You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Heart Anatomy WorksheetDocument3 pagesHeart Anatomy WorksheetUSMP FN ARCHIVOS100% (2)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Hospital Corpsman ManualDocument574 pagesHospital Corpsman ManualNes Navarro100% (1)
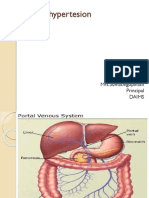
- Portal HypertensionDocument23 pagesPortal HypertensionSumathi GopinathNo ratings yet
- Assessment and Management of Women During Post-Natal PeriodDocument82 pagesAssessment and Management of Women During Post-Natal Periodsweta0% (1)
- Tabang Elementary School Science 4 Test on Digestion and Internal OrgansDocument6 pagesTabang Elementary School Science 4 Test on Digestion and Internal OrgansJanice Corañes100% (5)
- Marching Procedure For Tensegrity 2007Document26 pagesMarching Procedure For Tensegrity 2007Tensegrity WikiNo ratings yet
- Kent TensegritreeDocument12 pagesKent TensegritreeTensegrity WikiNo ratings yet
- Tensegrity Beam Wave Dynamics 2019Document20 pagesTensegrity Beam Wave Dynamics 2019Tensegrity WikiNo ratings yet
- NumerNumerical Algorithm For Tensegrity DynamicsDocument7 pagesNumerNumerical Algorithm For Tensegrity DynamicsTensegrity WikiNo ratings yet
- Tensegrity Systems Short Course Outline by Skelton, de OliveiraDocument3 pagesTensegrity Systems Short Course Outline by Skelton, de OliveiraTensegrity WikiNo ratings yet
- Pinuad Dynamics CRCDocument62 pagesPinuad Dynamics CRCTensegrity WikiNo ratings yet
- Tensegity Deployment by MasicDocument9 pagesTensegity Deployment by MasicTensegrity WikiNo ratings yet
- Dynamics and Control of Tensegrity SystemsDocument22 pagesDynamics and Control of Tensegrity SystemsTensegrity WikiNo ratings yet
- Growth-Adapted Tensegrity Structures Longman Skelton 2014Document147 pagesGrowth-Adapted Tensegrity Structures Longman Skelton 2014Tensegrity WikiNo ratings yet
- Biological Tensegrity Red Blood CellsDocument21 pagesBiological Tensegrity Red Blood CellsTensegrity Wiki100% (1)
- Path Planning For Tensegrity Deployment by Pinauda, Masica, SkeltonDocument12 pagesPath Planning For Tensegrity Deployment by Pinauda, Masica, SkeltonTensegrity WikiNo ratings yet
- Workshop Outline For Workshop On Tensegrity Systems With Skelton, de OliveiraDocument2 pagesWorkshop Outline For Workshop On Tensegrity Systems With Skelton, de OliveiraTensegrity WikiNo ratings yet
- Han LMI Struct Linear Cdc03Document6 pagesHan LMI Struct Linear Cdc03Tensegrity WikiNo ratings yet
- Weiwei FSN CDC42Document6 pagesWeiwei FSN CDC42Tensegrity WikiNo ratings yet
- Optimal Topology and Geometry For Controllable Tensegrity Systems by Jager, Masic, SkeltonDocument6 pagesOptimal Topology and Geometry For Controllable Tensegrity Systems by Jager, Masic, SkeltonTensegrity WikiNo ratings yet
- Prestress Tensegrity Structures by Masic, Skelton 2005Document16 pagesPrestress Tensegrity Structures by Masic, Skelton 2005Tensegrity WikiNo ratings yet
- Sensor or Actuator Selection by Skelton, Lia 2006Document8 pagesSensor or Actuator Selection by Skelton, Lia 2006Tensegrity WikiNo ratings yet
- Optimization of Class 2 Tensegrity Towers by Masic, SkeltonDocument12 pagesOptimization of Class 2 Tensegrity Towers by Masic, SkeltonTensegrity WikiNo ratings yet
- Equilibrium Conditions of A Tensegrity Structure by Williamson, Skelton, Han 2002Document18 pagesEquilibrium Conditions of A Tensegrity Structure by Williamson, Skelton, Han 2002Tensegrity WikiNo ratings yet
- An LMI Optimization Approach To The Design of Structured Linear ControllersDocument8 pagesAn LMI Optimization Approach To The Design of Structured Linear ControllersTensegrity WikiNo ratings yet
- Tensegrity Structures Prestressability Sultan, Skelton 2001Document16 pagesTensegrity Structures Prestressability Sultan, Skelton 2001Tensegrity WikiNo ratings yet
- Tensegrity Telescope DesignDocument2 pagesTensegrity Telescope DesignTensegrity WikiNo ratings yet
- Equilibrium Conditions of Class 1 Tensegrity Structures by Williamson, Skelton, HanDocument20 pagesEquilibrium Conditions of Class 1 Tensegrity Structures by Williamson, Skelton, HanTensegrity WikiNo ratings yet
- Intro To Mechanics of Tensegrity StructuresDocument6 pagesIntro To Mechanics of Tensegrity StructuresTensegrity WikiNo ratings yet
- Equilibrium Conditions of A Tensegrity StructureDocument21 pagesEquilibrium Conditions of A Tensegrity StructureTensegrity WikiNo ratings yet
- Control Synthesis For A Class of Light Robotic Tensegrity StructuresDocument7 pagesControl Synthesis For A Class of Light Robotic Tensegrity StructuresTensegrity WikiNo ratings yet
- Active Vibration Control of A Three-Stage Tensegrity Structure by Chan, Arbelaez, Bossens, SkeltonDocument7 pagesActive Vibration Control of A Three-Stage Tensegrity Structure by Chan, Arbelaez, Bossens, SkeltonTensegrity WikiNo ratings yet
- Stiffness of Planar Tensegrity Truss Topologies by Jager, Skelton 2006Document23 pagesStiffness of Planar Tensegrity Truss Topologies by Jager, Skelton 2006Tensegrity WikiNo ratings yet
- Optimization of Tensegrity Structures by Masic, Skelton, GillDocument17 pagesOptimization of Tensegrity Structures by Masic, Skelton, GillTensegrity WikiNo ratings yet
- Zoology - Lab ReportDocument3 pagesZoology - Lab ReportRonald Loreto MabajenNo ratings yet
- The Structure and Function of the Human HeartDocument22 pagesThe Structure and Function of the Human HeartNunoAlmeidaNo ratings yet
- Physiology of The Gastrointestinal Tract (Git)Document98 pagesPhysiology of The Gastrointestinal Tract (Git)jballungayNo ratings yet
- G6-Lesson PlanDocument4 pagesG6-Lesson PlanHansel RamosNo ratings yet
- Cell Structure and FunctionDocument10 pagesCell Structure and FunctionKanaga ValliNo ratings yet
- Test Bank For Understanding Current Procedural Terminology and Hcpcs Coding Systems 6th Edition by BowieDocument10 pagesTest Bank For Understanding Current Procedural Terminology and Hcpcs Coding Systems 6th Edition by Bowiepatriciajonesewrkbtsnfg100% (15)
- Maternal Ob NotesDocument103 pagesMaternal Ob NotesBoris OrbetaNo ratings yet
- Human Body Review TestDocument6 pagesHuman Body Review TestMrCiambroneNo ratings yet
- Clotting Factors List and Mnemonics For Easy Learning - All Medical StuffDocument10 pagesClotting Factors List and Mnemonics For Easy Learning - All Medical Stuffmonika ramosNo ratings yet
- Micr Biology: Chapter 17, Part ADocument17 pagesMicr Biology: Chapter 17, Part ARiaAfriyantiNo ratings yet
- Movement Organs in Humans and AnimalsDocument15 pagesMovement Organs in Humans and AnimalsDara FatimahNo ratings yet
- Science Grade 3 Life Unit 3L1Document22 pagesScience Grade 3 Life Unit 3L1Cristina Isabel Saavedra CuellesNo ratings yet
- Essay ImmuneDocument8 pagesEssay ImmuneAishah Najihah100% (1)
- Nesc 303 GuideDocument4 pagesNesc 303 GuideM MazloumNo ratings yet
- Oral & Maxillofacial Pathology II: Topic: Soft Tissue TumorsDocument123 pagesOral & Maxillofacial Pathology II: Topic: Soft Tissue TumorsDorina MalanceaNo ratings yet
- Nervous System-2022-23 Asi̇stDocument101 pagesNervous System-2022-23 Asi̇stYiğit GündüzNo ratings yet
- Animal Tissue - 2: Location FunctionDocument3 pagesAnimal Tissue - 2: Location FunctionKb AshishNo ratings yet
- Sci 5 PPT q2 WK 1 Day 1-5 (Autosaved)Document93 pagesSci 5 PPT q2 WK 1 Day 1-5 (Autosaved)renato.minaNo ratings yet
- Chapter12 Physical DevelopmentDocument38 pagesChapter12 Physical DevelopmentJhayCee100% (1)
- Fundamentals of Anatomy and Physiology: Course Description and ObjectivesDocument3 pagesFundamentals of Anatomy and Physiology: Course Description and Objectivesvamsi karna100% (1)
- Veterinary Comparative Anatomy - Exercise - The Integumentary SystemDocument6 pagesVeterinary Comparative Anatomy - Exercise - The Integumentary SystemYumeeeeeyNo ratings yet
- I-Platelet Pooling Set: Connect Up To 8 Buffy Coats Leukoreduction FilterDocument2 pagesI-Platelet Pooling Set: Connect Up To 8 Buffy Coats Leukoreduction FilterCampaign MediaNo ratings yet
- Protocol Book BWCT DominiqueS 2Document126 pagesProtocol Book BWCT DominiqueS 2N AaNo ratings yet
- Biology Mcqs Test For Entry Test by Dr. Sajid Ali TalpurDocument8 pagesBiology Mcqs Test For Entry Test by Dr. Sajid Ali TalpurDr. Sajid Ali Talpur100% (6)
- Strongylus (Uzal)Document16 pagesStrongylus (Uzal)Fredy MoralesNo ratings yet