You might also like
- 10 1152@japplphysiol 00907 2010Document11 pages10 1152@japplphysiol 00907 2010saulo freitas pereiraNo ratings yet
- Effects of Dietary Fat On Muscle Substrates,.11Document9 pagesEffects of Dietary Fat On Muscle Substrates,.11Saul NavaNo ratings yet
- Bio Qui MicaDocument9 pagesBio Qui MicaLuciana Santos da SilvaNo ratings yet
- Metabolic Adaptations To Endurance Training and NutritionDocument14 pagesMetabolic Adaptations To Endurance Training and NutritionValentina FernándezNo ratings yet
- Efectos Del Ayuno Previo Sobre La Oxidación de Grasas Durante El Ejercicio de ResistenciaDocument7 pagesEfectos Del Ayuno Previo Sobre La Oxidación de Grasas Durante El Ejercicio de ResistenciaRosaNo ratings yet
- Peri Workout NutritionDocument14 pagesPeri Workout NutritionLutz_KevinNo ratings yet
- Atualização em Nutrição EsportivaDocument42 pagesAtualização em Nutrição EsportivaHumberto GomesNo ratings yet
- Reducing The Intensity and Volume of Interval Training 1min X 1 MinDocument8 pagesReducing The Intensity and Volume of Interval Training 1min X 1 MinEduardo VieyraNo ratings yet
- Nutrition For The SprinterDocument12 pagesNutrition For The SprinterMarcelo NogueraNo ratings yet
- Articulo BioquimicaDocument11 pagesArticulo BioquimicaRafaAlvarezNo ratings yet
- Fat Loading For Endurance SportsDocument9 pagesFat Loading For Endurance SportsshaqtimNo ratings yet
- Effects of Protein Intake Prior To Carbohydrate-Restricted Endurance Exercise A Randomized Crossover TrialDocument13 pagesEffects of Protein Intake Prior To Carbohydrate-Restricted Endurance Exercise A Randomized Crossover TrialJuan Luis RuizNo ratings yet
- INGLES .. Mejor Rendimiento de Resistencia Mediante Periodización de La Ingesta de CHO.. Estrategia de ..SLEEP LOW..Document41 pagesINGLES .. Mejor Rendimiento de Resistencia Mediante Periodización de La Ingesta de CHO.. Estrategia de ..SLEEP LOW..Jony CostaNo ratings yet
- Adaptations To Skeletal Muscle With Endurance Exercise TrainingDocument5 pagesAdaptations To Skeletal Muscle With Endurance Exercise TrainingYo Vivo Fit Pablo y KarlaNo ratings yet
- Heavy-Resistance Exercise With or Without Nutritional Hormonal Responses To Consecutive Days ofDocument13 pagesHeavy-Resistance Exercise With or Without Nutritional Hormonal Responses To Consecutive Days ofmamithemadNo ratings yet
- High Intensity Aerobic TrainingDocument10 pagesHigh Intensity Aerobic TrainingdietabiosoficaNo ratings yet
- High-Intensity Interval Training: A Time-Efficient Strategy For Health Promotion?Document3 pagesHigh-Intensity Interval Training: A Time-Efficient Strategy For Health Promotion?neilh39No ratings yet
- The Role of Muscle Glycogen Content And.77Document48 pagesThe Role of Muscle Glycogen Content And.77sebastian valdebenitoNo ratings yet
- Nutrition For The SprinterDocument12 pagesNutrition For The Sprinterhrvoje09No ratings yet
- Nutrition tips for sprinters to build muscle and powerDocument18 pagesNutrition tips for sprinters to build muscle and powerscribdeseNo ratings yet
- The Impact of Metabolic Stress On Hormonal Responses and Muscular AdaptationsDocument10 pagesThe Impact of Metabolic Stress On Hormonal Responses and Muscular AdaptationsJoão NunesNo ratings yet
- Nutrition Guidelines for Marathon, Triathlon, and Cycling AthletesDocument9 pagesNutrition Guidelines for Marathon, Triathlon, and Cycling AthletescakebakeshakeNo ratings yet
- Nutrients: Carbohydrate Availability and Physical Performance: Physiological Overview and Practical RecommendationsDocument10 pagesNutrients: Carbohydrate Availability and Physical Performance: Physiological Overview and Practical RecommendationsX Adolfo Pulido Davila XNo ratings yet
- CipryanDocument10 pagesCipryan197Anzali CahyatiNo ratings yet
- Journal of The International Society of Sports NutritionDocument12 pagesJournal of The International Society of Sports NutritionMahdian RmqaNo ratings yet
- Polarized Training Has Greater Impact On Key Endurance Variables Than Threshold, High Intensity, or High Volume TrainingDocument9 pagesPolarized Training Has Greater Impact On Key Endurance Variables Than Threshold, High Intensity, or High Volume TrainingsergiNo ratings yet
- Levenhagen Et Al. (2001)Document13 pagesLevenhagen Et Al. (2001)testNo ratings yet
- Considerations For Protein Intake in Managing Weight Loss in AthletesDocument9 pagesConsiderations For Protein Intake in Managing Weight Loss in AthletesRoberto CasilloNo ratings yet
- Krista 2009Document9 pagesKrista 2009Vanessa BarrosNo ratings yet
- Can Drop Set Training Enhance Muscle Growth?: Strength and Conditioning Journal December 2017Document12 pagesCan Drop Set Training Enhance Muscle Growth?: Strength and Conditioning Journal December 2017EijunNo ratings yet
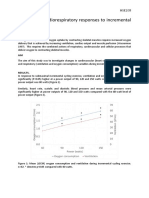
- Practical 4: Cardiorespiratory Responses To Incremental ExerciseDocument4 pagesPractical 4: Cardiorespiratory Responses To Incremental ExercisePATRICK OTIATONo ratings yet
- HIIT vs. Continuous Cardiovascular ExerciseDocument7 pagesHIIT vs. Continuous Cardiovascular ExerciseMarko ŠtambukNo ratings yet
- The Timing of Postexercise Protein Ingestion Is-Is Not ImportantDocument5 pagesThe Timing of Postexercise Protein Ingestion Is-Is Not ImportantEdoardo BianchiNo ratings yet
- Improving Endurance Performance WithDocument11 pagesImproving Endurance Performance WithmoggilavannanNo ratings yet
- Maximal Lipid Oxidation During Exercise: A Target For Individualizing Endurance Training in Obesity and Diabetes?Document2 pagesMaximal Lipid Oxidation During Exercise: A Target For Individualizing Endurance Training in Obesity and Diabetes?khaled safiNo ratings yet
- Low-Carbohydrate Training Increases Protein Requirements of Endurance AthletesDocument8 pagesLow-Carbohydrate Training Increases Protein Requirements of Endurance AthletesAlejandro Estrada RiosNo ratings yet
- Fatmax A New Concept To Optimize Fat Oxidation DurDocument6 pagesFatmax A New Concept To Optimize Fat Oxidation Durjesus.clemente.90No ratings yet
- 1 Sathish EtalDocument5 pages1 Sathish EtaleditorijmrhsNo ratings yet
- FFHP 6Document29 pagesFFHP 6Danny GoldstoneNo ratings yet
- El Efecto de La Suplementación Con Aceite de Pescado en Las Adaptaciones Inducidas Por El Entrenamiento de Resistencia 2023Document18 pagesEl Efecto de La Suplementación Con Aceite de Pescado en Las Adaptaciones Inducidas Por El Entrenamiento de Resistencia 2023RosaNo ratings yet
- Nutritional Interventions To Promote PosDocument191 pagesNutritional Interventions To Promote PosFelipe FerreiraNo ratings yet
- Effect of Suppl Timinsg&Resistance Excercise of Skeletal Muscle HypertrophyDocument9 pagesEffect of Suppl Timinsg&Resistance Excercise of Skeletal Muscle HypertrophyIñakiPosadasNo ratings yet
- Effect of Low-And High-Glycemic-lndex Meals On Metabolism and Performance During High-Intensity, Intermittent ExerciseDocument11 pagesEffect of Low-And High-Glycemic-lndex Meals On Metabolism and Performance During High-Intensity, Intermittent ExerciseChris HaNo ratings yet
- Strength Training and Weight Loss: Treinamento de Força e EmagrecimentoDocument4 pagesStrength Training and Weight Loss: Treinamento de Força e EmagrecimentoDevasyaNo ratings yet
- Nutrition and Hydration Issues For Combat Sport Athletes (Hoffman J.R.) (2011)Document9 pagesNutrition and Hydration Issues For Combat Sport Athletes (Hoffman J.R.) (2011)AnaNo ratings yet
- NutrientsDocument16 pagesNutrientsMarija NikolicNo ratings yet
- Two Weeks of High-Intensity Aerobic Interval Training Increases The Capacity For Fat Oxidation During Exercise in Women. Journal of ADocument9 pagesTwo Weeks of High-Intensity Aerobic Interval Training Increases The Capacity For Fat Oxidation During Exercise in Women. Journal of AFrancisco EspinozaNo ratings yet
- Ijms 24 05743Document14 pagesIjms 24 05743Antonio Luiz PereiraNo ratings yet
- Nutritional Aspects in Ultra-Endurance Exercise: Edith M. PetersDocument8 pagesNutritional Aspects in Ultra-Endurance Exercise: Edith M. PetersTBC TBCNo ratings yet
- Train Low Race HighDocument11 pagesTrain Low Race HighVasco PintadoNo ratings yet
- Fact #1: Wellness & Fitness-Body OptimizationDocument13 pagesFact #1: Wellness & Fitness-Body OptimizationCheeken CharliNo ratings yet
- Nutrients 13 02896Document16 pagesNutrients 13 02896Marija NikolicNo ratings yet
- Gonzalez Et Al. Amino AcidsDocument11 pagesGonzalez Et Al. Amino AcidsFrancisco Jose Albert GarciaNo ratings yet
- Physiological Adaptations To Low-Volume, High-Intensity Interval Training in Health and DiseaseDocument8 pagesPhysiological Adaptations To Low-Volume, High-Intensity Interval Training in Health and DiseasetomNo ratings yet
- Increased Fat Availability and Exercise PDFDocument7 pagesIncreased Fat Availability and Exercise PDFoskr1810No ratings yet
- Andrews Sedlock 2003 CHO Loading WomenDocument7 pagesAndrews Sedlock 2003 CHO Loading WomenVeronica ToddNo ratings yet
- Ijmrhs Vol 3 Issue 4Document294 pagesIjmrhs Vol 3 Issue 4editorijmrhsNo ratings yet
- Cafeina y Ejercicio 2Document10 pagesCafeina y Ejercicio 2Rodrigo Durán San MartínNo ratings yet
- Hepatokines - A Novel Group of Exercise FactorsDocument14 pagesHepatokines - A Novel Group of Exercise FactorsGabriela SolanoNo ratings yet
- Obesity - Short Scientific Findings to Ameliorate the Body WeightFrom EverandObesity - Short Scientific Findings to Ameliorate the Body WeightNo ratings yet
- Troublesome Verb Pairs 8Document3 pagesTroublesome Verb Pairs 8OscarTadlockNo ratings yet
- Dragon Head PrintableDocument1 pageDragon Head PrintableOscarTadlockNo ratings yet
- Ch. 5 Travel and Transportations Study SheetDocument2 pagesCh. 5 Travel and Transportations Study SheetOscarTadlockNo ratings yet
- All The Things You AreDocument1 pageAll The Things You AreOscarTadlockNo ratings yet
- Civil War Music and MotivationDocument16 pagesCivil War Music and MotivationOscarTadlockNo ratings yet
- Mahalia Jackson RWNBDocument11 pagesMahalia Jackson RWNBOscarTadlockNo ratings yet
- Civil War Music and MotivationDocument16 pagesCivil War Music and MotivationOscarTadlockNo ratings yet
- Stomach MeridianDocument1 pageStomach MeridianOscarTadlockNo ratings yet
- Power Forearms PDFDocument36 pagesPower Forearms PDFMike100% (2)
- 2 52 李慕華-課堂報告 痛經針灸報告Document2 pages2 52 李慕華-課堂報告 痛經針灸報告OscarTadlockNo ratings yet
- Glossary of Traditional Chinese Medicine TermsDocument7 pagesGlossary of Traditional Chinese Medicine TermsOscarTadlockNo ratings yet
- 痛經針灸報告Document1 page痛經針灸報告OscarTadlockNo ratings yet
- 莊子 essayDocument3 pages莊子 essayOscarTadlockNo ratings yet
- Lost World VocDocument3 pagesLost World VocOscarTadlockNo ratings yet
- 醫院服務人員英語對話手冊Document65 pages醫院服務人員英語對話手冊OscarTadlockNo ratings yet
- CM Considerations 4.10.14 FINALDocument21 pagesCM Considerations 4.10.14 FINALPaul Michael BalesNo ratings yet
- Huang Lao EssayDocument3 pagesHuang Lao EssayOscarTadlockNo ratings yet
- Taiwan HealthcareDocument21 pagesTaiwan HealthcareOscarTadlockNo ratings yet
- What Is A Normal Pulse Rate - Heart Matters Magazine BHFDocument1 pageWhat Is A Normal Pulse Rate - Heart Matters Magazine BHFabdullahNo ratings yet
- Cai Songfang Wujishi Breathing Exercises Medicine Health Publishing Co. 1994Document119 pagesCai Songfang Wujishi Breathing Exercises Medicine Health Publishing Co. 1994Amlen Singha100% (2)
- Design and Implementation of Medicine Reminder Box and Health CheckerDocument6 pagesDesign and Implementation of Medicine Reminder Box and Health CheckerIJRASETPublications100% (1)
- PE12 Q2 Module5Document18 pagesPE12 Q2 Module5Angel AquinoNo ratings yet
- 5257362Document1 page5257362Khan Hadiur RahmanNo ratings yet
- Advanced Life Support EMS ScenariosDocument147 pagesAdvanced Life Support EMS Scenariosfidelurtecho4881No ratings yet
- ASIM M9Homeo Homeostasis 5E TEACHER NOTES, Revised 10/2015Document6 pagesASIM M9Homeo Homeostasis 5E TEACHER NOTES, Revised 10/2015Joshua WoodsNo ratings yet
- Physical Activity Recommendations and Effects on HealthDocument12 pagesPhysical Activity Recommendations and Effects on HealthAroundGuyJNo ratings yet
- DLL Pe LasDocument8 pagesDLL Pe LasArgel Quilab LarongcoNo ratings yet
- 7 10 17Document2 pages7 10 17Josal Mariano JacintoNo ratings yet
- Module 1 Lesson 1 SPEAR 1 Fitness Assessment TestDocument15 pagesModule 1 Lesson 1 SPEAR 1 Fitness Assessment TestKlent CabrasNo ratings yet
- Paco Seirul-Lo Training The Aerobic Capacity of Distance Runners .5Document4 pagesPaco Seirul-Lo Training The Aerobic Capacity of Distance Runners .5Francisco Júlio de Sousa100% (1)
- Neonatal AsphyxiaDocument35 pagesNeonatal AsphyxiaAsha jilu67% (3)
- Survival Stress in Law EnforcementDocument25 pagesSurvival Stress in Law EnforcementFelipe PalmaNo ratings yet
- VHA-En-2016-Does Cat Attachment Have An Effect On Human Health - A Comparison Between Owners and VolunteersDocument12 pagesVHA-En-2016-Does Cat Attachment Have An Effect On Human Health - A Comparison Between Owners and VolunteersbrbgaralNo ratings yet
- Livestrong Elliptical Owner's Manual - English - French - SpanishDocument128 pagesLivestrong Elliptical Owner's Manual - English - French - SpanishPaul KennyNo ratings yet
- Personality Psychology 6th Edition Larsen Test BankDocument21 pagesPersonality Psychology 6th Edition Larsen Test Bankthoabangt69100% (31)
- Effectiveness of Bolus Administration of Continuous Intravenous Lidocaine On Pain Intensity After Mastectomy SurgeryDocument6 pagesEffectiveness of Bolus Administration of Continuous Intravenous Lidocaine On Pain Intensity After Mastectomy SurgeryInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- The Ergonomics of Sheep Shearing: Reducing Injuries for Agricultural WorkersDocument8 pagesThe Ergonomics of Sheep Shearing: Reducing Injuries for Agricultural WorkersPera DaviesNo ratings yet
- Windkessel Model 1Document13 pagesWindkessel Model 1Emmanuel PrahNo ratings yet
- Unipulse 400Document6 pagesUnipulse 400Eliel VelasquezNo ratings yet
- Neonatal Life Support UK Ressusstation Council G2015 - NLSDocument1 pageNeonatal Life Support UK Ressusstation Council G2015 - NLSBharat Kumar SharmaNo ratings yet
- Reviewer FITTDocument14 pagesReviewer FITTHilton D. CalawenNo ratings yet
- Horizon BSC 100 & 200 USER MANUAL PDFDocument31 pagesHorizon BSC 100 & 200 USER MANUAL PDFRayNo ratings yet
- Week 10 Worksheet AnswersDocument13 pagesWeek 10 Worksheet AnswersAb Staholic BoiiNo ratings yet
- Cycling Test ProtocolDocument7 pagesCycling Test ProtocolJonathan Bradley TepplerNo ratings yet
- Physiological and Psychological Fatigue in Extreme Conditions - Overtraining and Elite AthletesDocument9 pagesPhysiological and Psychological Fatigue in Extreme Conditions - Overtraining and Elite AthletesSalomé Horta LeytonNo ratings yet
- How to Read an ECG Step-by-Step GuideDocument15 pagesHow to Read an ECG Step-by-Step GuideCFTCNo ratings yet
- 15 Laws of TrainingDocument51 pages15 Laws of TrainingMichaelKahnertNo ratings yet
- Definition of Health Related Fitness: MapehDocument3 pagesDefinition of Health Related Fitness: MapehharijarenallNo ratings yet