You might also like
- Esructuras Cerebrales en La Percepción PDFDocument4 pagesEsructuras Cerebrales en La Percepción PDFJeimmy Garcia MarinNo ratings yet
- Estructuras Cerebrales Percepción, Atención y MemoriaDocument7 pagesEstructuras Cerebrales Percepción, Atención y MemoriaChecho AlegriaNo ratings yet
- Neuroplasticidad O.foster M.terzaghiDocument16 pagesNeuroplasticidad O.foster M.terzaghinicole50% (2)
- Corteza Prefrontal RostrolateralDocument9 pagesCorteza Prefrontal RostrolateralJulian David CuervoNo ratings yet
- NEUROPSICOLOGIA. Caso Practico-Pablo Ayala.Document27 pagesNEUROPSICOLOGIA. Caso Practico-Pablo Ayala.Psicologia Uac Valdivia UacNo ratings yet
- Las Redes AtencionalesDocument9 pagesLas Redes AtencionalesJaime Muñoz Insunza0% (1)
- Dialnet ComprendiendoElCodigoNeuronal 7855462Document6 pagesDialnet ComprendiendoElCodigoNeuronal 7855462Gustavo Alarcón ZbindenNo ratings yet
- 3 en EsDocument13 pages3 en EsLuis LeonNo ratings yet
- Funciones IntelectualesDocument5 pagesFunciones IntelectualesLRI19040 - CECILIA GARCIA NI�ONo ratings yet
- Tirapu y Muñoz, 2005 Memoria y Funciones EjecutivasDocument10 pagesTirapu y Muñoz, 2005 Memoria y Funciones Ejecutivaspame05220825No ratings yet
- Chen, JL, Zatorre, RJ y Penhune, VB (2008) ..En - EsDocument11 pagesChen, JL, Zatorre, RJ y Penhune, VB (2008) ..En - EsVerónica BenavidezNo ratings yet
- NeurocienciaDocument23 pagesNeurocienciaCarlos RuizNo ratings yet
- Lobulo FrontalDocument8 pagesLobulo FrontalAngel Daniel VásquezNo ratings yet
- Luria DnaDocument9 pagesLuria DnaJoha RocafuerteNo ratings yet
- NiendamDocument21 pagesNiendamMildred HickieNo ratings yet
- Cerebro y ConductaDocument18 pagesCerebro y ConductaSheila Canal SáezNo ratings yet
- 2015 Organización Motora Del Córtex Cerebral y El Papel Del Sistema de Las Neuronas Espejo. Repercusiones Clínicas para La RehabilitaciónDocument5 pages2015 Organización Motora Del Córtex Cerebral y El Papel Del Sistema de Las Neuronas Espejo. Repercusiones Clínicas para La RehabilitaciónCrisSanchezCarrascoNo ratings yet
- Los Lóbulos Temporales Anteriores y La Arquitectura Funcional de Traduccion PaperDocument8 pagesLos Lóbulos Temporales Anteriores y La Arquitectura Funcional de Traduccion PaperestefaniaNo ratings yet
- de La Especialización Funcional Al Conectoma Del Cerebro HumanoDocument25 pagesde La Especialización Funcional Al Conectoma Del Cerebro HumanoCoral LunaNo ratings yet
- Plasticidad Sináptica Duradera (LTP) : Un Punto de Partida para Entender Los Procesos de Aprendizaje y Memoria.Document51 pagesPlasticidad Sináptica Duradera (LTP) : Un Punto de Partida para Entender Los Procesos de Aprendizaje y Memoria.ILEANA PATRICIA VALENZUELA AYON100% (1)
- Seminario 4Document31 pagesSeminario 4David DiazNo ratings yet
- Analisis Neuropsicologico de Las Dificultades en La Lectoescritura, Quintanar PDFDocument13 pagesAnalisis Neuropsicologico de Las Dificultades en La Lectoescritura, Quintanar PDFJAZMIN ALEJANDRA HERNANDEZ - ENRIQUEZ100% (1)
- O 1hfppa0ajiqjr5f150915jf1k2ocDocument7 pagesO 1hfppa0ajiqjr5f150915jf1k2ocADRIANANo ratings yet
- Fisiología de La CogniciónDocument12 pagesFisiología de La CogniciónEdunexosNo ratings yet
- 1 Neurociencias GeneralidadesDocument6 pages1 Neurociencias GeneralidadesLaura LopezNo ratings yet
- Cuello CallosoDocument6 pagesCuello CallosoSandra MartinezNo ratings yet
- Teoria Disfuncion EjecutivaDocument24 pagesTeoria Disfuncion EjecutivaKatherine CuetoNo ratings yet
- Neurociencia. 1Document8 pagesNeurociencia. 1Ricardo Alejandro León MuñozNo ratings yet
- Cognitivas Lobulo Frontal PDFDocument5 pagesCognitivas Lobulo Frontal PDFVale DoteNo ratings yet
- Actividad de Reflexion 1Document3 pagesActividad de Reflexion 1Daniela Molina RuizNo ratings yet
- Neurocomputacion y Modelizacion de Procesos CognitivosDocument23 pagesNeurocomputacion y Modelizacion de Procesos CognitivosManuel LunaNo ratings yet
- Bateria de Luria DNA PDFDocument21 pagesBateria de Luria DNA PDFX.G100% (1)
- LIPINA (Conceptos Neurobiologícos Básicos)Document8 pagesLIPINA (Conceptos Neurobiologícos Básicos)c s tNo ratings yet
- Plasticidad Del Sistema Nervioso y Recuperación de FuncionesDocument20 pagesPlasticidad Del Sistema Nervioso y Recuperación de FuncionesErika Moreno de LeónNo ratings yet
- Contribución Del Cerebelo A Los Procesos Cognitivos AVANCES ACTUALESDocument40 pagesContribución Del Cerebelo A Los Procesos Cognitivos AVANCES ACTUALESAcintiaNo ratings yet
- Ensayo Modelos NeuropsicologicosDocument6 pagesEnsayo Modelos NeuropsicologicosClaudia MuñozNo ratings yet
- El Sistema de Atención Del Cerebro HumanoDocument22 pagesEl Sistema de Atención Del Cerebro HumanoAngel de la VegaNo ratings yet
- Algunos Conceptos Neurobiologicos Basicos - LIPINADocument10 pagesAlgunos Conceptos Neurobiologicos Basicos - LIPINAantugaiaNo ratings yet
- 1 Síndromes FrontalesDocument4 pages1 Síndromes FrontalesJonathan-Patty Rivero100% (1)
- Trabajo ForoDocument5 pagesTrabajo ForoKevin GuevaraNo ratings yet
- La Especialización Hemisférica y La Regulación de La Conducta Motora Desde La Perspectiva de La Neurociencia CognitivaDocument8 pagesLa Especialización Hemisférica y La Regulación de La Conducta Motora Desde La Perspectiva de La Neurociencia CognitivaldelblancodiezNo ratings yet
- Cómo Funcionan Los Circuitos CerebralesDocument7 pagesCómo Funcionan Los Circuitos CerebralesDaniel Cordero HernándezNo ratings yet
- Apunte de Aprendizaje 1Document12 pagesApunte de Aprendizaje 1Gabriela Iliana Toro OrtizNo ratings yet
- Funciones Cognitivas Del Lobulo FrontalDocument5 pagesFunciones Cognitivas Del Lobulo FrontalCésar Argumedos100% (1)
- Conceptos Fundamentales en La Teoria NeuropsicolgicaDocument8 pagesConceptos Fundamentales en La Teoria NeuropsicolgicaSandra MilenaNo ratings yet
- La Teoría Atencional de PosnerDocument15 pagesLa Teoría Atencional de PosnerHugo López100% (1)
- NeuroplasticidadDocument29 pagesNeuroplasticidadmauro100% (2)
- El CerebeloDocument7 pagesEl Cerebeloquispearapaabraham452No ratings yet
- Los Estudios Neurobiológicos de La ConductaDocument4 pagesLos Estudios Neurobiológicos de La ConductaItzel ChavezNo ratings yet
- Bases Teoricas y Clinicas Del Diagnostico y La Rehabilitacion NeuropsicologicaDocument8 pagesBases Teoricas y Clinicas Del Diagnostico y La Rehabilitacion NeuropsicologicaNtyFerNo ratings yet
- El Cerebelo y Sus FuncionesDocument7 pagesEl Cerebelo y Sus FuncionesAhura MazdaNo ratings yet
- Taller Corteza PrefrontalDocument2 pagesTaller Corteza PrefrontalIgnacio IvánNo ratings yet
- Compendio NeurpsicoDocument26 pagesCompendio NeurpsicodamianNo ratings yet
- L5-Mecanismos Elementales de La Plasticidad Del Sistema Nervioso CentralDocument9 pagesL5-Mecanismos Elementales de La Plasticidad Del Sistema Nervioso CentralJosé Carlos Bello LópezNo ratings yet
- Ritmos CerebralesDocument6 pagesRitmos CerebralescesardlNo ratings yet
- Biologia de Las Neuronas Espejo Relacion Con El Lenguaje, La Empatia y Determinados Trastornos de La Conducta.Document33 pagesBiologia de Las Neuronas Espejo Relacion Con El Lenguaje, La Empatia y Determinados Trastornos de La Conducta.Paula NapoliNo ratings yet
- Sistema nervioso: Neuroanatomía funcional y clínicaFrom EverandSistema nervioso: Neuroanatomía funcional y clínicaRating: 4 out of 5 stars4/5 (4)
- Doppler Vasos de Cuello Sistematica e InformeDocument4 pagesDoppler Vasos de Cuello Sistematica e Informefher14bcNo ratings yet
- Gastrulacion y EmbriologiaDocument17 pagesGastrulacion y EmbriologiaCecilia CVocalNo ratings yet
- YogumonDocument30 pagesYogumonMagica AlegriaNo ratings yet
- Relación Entre Grado de Anteposición de Cabeza y Cuello y Los Parámetros Cefalométricos de RocabadoDocument81 pagesRelación Entre Grado de Anteposición de Cabeza y Cuello y Los Parámetros Cefalométricos de RocabadoJose CollazosNo ratings yet
- Criterio MUSADocument37 pagesCriterio MUSAClia Maryluz100% (2)
- Policitemia NeonatalDocument20 pagesPolicitemia NeonatalMarco LojaNo ratings yet
- Capitulo 5 PDFDocument41 pagesCapitulo 5 PDFJULIANo ratings yet
- Equipos de Respiracion AutónomaDocument12 pagesEquipos de Respiracion AutónomaMarcelo Peñaloza TapiaNo ratings yet
- Parcial IIDocument8 pagesParcial IILorenaNo ratings yet
- Regulación de La Temperatura Corporal AbigailDocument9 pagesRegulación de La Temperatura Corporal AbigailIrving MayoNo ratings yet
- Fisiologia Emb PielDocument35 pagesFisiologia Emb PielRossana ValienteNo ratings yet
- Tema 6Document10 pagesTema 6Federico DcfNo ratings yet
- Denise A. Agnew - Corazones OcultosDocument26 pagesDenise A. Agnew - Corazones OcultosMew Rincone100% (1)
- Emociones y Sistema Inmune, PPT Autor: Dr. Orestes García FigueiredoDocument61 pagesEmociones y Sistema Inmune, PPT Autor: Dr. Orestes García FigueiredoNancySáezNo ratings yet
- Oido HistologiaDocument6 pagesOido HistologiaMaría Fernanda RiberaNo ratings yet
- 17 Paleopatoogia OralDocument11 pages17 Paleopatoogia OralhumbertoNo ratings yet
- AM C1 1 Clase1Document24 pagesAM C1 1 Clase1Sebastian Andre Pasten FuenzalidaNo ratings yet
- Fisiopatologia de La Gastritis y UlceraDocument12 pagesFisiopatologia de La Gastritis y UlceraANA PATRICIA MENDOZA MONTEALEGRENo ratings yet
- ESTRÉSDocument24 pagesESTRÉSEunice Zaragoza LealNo ratings yet
- La Investigación de La Hemoglobina y Los Orígenes de La Medicina MolecularDocument27 pagesLa Investigación de La Hemoglobina y Los Orígenes de La Medicina MolecularsalvadorNo ratings yet
- Iper SFC - Oficinas AdministrativasDocument3 pagesIper SFC - Oficinas AdministrativasTATITO030609101250% (2)
- Enfermedad Coronaria en Hombres y MujeresDocument6 pagesEnfermedad Coronaria en Hombres y MujeresDuncan MaxNo ratings yet
- PolimiositisDocument7 pagesPolimiositisapi-3698281100% (2)
- Grasa Despojos Visceras y SangreDocument12 pagesGrasa Despojos Visceras y Sangrejoel dante pomayay yaranga100% (1)
- Ajuste de Articulacion Sacroiliac ADocument7 pagesAjuste de Articulacion Sacroiliac Agustavogomez4505No ratings yet
- DKV LleidaDocument190 pagesDKV LleidaluckasaNo ratings yet
- Enzimas en ClínicaDocument45 pagesEnzimas en ClínicaLilian ArchivosNo ratings yet
- Ojo y PielDocument16 pagesOjo y PielJavier López AlisteNo ratings yet
- Farmacología y AerosolterapiaDocument81 pagesFarmacología y AerosolterapiaAlejandra VasquezNo ratings yet
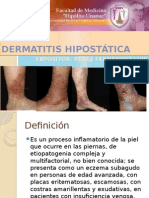
- Dermatitis HipostáticaDocument14 pagesDermatitis HipostáticaLuis Angel Perez FernandezNo ratings yet