You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Conference on Global Issues in Agricultural, Environmental and Forestry SciencesDocument16 pagesConference on Global Issues in Agricultural, Environmental and Forestry SciencesShirish NarnawareNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Physiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamDocument61 pagesPhysiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamZaigham HammadNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Muscle Energy Techniques ExplainedDocument2 pagesMuscle Energy Techniques ExplainedAtanu Datta100% (1)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
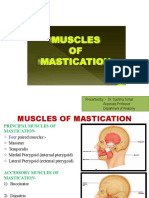
- Presented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyDocument20 pagesPresented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyCristine EchaveNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
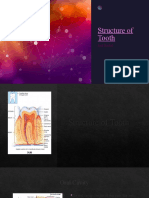
- Structure of Tooth m121Document13 pagesStructure of Tooth m121amirhossein vaeziNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- 2010 FulltextDocument410 pages2010 FulltextNajam Din SahitoNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Trunking CatalogDocument8 pagesTrunking CatalogDenuka PathiranaNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Correlation of Dynamic Strength in The Standing Calf Raise With Sprinting Performance in Consecutive Sections Up To 30 MetersDocument9 pagesCorrelation of Dynamic Strength in The Standing Calf Raise With Sprinting Performance in Consecutive Sections Up To 30 MetersJohn NixonNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Pain PathwayDocument17 pagesPain PathwaySalsabila Al-BasheerNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- Classification and Characterization of Microsatellite Instability Across 18 Cancer TypesDocument13 pagesClassification and Characterization of Microsatellite Instability Across 18 Cancer TypeshadymatrixNo ratings yet
- Ap Environmental Science Free-Response QuestionsDocument6 pagesAp Environmental Science Free-Response Questionsapi-234419767No ratings yet
- MYCOVIRODocument11 pagesMYCOVIROPau SorianoNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Life in The Deserts Worksheet 2Document2 pagesLife in The Deserts Worksheet 2Uma TiwariNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Reproductive Biology and Phylogeny of Birds PDFDocument624 pagesReproductive Biology and Phylogeny of Birds PDFFabricio GZNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Thesis Olof HerttingDocument57 pagesThesis Olof HerttingAjay IyerNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Characterization of Polydimethylsiloxane (PDMS) PropertiesDocument13 pagesCharacterization of Polydimethylsiloxane (PDMS) PropertiesEsteban ArayaNo ratings yet
- General Katalog PT. AmoebaDocument23 pagesGeneral Katalog PT. AmoebaMulyanaNo ratings yet
- Kenya's Natural Capital AtlasDocument139 pagesKenya's Natural Capital AtlasElvis Ole Sankale Ntimama100% (7)
- Field Inspection Techniques Ifad-CaspDocument17 pagesField Inspection Techniques Ifad-CaspNasiru Kura100% (1)
- Protochordata FIXDocument33 pagesProtochordata FIXSylvia AnggraeniNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Original RT-PCR ReportDocument1 pageOriginal RT-PCR ReportVishva IyerNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Lab Exercise 0ne: Carbohydrate Analysis Lab A.1 (Page 28)Document31 pagesLab Exercise 0ne: Carbohydrate Analysis Lab A.1 (Page 28)Goh Kae HorngNo ratings yet
- 2nd Periodic Test (Science)Document1 page2nd Periodic Test (Science)mosarbas0950% (2)
- Sickle Cell AnemiaDocument7 pagesSickle Cell AnemiaJuma AwarNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Growth and DevelopmentDocument10 pagesGrowth and DevelopmentPiyush DuttaNo ratings yet
- TC QMM 56942Document120 pagesTC QMM 56942Fernando R EpilNo ratings yet
- 03 Chapter 3Document148 pages03 Chapter 3arghaNo ratings yet
- When The Koel SingsDocument15 pagesWhen The Koel SingsAlfie Paduga BeguinaNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Book of Mythical BeastsDocument32 pagesBook of Mythical BeastsRishabhNo ratings yet
- Biomimicry Avjeet SinghDocument21 pagesBiomimicry Avjeet SinghAvjeet SinghNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)