You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Wolbachia - Master Manipulators of Invertebrate BiologyDocument12 pagesWolbachia - Master Manipulators of Invertebrate BiologyAlita BelleNo ratings yet
- Insect Symbiosis Volume 1Document270 pagesInsect Symbiosis Volume 1Ed Branco VictorNo ratings yet
- Wolbachia Write Up For IPTKFINAL PDFDocument13 pagesWolbachia Write Up For IPTKFINAL PDFdzc2016No ratings yet
- What Makes Community Engagement Effective2Document19 pagesWhat Makes Community Engagement Effective2dzc2016No ratings yet
- Wolbachia Strain Reduces Transmission of Zika VirusDocument7 pagesWolbachia Strain Reduces Transmission of Zika Virusdzc2016No ratings yet
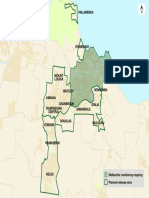
- Townsville Release MapDocument1 pageTownsville Release Mapdzc2016No ratings yet
- Community Participation in Dengue Control in BrazilDocument10 pagesCommunity Participation in Dengue Control in Brazildzc2016No ratings yet
- Invasion of Wolbachia at The Residential Block Level Is Associated With Local Abundance of Stegomyia and Property AttributesDocument8 pagesInvasion of Wolbachia at The Residential Block Level Is Associated With Local Abundance of Stegomyia and Property Attributesdzc2016No ratings yet
- Wolbachia SpreadDocument2 pagesWolbachia Spreaddzc2016No ratings yet
- Field Evaluation of The Establishment Wolbachia Project in VietnamDocument14 pagesField Evaluation of The Establishment Wolbachia Project in VietnamNik FajarNo ratings yet
- Broader Prevalence of Wolbachia in Insects Including Potential Human Disease VectorsDocument11 pagesBroader Prevalence of Wolbachia in Insects Including Potential Human Disease Vectorsdzc2016No ratings yet
- Establishment of A Wolbachia Superinfection in Aedes Aegypti Mosquitoes As A Potential Approach For Future Resistance ManagementDocument19 pagesEstablishment of A Wolbachia Superinfection in Aedes Aegypti Mosquitoes As A Potential Approach For Future Resistance Managementdzc2016No ratings yet
- Risk Associated With The Release of Wolbachia-Infected Aedes Aegypti Mosquitoes Into The Environment in An Effort To Control Dengue PDFDocument12 pagesRisk Associated With The Release of Wolbachia-Infected Aedes Aegypti Mosquitoes Into The Environment in An Effort To Control Dengue PDFdzc2016No ratings yet
- Designing A Community Engagement FrameworkDocument12 pagesDesigning A Community Engagement Frameworkdzc2016No ratings yet
- Wolbachia - ZikaDocument5 pagesWolbachia - Zikadzc2016No ratings yet
- Successful Establishment of Wolbachia in Aedes Populations To Surppress Dengue Transmission PDFDocument6 pagesSuccessful Establishment of Wolbachia in Aedes Populations To Surppress Dengue Transmission PDFdzc2016No ratings yet
- Stability of The Wmel Wolbachia Infection Following Invasion Into Aedes Aegypti Population PDFDocument9 pagesStability of The Wmel Wolbachia Infection Following Invasion Into Aedes Aegypti Population PDFdzc2016No ratings yet
- Wolbachia Write Up For IPTKFINAL PDFDocument13 pagesWolbachia Write Up For IPTKFINAL PDFdzc2016No ratings yet
- Successful Establishment of Wolbachia in Aedes Populations To Surppress Dengue TransmissionDocument1 pageSuccessful Establishment of Wolbachia in Aedes Populations To Surppress Dengue Transmissiondzc2016No ratings yet
- Successful Establishment of Wolbachia in Aedes Populations To Surppress Dengue Transmission PDFDocument6 pagesSuccessful Establishment of Wolbachia in Aedes Populations To Surppress Dengue Transmission PDFdzc2016No ratings yet
- Stability of The Wmel Wolbachia Infection Following Invasion Into Aedes Aegypti Population PDFDocument9 pagesStability of The Wmel Wolbachia Infection Following Invasion Into Aedes Aegypti Population PDFdzc2016No ratings yet
- Rear and Release A New Paradigm For Dengue ControlDocument5 pagesRear and Release A New Paradigm For Dengue Controldzc2016No ratings yet
- Risk Associated With The Release of Wolbachia-Infected Aedes Aegypti Mosquitoes Into The Environment in An Effort To Control Dengue PDFDocument12 pagesRisk Associated With The Release of Wolbachia-Infected Aedes Aegypti Mosquitoes Into The Environment in An Effort To Control Dengue PDFdzc2016No ratings yet
- Bibliografia GeneralDocument340 pagesBibliografia GeneralalfonsoNo ratings yet
- Kajian Pemanfaatan Wolbachia Terhadap Pengendalian DBD Studi Literatur Dan Studi Kasus Pemanfaatan Wolbachia Di YogyakartaDocument14 pagesKajian Pemanfaatan Wolbachia Terhadap Pengendalian DBD Studi Literatur Dan Studi Kasus Pemanfaatan Wolbachia Di Yogyakartajihan OktafianiNo ratings yet
- MiteDocument216 pagesMiteAnggry SolihinNo ratings yet
- BIOTECH - DOST Tawa Tawa Food SupplementDocument7 pagesBIOTECH - DOST Tawa Tawa Food SupplementJoven CedricNo ratings yet
- Fire Ant Control With Entomopathogens in The UsaDocument21 pagesFire Ant Control With Entomopathogens in The UsaRichar Manuel Simanca FontalvoNo ratings yet
- 2021 - Wolbachia Yogya Final Research Report - AWED CompressedDocument100 pages2021 - Wolbachia Yogya Final Research Report - AWED Compressed0645 AMNo ratings yet
- Ab182diseases of Monarchs and Other ButterfliesDocument6 pagesAb182diseases of Monarchs and Other ButterfliesGustavo ZaRoNo ratings yet
- Resistance To Insecticides in Anopheles Gambiae Complex in West Africa: A Review of The Current Situation and The Perspectives For Malaria ControlDocument13 pagesResistance To Insecticides in Anopheles Gambiae Complex in West Africa: A Review of The Current Situation and The Perspectives For Malaria ControlDuvánE.DueñasLópezNo ratings yet
- Arthropod Mass Rearing and Quality Control: Iobc OilbDocument67 pagesArthropod Mass Rearing and Quality Control: Iobc OilbFranJim FjdNo ratings yet
- Ecpe-Readvoc U4 SB Sample-UnitDocument12 pagesEcpe-Readvoc U4 SB Sample-UnitchristineNo ratings yet
- Sex Ratio DistortionDocument17 pagesSex Ratio DistortionAdam ZHUNo ratings yet
- Weekend Challenge 08 - KeyDocument5 pagesWeekend Challenge 08 - KeyQuỳnh NguyễnNo ratings yet
- Jurnal PoliketidaDocument43 pagesJurnal PoliketidaRama HidayatNo ratings yet
- A Climate Resilient Singapore For A Sustainable FutureDocument36 pagesA Climate Resilient Singapore For A Sustainable FutureHandikaNo ratings yet
- Be Uke Boom 2012rty54yDocument5 pagesBe Uke Boom 2012rty54ySyahroniNo ratings yet
- Mazur 2009Document2 pagesMazur 2009Syakti PerdanaNo ratings yet
- TephritidaeDocument86 pagesTephritidaePABLO FIGUEROA CARDENASNo ratings yet
- Definitions of Parasitism Considering Its Potentially Opposing Effects at Different Levels of Hierarchical OrganizationDocument8 pagesDefinitions of Parasitism Considering Its Potentially Opposing Effects at Different Levels of Hierarchical OrganizationwellingtonlimaNo ratings yet
- 01.wolbachia Masseliensis - YLDocument20 pages01.wolbachia Masseliensis - YLYounes LaidoudiNo ratings yet
- PNTD 0008805Document32 pagesPNTD 0008805Pawan MishraNo ratings yet
- Microbial Interactions PDFDocument12 pagesMicrobial Interactions PDFnabeela tabassumNo ratings yet
- 8d9c PDFDocument11 pages8d9c PDFJulian AshbourneNo ratings yet
- PARASITIC NEMATODES Molecular Biology, Biochemistry and ImmunologyDocument498 pagesPARASITIC NEMATODES Molecular Biology, Biochemistry and Immunologydouglasman100% (3)
- The Impact of Wolbachia On Virus InfectionDocument13 pagesThe Impact of Wolbachia On Virus InfectionShirly DiazgranadosNo ratings yet
- Nature Magazine 7158 - 2007-09-06Document125 pagesNature Magazine 7158 - 2007-09-06Roberto KlesNo ratings yet
- Lab 1: Arthropod Collection & Identification: Student GuideDocument15 pagesLab 1: Arthropod Collection & Identification: Student GuidePAU FRANCISCONo ratings yet
- A Review of The Natural Enemies of Beetles in The Subtribe Diabroticina (Coleoptera: Chrysomelidae) : Implications For Sustainable Pest ManagementDocument65 pagesA Review of The Natural Enemies of Beetles in The Subtribe Diabroticina (Coleoptera: Chrysomelidae) : Implications For Sustainable Pest ManagementBlue Dasher FarmNo ratings yet
- 05NTD 2022 - UPDATES IN VECTOR CONTROL IN MALAYSIA: What Is New?Document61 pages05NTD 2022 - UPDATES IN VECTOR CONTROL IN MALAYSIA: What Is New?Institute for Clinical ResearchNo ratings yet