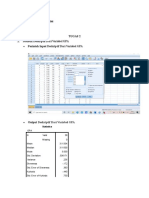
You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Seminar, Workshop, ConferenceDocument30 pagesSeminar, Workshop, ConferenceMutharasu SNo ratings yet
- Koine GreekDocument226 pagesKoine GreekΜάριος Αθανασίου100% (6)
- Stars and Galaxies 9th Edition Seeds Test BankDocument20 pagesStars and Galaxies 9th Edition Seeds Test Bankngocalmai0236h100% (32)
- His To GramsDocument15 pagesHis To GramsMaryam HasanNo ratings yet
- SuperDeck All ModelsDocument12 pagesSuperDeck All Modelsarthur chungNo ratings yet
- Curved Path of Electron A B: ST Mary's College 1Document5 pagesCurved Path of Electron A B: ST Mary's College 1elsie limNo ratings yet
- Panasonic MINAS A5Document478 pagesPanasonic MINAS A5Vaikuntam Ramamurthy100% (1)
- Din 2095Document42 pagesDin 2095Eduardo ValenciaNo ratings yet
- Nama: Ketut Dian Caturini NIM: 1813011007 Kelas: 7B Tugas 2Document3 pagesNama: Ketut Dian Caturini NIM: 1813011007 Kelas: 7B Tugas 2Sri AgustiniNo ratings yet
- DC Lecture Slides 1 - Information TheoryDocument22 pagesDC Lecture Slides 1 - Information TheoryG VenkateshNo ratings yet
- Money Calculations (F)Document14 pagesMoney Calculations (F)Pavan GhelaniNo ratings yet
- Mole Day Project Report RevisedDocument5 pagesMole Day Project Report Revisedapi-267301173No ratings yet
- Normal Motor and Sensory Nerve Conduction VelocityDocument3 pagesNormal Motor and Sensory Nerve Conduction Velocitycemm_11No ratings yet
- Wasif CVDocument2 pagesWasif CVTalha BaigNo ratings yet
- Mesh Based Multicast Routing ProtocolDocument10 pagesMesh Based Multicast Routing ProtocolArul JothiNo ratings yet
- Tetzaveh Parsha GematriaDocument1 pageTetzaveh Parsha GematriaRabbi Benyomin HoffmanNo ratings yet
- 2009-09-06 181738 Linsw RegDocument67 pages2009-09-06 181738 Linsw Regam1liNo ratings yet
- Heat & Mass Transfer PDFDocument2 pagesHeat & Mass Transfer PDFabyabraham_nytro50% (6)
- Carbon Compounds: Standard/ Class/ Grade - 10 SSC, CBSE - 8 ICSEDocument53 pagesCarbon Compounds: Standard/ Class/ Grade - 10 SSC, CBSE - 8 ICSEsaintEmNo ratings yet
- Aksa Ajd170Document5 pagesAksa Ajd170mhmmd14No ratings yet
- False: True True True TrueDocument2 pagesFalse: True True True TrueSuubi brianNo ratings yet
- Slab DesignDocument96 pagesSlab Designdilrangi100% (2)
- Properties of Solutions: Electrolytes and Non-Electrolytes by Dan HolmquistDocument4 pagesProperties of Solutions: Electrolytes and Non-Electrolytes by Dan HolmquistPaul Schumann50% (2)
- DeskView Client 6 45 enDocument166 pagesDeskView Client 6 45 enRazvan22081997No ratings yet
- SAE-J1850 Communication Protocol Conformity Transmission Controller For Automotive LANDocument59 pagesSAE-J1850 Communication Protocol Conformity Transmission Controller For Automotive LANMa Ngoc TrungNo ratings yet
- Sap Basis Transaction CodesDocument2 pagesSap Basis Transaction CodeskatrinbreaksNo ratings yet
- Market Manipulation Rules and IPO Underpricing: Huu - Duong@monash - EduDocument54 pagesMarket Manipulation Rules and IPO Underpricing: Huu - Duong@monash - EduTI Logic ?? sobayedNo ratings yet
- BXE Experiment No.3Document8 pagesBXE Experiment No.3DsgawaliNo ratings yet
- T60 Transformer Management Relay: UR Series Instruction ManualDocument404 pagesT60 Transformer Management Relay: UR Series Instruction Manualeng amadaNo ratings yet
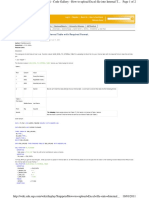
- How To Upload Excel File Into Internal Table With Required FormatDocument2 pagesHow To Upload Excel File Into Internal Table With Required FormatErick ViteNo ratings yet