You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Gardes1993 Primers de FungosDocument6 pagesGardes1993 Primers de FungosDiego De Araujo FrazilioNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Occurrence and Function of Yeast in Asian Indigenous Fermented Foods.Document10 pagesOccurrence and Function of Yeast in Asian Indigenous Fermented Foods.J Alberto LucasNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Biodiversity of Non-Saccharomyces Yeast in Distilleries of The La Mancha Region (Spain) .Document11 pagesBiodiversity of Non-Saccharomyces Yeast in Distilleries of The La Mancha Region (Spain) .J Alberto LucasNo ratings yet
- Ponting, Et Al 2009. Evolution and Functions of Long Noncoding RNAs.Document13 pagesPonting, Et Al 2009. Evolution and Functions of Long Noncoding RNAs.J Alberto LucasNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Function of Yeast Species and Strains in Wine FlavourDocument12 pagesFunction of Yeast Species and Strains in Wine FlavourisatburNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Baleiras-Cuoto Et Al. 1996 - Evaluation of Molecular Typing Techniques To Assign Genetic Diversity Among Saccharomyces Cerevisiae Strains.Document6 pagesBaleiras-Cuoto Et Al. 1996 - Evaluation of Molecular Typing Techniques To Assign Genetic Diversity Among Saccharomyces Cerevisiae Strains.J Alberto LucasNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Furstenberg-Hagg Et Al. 2013. Plant Defense Against Insect Herbivores.Document56 pagesFurstenberg-Hagg Et Al. 2013. Plant Defense Against Insect Herbivores.J Alberto LucasNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Fermentations in World Food Processing.Document10 pagesFermentations in World Food Processing.J Alberto LucasNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Cagno, Et Al. 2010 - Taxonomic Structure of The Yeast and Lactic Acid Bacteria Microbiota of Pineapple (Anana Comosus L. Merr) and Use of Autochthonous Starters For Minimally Processing.Document9 pagesCagno, Et Al. 2010 - Taxonomic Structure of The Yeast and Lactic Acid Bacteria Microbiota of Pineapple (Anana Comosus L. Merr) and Use of Autochthonous Starters For Minimally Processing.J Alberto LucasNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Arnegard, Et Al 2014. Genetics of Ecological Divergence During Speciation.Document17 pagesArnegard, Et Al 2014. Genetics of Ecological Divergence During Speciation.J Alberto LucasNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Amzad and Rahman, 2011 - Total Phenolics, Flavonoids and Antioxidant Activity of Tropical Fruit Pineapple.Document5 pagesAmzad and Rahman, 2011 - Total Phenolics, Flavonoids and Antioxidant Activity of Tropical Fruit Pineapple.J Alberto Lucas100% (1)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Phyto 96 0195Document12 pagesPhyto 96 0195J Alberto LucasNo ratings yet
- Gonzalez, Et Al. 2006. Molecular Identification and Characterization of Wine Yeast Isolated From Tenerife (Canarias Island, Spain)Document8 pagesGonzalez, Et Al. 2006. Molecular Identification and Characterization of Wine Yeast Isolated From Tenerife (Canarias Island, Spain)J Alberto LucasNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Blandino, Et Al. 2003. Cereal-Based Fermented Foods and Beverages.Document17 pagesBlandino, Et Al. 2003. Cereal-Based Fermented Foods and Beverages.J Alberto LucasNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Evolutionary Ecology of Ethanol Production and AlcoholismDocument7 pagesThe Evolutionary Ecology of Ethanol Production and AlcoholismJ Alberto LucasNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Biology of Pineapple (Ananas comosus var. comosusDocument43 pagesThe Biology of Pineapple (Ananas comosus var. comosusJ Alberto LucasNo ratings yet
- 1 s2.0 S0168160502001940 MainDocument9 pages1 s2.0 S0168160502001940 MainJ Alberto LucasNo ratings yet
- Monoterpene Barkbeetles AdamsDocument10 pagesMonoterpene Barkbeetles AdamsJ Alberto LucasNo ratings yet
- Algebra PracticeDocument3 pagesAlgebra PracticeJ Alberto LucasNo ratings yet
- 1 s2.0 0168160594900612 MainDocument9 pages1 s2.0 0168160594900612 MainJ Alberto LucasNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- 1 s2.0 S016816050500468X MainDocument11 pages1 s2.0 S016816050500468X MainJ Alberto LucasNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Poty VirusDocument24 pagesPoty VirusJ Alberto LucasNo ratings yet
- Beginnings of Microbiology and Biochemistry - The Contribution of Yeast Research.Document11 pagesBeginnings of Microbiology and Biochemistry - The Contribution of Yeast Research.J Alberto LucasNo ratings yet
- Simplified mtDNA analysis for food yeast identificationDocument7 pagesSimplified mtDNA analysis for food yeast identificationJ Alberto LucasNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- 1 s2.0 S016816050500468X MainDocument11 pages1 s2.0 S016816050500468X MainJ Alberto LucasNo ratings yet
- A Review of Rapid Metods For The Analysis of Ins Zheng, M. Z. Et Al. 2006Document13 pagesA Review of Rapid Metods For The Analysis of Ins Zheng, M. Z. Et Al. 2006J Alberto LucasNo ratings yet
- Encyclopedia of Dinosaurs and Prehistoric LifeDocument378 pagesEncyclopedia of Dinosaurs and Prehistoric LifeJ Alberto Lucas75% (12)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- MODULE 3 - Revisiting The ClassicsDocument22 pagesMODULE 3 - Revisiting The ClassicsSneha MichaelNo ratings yet
- Mark Scheme (Results) : Summer 2018Document22 pagesMark Scheme (Results) : Summer 2018RafaNo ratings yet
- Using ABAQUS to Reduce Tire Belt Edge StressesDocument10 pagesUsing ABAQUS to Reduce Tire Belt Edge StressesGautam SharmaNo ratings yet
- DFA vs NLRC Immunity DisputeDocument7 pagesDFA vs NLRC Immunity DisputeThrees SeeNo ratings yet
- FCE - Exam: B2 First For SchoolsDocument26 pagesFCE - Exam: B2 First For SchoolsNele Loubele PrivateNo ratings yet
- Compact Concealed Handgun Comparison Chart (Illustrated) - 2011Document10 pagesCompact Concealed Handgun Comparison Chart (Illustrated) - 2011KomodowaranNo ratings yet
- Students' Perception of Online English Learning During Covid-19 PandemicDocument5 pagesStudents' Perception of Online English Learning During Covid-19 Pandemicfadhilah santriNo ratings yet
- Pamela Presentation PDFDocument8 pagesPamela Presentation PDFawtshfhdNo ratings yet
- Organization and Operation of Partnerships and LLPsDocument50 pagesOrganization and Operation of Partnerships and LLPsAliaa Hosam100% (1)
- PBA Physics SSC-I FinalDocument7 pagesPBA Physics SSC-I Finalrayyanhashmi93No ratings yet
- The Myth of Scientific Miracles in The KoranDocument13 pagesThe Myth of Scientific Miracles in The KoranDoctor Jones33% (3)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Unit 11 Multiple Correlation: StructureDocument18 pagesUnit 11 Multiple Correlation: StructurePranav ViswanathanNo ratings yet
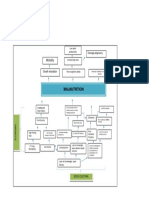
- Problem Tree Analysis MalnutritionDocument1 pageProblem Tree Analysis MalnutritionMho Dsb50% (6)
- Pon Vidyashram Group of Cbse Schools STD 8 SCIENCE NOTES (2020-2021)Document3 pagesPon Vidyashram Group of Cbse Schools STD 8 SCIENCE NOTES (2020-2021)Bharath Kumar 041No ratings yet
- Indikasi, Teknik Pembuatan Stoma, ReanastomosispptDocument43 pagesIndikasi, Teknik Pembuatan Stoma, ReanastomosispptErwin Aritama Ismail100% (1)
- FORMS Transaction DisputeDocument1 pageFORMS Transaction DisputeKristine Cruz-MaltoNo ratings yet
- Health Communication BJMCDocument42 pagesHealth Communication BJMCxyz_171274767100% (3)
- Mama and Mommy and Me in The Middle Press ReleaseDocument2 pagesMama and Mommy and Me in The Middle Press ReleaseCandlewick Press0% (1)
- A2 AQA History Britain NotesDocument7 pagesA2 AQA History Britain Notesjazzybob0% (1)
- NATO Intelligence Exploitation of The InternetDocument104 pagesNATO Intelligence Exploitation of The Internetmauro.pennacchietti100% (1)
- Public Service Commission, West Bengal: Notice Inviting Expression of Interest For Enlistment As LawyersDocument3 pagesPublic Service Commission, West Bengal: Notice Inviting Expression of Interest For Enlistment As LawyersITI JobNo ratings yet
- AnalogyDocument50 pagesAnalogyNathaniel Campeciño89% (9)
- The Cell Cycle: How Cells Grow and DivideDocument21 pagesThe Cell Cycle: How Cells Grow and DivideAqsa YaminNo ratings yet
- Overexploitation of Parrots in The NeotropicsDocument6 pagesOverexploitation of Parrots in The NeotropicsElenananaNo ratings yet
- 5 All India Mobile Database SampleDocument15 pages5 All India Mobile Database Sampleali khan Saifi100% (1)
- BioassayDocument38 pagesBioassayMuhammad Masoom AkhtarNo ratings yet
- Environmental Sustainability Assessment of An Academic Institution in Calamba CityDocument29 pagesEnvironmental Sustainability Assessment of An Academic Institution in Calamba CityAmadeus Fernando M. PagenteNo ratings yet
- Miro's Response To MotionDocument6 pagesMiro's Response To Motional_crespoNo ratings yet
- 1 Calimutan v. People - G.R. No. 152133Document1 page1 Calimutan v. People - G.R. No. 152133eiram23100% (1)
- UP Law Bar Exam Suggested AnswersDocument16 pagesUP Law Bar Exam Suggested AnswersprincessmagpatocNo ratings yet
- Forever Strong: A New, Science-Based Strategy for Aging WellFrom EverandForever Strong: A New, Science-Based Strategy for Aging WellNo ratings yet
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Periodic Tales: A Cultural History of the Elements, from Arsenic to ZincFrom EverandPeriodic Tales: A Cultural History of the Elements, from Arsenic to ZincRating: 3.5 out of 5 stars3.5/5 (137)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Metabolism Revolution: Lose 14 Pounds in 14 Days and Keep It Off for LifeFrom EverandMetabolism Revolution: Lose 14 Pounds in 14 Days and Keep It Off for LifeNo ratings yet
- Alex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessFrom EverandAlex & Me: How a Scientist and a Parrot Discovered a Hidden World of Animal Intelligence—and Formed a Deep Bond in the ProcessNo ratings yet
- Summary of Mary Claire Haver's The Galveston DietFrom EverandSummary of Mary Claire Haver's The Galveston DietRating: 5 out of 5 stars5/5 (1)