You might also like
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Fds-Ofite Edta 0,1MDocument7 pagesFds-Ofite Edta 0,1MVeinte Años Sin VosNo ratings yet
- Delusion in SocietyDocument2 pagesDelusion in SocietyGasimovskyNo ratings yet
- Review On AlgebraDocument29 pagesReview On AlgebraGraziela GutierrezNo ratings yet
- What's New in CAESAR II: Piping and Equipment CodesDocument1 pageWhat's New in CAESAR II: Piping and Equipment CodeslnacerNo ratings yet
- IEC TC 56 Dependability PDFDocument8 pagesIEC TC 56 Dependability PDFsaospieNo ratings yet
- 2016 Closing The Gap ReportDocument64 pages2016 Closing The Gap ReportAllan ClarkeNo ratings yet
- Cisco BGP ASPATH FilterDocument115 pagesCisco BGP ASPATH FilterHalison SantosNo ratings yet
- Escaner Electromagnético de Faja Transportadora-Steel SPECTDocument85 pagesEscaner Electromagnético de Faja Transportadora-Steel SPECTEdwin Alfredo Eche QuirozNo ratings yet
- The Influence of Irish Monks On Merovingian Diocesan Organization-Robbins BittermannDocument15 pagesThe Influence of Irish Monks On Merovingian Diocesan Organization-Robbins BittermanngeorgiescuNo ratings yet
- WarringFleets Complete PDFDocument26 pagesWarringFleets Complete PDFlingshu8100% (1)
- Thesis PaperDocument53 pagesThesis PaperAnonymous AOOrehGZAS100% (1)
- IMCI Chart BookletDocument43 pagesIMCI Chart Bookletmysticeyes_17100% (1)
- Health Post - Exploring The Intersection of Work and Well-Being - A Guide To Occupational Health PsychologyDocument3 pagesHealth Post - Exploring The Intersection of Work and Well-Being - A Guide To Occupational Health PsychologyihealthmailboxNo ratings yet
- Hidrl1 PDFDocument7 pagesHidrl1 PDFRajesh Kumar100% (1)
- Webinar Gizi - Patho StuntingDocument16 pagesWebinar Gizi - Patho StuntingMiftahul HikmahNo ratings yet
- Toeic: Check Your English Vocabulary ForDocument41 pagesToeic: Check Your English Vocabulary ForEva Ibáñez RamosNo ratings yet
- RFID Seminar AbstractDocument2 pagesRFID Seminar Abstractanushabhagawath80% (5)
- ReadmeDocument2 pagesReadmechethan100% (1)
- ChatGpt PDFDocument19 pagesChatGpt PDFsanx2014100% (1)
- Assistant Cook Learner Manual EnglishDocument152 pagesAssistant Cook Learner Manual EnglishSang Putu Arsana67% (3)
- Lecture 2 Effects of Operating Conditions in VCCDocument9 pagesLecture 2 Effects of Operating Conditions in VCCDeniell Joyce MarquezNo ratings yet
- Friction: Ultiple Hoice UestionsDocument5 pagesFriction: Ultiple Hoice Uestionspk2varmaNo ratings yet
- Final Project Strategic ManagementDocument2 pagesFinal Project Strategic ManagementMahrukh RasheedNo ratings yet
- Baseline Scheduling Basics - Part-1Document48 pagesBaseline Scheduling Basics - Part-1Perwaiz100% (1)
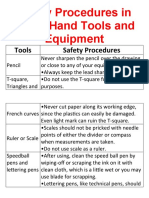
- Safety Procedures in Using Hand Tools and EquipmentDocument12 pagesSafety Procedures in Using Hand Tools and EquipmentJan IcejimenezNo ratings yet
- Physics Blue Print 1 Class XI Half Yearly 23Document1 pagePhysics Blue Print 1 Class XI Half Yearly 23Nilima Aparajita SahuNo ratings yet
- Recitation Math 001 - Term 221 (26166)Document36 pagesRecitation Math 001 - Term 221 (26166)Ma NaNo ratings yet
- Man As God Created Him, ThemDocument3 pagesMan As God Created Him, ThemBOEN YATORNo ratings yet
- Micro EvolutionDocument9 pagesMicro EvolutionBryan TanNo ratings yet
- Iguana Joe's Lawsuit - September 11, 2014Document14 pagesIguana Joe's Lawsuit - September 11, 2014cindy_georgeNo ratings yet