You might also like
- FraudDocument77 pagesFraudTan Siew Li100% (2)
- Chess Handbook For Parents and Coaches: Ronn MunstermanDocument29 pagesChess Handbook For Parents and Coaches: Ronn MunstermanZull Ise HishamNo ratings yet
- Grade 2 - PAN-ASSESSMENT-TOOLDocument5 pagesGrade 2 - PAN-ASSESSMENT-TOOLMaestro Varix100% (4)
- Adult Neurogenesis in the Hippocampus: Health, Psychopathology, and Brain DiseaseFrom EverandAdult Neurogenesis in the Hippocampus: Health, Psychopathology, and Brain DiseaseJuan J. CanalesNo ratings yet
- Felomino Urbano vs. IAC, G.R. No. 72964, January 7, 1988 ( (157 SCRA 7)Document1 pageFelomino Urbano vs. IAC, G.R. No. 72964, January 7, 1988 ( (157 SCRA 7)Dwight LoNo ratings yet
- SCIENCE 5 PPT Q3 W6 - Parts of An Electric CircuitDocument24 pagesSCIENCE 5 PPT Q3 W6 - Parts of An Electric CircuitDexter Sagarino100% (1)
- Office of The Court Administrator v. de GuzmanDocument7 pagesOffice of The Court Administrator v. de GuzmanJon Joshua FalconeNo ratings yet
- Url-2003 Parvovirus en Neuronas de GatoDocument5 pagesUrl-2003 Parvovirus en Neuronas de GatocristianNo ratings yet
- Cellular and Subcellular Specification of Na, K-ATPase A and B Isoforms in The Postnatal Development of Mouse RetinaDocument12 pagesCellular and Subcellular Specification of Na, K-ATPase A and B Isoforms in The Postnatal Development of Mouse RetinaJose GarciaNo ratings yet
- Apoptosis in Natural Rabies Virus Infection in DogsDocument5 pagesApoptosis in Natural Rabies Virus Infection in DogspapiipiiNo ratings yet
- 6142 FullDocument12 pages6142 FullAmol MundeNo ratings yet
- FN_Art_10047-1Document11 pagesFN_Art_10047-1Carmen NeacsuNo ratings yet
- Light-Induced Photon Emission by Mammalian CellsDocument5 pagesLight-Induced Photon Emission by Mammalian CellsAntonis TzambazakisNo ratings yet
- Stemcells 2006-0409Document9 pagesStemcells 2006-0409dupuytrenNo ratings yet
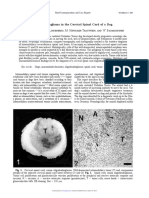
- Oligodendroglioma in The Cervical Spinal Cord of A Dog: T. M, A. M - L, M. H - T, W. BDocument3 pagesOligodendroglioma in The Cervical Spinal Cord of A Dog: T. M, A. M - L, M. H - T, W. BSamir BazanNo ratings yet
- 2010 - The Zebrafish As A Model Organism For The Study of Apoptosis PDFDocument19 pages2010 - The Zebrafish As A Model Organism For The Study of Apoptosis PDFSebastián PiernagordaNo ratings yet
- TMP 5 CCDocument13 pagesTMP 5 CCFrontiersNo ratings yet
- Basofilos 1Document8 pagesBasofilos 1Natalia Cano GNo ratings yet
- FinalCorona NanniDocument3 pagesFinalCorona Nannimnanni1No ratings yet
- Doença de Batten 2004Document9 pagesDoença de Batten 2004Ana Paula BoscatoNo ratings yet
- Apoptosis in The NcsDocument8 pagesApoptosis in The NcsMiguel Angel Villarreal YbazetaNo ratings yet
- Transcription of The SCL TAL1 Interrupting Locus Stil - 2014 - Journal of BioDocument7 pagesTranscription of The SCL TAL1 Interrupting Locus Stil - 2014 - Journal of BioMary GarridoNo ratings yet
- Immunohistochemical Characterization of A Renal Nephroblastoma in A Trp53-Mutant and Prolyl Isomerase 1-Deficient MouseDocument5 pagesImmunohistochemical Characterization of A Renal Nephroblastoma in A Trp53-Mutant and Prolyl Isomerase 1-Deficient Mouselily rogaNo ratings yet
- Primary T-Cell Lymphoma of The Central 2001Document4 pagesPrimary T-Cell Lymphoma of The Central 2001emmanouelaapostolopoulouNo ratings yet
- Chat 5HT Brain Stem Mouse AnatomyDocument35 pagesChat 5HT Brain Stem Mouse AnatomyRam KNo ratings yet
- tmp337B TMPDocument29 pagestmp337B TMPFrontiersNo ratings yet
- Neuroprotective Dopamine - FullDocument7 pagesNeuroprotective Dopamine - FullmorexyzNo ratings yet
- Canine DistemperDocument16 pagesCanine DistemperFelipe GonzalezNo ratings yet
- Articulo Neuro 1Document12 pagesArticulo Neuro 1Victor CuellarNo ratings yet
- Ref 5Document11 pagesRef 5Tiago BaraNo ratings yet
- Mitochondrial Respiration and ATP Production Are Significantly Impaired in Striatal Cells Expressing Mutant HuntingtinDocument11 pagesMitochondrial Respiration and ATP Production Are Significantly Impaired in Striatal Cells Expressing Mutant HuntingtinNICOLE HSIAO-SANCHEZNo ratings yet
- Pi Is 0021925819770471Document6 pagesPi Is 0021925819770471ivette ramirezNo ratings yet
- Pallatto Et Al-2018-Veterinary Clinical PathologyDocument4 pagesPallatto Et Al-2018-Veterinary Clinical Pathologygranulomatous pneumoniaNo ratings yet
- Https:journals Sagepub Com:doi:pdf:10 1354:vp 37-6-650Document3 pagesHttps:journals Sagepub Com:doi:pdf:10 1354:vp 37-6-650Lucas XavierNo ratings yet
- Animal Model of EADocument13 pagesAnimal Model of EAzigurat00No ratings yet
- Balasubramanian 2013 Cloning and Molecular Analysis of The Aspartic Protease Sc-ASP110 GeneDocument8 pagesBalasubramanian 2013 Cloning and Molecular Analysis of The Aspartic Protease Sc-ASP110 GeneIliana Zazueta OjedaNo ratings yet
- Villeda, 2011 PDFDocument7 pagesVilleda, 2011 PDFDaniel TorresNo ratings yet
- tmp1CA5 TMPDocument10 pagestmp1CA5 TMPFrontiersNo ratings yet
- Del ValleDocument4 pagesDel ValleEva Del ValleNo ratings yet
- Somatotroph Pituitary Tumors in Budgerigars (Melopsittacus Undulatus)Document5 pagesSomatotroph Pituitary Tumors in Budgerigars (Melopsittacus Undulatus)Firoz RezaNo ratings yet
- Differential Transcripts TheDocument5 pagesDifferential Transcripts TheGuhan KANo ratings yet
- Obe. Uz) .: RAT (P CtmraoraqDocument1 pageObe. Uz) .: RAT (P CtmraoraqKundanNo ratings yet
- Hepatology - 2008 - Yovchev - Identification of Adult Hepatic Progenitor Cells Capable of Repopulating Injured Rat Liver6Document12 pagesHepatology - 2008 - Yovchev - Identification of Adult Hepatic Progenitor Cells Capable of Repopulating Injured Rat Liver6relbelkasyNo ratings yet
- Overexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 CellsDocument7 pagesOverexpression of Hematopoietically Expressed Homeoprotein Induces Nonapoptotic Cell Death in Mouse Prechondrogenic ATDC5 CellsCamilo CabreraNo ratings yet
- UCHL1 TesDocument7 pagesUCHL1 TesClint KashNo ratings yet
- Jin Wan, Hua Zheng, Hong-Lei Xiao, Zhen-Jue She and Guo-Min Zhou - Sonic Hedgehog Promotes Stem-Cell Potential of Muller Glia in The Mammalian RetinaDocument8 pagesJin Wan, Hua Zheng, Hong-Lei Xiao, Zhen-Jue She and Guo-Min Zhou - Sonic Hedgehog Promotes Stem-Cell Potential of Muller Glia in The Mammalian RetinaHutsDMNo ratings yet
- 1993 Chun Han Chung Waessle JCompNeurolDocument12 pages1993 Chun Han Chung Waessle JCompNeuroljmcanadaNo ratings yet
- Astrocytes and Epilepsy: Nihal C. de Lanerolle, Tih-Shih Lee, and Dennis D. SpencerDocument15 pagesAstrocytes and Epilepsy: Nihal C. de Lanerolle, Tih-Shih Lee, and Dennis D. SpencerErsin KarataşNo ratings yet
- Biochimie Volume 81 Issue 6-Supp-S1 1999 (Doi 10.1016/s0300-9084 (99) 80112-6) - The Sir Hans Krebs LectureDocument390 pagesBiochimie Volume 81 Issue 6-Supp-S1 1999 (Doi 10.1016/s0300-9084 (99) 80112-6) - The Sir Hans Krebs LectureAllcrissNo ratings yet
- Yi_et_al-Glia 2017Document19 pagesYi_et_al-Glia 2017Daniela HenríquezNo ratings yet
- Maduracion de La AmigadalaDocument15 pagesMaduracion de La AmigadalaAracelis Calzadilla NúñezNo ratings yet
- Senile Plaque Neurites Alzheimer Disease Accumulate Amyloid Precursor ProteinDocument5 pagesSenile Plaque Neurites Alzheimer Disease Accumulate Amyloid Precursor Proteinnegru paulNo ratings yet
- Aaas 2011 MeetingDocument1 pageAaas 2011 Meetingapi-234075032No ratings yet
- EXOSC8Document6 pagesEXOSC8vuhaipham1407No ratings yet
- Li 07 StemcellsDocument15 pagesLi 07 Stemcellsapi-3821304No ratings yet
- Current Status and Challenges of Stem Cell Treatment For Alzheimer's DiseaseDocument19 pagesCurrent Status and Challenges of Stem Cell Treatment For Alzheimer's DiseaseHelical Nail FemurNo ratings yet
- Evidence for Human Lung Stem Cells: Identification and CharacterizationDocument11 pagesEvidence for Human Lung Stem Cells: Identification and CharacterizationChairunisa AnggrainiNo ratings yet
- Glial Cells Shape Brain Physiology and PathologyDocument12 pagesGlial Cells Shape Brain Physiology and PathologyBernardo LeñeroNo ratings yet
- Neun, A Neuronal Specific Nuclear Protein in VertebratesDocument11 pagesNeun, A Neuronal Specific Nuclear Protein in VertebratesAditya KhemkaNo ratings yet
- Elevated Gene Expression Levels Distinguish Human Distinguish Human From Non HumansCLAVEEEEEEEEEDocument6 pagesElevated Gene Expression Levels Distinguish Human Distinguish Human From Non HumansCLAVEEEEEEEEELina María SalgueroNo ratings yet
- Upregulated Acetylcholine Synthesis During Early Differentiation in The Embryonic Stem Cell Line Cgr8Document5 pagesUpregulated Acetylcholine Synthesis During Early Differentiation in The Embryonic Stem Cell Line Cgr8JasonNo ratings yet
- NHE6 PaperDocument28 pagesNHE6 PaperJon DrdNo ratings yet
- Restraint Stress Affects Hippocampal CelDocument4 pagesRestraint Stress Affects Hippocampal CelJean Pierre Chastre LuzaNo ratings yet
- Accumulation of Abnormally Processed Tau Protein in Neuronal Cells As A Biomarker For DementiaDocument20 pagesAccumulation of Abnormally Processed Tau Protein in Neuronal Cells As A Biomarker For Dementiakj185No ratings yet
- Citosol Triggers Apoptosis in Leukemia CellsDocument1 pageCitosol Triggers Apoptosis in Leukemia CellsAlondra MaldonadoNo ratings yet
- Animal Models and Hypoxia: Proceedings of an International Symposium on Animal Models and Hypoxia, Held at Wiesbaden, Federal Republic of Germany, 19 November 1979From EverandAnimal Models and Hypoxia: Proceedings of an International Symposium on Animal Models and Hypoxia, Held at Wiesbaden, Federal Republic of Germany, 19 November 1979V. StefanovichNo ratings yet
- Lentivirus Production ProtocolDocument11 pagesLentivirus Production ProtocoldupuytrenNo ratings yet
- Eview: Alternative Polyadenylation: A Twist On mRNA 3 End FormationDocument9 pagesEview: Alternative Polyadenylation: A Twist On mRNA 3 End FormationdupuytrenNo ratings yet
- Adenovirus Production ProtocolDocument13 pagesAdenovirus Production ProtocoldupuytrenNo ratings yet
- Genome-Wide Alternative Polyadenylation in Animals: Insights From High-Throughput TechnologiesDocument10 pagesGenome-Wide Alternative Polyadenylation in Animals: Insights From High-Throughput TechnologiesdupuytrenNo ratings yet
- Fluorescence and Fluorescence ApplicationsDocument13 pagesFluorescence and Fluorescence ApplicationsPaulraj Mosae SelvakumarNo ratings yet
- Simplicon Rna Reprogramming - PrinterviewDocument5 pagesSimplicon Rna Reprogramming - PrinterviewdupuytrenNo ratings yet
- Non-Genetic Inheritance Via The Male Germline in Mammals: ReviewDocument7 pagesNon-Genetic Inheritance Via The Male Germline in Mammals: ReviewdupuytrenNo ratings yet
- Heat Inactivation ProtocolDocument1 pageHeat Inactivation ProtocoldupuytrenNo ratings yet
- Direct Comparison of Dna Versus Mrna On The Efficiency of Induced Pluripotent Stem Cells Generation From Human Primary FibroblastsDocument7 pagesDirect Comparison of Dna Versus Mrna On The Efficiency of Induced Pluripotent Stem Cells Generation From Human Primary FibroblastsdupuytrenNo ratings yet
- Microtubules in SynapseDocument6 pagesMicrotubules in SynapsedupuytrenNo ratings yet
- Dynamics of Epigenetic Phenomena: Intergenerational and Intragenerational Phenotype Washout'Document8 pagesDynamics of Epigenetic Phenomena: Intergenerational and Intragenerational Phenotype Washout'dupuytrenNo ratings yet
- An Integrative Framework For Understanding The Mechanisms and Multigenerational Consequences of Transgenerational PlasticityDocument22 pagesAn Integrative Framework For Understanding The Mechanisms and Multigenerational Consequences of Transgenerational PlasticitydupuytrenNo ratings yet
- Adhesion Signalling Complexes: PrimerDocument5 pagesAdhesion Signalling Complexes: PrimerdupuytrenNo ratings yet
- 5550 FullDocument8 pages5550 FulldupuytrenNo ratings yet
- Uncovering Cell Biology in The Third DimensionDocument5 pagesUncovering Cell Biology in The Third DimensiondupuytrenNo ratings yet
- Stemcells 2006-0409Document9 pagesStemcells 2006-0409dupuytrenNo ratings yet
- Commercialization of Organoids: ForumDocument4 pagesCommercialization of Organoids: ForumdupuytrenNo ratings yet
- Focal Adhesions in Osteoneogenesis: Special Issue Paper 1441Document13 pagesFocal Adhesions in Osteoneogenesis: Special Issue Paper 1441dupuytrenNo ratings yet
- Echinocandins ReviewDocument11 pagesEchinocandins ReviewdupuytrenNo ratings yet
- Perspectives: Applying CRISPR-Cas9 Tools To Identify and Characterize Transcriptional EnhancersDocument8 pagesPerspectives: Applying CRISPR-Cas9 Tools To Identify and Characterize Transcriptional EnhancersdupuytrenNo ratings yet
- Studying Hallmarks of Cancer - AbcamDocument11 pagesStudying Hallmarks of Cancer - AbcamdupuytrenNo ratings yet
- Pioneer Transcription Factors in Cell Fate ControlDocument5 pagesPioneer Transcription Factors in Cell Fate ControldupuytrenNo ratings yet
- Cellular Viability - Calcein / Propidium Iodide: We've Performed This Assay On The Following Cell TypesDocument1 pageCellular Viability - Calcein / Propidium Iodide: We've Performed This Assay On The Following Cell TypesdupuytrenNo ratings yet
- New Drugs 2017Document2 pagesNew Drugs 2017dupuytrenNo ratings yet
- Plan the retrosynthesis of target moleculesDocument3 pagesPlan the retrosynthesis of target moleculesJaya KumarNo ratings yet
- 8927 FullDocument5 pages8927 FulldupuytrenNo ratings yet
- Treatment of Antineutrophil Cytoplasmic Antibodyassociated VasculitisDocument8 pagesTreatment of Antineutrophil Cytoplasmic Antibodyassociated VasculitisdupuytrenNo ratings yet
- Preliminary MatterDocument26 pagesPreliminary MatterdupuytrenNo ratings yet
- Neuroview: How To Be A Graduate AdviseeDocument3 pagesNeuroview: How To Be A Graduate AdviseedupuytrenNo ratings yet
- Cold Spring Harb Protoc-2011-Matheu-pdb - Prot5565Document6 pagesCold Spring Harb Protoc-2011-Matheu-pdb - Prot5565dupuytrenNo ratings yet
- Unofficial Transcript - Printer FriendlyDocument4 pagesUnofficial Transcript - Printer Friendlyapi-251794642No ratings yet
- Pure TheoriesDocument5 pagesPure Theorieschristine angla100% (1)
- TOPIC - 1 - Intro To Tourism PDFDocument16 pagesTOPIC - 1 - Intro To Tourism PDFdevvy anneNo ratings yet
- Konsep SRA (Sekolah Ramah Anak) Dalam Membentuk Budaya Islami Di Sekolah DasarDocument10 pagesKonsep SRA (Sekolah Ramah Anak) Dalam Membentuk Budaya Islami Di Sekolah Dasarsupriyono hasanNo ratings yet
- Chemistry Sample Paper 2021-22Document16 pagesChemistry Sample Paper 2021-22sarthak MongaNo ratings yet
- Career Guidance Program Modules MonitoringDocument7 pagesCareer Guidance Program Modules MonitoringJevin GonzalesNo ratings yet
- Preterm Labour: Muhammad Hanif Final Year MBBSDocument32 pagesPreterm Labour: Muhammad Hanif Final Year MBBSArslan HassanNo ratings yet
- Sadhu or ShaitaanDocument3 pagesSadhu or ShaitaanVipul RathodNo ratings yet
- Helical Antennas: Circularly Polarized, High Gain and Simple to FabricateDocument17 pagesHelical Antennas: Circularly Polarized, High Gain and Simple to FabricatePrasanth KumarNo ratings yet
- First Time Login Guidelines in CRMDocument23 pagesFirst Time Login Guidelines in CRMSumeet KotakNo ratings yet
- Ettercap PDFDocument13 pagesEttercap PDFwyxchari3No ratings yet
- Term2 WS7 Revision2 PDFDocument5 pagesTerm2 WS7 Revision2 PDFrekhaNo ratings yet
- Cloud Computing Basics and Service ModelsDocument29 pagesCloud Computing Basics and Service ModelsBhupendra singh TomarNo ratings yet
- GPAODocument2 pagesGPAOZakariaChardoudiNo ratings yet
- Electric Vehicles PresentationDocument10 pagesElectric Vehicles PresentationKhagesh JoshNo ratings yet
- What Is Innovation and Its Characteristics of InnovationDocument4 pagesWhat Is Innovation and Its Characteristics of InnovationMohd TauqeerNo ratings yet
- MagnitismDocument3 pagesMagnitismapi-289032603No ratings yet
- MES - Project Orientation For Night Study - V4Document41 pagesMES - Project Orientation For Night Study - V4Andi YusmarNo ratings yet
- Inver Powderpaint SpecirficationsDocument2 pagesInver Powderpaint SpecirficationsArun PadmanabhanNo ratings yet
- God Love Value CoreDocument11 pagesGod Love Value CoreligayaNo ratings yet
- A Study On Waves of FeminismDocument3 pagesA Study On Waves of FeminismLusekero MwangondeNo ratings yet
- Derivatives 17 Session1to4Document209 pagesDerivatives 17 Session1to4anon_297958811No ratings yet
- Transportation ProblemDocument12 pagesTransportation ProblemSourav SahaNo ratings yet
- Zeng 2020Document11 pagesZeng 2020Inácio RibeiroNo ratings yet