You might also like
- Biografia de Charles DarwinDocument4 pagesBiografia de Charles DarwinÄdëlä Gönzälëz100% (5)
- Lesiones de La Mano y MuñecaDocument11 pagesLesiones de La Mano y MuñecaAdriana Magaly Fernandez Laconich100% (1)
- Escalas Del DolorDocument34 pagesEscalas Del DolorFabiola Garcia100% (1)
- 2 DICCIONARIO FISIOTERAPEUTICOkatoDocument19 pages2 DICCIONARIO FISIOTERAPEUTICOkatoJonathan Kato Tintaya TintayaNo ratings yet
- BIORRITMODocument44 pagesBIORRITMOIbeth Liliana J. Calderón100% (1)
- Constancia de Trabajo - CasDocument16 pagesConstancia de Trabajo - CasOscar Ramirez100% (1)
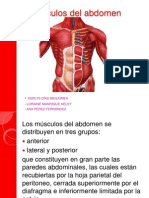
- Musculos Del AbdomenDocument13 pagesMusculos Del AbdomenGustavo Robelly100% (1)
- Bases Anatomia FisiologiaDocument28 pagesBases Anatomia Fisiologiacelso1966No ratings yet
- TEMA 4.LAS CARACTERISTICAS DEL MOVIMIENTO Y LA BIOMECÁNICAdocDocument13 pagesTEMA 4.LAS CARACTERISTICAS DEL MOVIMIENTO Y LA BIOMECÁNICAdocGabriel Blanco GarridoNo ratings yet
- Genética y Su ImportanciaDocument61 pagesGenética y Su ImportanciaDaniellaNo ratings yet
- Actividad 7 Francisco Javier Díaz González PDFDocument1 pageActividad 7 Francisco Javier Díaz González PDFJDNo ratings yet
- Rcqconsolidación Del Hueso Mark DuttonDocument61 pagesRcqconsolidación Del Hueso Mark DuttonAldo NavarreteNo ratings yet
- Articulo AmputadoDocument10 pagesArticulo AmputadoLtf Oswaldo Guerrero IslasNo ratings yet
- Estimulacion Electrica para Contraccion MuscularDocument90 pagesEstimulacion Electrica para Contraccion MuscularPILAR HERRERANo ratings yet
- Contreras, J. (Comp.) - Alimentación y Cultura. Necesidades, Gustos y CostumbresDocument367 pagesContreras, J. (Comp.) - Alimentación y Cultura. Necesidades, Gustos y CostumbresRael ReaNo ratings yet
- ProteomicaDocument154 pagesProteomicaSangel López100% (1)
- La Movilidad ArticularDocument4 pagesLa Movilidad ArticularDilmar Peraza Dominguez100% (1)
- Enzimas: cómo funcionanDocument13 pagesEnzimas: cómo funcionanRicardo FloresNo ratings yet
- Parafina, Fluidoterapia y UltrasonidoDocument19 pagesParafina, Fluidoterapia y UltrasonidoLuisa Bautista CampanurNo ratings yet
- Órganos de Los SentidosDocument9 pagesÓrganos de Los SentidosFacsury Sanjuan MezaNo ratings yet
- El Modo de Ser Propio Del HombreDocument77 pagesEl Modo de Ser Propio Del HombreXavier Vargas Beal100% (3)
- Beneficios de Las Enzimas en El Organismo PDFDocument7 pagesBeneficios de Las Enzimas en El Organismo PDFNoemi Torrez100% (1)
- Manual Manejo Avanzado Tejidos BlandosDocument92 pagesManual Manejo Avanzado Tejidos Blandoskoke423No ratings yet
- Practica 1 Reporte PDFDocument10 pagesPractica 1 Reporte PDFNatalia J PadillaNo ratings yet
- Responsabilidad GerencialDocument13 pagesResponsabilidad GerencialliliNo ratings yet
- Tesis Etnografia de AyutuxtepequeDocument110 pagesTesis Etnografia de AyutuxtepequeNaty Navarrete100% (1)
- Quiste GanglionarDocument10 pagesQuiste GanglionarAlejandro Morales100% (1)
- Presentacion FisioDocument11 pagesPresentacion FisioDiana CastilloNo ratings yet
- Coordinacion y Equilibrio / FisiologiaDocument2 pagesCoordinacion y Equilibrio / FisiologiaPaoNo ratings yet
- Glosario para Leer El Arbol Del ConocimientoDocument28 pagesGlosario para Leer El Arbol Del ConocimientoEnid Sofía Zúñiga MurilloNo ratings yet
- CadenasCinemáticasDocument15 pagesCadenasCinemáticasgenesis gonzalezNo ratings yet
- Estructura y Funcion ArticularDocument48 pagesEstructura y Funcion ArticularDimitriNo ratings yet
- Enzimas ReguladorasDocument4 pagesEnzimas ReguladorasLucas Tapia Parra0% (1)
- Cuerpo HumanoDocument59 pagesCuerpo HumanoVíctor CatalàNo ratings yet
- Arteria SubclaviaDocument37 pagesArteria SubclaviaCésar Martínez MorenoNo ratings yet
- CFC - Músculos Del AbdomenDocument57 pagesCFC - Músculos Del AbdomenjuanNo ratings yet
- Qué Es La KinesiologíaDocument2 pagesQué Es La Kinesiologíacristian f plataNo ratings yet
- El Aplastamiento VertebralDocument2 pagesEl Aplastamiento VertebralWilly PuenteNo ratings yet
- Maniobra de Nelaton para reducir luxación ATMDocument13 pagesManiobra de Nelaton para reducir luxación ATMjunior tenicela torres100% (1)
- TermoterapiaDocument8 pagesTermoterapiaNestor HernandezNo ratings yet
- Descanso, sueño y alivio del dolor: factores y fasesDocument17 pagesDescanso, sueño y alivio del dolor: factores y fasesEver Romero100% (1)
- Músculos antebrazo manoDocument16 pagesMúsculos antebrazo manoEmberson Vela GavilanesNo ratings yet
- Región GlúteaDocument23 pagesRegión GlúteaBrianda MoralesNo ratings yet
- Biografia de Angel Dias Barriga CasaresDocument108 pagesBiografia de Angel Dias Barriga CasaresmarcosintelNo ratings yet
- FNPDocument7 pagesFNPIsrael PapinuttiNo ratings yet
- Alteración de La Piel # 2: Los GluteosDocument28 pagesAlteración de La Piel # 2: Los GluteosJuan GarciaNo ratings yet
- Introducción A La Terapia Física-ApunteDocument31 pagesIntroducción A La Terapia Física-ApunteJC XmanNo ratings yet
- BIOMEC-CLAS-001 IntroducciónDocument46 pagesBIOMEC-CLAS-001 IntroduccióndanNo ratings yet
- Cadena GanglionarDocument17 pagesCadena GanglionarYovana Carolina Corimanya MoranNo ratings yet
- Estiramiento MuscularDocument17 pagesEstiramiento MuscularOctimuzzNo ratings yet
- Cambio PosturalDocument9 pagesCambio PosturalHenry BarrenoNo ratings yet
- Evaluación de MovilidadDocument14 pagesEvaluación de MovilidadFlorChepeChumpitaz100% (1)
- Guia Reflejos 2020Document13 pagesGuia Reflejos 2020Esteban Araya AcevedoNo ratings yet
- Evaluación Módulo 3 Curso Masoterapia GeneralDocument2 pagesEvaluación Módulo 3 Curso Masoterapia GeneralIsabel Diaz100% (1)
- Espectro acústico zonas ultrasonidos efectos indicacionesDocument11 pagesEspectro acústico zonas ultrasonidos efectos indicacionesAlex Delgado RmzNo ratings yet
- 25.volumenes y Capacidades PulmonaresDocument28 pages25.volumenes y Capacidades Pulmonarestkmon22No ratings yet
- Músculos abdominales: grupos, funciones y descripciónDocument13 pagesMúsculos abdominales: grupos, funciones y descripciónKevin Manrique100% (1)
- Lesiones Del HombroDocument24 pagesLesiones Del HombroMariano Leiva100% (2)
- Planos anatómicos referencias espacialesDocument8 pagesPlanos anatómicos referencias espacialesSergio Franco RodríguezNo ratings yet
- Músculos Del PiéDocument3 pagesMúsculos Del PiéBriggi CastilloNo ratings yet
- Biomecanica Del CodoDocument15 pagesBiomecanica Del CodoBryan FreireNo ratings yet
- Fundamentos en FisioterapiaDocument8 pagesFundamentos en FisioterapiaOscar Pinto MorenoNo ratings yet
- Expo Mckenzie KinesioDocument24 pagesExpo Mckenzie KinesioYurii Gutiérrez SolanoNo ratings yet
- Actividad Física y Dosificación DelDocument22 pagesActividad Física y Dosificación DelYohary Andres Robles MorenoNo ratings yet
- ApuntesDocument175 pagesApuntesMerche BorregueroNo ratings yet
- Biomecánica del hueso en la Universidad Nacional de TrujilloDocument22 pagesBiomecánica del hueso en la Universidad Nacional de TrujilloAnthonyVelasquezMirandaNo ratings yet
- Curso de Cinesiterapia: Especialidad FisioterapiaDocument18 pagesCurso de Cinesiterapia: Especialidad FisioterapiaJesús MorenoNo ratings yet
- Practica 7 EcologíaDocument7 pagesPractica 7 EcologíaCarlos CőrdovaNo ratings yet
- Principios Generales de Biología de La ConservaciónDocument11 pagesPrincipios Generales de Biología de La ConservaciónDan Pinzón-GómezNo ratings yet
- INFORME DS-076-2020-Hospital ll-1Document18 pagesINFORME DS-076-2020-Hospital ll-1Oscar RamirezNo ratings yet
- Proyecto de Investigación - Oscar Ramirez de La CruzDocument75 pagesProyecto de Investigación - Oscar Ramirez de La CruzOscar RamirezNo ratings yet
- Monografia - Gerencia EmpresarialDocument2 pagesMonografia - Gerencia EmpresarialOscar RamirezNo ratings yet
- 2.-Elaborando Programa de Ventas PDFDocument3 pages2.-Elaborando Programa de Ventas PDFOscar RamirezNo ratings yet
- Articulo Científico - Oscar Ramirez-1Document12 pagesArticulo Científico - Oscar Ramirez-1Oscar RamirezNo ratings yet
- 1.-Programa de Servicio Social de Atención Primaria de La Salud1 PDFDocument10 pages1.-Programa de Servicio Social de Atención Primaria de La Salud1 PDFOscar RamirezNo ratings yet
- 55%-Guardias - Progreso - Febrero - Marzo - 2017Document6 pages55%-Guardias - Progreso - Febrero - Marzo - 2017Oscar RamirezNo ratings yet
- Articulo Científico - Oscar Ramirez-1Document12 pagesArticulo Científico - Oscar Ramirez-1Oscar RamirezNo ratings yet
- Catálogo de Perfiles de Puestos-4Document646 pagesCatálogo de Perfiles de Puestos-4Oscar RamirezNo ratings yet
- La Planificacion EmpresarialDocument16 pagesLa Planificacion EmpresarialNoe QRNo ratings yet
- Planeacion Estrategica PDFDocument11 pagesPlaneacion Estrategica PDFjagomegaNo ratings yet
- Actividad I - Fuerza de VentasDocument5 pagesActividad I - Fuerza de VentasOscar RamirezNo ratings yet
- Gestión de calidad y servicio al cliente en cooperativa de TocacheDocument2 pagesGestión de calidad y servicio al cliente en cooperativa de TocacheOscar RamirezNo ratings yet
- Practicas Preprofesionales II-Administracion.Document55 pagesPracticas Preprofesionales II-Administracion.Oscar RamirezNo ratings yet
- Urgencias NeurologicasDocument47 pagesUrgencias NeurologicasOscar RamirezNo ratings yet
- Diabetes, Que ComerDocument3 pagesDiabetes, Que ComerOscar RamirezNo ratings yet
- Evolucion de Auditoria AdministrativaDocument23 pagesEvolucion de Auditoria AdministrativaOscar Ramirez100% (2)
- Teoria EcologicaDocument8 pagesTeoria EcologicaOscar RamirezNo ratings yet
- Teoria EcologicaDocument8 pagesTeoria EcologicaOscar RamirezNo ratings yet
- Texto Liderazgo II AdmDocument144 pagesTexto Liderazgo II AdmOscar RamirezNo ratings yet
- Es Tic, Ventajas y DesventajasDocument6 pagesEs Tic, Ventajas y DesventajasOscar RamirezNo ratings yet
- Directiva General Del Proceso de Planeamiento Estratégico - Sistema Nacional de Planeamiento Estratégico"-SINAPDocument24 pagesDirectiva General Del Proceso de Planeamiento Estratégico - Sistema Nacional de Planeamiento Estratégico"-SINAPTeresa Clotilde Ojeda SánchezNo ratings yet
- Diabetes, Que ComerDocument3 pagesDiabetes, Que ComerOscar RamirezNo ratings yet
- MAPA ConceptualDocument2 pagesMAPA ConceptualOscar RamirezNo ratings yet
- MOF Informatica 2014Document13 pagesMOF Informatica 2014Oscar RamirezNo ratings yet
- Propiedades y beneficios de las Hojas de CocaDocument6 pagesPropiedades y beneficios de las Hojas de CocaOscar RamirezNo ratings yet
- Documento Tecnico F 18 11 2014 PDFDocument96 pagesDocumento Tecnico F 18 11 2014 PDFOscar RamirezNo ratings yet
- Planeamiento Multianual Inversiones San MartinDocument19 pagesPlaneamiento Multianual Inversiones San MartinOscar RamirezNo ratings yet
- Glándula pineal: aspectos anatómicos y funciones de la melatoninaDocument14 pagesGlándula pineal: aspectos anatómicos y funciones de la melatoninaVerónica Isabel Rincón BelandriaNo ratings yet
- M16S1AI1Document3 pagesM16S1AI1manu bbNo ratings yet
- ANALISIS - DE - MICROORGANISMOS - AEROBIOS Practica 4Document2 pagesANALISIS - DE - MICROORGANISMOS - AEROBIOS Practica 4jenny paredes varelaNo ratings yet
- Foro 12 Sistema EndocrinoDocument8 pagesForo 12 Sistema EndocrinoAslhy Lizetti Hidalgo ArcelaNo ratings yet
- Desde Que El Hombre Tuvo La Capacidad de Pesar y de RazonarDocument4 pagesDesde Que El Hombre Tuvo La Capacidad de Pesar y de RazonarPlacido PajaNo ratings yet
- Daniel Felipe Angaita B.: Virus Del Papiloma HumanoDocument8 pagesDaniel Felipe Angaita B.: Virus Del Papiloma Humanoluz stella ballesteros duarteNo ratings yet
- Conciencia y Azar PDFDocument76 pagesConciencia y Azar PDFedgardo2050No ratings yet
- Problematica Medicolegal Del Shock AnafilacticoDocument10 pagesProblematica Medicolegal Del Shock AnafilacticoElvia RendónNo ratings yet
- Colegio La Sagrda Familia Examen de Octavo 2 PeriodoDocument3 pagesColegio La Sagrda Familia Examen de Octavo 2 PeriodoColegio La Sagrada FamiliaNo ratings yet
- Postulado de La Teoría CelularDocument3 pagesPostulado de La Teoría CelularPaula Pineda de NavasNo ratings yet
- Miastenia Gravis y MiopatiasDocument16 pagesMiastenia Gravis y MiopatiasBryant Manuel Ortiz AquinoNo ratings yet
- PROYECTO 1. Descubro Mi Cuerpo. Parte 1Document11 pagesPROYECTO 1. Descubro Mi Cuerpo. Parte 1Cecy SolanoNo ratings yet
- Panleucopenia FelinaDocument2 pagesPanleucopenia FelinaDavid ElihúNo ratings yet
- Coadyuvantes AgricolasDocument5 pagesCoadyuvantes AgricolasmantecoNo ratings yet
- Adaptaciones Biologicas DécimoDocument6 pagesAdaptaciones Biologicas DécimoLuismi BNo ratings yet
- Anexos Micro 2Document11 pagesAnexos Micro 2Bladimir Jesús Ccama CutipaNo ratings yet
- Planificacion AnualDocument5 pagesPlanificacion AnualChoni RescuerNo ratings yet
- Introductorio - Histología HumanaDocument6 pagesIntroductorio - Histología HumanaVíctor Seminario MarceloNo ratings yet
- Practica N 8 Meristema y ParenquimaDocument4 pagesPractica N 8 Meristema y ParenquimaBuka JosueNo ratings yet
- Injerto de PielDocument24 pagesInjerto de PielJosé Juan Mtz ChzNo ratings yet
- Enfermedades EndocrinasDocument5 pagesEnfermedades EndocrinashectorNo ratings yet
- Preguntas Cortas Estructura BioquímicaDocument6 pagesPreguntas Cortas Estructura BioquímicafuggiaNo ratings yet