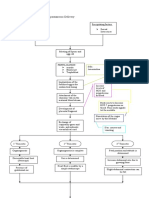
You might also like
- Physiology of Normal Spontaneous DeliveryDocument2 pagesPhysiology of Normal Spontaneous DeliverySummer Rain100% (2)
- Nonextraction Treatment: by Norman M. Cetlin, Raffaele SpenaDocument78 pagesNonextraction Treatment: by Norman M. Cetlin, Raffaele SpenaMuhammad AsimNo ratings yet
- Opus LoopDocument9 pagesOpus LoopMuhammad AsimNo ratings yet
- Genetic Basis of Tooth Development and Dental DefectsDocument5 pagesGenetic Basis of Tooth Development and Dental DefectsJhon Mario Pinto RamirezNo ratings yet
- Thesleff 2018Document5 pagesThesleff 2018jalozanorNo ratings yet
- Genetics of AbnormalitiesDocument11 pagesGenetics of AbnormalitiesLjubomirErdoglijaNo ratings yet
- Epithelial-Mesenchymal Signalling Regulating Tooth MorphogenesisDocument2 pagesEpithelial-Mesenchymal Signalling Regulating Tooth MorphogenesisalifNo ratings yet
- Molecular Basis of Craniofacial Development and GrowthDocument2 pagesMolecular Basis of Craniofacial Development and GrowthCornelia GheorghitaNo ratings yet
- Developmental Genetics and Pharmacogeneticsdoc4301Document8 pagesDevelopmental Genetics and Pharmacogeneticsdoc4301sayednourNo ratings yet
- Genetics in OrthodonticsDocument56 pagesGenetics in OrthodonticsSadhu AbhijeetNo ratings yet
- Craniofacial Biology - Gene Expression During Tooth DevelopmentDocument3 pagesCraniofacial Biology - Gene Expression During Tooth DevelopmentzeljkostojakovicNo ratings yet
- Cobourne (1999) The Genetic Control of Early OdontogenesisDocument0 pagesCobourne (1999) The Genetic Control of Early OdontogenesisMaja Maja BułkaNo ratings yet
- Microarray Evaluation of Age-Related Changes in Human Dental PulpDocument7 pagesMicroarray Evaluation of Age-Related Changes in Human Dental PulpnagygeNo ratings yet
- Osseointegration Comunication Cell 2012Document9 pagesOsseointegration Comunication Cell 2012César Rafael Barrera BenavidesNo ratings yet
- Genetics in Pediatric Dentistry - A Review: Drifzah, DR - Divyaprahlad, DR - Priya Subramaniam, DR - Kinjal ShahDocument9 pagesGenetics in Pediatric Dentistry - A Review: Drifzah, DR - Divyaprahlad, DR - Priya Subramaniam, DR - Kinjal Shahanusha guptaNo ratings yet
- Bone and Developement BookDocument321 pagesBone and Developement BookGiannis Iakovou100% (4)
- Epigenetics - A New Frontier in DentistryDocument11 pagesEpigenetics - A New Frontier in DentistryDiego Andres Hincapie HerreraNo ratings yet
- Upregulation of Phosphorylated HSP27, PRDX2, GRP75, GRP78 and GRP94 in Acquired Middle Ear Cholesteatoma GrowthDocument21 pagesUpregulation of Phosphorylated HSP27, PRDX2, GRP75, GRP78 and GRP94 in Acquired Middle Ear Cholesteatoma Growthprofesor conanNo ratings yet
- Role of Genetics in Orthodontics - Simplified ConceptDocument5 pagesRole of Genetics in Orthodontics - Simplified ConceptInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- 954 1638 1 SM PDFDocument6 pages954 1638 1 SM PDFIndah AmaliaNo ratings yet
- Epigenetics and Psychoneuroinmunology. Mechanims and ModelsDocument15 pagesEpigenetics and Psychoneuroinmunology. Mechanims and ModelsSparkniano HeNo ratings yet
- Prenatal PDFDocument56 pagesPrenatal PDFJohn MccormickNo ratings yet
- Comparative Gene Expression Analysis of Genital TubercleDocument29 pagesComparative Gene Expression Analysis of Genital TubercleAnonymous rprdjdFMNzNo ratings yet
- World's Largest Science, Technology & Medicine Open Access Book PublisherDocument30 pagesWorld's Largest Science, Technology & Medicine Open Access Book PublisherVeronica ArchiekinsNo ratings yet
- Dev Bio IntroductoryDocument17 pagesDev Bio Introductoryreynaldo banaria jrNo ratings yet
- Dimorfismo SexualDocument21 pagesDimorfismo SexualDiana CortesNo ratings yet
- Developmental Dynamics - 2016 - Christ - LRP2 An Auxiliary Receptor That Controls Sonic Hedgehog Signaling in DevelopmentDocument11 pagesDevelopmental Dynamics - 2016 - Christ - LRP2 An Auxiliary Receptor That Controls Sonic Hedgehog Signaling in Developmenthennysusanto18No ratings yet
- V 014 Supp 02 A 015Document1 pageV 014 Supp 02 A 015Isaac Ooi Poh JitNo ratings yet
- Celulas Madre en Los Dientes-Dental Cell Type Atlas Reveals Stem and Differentiated Cell Types in Mouse and Human TeethDocument18 pagesCelulas Madre en Los Dientes-Dental Cell Type Atlas Reveals Stem and Differentiated Cell Types in Mouse and Human TeethSusana FariasNo ratings yet
- Reiterative Signaling and Patterning During MammalianDocument11 pagesReiterative Signaling and Patterning During MammalianJhon Mario Pinto RamirezNo ratings yet
- Basic Sciences (Anatomy, Physiology), Management of Medically Compromised Patients and Drug InteractionsDocument64 pagesBasic Sciences (Anatomy, Physiology), Management of Medically Compromised Patients and Drug InteractionsNancyLiaoNo ratings yet
- Cleft Lip and Palate: Etiological Factors, A ReviewDocument8 pagesCleft Lip and Palate: Etiological Factors, A ReviewAbu-Hussein MuhamadNo ratings yet
- Genetics and Orthodontics-FinalDocument96 pagesGenetics and Orthodontics-FinalRaj SinghNo ratings yet
- Hypodontia: An Update On It 'S EtiologyDocument7 pagesHypodontia: An Update On It 'S Etiologyionut1ilie-1No ratings yet
- Cell Lines and Primary Cell Cultures in The Study of Bone Cell Biology PDFDocument24 pagesCell Lines and Primary Cell Cultures in The Study of Bone Cell Biology PDFFilipa PereiraNo ratings yet
- Developmental GeneticsDocument20 pagesDevelopmental GeneticsabhishekNo ratings yet
- NIH Public Access: Author ManuscriptDocument25 pagesNIH Public Access: Author ManuscriptMaria SilvaNo ratings yet
- Abnormalities in Cartilage and Bone Development in The Apert Syndrome FGFR2+/S252W MouDocument12 pagesAbnormalities in Cartilage and Bone Development in The Apert Syndrome FGFR2+/S252W MouMelissa SanchezNo ratings yet
- Growth and Transcription Factors in Tooth DevelopmDocument15 pagesGrowth and Transcription Factors in Tooth DevelopmM.Firwan KhalidNo ratings yet
- Jacobs, Ku, Que - 2012 - Genetic and Cellular Mechanisms Regulating Anterior Foregut and Esophageal DevelopmentDocument11 pagesJacobs, Ku, Que - 2012 - Genetic and Cellular Mechanisms Regulating Anterior Foregut and Esophageal DevelopmentCarly LeungNo ratings yet
- TGF-B Family Signaling in Mesenchymal DifferentiationDocument55 pagesTGF-B Family Signaling in Mesenchymal DifferentiationJosé Pablo IglesiasNo ratings yet
- Jhoanna Rein S. Duzon 4B4Document8 pagesJhoanna Rein S. Duzon 4B4Jhoanna Rein DuzonNo ratings yet
- Regulatory Mechanisms of Soft Palate Development and MalformationsDocument9 pagesRegulatory Mechanisms of Soft Palate Development and MalformationsMuhammad Alif SatrioNo ratings yet
- MouseDocument14 pagesMouseapi-345977843No ratings yet
- EpigeneticsDocument5 pagesEpigeneticsBioBotimy100% (1)
- tmp43CF TMPDocument11 pagestmp43CF TMPFrontiersNo ratings yet
- Genetics in HumansDocument3 pagesGenetics in HumansArif HidyawanNo ratings yet
- Epigenetic Eti Old IDocument10 pagesEpigenetic Eti Old IStella IguiniNo ratings yet
- Fibrous Dysplasia-A Review Article: October 2018Document8 pagesFibrous Dysplasia-A Review Article: October 2018Phúc BùiNo ratings yet
- Omics IntroductionDocument25 pagesOmics IntroductionMarlvin PrimeNo ratings yet
- A Curriculum Vitae of Teeth Evolution, Generation, RegenerationDocument18 pagesA Curriculum Vitae of Teeth Evolution, Generation, RegenerationDulce MartinezNo ratings yet
- Principles of Genetics:: Genetic Factors and Inherited Factors, Its Role On Cranio-Facial AbnormalitiesDocument12 pagesPrinciples of Genetics:: Genetic Factors and Inherited Factors, Its Role On Cranio-Facial AbnormalitiesJohn CenaNo ratings yet
- The Life Histories of GenesDocument3 pagesThe Life Histories of Genesjyotirmoy21No ratings yet
- Developmental Human Leydig CellDocument9 pagesDevelopmental Human Leydig CellDelavemia RNo ratings yet
- Genes Dev. 2015 Quesnel Vallières 746 59Document15 pagesGenes Dev. 2015 Quesnel Vallières 746 59Misael G JordanNo ratings yet
- 7 Gendev PDFDocument12 pages7 Gendev PDFJohn DeHansNo ratings yet
- #3 PalatogenesisDocument14 pages#3 PalatogenesisValentina MoralesNo ratings yet
- Seven Types of Pleiotropy: Jonathan HodgkinDocument5 pagesSeven Types of Pleiotropy: Jonathan HodgkinAriyo 'Jhon Jhay' IbukunNo ratings yet
- Genetics of Dentofacial and Orthodontic AbnormalitiesDocument6 pagesGenetics of Dentofacial and Orthodontic AbnormalitiesMariana SantosNo ratings yet
- Differential Gene Expression and DevelopmentDocument9 pagesDifferential Gene Expression and DevelopmentShaher Bano Mirza100% (1)
- Beyond DNA: The Epigenetic Revolution: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchFrom EverandBeyond DNA: The Epigenetic Revolution: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchNo ratings yet
- Beyond DNA: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchFrom EverandBeyond DNA: From Cellular Mechanisms to Environmental Factors: How Epigenetics Shapes Our Biological Destiny and its Implications for Health, Behavior, and the Future of ResearchNo ratings yet
- The Dental Pulp: Biology, Pathology, and Regenerative TherapiesFrom EverandThe Dental Pulp: Biology, Pathology, and Regenerative TherapiesNo ratings yet
- Diagnostic Facial Tri4Document17 pagesDiagnostic Facial Tri4Muhammad AsimNo ratings yet
- The ABO's 75th Anniversary: A Retrospective Glance at Progress in The Last Quarter CenturyDocument5 pagesThe ABO's 75th Anniversary: A Retrospective Glance at Progress in The Last Quarter CenturyMuhammad AsimNo ratings yet
- OrthodonticsDocument21 pagesOrthodonticsMuhammad AsimNo ratings yet
- Effect of Micro-Osteoperforations On The Rate of Tooth MovementDocument20 pagesEffect of Micro-Osteoperforations On The Rate of Tooth MovementMuhammad AsimNo ratings yet
- How Does The Amount of Surgical Insult Affect Bone Around Moving Teeth?Document8 pagesHow Does The Amount of Surgical Insult Affect Bone Around Moving Teeth?Muhammad AsimNo ratings yet
- Effect of Piezopuncture On Tooth Movement and Bone Remodeling in DogsDocument26 pagesEffect of Piezopuncture On Tooth Movement and Bone Remodeling in DogsMuhammad AsimNo ratings yet
- Adult OrthodonticsDocument16 pagesAdult OrthodonticsMuhammad AsimNo ratings yet
- Comparative Distalization Effects of Jones Jig and PendulumDocument30 pagesComparative Distalization Effects of Jones Jig and PendulumMuhammad AsimNo ratings yet
- Orthodontic Basic Priciples in Treatment PlanningDocument61 pagesOrthodontic Basic Priciples in Treatment PlanningMuhammad Asim100% (2)
- MVUPGO 28 (Embryology)Document41 pagesMVUPGO 28 (Embryology)Muhammed AbdulmajeedNo ratings yet
- Teacher: DR Gouri Course: Zoology Hons. 6 Sem Paper: Developmental BiologyDocument17 pagesTeacher: DR Gouri Course: Zoology Hons. 6 Sem Paper: Developmental BiologyApratim SinghNo ratings yet
- Stem Cell Research Visual AidDocument8 pagesStem Cell Research Visual Aidapi-376567878No ratings yet
- Human Genome Project: Prepared By: LJ SilvestreDocument28 pagesHuman Genome Project: Prepared By: LJ SilvestreLJ SilvestreNo ratings yet
- Grade 5 LP4 Reproduction Sept24 PDFDocument27 pagesGrade 5 LP4 Reproduction Sept24 PDFWyenne Rose TorradoNo ratings yet
- Embryology TerminologiesDocument2 pagesEmbryology TerminologiesSomaya QuaziNo ratings yet
- Instant Download Ebook PDF Biological Science 2nd Canadian Edition PDF ScribdDocument41 pagesInstant Download Ebook PDF Biological Science 2nd Canadian Edition PDF Scribdmatthew.weimar370100% (40)
- Embryology: Ahlam HarasaniDocument67 pagesEmbryology: Ahlam HarasanibeylaNo ratings yet
- Plant Tissue CultureDocument26 pagesPlant Tissue CultureNAVEENANo ratings yet
- L6-Steczina, Sonette BIOEN345 Lecture 20200415Document22 pagesL6-Steczina, Sonette BIOEN345 Lecture 20200415asdfghjklNo ratings yet
- The Ethics of CloningDocument5 pagesThe Ethics of CloningUpai MbembNo ratings yet
- Training Manual On Vegetable Seed Treatment and ConditioningDocument38 pagesTraining Manual On Vegetable Seed Treatment and ConditioningMuhammad Boota SarwarNo ratings yet
- Cardiac RegenerationDocument5 pagesCardiac RegenerationBenyam ZenebeNo ratings yet
- Hall, Young, Kenway - 2002 - Manual For The Determination of Egg Fertility in Penaeus Monodon - PDF TraducidoDocument54 pagesHall, Young, Kenway - 2002 - Manual For The Determination of Egg Fertility in Penaeus Monodon - PDF Traducidoelina del mar rodriguez barbosaNo ratings yet
- Tudung AkarDocument17 pagesTudung AkarSyifa SariNo ratings yet
- Chapter 14 - Reproduction in HumansDocument44 pagesChapter 14 - Reproduction in HumansJnr DogheadNo ratings yet
- EmbryologyDocument61 pagesEmbryologyAmritha Anand100% (2)
- Early Development and Axis Formation in Amphibians and FishDocument3 pagesEarly Development and Axis Formation in Amphibians and FishJohnkevin NocheNo ratings yet
- 12 Biology NcertSolutions Chapter 3Document11 pages12 Biology NcertSolutions Chapter 3vijayan vijayanNo ratings yet
- 2.1 One Pager Female Reproduction MEMODocument1 page2.1 One Pager Female Reproduction MEMORudzi UdziNo ratings yet
- Stem CellsDocument20 pagesStem CellsalibrownNo ratings yet
- Dr. Bapi Ghosh Assistant Professor Department of Botany (Dodl)Document45 pagesDr. Bapi Ghosh Assistant Professor Department of Botany (Dodl)Project ICTNo ratings yet
- Fluorescence Activated Cell SortingDocument6 pagesFluorescence Activated Cell SortingAjit YadavNo ratings yet
- Gut Tube and Body CavitiesDocument22 pagesGut Tube and Body CavitiesKeysha YbanezNo ratings yet
- Embryology MCQ ReviewDocument58 pagesEmbryology MCQ ReviewAriba Asif100% (1)
- Nómina Embriológica-2006 PDFDocument47 pagesNómina Embriológica-2006 PDFTom ArrolloNo ratings yet
- Somatic Embryogenesis NotesDocument3 pagesSomatic Embryogenesis NotesjyomamsNo ratings yet
- Developmental Biology GametogenesisDocument14 pagesDevelopmental Biology Gametogenesispragyamaharjan2No ratings yet