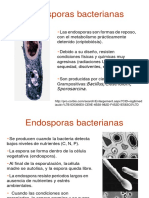
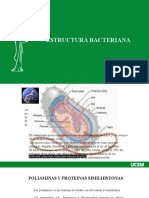
You might also like
- La Endospora Bacteriana y La EsporulaciónDocument14 pagesLa Endospora Bacteriana y La EsporulaciónTerry Huaman Che100% (1)
- La Endospora Bacteriana y La EsporulaciónDocument10 pagesLa Endospora Bacteriana y La EsporulaciónLeo OlivoNo ratings yet
- Endospora Bacteriana y EsporulacionDocument18 pagesEndospora Bacteriana y Esporulacioncaty100% (22)
- Taller 1 MBUM 1001 EsporulaciónDocument7 pagesTaller 1 MBUM 1001 EsporulaciónDuerre WatsonNo ratings yet
- Esporulación BacterianaDocument7 pagesEsporulación BacterianaSol De la PeñaNo ratings yet
- Esporas 20132Document44 pagesEsporas 20132Will ACNo ratings yet
- Bacterias TP 9 Material de EstudioDocument11 pagesBacterias TP 9 Material de Estudionicov04No ratings yet
- Celula Procariota MARTINDocument30 pagesCelula Procariota MARTINYAHAIRA TRUJILLO VENCESNo ratings yet
- Esporas y ExosporasDocument4 pagesEsporas y ExosporasAndrea Jimenez100% (2)
- Clase 2Document9 pagesClase 2Sofia Leiva JaraNo ratings yet
- 15 08 21 NucleoideDocument4 pages15 08 21 NucleoideKatiAriasNo ratings yet
- EsporasDocument45 pagesEsporasenriquejoverNo ratings yet
- Estudio de La Celula Procariota y EucariotaDocument5 pagesEstudio de La Celula Procariota y Eucariotabrydel93% (30)
- Esporas BacterianasDocument38 pagesEsporas BacterianasTavoNo ratings yet
- Clase 2 - La Ciencia de La Microbiología, ContDocument10 pagesClase 2 - La Ciencia de La Microbiología, ContMace AsheNo ratings yet
- Morfología y Estructura Bacteriana-DiapositivaDocument39 pagesMorfología y Estructura Bacteriana-DiapositivaFernanda Vilches67% (3)
- EndosporasDocument5 pagesEndosporasMaria Camila Cabarcas RamosNo ratings yet
- Resumen Celula Procariota TrabajoDocument9 pagesResumen Celula Procariota TrabajoCristian Antoni Sanchez ChoqueNo ratings yet
- Unidad 2 - MicrobiologiaDocument17 pagesUnidad 2 - MicrobiologiaLuna WeckNo ratings yet
- Estructura de Las BacteriasDocument5 pagesEstructura de Las BacteriasDulcesita Jackeline Gomez ZumaetaNo ratings yet
- Tema 3 - Características de Los Microorganismos ProcariotasDocument10 pagesTema 3 - Características de Los Microorganismos ProcariotasInes BadiaNo ratings yet
- Ciclo Explicado de La Esporulación Celular.Document34 pagesCiclo Explicado de La Esporulación Celular.LALUKIS100% (1)
- Espiras BacterianasDocument18 pagesEspiras BacterianasEstefanni SanchezNo ratings yet
- Anatomia Bacteriana.Document6 pagesAnatomia Bacteriana.Rosmely Rojas50% (2)
- Célula ProcariotaDocument15 pagesCélula Procariotaleo apazaNo ratings yet
- Resumen Sordelli (Caps 1,2,3,5 y 6)Document26 pagesResumen Sordelli (Caps 1,2,3,5 y 6)Ricardo AguileraNo ratings yet
- La Celula - LlojaaaaaaaaaaaDocument112 pagesLa Celula - LlojaaaaaaaaaaasoldjairNo ratings yet
- Estructuras CelulareDocument9 pagesEstructuras CelulareAndry MartinezNo ratings yet
- Estructura BacterianaDocument28 pagesEstructura BacterianaBoris MaldonadoNo ratings yet
- Tema 11. La Célula ProcariotaDocument6 pagesTema 11. La Célula ProcariotaCristina Agudelo RamírezNo ratings yet
- Taller Morfología y Estructura BacterianaDocument8 pagesTaller Morfología y Estructura BacterianaMelissa Tamayo GómezNo ratings yet
- Unidad II Características de Las BacteriasDocument67 pagesUnidad II Características de Las BacteriasKuzzy Luzzy TorresNo ratings yet
- Informe #08Document18 pagesInforme #08Antonio Bladimir Talledo TrellesNo ratings yet
- Copia de Tema 6 ORGANIZACION CELULARDocument53 pagesCopia de Tema 6 ORGANIZACION CELULARAMIRA MOHAMED EL HARRAKNo ratings yet
- Bacterias UBA (Todo en Un Documento)Document120 pagesBacterias UBA (Todo en Un Documento)Andres LinaresNo ratings yet
- ESPORAS BACTERIANAS KKKKDocument9 pagesESPORAS BACTERIANAS KKKKNuria Sanchez AndiaNo ratings yet
- As Capsula y Pared BacterianaDocument13 pagesAs Capsula y Pared BacterianaLaura Elizabeth AbadNo ratings yet
- Wuolah Free TEMA 3 8Document32 pagesWuolah Free TEMA 3 8carlos ANo ratings yet
- Capitulo 26Document9 pagesCapitulo 26Monse BeltranNo ratings yet
- Preinforme Células Vegetales e InclusionesDocument9 pagesPreinforme Células Vegetales e InclusionesDavid Vinson100% (2)
- Tema 4 - División ProcariotaDocument4 pagesTema 4 - División Procariotaadrima17No ratings yet
- Biologia Comun Clase 4 CelulasDocument12 pagesBiologia Comun Clase 4 CelulasMarcelaNo ratings yet
- 2i EndosporasBRV PDFDocument48 pages2i EndosporasBRV PDFLeslie TrejoNo ratings yet
- Microbiologia y Parasitologia Guia 1Document5 pagesMicrobiologia y Parasitologia Guia 1saraNo ratings yet
- Sesion 02Document42 pagesSesion 02Melissa ChavezNo ratings yet
- Bacterias y ArqueobacteriasDocument7 pagesBacterias y ArqueobacteriasIsaNo ratings yet
- 02 Celula ProcarioticaDocument32 pages02 Celula ProcarioticaCristhian FuentesNo ratings yet
- Objetivo 3. BacteriasDocument23 pagesObjetivo 3. Bacteriasmusicart30008394100% (3)
- Esporas BacterianasDocument2 pagesEsporas BacterianasÄngïe HënäöNo ratings yet
- Citoplasma BacterianoDocument17 pagesCitoplasma Bacterianol3anngNo ratings yet
- Célula - Medicina 2021Document16 pagesCélula - Medicina 2021Carolina MartinezNo ratings yet
- Célula ProcariotaDocument5 pagesCélula ProcariotacecinoreroNo ratings yet
- Procariota Estructuras Mem, Microcorpatimientos, InclusionesDocument11 pagesProcariota Estructuras Mem, Microcorpatimientos, InclusionesAngela HernandezNo ratings yet
- Anatomia Funcional de Las Celulas ProcariotasDocument20 pagesAnatomia Funcional de Las Celulas ProcariotasNitgma DcNo ratings yet
- Estructura Bacteriana Corpusculos Metacromaticos y EsporasDocument22 pagesEstructura Bacteriana Corpusculos Metacromaticos y EsporasMejia Jhenny YaneNo ratings yet
- Medición Del CrecimientoDocument3 pagesMedición Del CrecimientoMarcoNo ratings yet
- Análisis funcional y microbiológico de derivados lácteos y cárnicosFrom EverandAnálisis funcional y microbiológico de derivados lácteos y cárnicosNo ratings yet
- Biología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularFrom EverandBiología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularNo ratings yet
- Acceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012From EverandAcceso a Universidad para Mayores de 25 años. Biología.: Solucionario Pruebas 2006-2012No ratings yet
- Acceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017From EverandAcceso a Universidad para Mayores de 25 años. Biología 2013-2017.: Solucionario Pruebas 2013-2017No ratings yet
- Negocios Verdes Actividad de ReflexionDocument5 pagesNegocios Verdes Actividad de Reflexionthalia jimenez riveraNo ratings yet
- Clase 1 ExplanacionesDocument14 pagesClase 1 ExplanacionesRicardo JesúsNo ratings yet
- Caracterización de YogurtDocument63 pagesCaracterización de YogurtHellenzitah Guarniz JulcaNo ratings yet
- Clase 2.2 Clasificación de Las Rocas 25 05 22Document37 pagesClase 2.2 Clasificación de Las Rocas 25 05 22Pame JácomeNo ratings yet
- ACLS Guia de Estudio 1 Ed Esp 2018 Ebook-1 UnlockedDocument192 pagesACLS Guia de Estudio 1 Ed Esp 2018 Ebook-1 UnlockedPaulina SuquiNo ratings yet
- TalasemiaDocument2 pagesTalasemiaanon-150354No ratings yet
- Minuta Deteccion H. ArmigeraDocument3 pagesMinuta Deteccion H. ArmigeraRoberto SaldiaNo ratings yet
- Funciones VectorialesDocument18 pagesFunciones VectorialesElvimar YajureNo ratings yet
- Mezcladores y Agitadores en El Proceso Del RefrescoDocument7 pagesMezcladores y Agitadores en El Proceso Del RefrescoGiovanniMartinezNo ratings yet
- Fallas Del Proceso Sancionatorio Ambiental en ColombiaDocument4 pagesFallas Del Proceso Sancionatorio Ambiental en ColombiaYesica Hernandez BotiaNo ratings yet
- Candado Seguridad Gpo - ImexDocument1 pageCandado Seguridad Gpo - ImexPsIndustrial Del Noreste SA de CVNo ratings yet
- UNI-Examen Fisica 2008 I - SolucionarioDocument12 pagesUNI-Examen Fisica 2008 I - Solucionarioprofe.alex100% (2)
- Eva. Acumulativa-8º-Cuarto PeriodoDocument5 pagesEva. Acumulativa-8º-Cuarto Periodojose fernandoNo ratings yet
- Enfoque PsicoanalíticoDocument4 pagesEnfoque PsicoanalíticochiiNo ratings yet
- Cuarto. Guía 5. Unidad 2 Análisis de Textos Narrativos PDFDocument3 pagesCuarto. Guía 5. Unidad 2 Análisis de Textos Narrativos PDFAnge EspinozaNo ratings yet
- El Planeta Marte PDFDocument15 pagesEl Planeta Marte PDFmanuel rosilesNo ratings yet
- El Gato Medicina Clínica y Manejo Parte 3Document50 pagesEl Gato Medicina Clínica y Manejo Parte 3Raul Hernandez SotoNo ratings yet
- Solucionario Dominios de Fisica 10 - EduvisiónDocument286 pagesSolucionario Dominios de Fisica 10 - EduvisiónRafael Eduardo Cordoba80% (15)
- Taller 2 Tec. Profesional en Prevención de Riesgos LaboralesDocument2 pagesTaller 2 Tec. Profesional en Prevención de Riesgos LaboralesAndrea ChavarroNo ratings yet
- Tema Tesis UdchDocument16 pagesTema Tesis UdchJoseSilvaNo ratings yet
- 7 DesperdiciosDocument9 pages7 DesperdiciosEstefanía Salazar RamírezNo ratings yet
- Analisis Dioses y Héroes de La Mitología GriegaDocument24 pagesAnalisis Dioses y Héroes de La Mitología Griegaefrain100% (4)
- Precisión de Los Mapas y Planos.Document2 pagesPrecisión de Los Mapas y Planos.Javier Yribarren MondejarNo ratings yet
- Calculo de EvaporacionDocument30 pagesCalculo de EvaporacionCarlos Jesus Moya EgoavilNo ratings yet
- GitlabDocument5 pagesGitlabVanessa MendozaNo ratings yet
- Armando Valdes Resumen Del Libro Embriologia PDFDocument9 pagesArmando Valdes Resumen Del Libro Embriologia PDFOmar PortelaNo ratings yet
- El NitrógenoDocument8 pagesEl Nitrógenoelvizito1991No ratings yet
- Laguna de Los MilagrosDocument11 pagesLaguna de Los MilagrosBeatriz100% (1)
- Tesis CalzatureDocument108 pagesTesis CalzatureFranklin Junior Sanchez MonjaNo ratings yet
- La BrujulaDocument42 pagesLa BrujulaFrancisco David Medrano RiveraNo ratings yet