You might also like
- Dynamic Model of CHO Cell Metabolism PDFDocument17 pagesDynamic Model of CHO Cell Metabolism PDFIdalba SouzaNo ratings yet
- q3 Basic CalculusDocument8 pagesq3 Basic CalculusDon Aldrich SantosNo ratings yet
- Gas Turbine Fuel SelectionDocument8 pagesGas Turbine Fuel Selectionblooom_00100% (1)
- Atpdt 9 Vcu 20103Document158 pagesAtpdt 9 Vcu 20103JenderalKancilNo ratings yet
- Analytic Thinking and Presentation For Intelligence Analysis Training Handbook CIA 88ppDocument88 pagesAnalytic Thinking and Presentation For Intelligence Analysis Training Handbook CIA 88ppjtl_goose100% (4)
- Protein Purification - CACDocument21 pagesProtein Purification - CACAhammed Abu Dil100% (1)
- Cell Biology Assays: ProteinsFrom EverandCell Biology Assays: ProteinsFanny JaulinNo ratings yet
- Jobses 1Document10 pagesJobses 1Mindy MunozNo ratings yet
- A Growth Kinetic Model of Cultures On Cheese Whey As SubstrateDocument6 pagesA Growth Kinetic Model of Cultures On Cheese Whey As Substratewp_dessyNo ratings yet
- Ba VariableDocument8 pagesBa VariableDavid SantosNo ratings yet
- Lei 2001Document17 pagesLei 2001andres granadosNo ratings yet
- Jurnal TFDocument6 pagesJurnal TFDhian ZhahirahNo ratings yet
- A Structured Model For Penicillin Production On Mixed SubstratesDocument11 pagesA Structured Model For Penicillin Production On Mixed SubstratesTela24No ratings yet
- Simulation of Enzymatic Hydrolysis of Wheat Straw Using Autohydrolysis and OrganosolvDocument1 pageSimulation of Enzymatic Hydrolysis of Wheat Straw Using Autohydrolysis and OrganosolvSemana da Escola de Engenharia da Universidade do MinhoNo ratings yet
- Akesson01b PDFDocument6 pagesAkesson01b PDFajayterdal8471No ratings yet
- Product and Cell Yield CoefficientsDocument2 pagesProduct and Cell Yield CoefficientsBeauregardNo ratings yet
- Metabolic Control Theory: Its Role in Microbiology and BiotechnologyDocument16 pagesMetabolic Control Theory: Its Role in Microbiology and BiotechnologyEstalin CaizapantaNo ratings yet
- Quickly SlidesCarnivalDocument18 pagesQuickly SlidesCarnivalJILLIAN DALUPONo ratings yet
- Towards Energy-Based Dynamic Optimization of Monoclonal Antibody Producing GS-NS0 CulturesDocument6 pagesTowards Energy-Based Dynamic Optimization of Monoclonal Antibody Producing GS-NS0 CulturesaminghostNo ratings yet
- Fuhrer 2011Document7 pagesFuhrer 2011CoNo ratings yet
- What Is Flux Balance Analysis? Supplementary Tutorial: Beginner COBRA MethodsDocument28 pagesWhat Is Flux Balance Analysis? Supplementary Tutorial: Beginner COBRA MethodsMridula PrasadNo ratings yet
- LT2 - Modelo Cinetico Pseudo-Intrinseco de La Bioconversion de CetonaDocument9 pagesLT2 - Modelo Cinetico Pseudo-Intrinseco de La Bioconversion de CetonaMafer CVNo ratings yet
- Xo Mics To ModelDocument61 pagesXo Mics To ModelVijendra KavatalkarNo ratings yet
- Analysis of Conversion and Operation Strategies For Enzymatic Hydrolysis of Lignocellulosic Biomass in A Series of Cstrs With Distributed FeedingDocument10 pagesAnalysis of Conversion and Operation Strategies For Enzymatic Hydrolysis of Lignocellulosic Biomass in A Series of Cstrs With Distributed FeedingVictor PuglieseNo ratings yet
- Immobilized Enzyme Reactor 1 PDFDocument5 pagesImmobilized Enzyme Reactor 1 PDFPrajakta NakateNo ratings yet
- CHEN 509 Tutorial Revised - 2024Document5 pagesCHEN 509 Tutorial Revised - 2024Enemona ShellengNo ratings yet
- Bioautomation, 2007, 7, 34 - 45Document12 pagesBioautomation, 2007, 7, 34 - 45olympia1047No ratings yet
- Biotechnology: Economic Evaluation of Alternative Ethanol Fermentation ProcessesDocument23 pagesBiotechnology: Economic Evaluation of Alternative Ethanol Fermentation ProcessesMary Grace VelitarioNo ratings yet
- V LyubenovaDocument5 pagesV LyubenovaMARKASGEORGENo ratings yet
- Extended AbstractDocument14 pagesExtended AbstractVenkata Suryanarayana GorleNo ratings yet
- Optimal Biocatalyst Loading in A Fixed BedDocument7 pagesOptimal Biocatalyst Loading in A Fixed BedEleazar EscamillaNo ratings yet
- Paper 1 Bio 1 LabDocument4 pagesPaper 1 Bio 1 Labheidia92No ratings yet
- Metabolomics: Topics in Chemical Biology: Advanced ArticleDocument10 pagesMetabolomics: Topics in Chemical Biology: Advanced ArticleazzaassNo ratings yet
- Zhang - 2017 - Quantification of Lactate Isotopologues and IsotopomersDocument12 pagesZhang - 2017 - Quantification of Lactate Isotopologues and IsotopomersMokr Guerrero EsperanzaNo ratings yet
- Enzyme CatalysisDocument15 pagesEnzyme CatalysisMaelyn Nicole RominNo ratings yet
- Effect of PH On Solvent Flux During Stirred Ultrafiltration of ProteinsDocument8 pagesEffect of PH On Solvent Flux During Stirred Ultrafiltration of ProteinsRoxana ElenaNo ratings yet
- Dynamic Modeling of The Central Carbon Metabolism of Escherichia ColiDocument21 pagesDynamic Modeling of The Central Carbon Metabolism of Escherichia ColiclapatriciasaNo ratings yet
- tmp1DDA TMPDocument4 pagestmp1DDA TMPFrontiersNo ratings yet
- Abstract 2009 ZurigoDocument1 pageAbstract 2009 Zurigoalan1_white2No ratings yet
- Improvements in Titer, Productivity, and Yield Using Solka-Floc For Cellulase ProductionDocument16 pagesImprovements in Titer, Productivity, and Yield Using Solka-Floc For Cellulase Productionguime27No ratings yet
- Journal of Life Sciences & BiomedicineDocument10 pagesJournal of Life Sciences & BiomedicineBiomedicince journalNo ratings yet
- Linjianqiang 6Document8 pagesLinjianqiang 6Med Ramy Gaddem EssefiNo ratings yet
- Mathematical ModelingDocument7 pagesMathematical ModelingalirezamdfNo ratings yet
- Protein - Ligand Binding With The Coarse-Grained Martini ModelDocument11 pagesProtein - Ligand Binding With The Coarse-Grained Martini Modelchrisjorg@gmail.comNo ratings yet
- Aiba 1968 Kinetics of Product Inhibition in Alcohol FermentationDocument20 pagesAiba 1968 Kinetics of Product Inhibition in Alcohol Fermentationrishu2525No ratings yet
- Alex - S Protein Folding For SCIENCE EPQ StuffDocument3 pagesAlex - S Protein Folding For SCIENCE EPQ StuffFunkymaleNo ratings yet
- Determination of Protein Content SpectrophotometricallyDocument10 pagesDetermination of Protein Content SpectrophotometricallyTsabit AlbananiNo ratings yet
- Pfeffer Et Al. - 2011 - Modeling and Measuring Intracellular Fluxes of SecDocument11 pagesPfeffer Et Al. - 2011 - Modeling and Measuring Intracellular Fluxes of SecAnaNo ratings yet
- 2021 12 05 471285v2 FullDocument27 pages2021 12 05 471285v2 Fullfatinnafis27No ratings yet
- Modeling and Parameter Estimation of A Nonlinear Switching Systemin Fed-Batch Culture With PH FeedbackDocument11 pagesModeling and Parameter Estimation of A Nonlinear Switching Systemin Fed-Batch Culture With PH FeedbackDeepak KumarNo ratings yet
- Arndt Et Al - 2005Document8 pagesArndt Et Al - 2005Felipe MatheusNo ratings yet
- A Single Nutrient Feed Supports Both Chemically Defined NS0 and CHODocument11 pagesA Single Nutrient Feed Supports Both Chemically Defined NS0 and CHOMingzhong ChenNo ratings yet
- Cell kinetics-IIIDocument12 pagesCell kinetics-IIIVineet Kumar ThakurNo ratings yet
- ELETROFORESEPRECIPITAÇÃODocument12 pagesELETROFORESEPRECIPITAÇÃOBruna BarbosaNo ratings yet
- 2 Biochemical Engineering CH 2Document54 pages2 Biochemical Engineering CH 2Fasil ManNo ratings yet
- International Journal of EngineeringDocument10 pagesInternational Journal of EngineeringChokri ChakiirNo ratings yet
- Fed-Batch Culture Technology: Biotech. Adv. 0734-9750/86Document12 pagesFed-Batch Culture Technology: Biotech. Adv. 0734-9750/86marco hernandezNo ratings yet
- Articles: Neural Network Prediction of Biomass Digestibility Based On Structural FeaturesDocument10 pagesArticles: Neural Network Prediction of Biomass Digestibility Based On Structural FeaturesSaif EvonyNo ratings yet
- Using Modelica-Matlab For Parameter Estimation in A Bioethanol Fermentation ModelDocument13 pagesUsing Modelica-Matlab For Parameter Estimation in A Bioethanol Fermentation ModelwariascNo ratings yet
- 8.3 Fed Batch Reactors 8.3.1 Variable Volume Fermentation (VARVOL and Varvold)Document34 pages8.3 Fed Batch Reactors 8.3.1 Variable Volume Fermentation (VARVOL and Varvold)Hana HamidNo ratings yet
- Fermentation Process KineticsDocument7 pagesFermentation Process KineticsDillip_subuNo ratings yet
- Problem Set 3Document2 pagesProblem Set 3Nigel Kow0% (1)
- Activity 1 Merge-KMJ32503 - 230406 - 125701Document25 pagesActivity 1 Merge-KMJ32503 - 230406 - 125701Bharrathi RagupathyNo ratings yet
- MillatDocument22 pagesMillatMARIA DE LA PAZ GRAJEDA PINEDANo ratings yet
- International Journal of Infectious Diseases: EditorialDocument5 pagesInternational Journal of Infectious Diseases: Editorialblooom_00No ratings yet
- NIH Public Access: Current Concepts in Host-Microbe Interaction Leading To Pneumococcal PneumoniaDocument15 pagesNIH Public Access: Current Concepts in Host-Microbe Interaction Leading To Pneumococcal Pneumoniablooom_00No ratings yet
- Previews: Living in Peace: Host-Microbiota Mutualism in The SkinDocument2 pagesPreviews: Living in Peace: Host-Microbiota Mutualism in The Skinblooom_00No ratings yet
- Biosensors and Bioelectronics: Meng Wang, Rui Zhang, Jinming LiDocument18 pagesBiosensors and Bioelectronics: Meng Wang, Rui Zhang, Jinming Liblooom_00No ratings yet
- The Negative Impact of Antibiotic Resistance: ReviewDocument7 pagesThe Negative Impact of Antibiotic Resistance: Reviewblooom_00No ratings yet
- Miscible Ethanol Ether Chloroform Ethyl Cellulose Polyvinyl Butyral Ethanol Methanol Aqueous Solutions Sodium ChlorideDocument1 pageMiscible Ethanol Ether Chloroform Ethyl Cellulose Polyvinyl Butyral Ethanol Methanol Aqueous Solutions Sodium Chlorideblooom_00No ratings yet
- Heart Transplantation: Pediatric RecipientsDocument199 pagesHeart Transplantation: Pediatric Recipientsblooom_00No ratings yet
- Health Services and Outcomes Research: Cardiac Complications in Patients With Community-Acquired PneumoniaDocument9 pagesHealth Services and Outcomes Research: Cardiac Complications in Patients With Community-Acquired Pneumoniablooom_00No ratings yet
- MordorDocument2 pagesMordorblooom_00No ratings yet
- Butane (Butane or Isobutane (Also Called "Methylpropane"), or To A Mixture of TheseDocument2 pagesButane (Butane or Isobutane (Also Called "Methylpropane"), or To A Mixture of Theseblooom_00No ratings yet
- Japanese Cinema - Film Style and National Character - Donald Richie PDFDocument291 pagesJapanese Cinema - Film Style and National Character - Donald Richie PDFilgrandereNo ratings yet
- TurmericDocument6 pagesTurmericblooom_00No ratings yet
- Research MgPropertiesDocument8 pagesResearch MgPropertiesblooom_00No ratings yet
- Name: Charina Malolot-Villalon Exercise 4: Descriptive Statistics Instruction: Download Population Data at The Barangay Level of Your RespectiveDocument2 pagesName: Charina Malolot-Villalon Exercise 4: Descriptive Statistics Instruction: Download Population Data at The Barangay Level of Your RespectiveChrina MalolotNo ratings yet
- 84 Picon NunezDocument6 pages84 Picon Nunez1940LaSalleNo ratings yet
- Course Outline: Course Title: Linear Algebra Course Code: EDME3201 (ME32A) Level: 3 Semester: 3 Credits: 3Document3 pagesCourse Outline: Course Title: Linear Algebra Course Code: EDME3201 (ME32A) Level: 3 Semester: 3 Credits: 3Perry SinNo ratings yet
- Coeficientes A B Y C para TermistoresDocument4 pagesCoeficientes A B Y C para TermistorescarlosNo ratings yet
- 2nd Quaarter ExamDocument2 pages2nd Quaarter ExamJoana Marie DeontoyNo ratings yet
- Retaining Wall-Masonry Design and Calculation SpreadsheetDocument5 pagesRetaining Wall-Masonry Design and Calculation SpreadsheetTECHNOCONS CONST.No ratings yet
- Bcs Higher Education Qualifications Bcs Level 5 Diploma in ItDocument3 pagesBcs Higher Education Qualifications Bcs Level 5 Diploma in ItDesizm .ComNo ratings yet
- Morelli 2013Document4 pagesMorelli 2013farrukhNo ratings yet
- Haskell FunctionalDocument22 pagesHaskell FunctionaldfjhNo ratings yet
- Mathematics 2023 (65-4-2) Set-2Document49 pagesMathematics 2023 (65-4-2) Set-2prajjwalroyroyNo ratings yet
- The Sine Rule Worksheet #01, Shape & Space Revision From GCSE Maths TutorDocument2 pagesThe Sine Rule Worksheet #01, Shape & Space Revision From GCSE Maths TutorgcsemathstutorNo ratings yet
- Es 13 Prob Set 1Document5 pagesEs 13 Prob Set 1Julian de LaraNo ratings yet
- Notes For Students Xi (Programming and Computational Thinking) PDFDocument106 pagesNotes For Students Xi (Programming and Computational Thinking) PDFsuryansh tyagi0% (2)
- Solving Accelerated Motion ProblemsDocument23 pagesSolving Accelerated Motion ProblemsrhazoorNo ratings yet
- Lec1.2 IITM Week 1 Stats2Document9 pagesLec1.2 IITM Week 1 Stats2samudraneel05No ratings yet
- 3 - 2 Logarithmic FNS and Their Graphs PDFDocument29 pages3 - 2 Logarithmic FNS and Their Graphs PDFAikaterine Simbajon GabisayNo ratings yet
- Eda Module 3 AnsDocument11 pagesEda Module 3 AnsNiño AntoninoNo ratings yet
- Engineering Mathematics IIDocument102 pagesEngineering Mathematics IIIsaac P PlanNo ratings yet
- Bertram and Rehdanz - 2014 - The Role of Urban Green Space On WBDocument33 pagesBertram and Rehdanz - 2014 - The Role of Urban Green Space On WBIsabella Valencia VernazaNo ratings yet
- Lecture10 - FlowCharts IDocument41 pagesLecture10 - FlowCharts IFatima AqeelNo ratings yet
- USAGE Clause in CobolDocument3 pagesUSAGE Clause in CobolRammurthyNo ratings yet
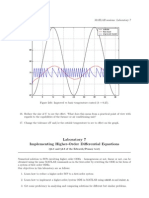
- LAB07Document7 pagesLAB07jlbalbNo ratings yet
- 20itt43 Daa Unit IDocument122 pages20itt43 Daa Unit Irt.cseNo ratings yet
- RESEARCH WORK 2 AlejandroDocument28 pagesRESEARCH WORK 2 AlejandroAngelica AlejandroNo ratings yet
- Chapter 4 Decision Support System (Dr. Mahmoud Mostafa)Document32 pagesChapter 4 Decision Support System (Dr. Mahmoud Mostafa)JonathanHindiNo ratings yet
- mpm2d1 Course Overview 2017Document2 pagesmpm2d1 Course Overview 2017api-235072058No ratings yet
- Go Math Homework 1st GradeDocument7 pagesGo Math Homework 1st Gradeafnaewonaoejmm100% (1)