuideDocument25 pages2019 Indiana Dunes Birding Festival GuideBrad Bumgardner100% (8)
- (Edu - Joshuatly.com) Trial Kelantan STPM 2012 Biology (0FF2679E)Document0 pages(Edu - Joshuatly.com) Trial Kelantan STPM 2012 Biology (0FF2679E)otpengNo ratings yet
- Phylum Porifera: / P Ɒ R Ɪ F ƏR ƏDocument4 pagesPhylum Porifera: / P Ɒ R Ɪ F ƏR ƏPeterjohn QuimingNo ratings yet
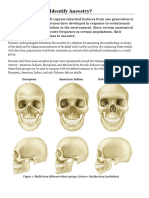
- Activity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanDocument5 pagesActivity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanpoovenmoonoosamyNo ratings yet
- Animal KingdomDocument2 pagesAnimal KingdomDébàshis DashNo ratings yet
- Kedah Matriculation College: BiodiversityDocument12 pagesKedah Matriculation College: BiodiversityMaisarahNo ratings yet
- Biodiversity of The National Zoo (Sri Lanka)Document9 pagesBiodiversity of The National Zoo (Sri Lanka)D.M.S. Suranjan KarunarathnaNo ratings yet
- Malacologia 16264 InstDocument532 pagesMalacologia 16264 InstniccosanNo ratings yet
- Usborne - Baby - S Book of RhymesDocument37 pagesUsborne - Baby - S Book of Rhymessugest270383815No ratings yet
- Npre2007241 2 PDFDocument134 pagesNpre2007241 2 PDFranjeet kumarNo ratings yet