You might also like
- Non-Target Effects of Clothianidin On Monarch ButterfliesDocument4 pagesNon-Target Effects of Clothianidin On Monarch ButterfliesBlue Dasher FarmNo ratings yet
- Trading Biodiversity For Pest Problems: ResearcharticleDocument5 pagesTrading Biodiversity For Pest Problems: ResearcharticleBlue Dasher FarmNo ratings yet
- Understanding and Enhancing Soil Biological Health: The Solution For Reversing Soil DegradationDocument40 pagesUnderstanding and Enhancing Soil Biological Health: The Solution For Reversing Soil DegradationBlue Dasher FarmNo ratings yet
- Insecticide Use and Crop Selection in Regions With High GM Adoption RatesDocument11 pagesInsecticide Use and Crop Selection in Regions With High GM Adoption RatesBlue Dasher FarmNo ratings yet
- Tracking Predation of Subterranean Pests: Digestion of Corn Rootworm DNA by A Generalist MiteDocument5 pagesTracking Predation of Subterranean Pests: Digestion of Corn Rootworm DNA by A Generalist MiteBlue Dasher FarmNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- 70 Valves SolenoidDocument105 pages70 Valves SolenoidrizalNo ratings yet
- The JHipster Mini Book 2Document129 pagesThe JHipster Mini Book 2tyulist100% (1)
- Stearns 87700 Series Parts ListDocument4 pagesStearns 87700 Series Parts ListYorkistNo ratings yet
- IKEA SHANGHAI Case StudyDocument5 pagesIKEA SHANGHAI Case StudyXimo NetteNo ratings yet
- 2021-03 Trophy LagerDocument11 pages2021-03 Trophy LagerAderayo OnipedeNo ratings yet
- Existential ThreatsDocument6 pagesExistential Threatslolab_4No ratings yet
- Android Developer PDFDocument2 pagesAndroid Developer PDFDarshan ChakrasaliNo ratings yet
- Monergism Vs SynsergismDocument11 pagesMonergism Vs SynsergismPam AgtotoNo ratings yet
- Oracle Forms & Reports 12.2.1.2.0 - Create and Configure On The OEL 7Document50 pagesOracle Forms & Reports 12.2.1.2.0 - Create and Configure On The OEL 7Mario Vilchis Esquivel100% (1)
- Space Hulk - WDDocument262 pagesSpace Hulk - WDIgor Baranenko100% (1)
- Career Level Diagram - V5Document1 pageCareer Level Diagram - V5Shivani RaikwarNo ratings yet
- Lesson PlanDocument2 pagesLesson Plannicole rigonNo ratings yet
- Recommendations For Students With High Functioning AutismDocument7 pagesRecommendations For Students With High Functioning AutismLucia SaizNo ratings yet
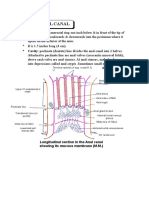
- Anatomy Anal CanalDocument14 pagesAnatomy Anal CanalBela Ronaldoe100% (1)
- Chapter13 PDFDocument34 pagesChapter13 PDFAnastasia BulavinovNo ratings yet
- 74HC00D 74HC00D 74HC00D 74HC00D: CMOS Digital Integrated Circuits Silicon MonolithicDocument8 pages74HC00D 74HC00D 74HC00D 74HC00D: CMOS Digital Integrated Circuits Silicon MonolithicAssistec TecNo ratings yet
- The Use of Air Cooled Heat Exchangers in Mechanical Seal Piping Plans - SnyderDocument7 pagesThe Use of Air Cooled Heat Exchangers in Mechanical Seal Piping Plans - SnyderJaime Ocampo SalgadoNo ratings yet
- How To Identify MQ Client Connections and Stop ThemDocument26 pagesHow To Identify MQ Client Connections and Stop ThemPurushotham100% (1)
- in Strategic Management What Are The Problems With Maintaining A High Inventory As Experienced Previously With Apple?Document5 pagesin Strategic Management What Are The Problems With Maintaining A High Inventory As Experienced Previously With Apple?Priyanka MurthyNo ratings yet
- Review1 ScheduleDocument3 pagesReview1 Schedulejayasuryam.ae18No ratings yet
- Electro Fashion Sewable LED Kits WebDocument10 pagesElectro Fashion Sewable LED Kits WebAndrei VasileNo ratings yet
- (500eboard) Version Coding Model 140 As of MY 1995Document1 page(500eboard) Version Coding Model 140 As of MY 1995Saimir SaliajNo ratings yet
- Psychological Contract Rousseau PDFDocument9 pagesPsychological Contract Rousseau PDFSandy KhanNo ratings yet
- DQ Vibro SifterDocument13 pagesDQ Vibro SifterDhaval Chapla67% (3)
- g6 - AFA - Q1 - Module 6 - Week 6 FOR TEACHERDocument23 pagesg6 - AFA - Q1 - Module 6 - Week 6 FOR TEACHERPrincess Nicole LugtuNo ratings yet
- Week 7Document24 pagesWeek 7Priyank PatelNo ratings yet
- MRBR ATR 72 Rev18Document424 pagesMRBR ATR 72 Rev18Juan Forero OrtizNo ratings yet
- Ib History Command Term PostersDocument6 pagesIb History Command Term Postersapi-263601302100% (4)
- Chronic Kidney DiseaseDocument15 pagesChronic Kidney Diseaseapi-270623039No ratings yet
- UTP Student Industrial ReportDocument50 pagesUTP Student Industrial ReportAnwar HalimNo ratings yet