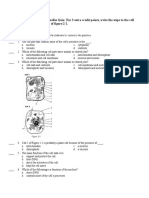
You might also like
- Chromosone AnalysisDocument234 pagesChromosone AnalysisSpataru VasileNo ratings yet
- Microbial PhysiologyDocument41 pagesMicrobial PhysiologyLiza Visbal- DionaldoNo ratings yet
- Form 4 Biology Test 3Document22 pagesForm 4 Biology Test 3Ventus TanNo ratings yet
- Unit 9 Lipid MetabolismDocument28 pagesUnit 9 Lipid MetabolismAthina100% (1)
- Biochemistry of Cheese RipeningDocument19 pagesBiochemistry of Cheese RipeningKenesei GyörgyNo ratings yet
- Sarcoma de Ewing S NEJMDocument11 pagesSarcoma de Ewing S NEJMbanzethyskyNo ratings yet
- GENBIO 1 Q1 Periodic TestDocument6 pagesGENBIO 1 Q1 Periodic TestCelsa ChicoNo ratings yet
- Hill's ReactionDocument11 pagesHill's ReactionTiyah TimothyNo ratings yet
- Jurnal Genetikaenglish 2Document9 pagesJurnal Genetikaenglish 2Leonard SitumorangNo ratings yet
- JCB 202205137Document26 pagesJCB 202205137Aleksay NesmashnyyNo ratings yet
- Vale Cell 2003 Review Molecular Motor Toolbox For Intracellular TransportDocument14 pagesVale Cell 2003 Review Molecular Motor Toolbox For Intracellular TransportSagarNo ratings yet
- Tugas Take Home GodfatherDocument27 pagesTugas Take Home GodfatherDigdo SudigyoNo ratings yet
- Emerging Roles of Mitochondria in The Evolution, Biogenesis, and Function of PeroxisomesDocument13 pagesEmerging Roles of Mitochondria in The Evolution, Biogenesis, and Function of PeroxisomeslistyNo ratings yet
- The Protein Import Machinery of Mitochondria-A Regulatory Hub in Metabolism, Stress, and DiseaseDocument16 pagesThe Protein Import Machinery of Mitochondria-A Regulatory Hub in Metabolism, Stress, and Diseaseraffaelec123No ratings yet
- Cores of Reproducibility in Physiology: CORP: Using Transgenic Mice To Study Skeletal Muscle Physiology - PMCDocument26 pagesCores of Reproducibility in Physiology: CORP: Using Transgenic Mice To Study Skeletal Muscle Physiology - PMCd2cc8hbcjcNo ratings yet
- Amino Acid ConservatismDocument18 pagesAmino Acid Conservatismjyotirmoy21No ratings yet
- Computational Biology and Chemistry: Tikam Chand Dakal, Rajender Kumar, Dindial RamotarDocument11 pagesComputational Biology and Chemistry: Tikam Chand Dakal, Rajender Kumar, Dindial RamotarMATIAS MARAMBIO SOTONo ratings yet
- Acb 43 97Document13 pagesAcb 43 97maryjoseNo ratings yet
- Connecting Two Arrays The Emerging Role of Actin Microtubule Cross Linking ProteinsDocument7 pagesConnecting Two Arrays The Emerging Role of Actin Microtubule Cross Linking ProteinsMaria Laura VilchezNo ratings yet
- The Planetary Biology of Cytochrome P450 Aromatases 2004Document14 pagesThe Planetary Biology of Cytochrome P450 Aromatases 2004Nimra Naveed ShaikhNo ratings yet
- If You Understand How Proteins WorkDocument4 pagesIf You Understand How Proteins WorkMah Song WeiNo ratings yet
- HHS Public Access: Vesicular Transport Earns A NobelDocument5 pagesHHS Public Access: Vesicular Transport Earns A NobelJoha MarinNo ratings yet
- SPERMATOGENESIS Spermatids Zirkin and Goldberg 2018Document5 pagesSPERMATOGENESIS Spermatids Zirkin and Goldberg 2018LudimilaNo ratings yet
- Paper 4Document7 pagesPaper 4Nguyen Trinh Trung DuongNo ratings yet
- Advanced Medicineprize2013Document6 pagesAdvanced Medicineprize2013MairaBarbosaNo ratings yet
- Mitotic CheckpointsDocument32 pagesMitotic CheckpointsmikefirminoNo ratings yet
- Mik Roar RayDocument39 pagesMik Roar RayDiah AmilNo ratings yet
- Itagaki 2012Document6 pagesItagaki 2012Mohammad Nazmul IslamNo ratings yet
- Dunn 2008Document25 pagesDunn 2008amandaljmonteNo ratings yet
- Female and Male Gamete Mitochondria Are Distinct and Complementary in Transcription, Structure, and Genome FunctionDocument9 pagesFemale and Male Gamete Mitochondria Are Distinct and Complementary in Transcription, Structure, and Genome FunctionardirshajiNo ratings yet
- Accepted Manuscript: Genomics, Proteomics & BioinformaticsDocument25 pagesAccepted Manuscript: Genomics, Proteomics & BioinformaticsromanNo ratings yet
- Autophagy and Disease: Always Two Sides To A Problem: Invited ReviewDocument19 pagesAutophagy and Disease: Always Two Sides To A Problem: Invited ReviewCLPHtheoryNo ratings yet
- Genome-Scale MetabolicDocument6 pagesGenome-Scale MetabolicMohamed BahgatNo ratings yet
- Cell Biology of Prokaryotic OrganellesDocument9 pagesCell Biology of Prokaryotic OrganellesHafsha May AsedreNo ratings yet
- MetabolomicsDocument12 pagesMetabolomicswatson191No ratings yet
- Important Questions For Cell BiologyDocument16 pagesImportant Questions For Cell BiologyTambudzai RazyNo ratings yet
- Sitokinesis Pada Proses Pembelahan Sel MamaliaDocument6 pagesSitokinesis Pada Proses Pembelahan Sel MamaliaAnonymous AkCt1GONo ratings yet
- Field JCellBiol 2011Document10 pagesField JCellBiol 2011huouinkyoumaNo ratings yet
- 69 PDFDocument10 pages69 PDFsriNo ratings yet
- Metabolic Fingerprints of Human Primary Endothelial and Fibroblast CellsDocument12 pagesMetabolic Fingerprints of Human Primary Endothelial and Fibroblast CellsDavid Galiano LatorreNo ratings yet
- Review: Mitochondrial Regulation in Pluripotent Stem CellsDocument8 pagesReview: Mitochondrial Regulation in Pluripotent Stem CellsSamile AmorimNo ratings yet
- Ijms 10 02611Document22 pagesIjms 10 02611Laith BioNo ratings yet
- Chromatophore Manuscript Final 03 24 19Document25 pagesChromatophore Manuscript Final 03 24 19SigmundNo ratings yet
- Meiosis PDFDocument54 pagesMeiosis PDFYusrahanna daulayNo ratings yet
- MBT31 - Proteomics NotesDocument40 pagesMBT31 - Proteomics Notesprb80730No ratings yet
- Introduction To Protein Folding For Physicists: 1 Why Study Proteins?Document53 pagesIntroduction To Protein Folding For Physicists: 1 Why Study Proteins?Alishba Faixan100% (1)
- Unesco - Eolss Sample Chapters: Evolutionary and Molecular TaxonomyDocument9 pagesUnesco - Eolss Sample Chapters: Evolutionary and Molecular Taxonomycynthiachepkoech343No ratings yet
- Nihms 421400Document16 pagesNihms 421400Sergeat18BNo ratings yet
- Optimization of Protein Expression in Mammalian CellsDocument28 pagesOptimization of Protein Expression in Mammalian Cellsmatheus.biotechNo ratings yet
- Paper de CitoesqueletoDocument4 pagesPaper de CitoesqueletoPerfil ProveedoresNo ratings yet
- Introduction To The Histology and Cell Biology of The Actin CytoskeletonDocument2 pagesIntroduction To The Histology and Cell Biology of The Actin CytoskeletonpajtgocjNo ratings yet
- The Proteomics of Plant Cell Membranes: Proteomics Selaput Sel (Pabrik/tumbuhan)Document15 pagesThe Proteomics of Plant Cell Membranes: Proteomics Selaput Sel (Pabrik/tumbuhan)Depia RohmahNo ratings yet
- Endosomal Pathway Role of TRP Channels in The Regulation of TheDocument10 pagesEndosomal Pathway Role of TRP Channels in The Regulation of ThealrokkNo ratings yet
- Elife-73808 3Document44 pagesElife-73808 3Cdotc CprimecNo ratings yet
- NIH Public Access: Author ManuscriptDocument31 pagesNIH Public Access: Author ManuscriptHutsDMNo ratings yet
- MSB 13 904Document25 pagesMSB 13 904Firda ZainabNo ratings yet
- Intro To Protein FoldingDocument53 pagesIntro To Protein FoldingRickey Castro100% (1)
- Art:10.1007/s11738 010 0538 2 PDFDocument12 pagesArt:10.1007/s11738 010 0538 2 PDFPedro ConfortNo ratings yet
- Crystals 13 00071Document11 pagesCrystals 13 00071Vaishnav SankarkNo ratings yet
- Searching For Folded Proteins And: in Vitro in SilicoDocument8 pagesSearching For Folded Proteins And: in Vitro in SilicoVenkata Suryanarayana GorleNo ratings yet
- The Mechanisms of Vesicle Budding and Fussion - Bonifacino 04Document14 pagesThe Mechanisms of Vesicle Budding and Fussion - Bonifacino 04Anahi FranchiNo ratings yet
- Molecular Basis of Morphogenesis During Vertebrate GastrulationDocument11 pagesMolecular Basis of Morphogenesis During Vertebrate GastrulationVero Claros TapiaNo ratings yet
- 2017 - Cre-loxP-Mediated Recombination - General Principles and Experimental ConsiderationsDocument12 pages2017 - Cre-loxP-Mediated Recombination - General Principles and Experimental ConsiderationsLucas PaulaNo ratings yet
- Single-Cell Metabolic Profiling of Human Cytotoxic T Cells: ArticlesDocument19 pagesSingle-Cell Metabolic Profiling of Human Cytotoxic T Cells: ArticlesShivaprakash Jagalur MuttNo ratings yet
- Single-Molecule Kinetics in Living CellsDocument32 pagesSingle-Molecule Kinetics in Living Cellstedy ekoNo ratings yet
- табл 11 Т№ 5Document1 pageтабл 11 Т№ 5Sergeat18BNo ratings yet
- Aims and Types of PatrolsDocument2 pagesAims and Types of PatrolsSergeat18BNo ratings yet
- Maintain A PK Machine GunDocument8 pagesMaintain A PK Machine GunSergeat18BNo ratings yet
- 0204SAA15HODocument1 page0204SAA15HOSergeat18BNo ratings yet
- Includepicture "../../../Temp/Finereader10/Media/Image6.Png" / Mergeformat Includepicture "../../../Temp/Finereader10/Media/Image6.Png" / MergeformatDocument1 pageIncludepicture "../../../Temp/Finereader10/Media/Image6.Png" / Mergeformat Includepicture "../../../Temp/Finereader10/Media/Image6.Png" / MergeformatSergeat18BNo ratings yet
- 0204saa12ho PDFDocument1 page0204saa12ho PDFSergeat18BNo ratings yet
- Aids To Judging Distance: Key Ranges. If You KnowDocument1 pageAids To Judging Distance: Key Ranges. If You KnowSergeat18BNo ratings yet
- 0106FC14HODocument1 page0106FC14HOSergeat18BNo ratings yet
- Duties of A Sentry: Challenge Procedure You Need To KnowDocument1 pageDuties of A Sentry: Challenge Procedure You Need To KnowSergeat18BNo ratings yet
- 0204saa17ho PDFDocument1 page0204saa17ho PDFSergeat18BNo ratings yet
- Other Aids To Judging DistanceDocument1 pageOther Aids To Judging DistanceSergeat18BNo ratings yet
- 0106FC07HODocument1 page0106FC07HOSergeat18BNo ratings yet
- Map Scales and Measuring DistancesDocument14 pagesMap Scales and Measuring DistancesSergeat18BNo ratings yet
- Finding Your Position From Local Detail: 0305MAC01OHDocument1 pageFinding Your Position From Local Detail: 0305MAC01OHSergeat18BNo ratings yet
- 0106fc02ho PDFDocument1 page0106fc02ho PDFSergeat18BNo ratings yet
- 0106FC12 13hoDocument2 pages0106FC12 13hoSergeat18BNo ratings yet
- 0106fc02ho PDFDocument1 page0106fc02ho PDFSergeat18BNo ratings yet
- 0205mac07ho PDFDocument1 page0205mac07ho PDFSergeat18BNo ratings yet
- 0205mac07ho PDFDocument1 page0205mac07ho PDFSergeat18BNo ratings yet
- 0205mac07ho PDFDocument1 page0205mac07ho PDFSergeat18BNo ratings yet
- 0105mac04ho PDFDocument1 page0105mac04ho PDFSergeat18BNo ratings yet
- Bearings: Types of BearingDocument1 pageBearings: Types of BearingSergeat18BNo ratings yet
- Finding North by The Pole Star: 3 Star Map and Compass Lesson 4Document8 pagesFinding North by The Pole Star: 3 Star Map and Compass Lesson 4Sergeat18BNo ratings yet
- Introduction, Types of Maps, Reliability and CareDocument1 pageIntroduction, Types of Maps, Reliability and CareSergeat18BNo ratings yet
- Setting A Map by Compass: 0205MAC07HODocument1 pageSetting A Map by Compass: 0205MAC07HOSergeat18BNo ratings yet
- Reliability and Marginal InformationDocument1 pageReliability and Marginal InformationSergeat18BNo ratings yet
- Grid References: NorthingsDocument2 pagesGrid References: NorthingsSergeat18BNo ratings yet
- Map Scales and Measuring DistancesDocument1 pageMap Scales and Measuring DistancesSergeat18BNo ratings yet
- Bearings: Measuring A Magnetic BearingDocument1 pageBearings: Measuring A Magnetic BearingSergeat18BNo ratings yet
- BIO101 Summary 5Document5 pagesBIO101 Summary 5LoempersNo ratings yet
- Trusight One Sequencing Panel Reference Guide 15046431 03 PDFDocument37 pagesTrusight One Sequencing Panel Reference Guide 15046431 03 PDFjoda23No ratings yet
- Mol. Basis Cheat SheetDocument3 pagesMol. Basis Cheat SheetShradha SharmaNo ratings yet
- Metabolism of XenobioticsDocument30 pagesMetabolism of XenobioticsCzarina RiveraNo ratings yet
- How To Write A Lab ReportDocument8 pagesHow To Write A Lab ReportEthan ZhengNo ratings yet
- Microbial PhysiologyDocument11 pagesMicrobial Physiologybelhadriahmed_837375No ratings yet
- DNA Paper Models ActivityDocument8 pagesDNA Paper Models ActivityLouise Meara SeveroNo ratings yet
- Topic 1.3 Revision Practice (Ans)Document3 pagesTopic 1.3 Revision Practice (Ans)Mahina WongNo ratings yet
- Comprehensive Drug Study Ketoanalogue PDF FreeDocument2 pagesComprehensive Drug Study Ketoanalogue PDF FreeYuuki Chitose (tai-kun)No ratings yet
- 2.2 Biological Molecules - Proteins QPDocument24 pages2.2 Biological Molecules - Proteins QPsuccesshustlerclubNo ratings yet
- Evaluation Enzyme Inhibitors Drug Discovery PDFDocument2 pagesEvaluation Enzyme Inhibitors Drug Discovery PDFKarlaNo ratings yet
- Prescott Harley Klein S Microbiology 7th Edition - (Nosnibor) Ebook PDF - 212-245 PDFDocument34 pagesPrescott Harley Klein S Microbiology 7th Edition - (Nosnibor) Ebook PDF - 212-245 PDFAli AwanNo ratings yet
- Biol 2334 - Molecular Biology Chapter 5 Worksheet - Replication & RepairDocument6 pagesBiol 2334 - Molecular Biology Chapter 5 Worksheet - Replication & RepairJohn Paradise100% (1)
- Biochem Transes XXXDocument59 pagesBiochem Transes XXXJanica Pauline DaydayNo ratings yet
- Medical Progress: Review ArticlesDocument15 pagesMedical Progress: Review ArticlesFeri EspinosaSerranoNo ratings yet
- Chapter 7 Section 2Document3 pagesChapter 7 Section 2Pak RisNo ratings yet
- Mass-Spectrometry-Based Draft of TheDocument28 pagesMass-Spectrometry-Based Draft of ThePurboPathikNo ratings yet
- Quiz in Lipids (G1)Document3 pagesQuiz in Lipids (G1)Wayne David C. PadullonNo ratings yet
- Chapter - 2 PDFDocument32 pagesChapter - 2 PDFnikhilNo ratings yet
- Chemicals of LifeDocument44 pagesChemicals of Lifes17m0582No ratings yet
- SundariDocument8 pagesSundariRani Eva DewiNo ratings yet
- IB Biology SL Topics Study GuideDocument65 pagesIB Biology SL Topics Study GuideSai 0235No ratings yet
- 14 - Makalah AlelopatiDocument18 pages14 - Makalah Alelopaticutazuraizatul nufusNo ratings yet